Solute Transport and Transformation in an Intermittent, Headwater Mountain Stream with Diurnal Discharge Fluctuations

 , ,
, ,  , , , ,
, , , ,
Abstract
1. Introduction
2. Methods
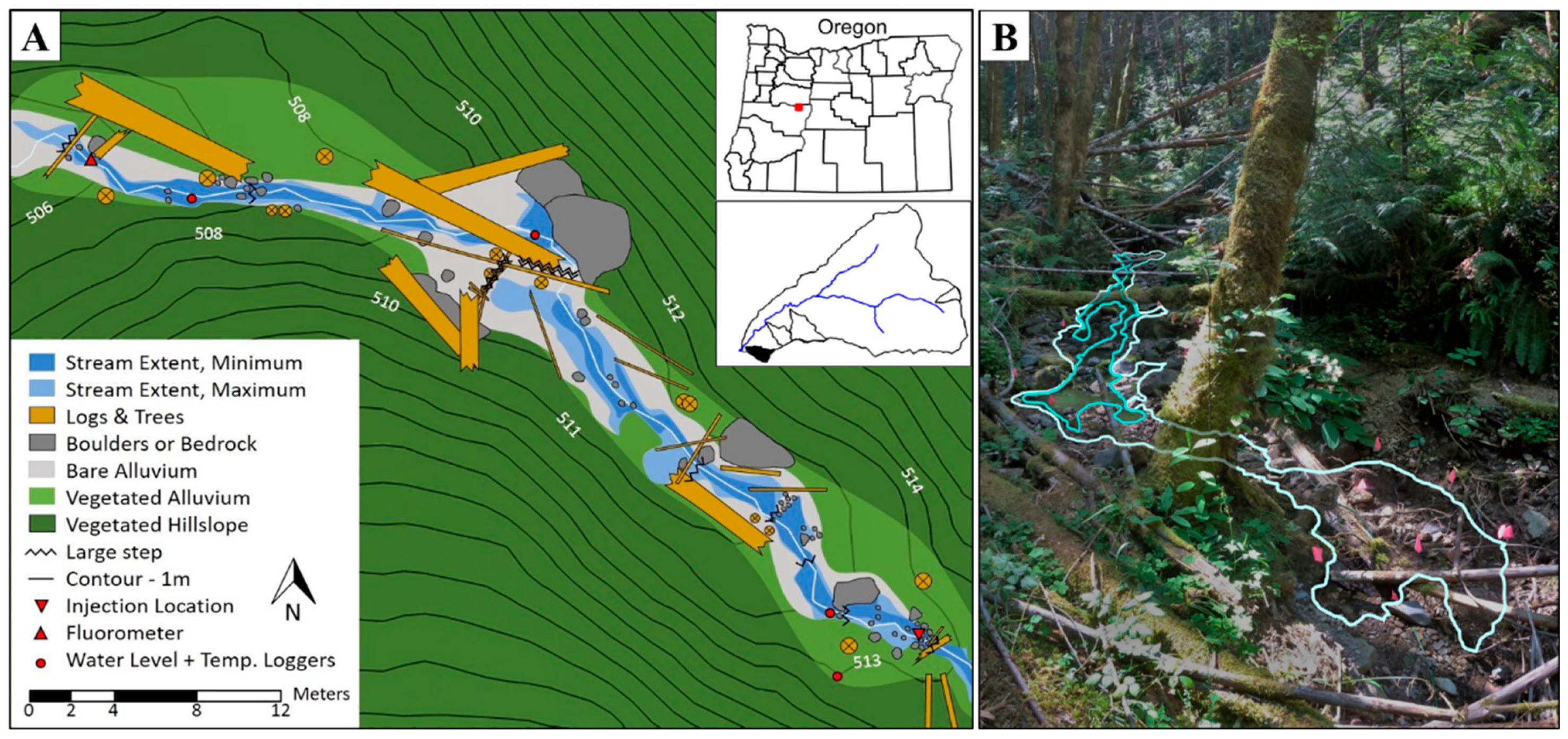
2.1. Site Description
2.2. Stream Solute Tracer Experiments
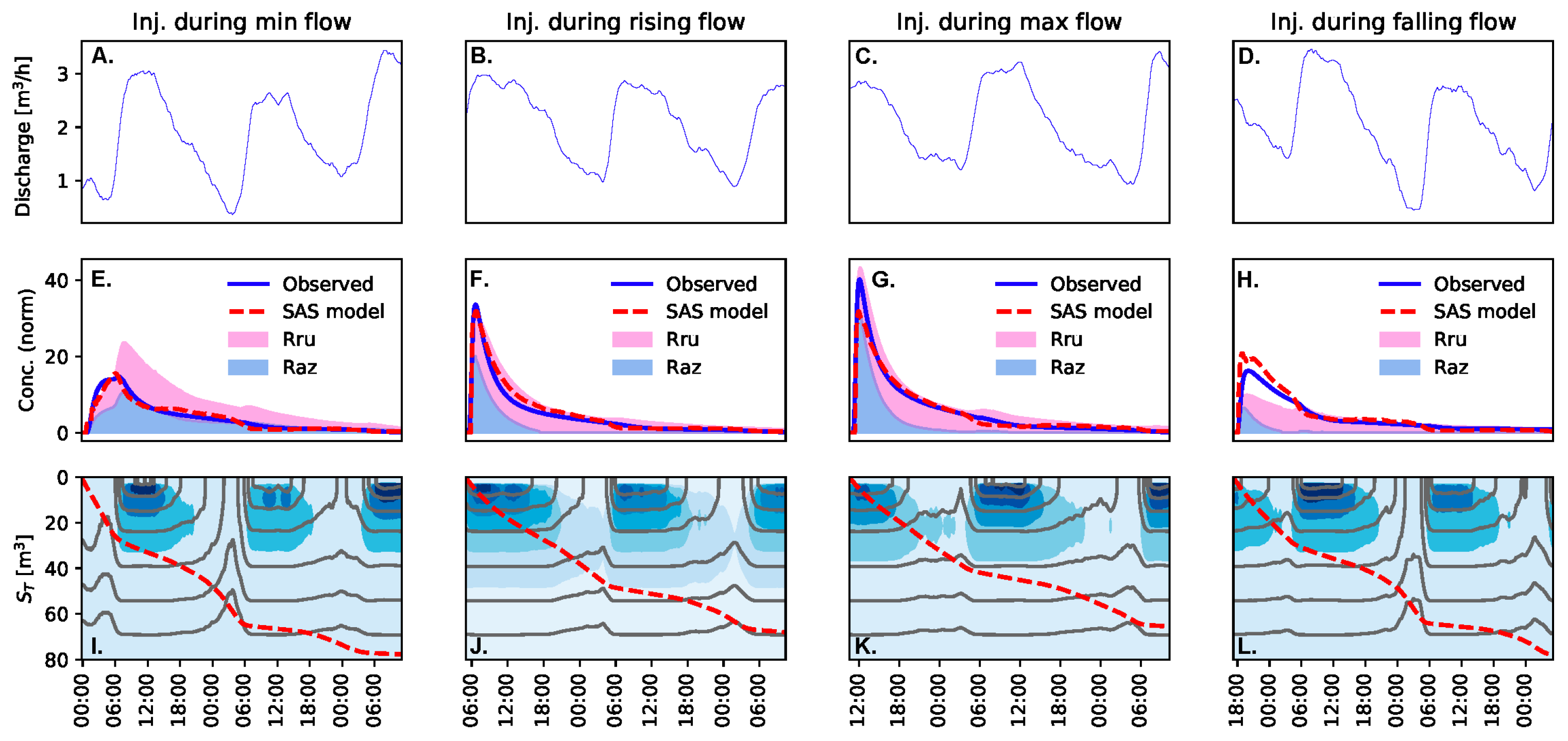
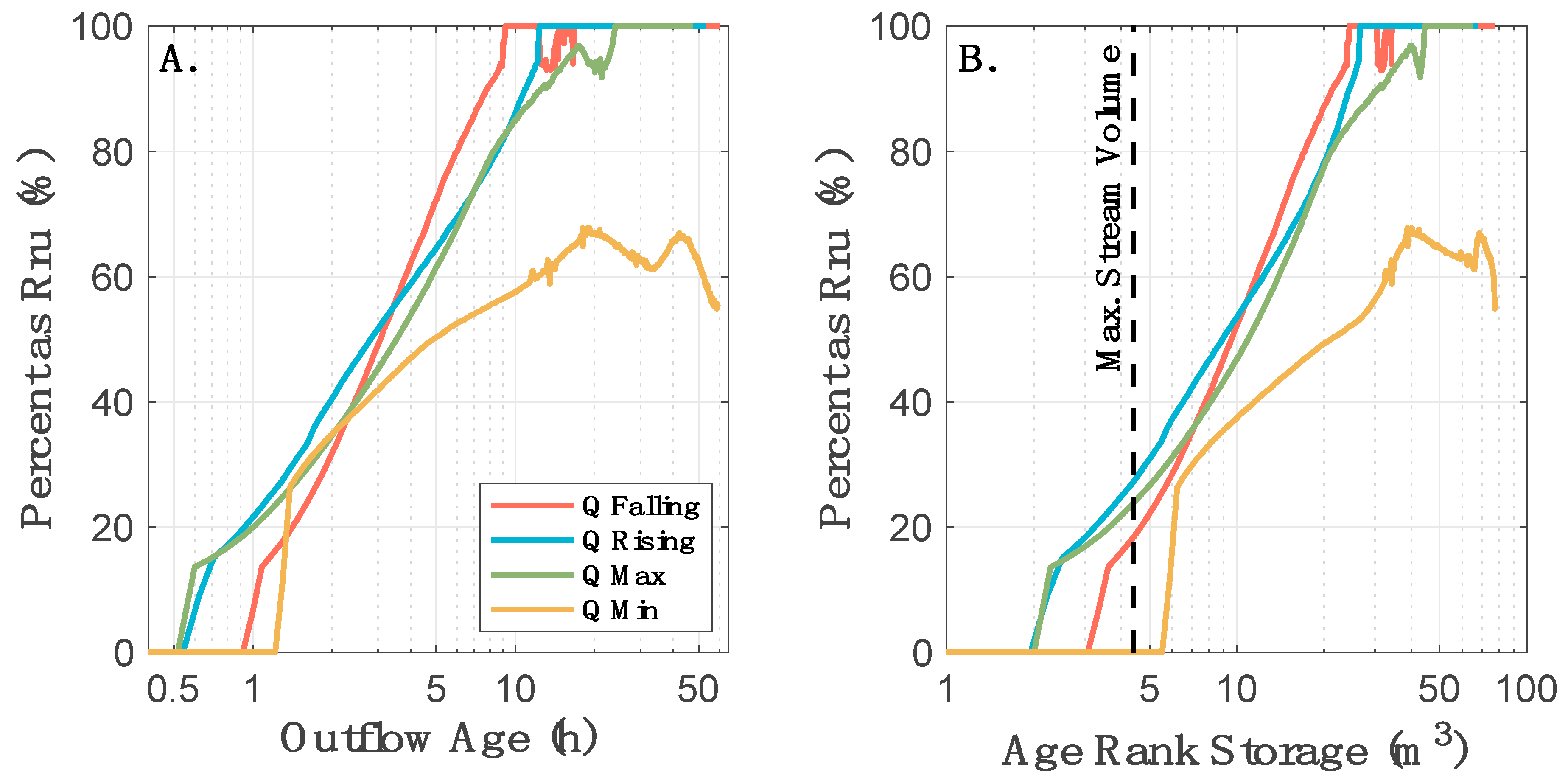
2.3. StorAge Selection Modeling
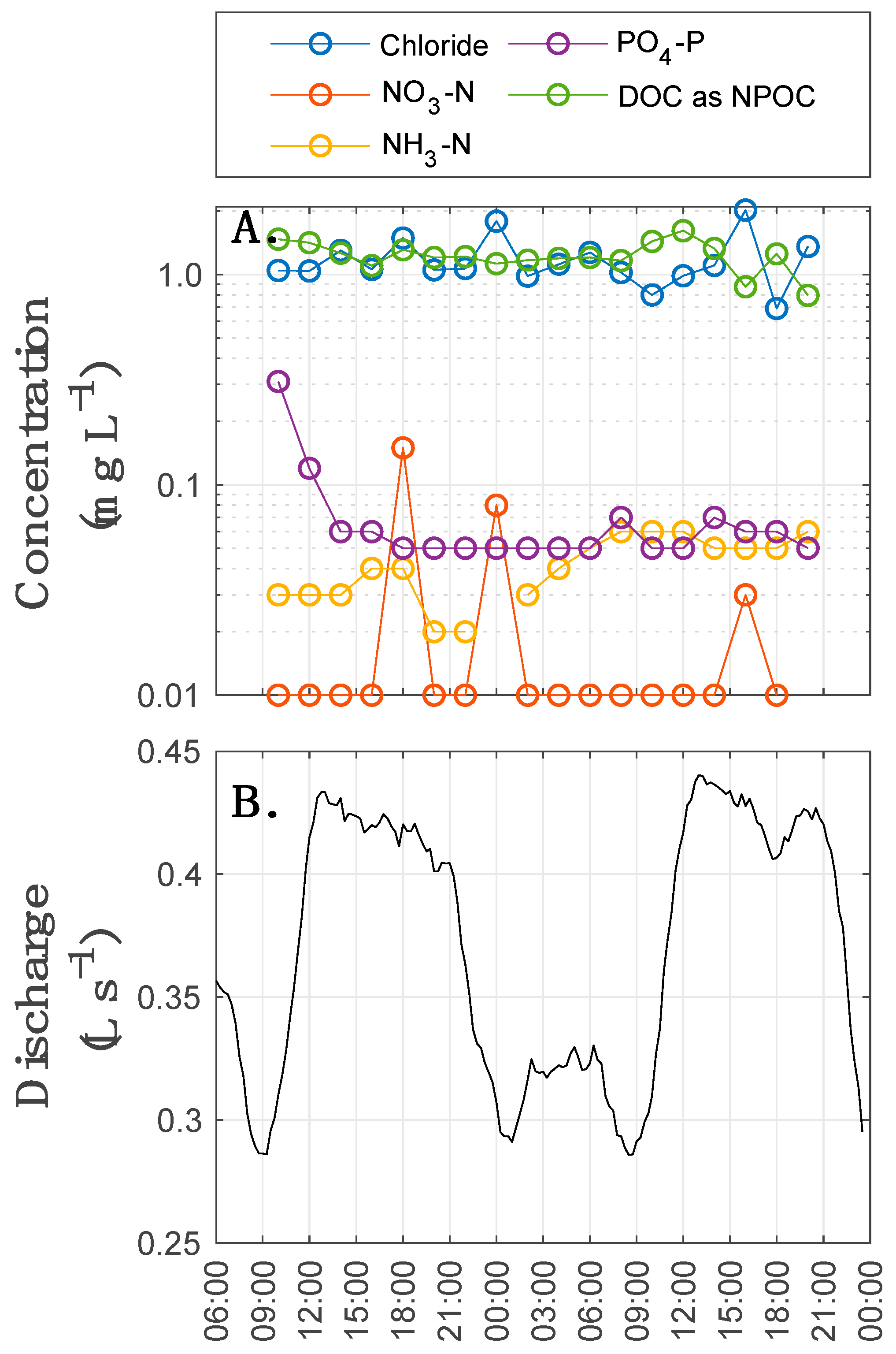
2.4. In-Stream Biogeochemistry
3. Results and Discussion
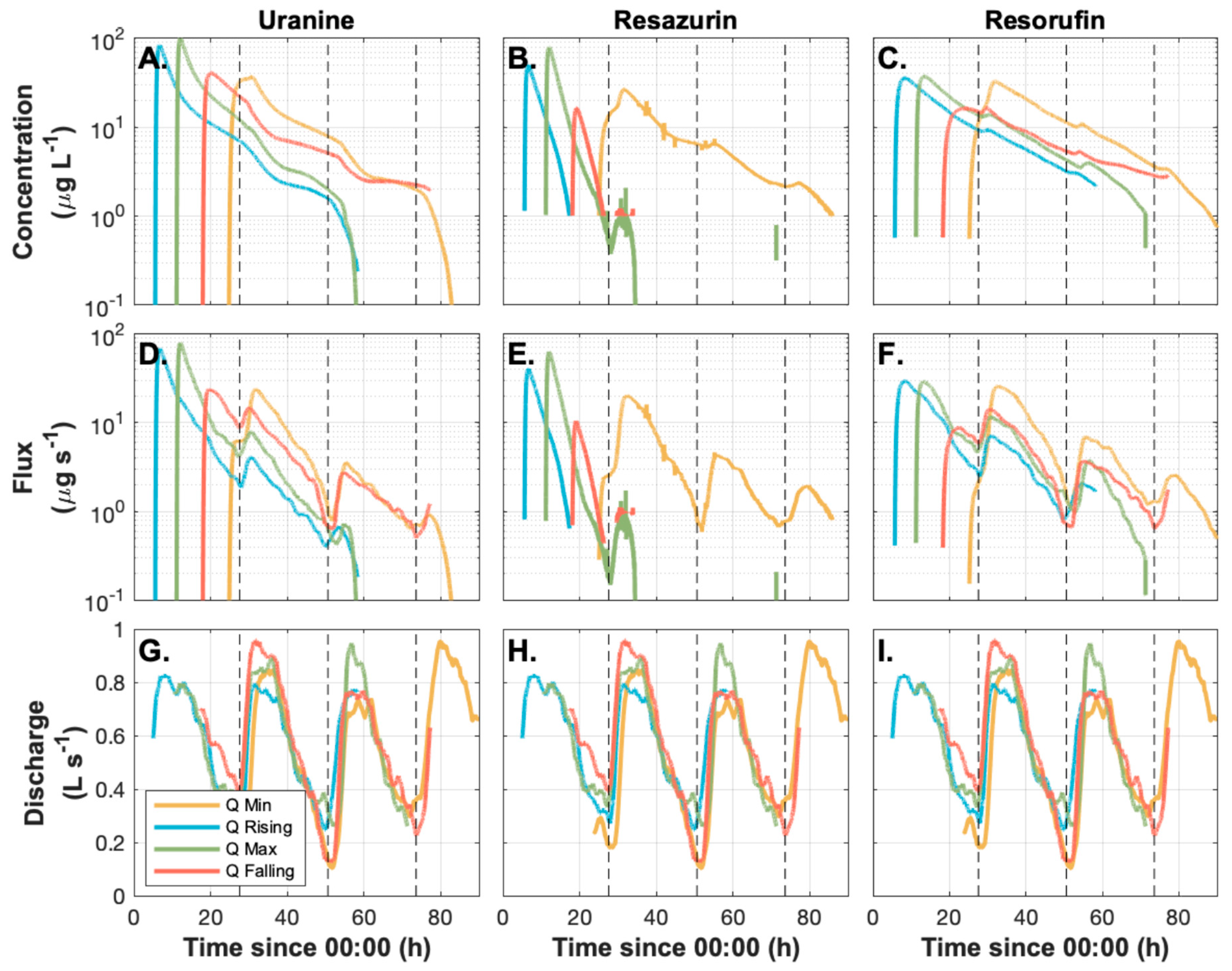
3.1. Conservative Solute Tracer Transport Fluctuates Consistently with Unsteady Discharge
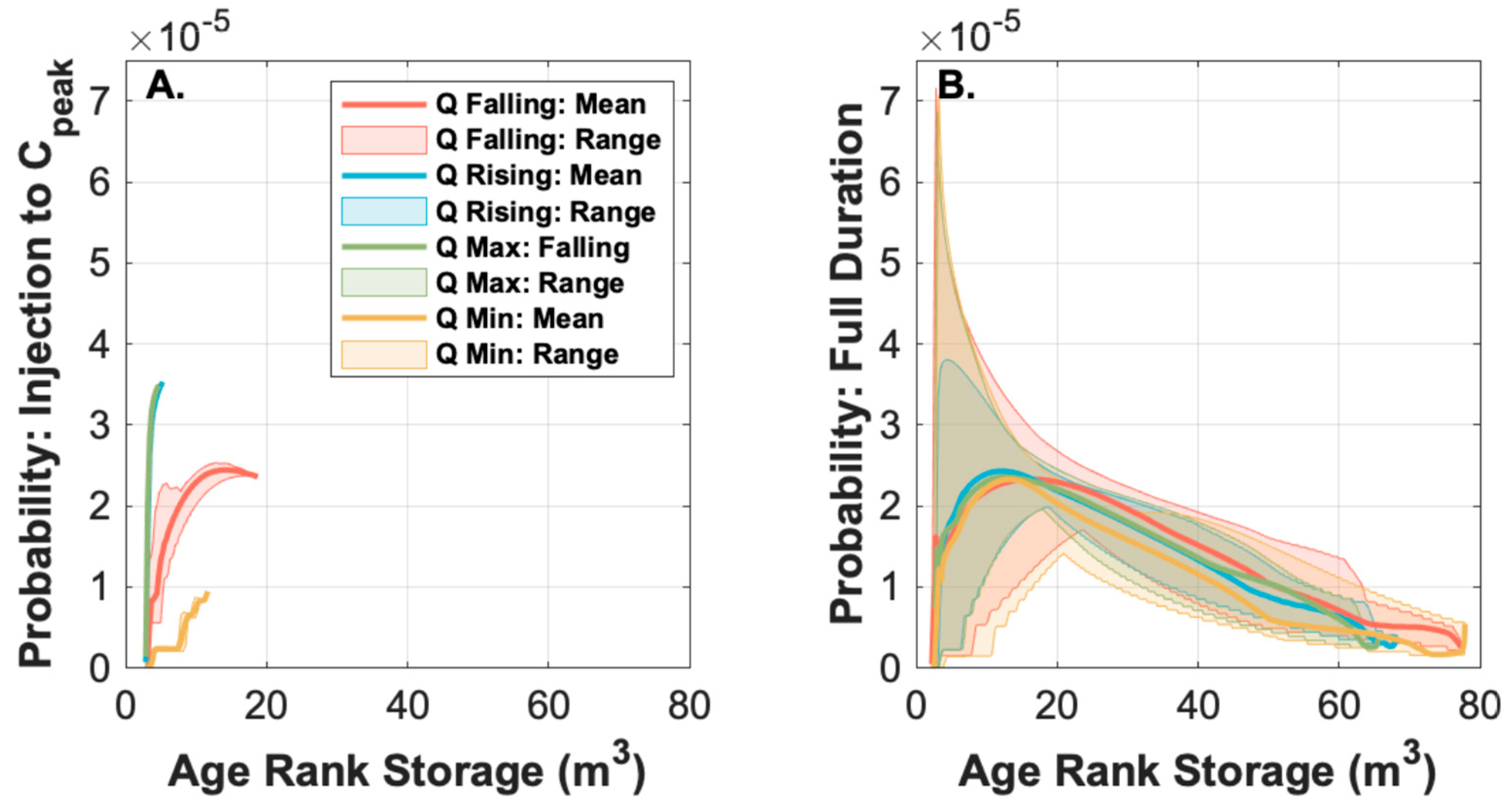
3.2. Observed Variation in Solute Transport is Primarily Due to Storage Release Timing Relative to an Oscillating Diurnal Cycle in the Underlying Transport Processes
3.3. Spatial and Temporal Variation in Raz-to-Rru Transformation
3.3.1. Rru Production Does Not Appear to Change with Discharge
3.3.2. Older Storage is Not as Metabolically Active as Younger Storage
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boano, F.; Harvey, J.W.; Marion, A.; Packman, A.I.; Revelli, R.; Ridolfi, L.; Worman, A. Hyporheic flow and transport processes: Mechanisms, models, and biogeochemical implications. Rev. Geophys. 2014, 52, 2012RG000417. [Google Scholar] [CrossRef]
- Maher, K. The role of fluid residence time and topographic scales in determining chemical fluxes from landscapes. Earth Planet. Sci. Lett. 2011, 312, 48–58. [Google Scholar] [CrossRef]
- Bardini, L.; Boano, F.; Cardenas, M.B.; Revelli, R.; Ridolfi, L. Nutrient cycling in bedform induced hyporheic zones. Geochim. Cosmochim. Acta 2012, 84, 47–61. [Google Scholar] [CrossRef]
- Harvey, J.; Gomez-Velez, J.; Schmadel, N.M.; Scott, D.; Boyer, E.; Alexander, R.; Eng, K.; Golden, H.; Kettner, A.; Konrad, C.; et al. How Hydrologic Connectivity Regulates Water Quality in River Corridors. J. Am. Water Resour. Assoc. 2019, 55, 369–381. [Google Scholar] [CrossRef]
- Ward, A.S.; Schmadel, N.M.; Wondzell, S.M. Time-Variable Transit Time Distributions in the Hyporheic Zone of a Headwater Mountain Stream. Water Resour. Res. 2018. [Google Scholar] [CrossRef]
- Van Meerveld, H.J.I.; Kirchner, J.W.; Vis, M.J.P.; Assendelft, R.S.; Seibert, J. Expansion and contraction of the flowing stream network changes hillslope flowpath lengths and the shape of the travel time distribution. Hydrol. Earth Syst. Sci. Discuss. 2019, 2006, 1–18. [Google Scholar] [CrossRef]
- Pinay, G.; Clément, J.C.; Naiman, R.J. Basic Principles and Ecological Consequences of Changing Water Regimes on Nitrogen Cycling in Fluvial Systems. Environ. Manage. 2002, 30, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.S.; Gooseff, M.N.; Voltz, T.J.; Fitzgerald, M.; Singha, K.; Zarnetske, J.P. How does rapidly changing discharge during storm events affect transient storage and channel water balance in a headwater mountain stream? Water Resour. Res. 2013, 49, 5473–5486. [Google Scholar] [CrossRef]
- Dudley-Southern, M.; Binley, A. Temporal responses of groundwater-surface water exchange to successive storm events. Water Resour. Res. 2015, 51, 1112–1126. [Google Scholar] [CrossRef]
- Schmadel, N.M.; Ward, A.S.; Lowry, C.S.; Malzone, J.M. Hyporheic exchange controlled by dynamic hydrologic boundary conditions. Geophys. Res. Lett. 2016, 43, 4408–4417. [Google Scholar] [CrossRef]
- Malzone, J.M.; Anseeuw, S.K.; Lowry, C.S.; Allen-King, R. Temporal Hyporheic Zone Response to Water Table Fluctuations. Groundwater 2015, 54, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Malzone, J.M.; Lowry, C.S.; Ward, A.S. Response of the hyporheic zone to transient groundwater fluctuations on the annual and storm event time scales. Water Resour. Res. 2016, 52, 1–20. [Google Scholar] [CrossRef]
- Sawyer, A.H.; Shi, F.; Kirby, J.T.; Michael, H.A. Dynamic response of surface water-groundwater exchange to currents, tides, and waves in a shallow estuary. J. Geophys. Res. Ocean. 2013, 118, 1749–1758. [Google Scholar] [CrossRef]
- Musial, C.; Sawyer, A.H.; Barnes, R.; Bray, S.; Knights, D. Surface water–groundwater exchange dynamics in a tidal freshwater zone. Hydrol. Process. 2016, 30, 739–750. [Google Scholar] [CrossRef]
- Bianchin, M.S.; Smith, L.; Beckie, R.D. Defining the hyporheic zone in a large tidally influenced river. J. Hydrol. 2011, 406, 16–29. [Google Scholar] [CrossRef]
- Knights, D.; Sawyer, A.H.; Barnes, R.T.; Musial, C.T.; Bray, S. Tidal controls on riverbed denitrification along a tidal freshwater zone. Water Resour. Res. 2017, 53, 799–816. [Google Scholar] [CrossRef]
- Payn, R.A.; Gooseff, M.N.; McGlynn, B.L.; Bencala, K.E.; Wondzell, S.M. Channel water balance and exchange with subsurface flow along a mountain headwater stream in Montana, United States. Water Resour. Res. 2009, 45. [Google Scholar] [CrossRef]
- Ward, A.S.; Payn, R.A.; Gooseff, M.N.; McGlynn, B.L.; Bencala, K.E.; Kelleher, C.A.; Wondzell, S.M.; Wagener, T. Variations in surface water-ground water interactions along a headwater mountain stream: Comparisons between transient storage and water balance analyses. Water Resour. Res. 2013, 49, 3359–3374. [Google Scholar] [CrossRef]
- Ward, A.S.; Fitzgerald, M.; Gooseff, M.N.; Voltz, T.J.; Binley, A.M.; Singha, K. Hydrologic and geomorphic controls on hyporheic exchange during base flow recession in a headwater mountain stream. Water Resour. Res. 2012, 48, W04513. [Google Scholar] [CrossRef]
- Ward, A.S.; Gooseff, M.N.; Fitzgerald, M.; Voltz, T.J.; Singha, K. Spatially distributed characterization of hyporheic solute transport during baseflow recession in a headwater mountain stream using electrical geophysical imaging. J. Hydrol. 2014, 517, 362–377. [Google Scholar] [CrossRef]
- Wondzell, S.M.; Swanson, F.J. Seasonal and Storm Dynamics of the Hyporheic Zone of a 4th-Order Mountain Stream. I: Hydrologic Processes. J. North Am. Beth. Soc. 1996, 15, 3–19. [Google Scholar]
- Voltz, T.J.; Gooseff, M.N.; Ward, A.S.; Singha, K.; Fitzgerald, M.; Wagener, T. Riparian hydraulic gradient and stream-groundwater exchange dynamics in steep headwater valleys. J. Geophys. Res. Earth Surf. 2013, 118, 953–969. [Google Scholar]
- Ward, A.S.; Schmadel, N.M.; Wondzell, S.M. Simulation of dynamic expansion, contraction, and connectivity in a mountain stream network. Adv. Water Resour. 2018, 114, 64–82. [Google Scholar]
- Bond, B.J.; Jones, J.A.; Moore, G.; Phillips, N.; Post, D.; McDonnell, J.J. The zone of vegetation influence on baseflow revealed by diel patterns of streamflow and vegetation water use in a headwater basin. Hydrol. Process. 2002, 16, 1671–1677. [Google Scholar]
- Koch, J.C.; McKnight, D.M.; Neupauer, R.M. Simulating unsteady flow, anabranching, and hyporheic dynamics in a glacial meltwater stream using a coupled surface water routing and groundwater flow model. Water Resour. Res. 2011, 47, 1–15. [Google Scholar] [CrossRef]
- Wlostowski, A.N.; Gooseff, M.N.; McKnight, D.M.; Lyons, W.B. Transit Times and Rapid Chemical Equilibrium Explain Chemostasis in Glacial Meltwater Streams in the McMurdo Dry Valleys, Antarctica. Geophys. Res. Lett. 2018, 45, 13322–13331. [Google Scholar] [CrossRef]
- Lowry, C.S.; Deems, J.S.; Loheide, S.P.; Lundquist, J.D. Linking snowmelt-derived fluxes and groundwater flow in a high elevation meadow system, Sierra Nevada Mountains, California. Hydrol. Process. 2010, 24, 2821–2833. [Google Scholar]
- Ii, S.P.L.; Lundquist, J.D. Snowmelt-induced diel fluxes through the hyporheic zone. Water Resour. Res. 2009, 45, 1–9. [Google Scholar]
- Sawyer, A.H.; Cardenas, M.B.; Bomar, A.; Mackey, M. Impact of dam operations on hyporheic exchange in the riparian zone of a regulated river. Hydrol. Process. 2009, 23, 2129–2137. [Google Scholar]
- Gerecht, K.E.; Cardenas, M.B.; Guswa, A.J.; Sawyer, A.H.; Nowinski, J.D.; Swanson, T.E. Dynamics of hyporheic flow and heat transport across a bed-to-bank continuum in a large regulated river. Water Resour. Res. 2011, 47, W03524. [Google Scholar] [CrossRef]
- Hancock, P.J.; Boulton, A.J. The effects of an environmental flow release on water quality in the hyporheic zone of the Hunter River, Australia. Hydrobiologia 2005, 552, 75–85. [Google Scholar] [CrossRef]
- Liu, D.; Zhao, J.; Chen, X.; Li, Y.; Weiyan, S.; Feng, M. Dynamic processes of hyporheic exchange and temperature distribution in the riparian zone in response to dam-induced water fluctuations. Geosci. J. 2018, 22, 465–475. [Google Scholar] [CrossRef]
- Paul, M.T.; Meyer, J.L. Streams in the Urban Landscape. Annu. Rev. Ecol. Evol. Syst. 2001, 32, 333–365. [Google Scholar] [CrossRef]
- Butturini, A.; Sabater, F. Importance of transient storage zones for ammonium and phosphate retention in a sandy-bottom Mediterranean stream. Freshw. Biol. 1999, 41, 593–603. [Google Scholar] [CrossRef]
- Fabian, M.W.; Endreny, T.A.; Bottacin-Busolin, A.; Lautz, L.K. Seasonal variation in cascade-driven hyporheic exchange, northern Honduras. Hydrol. Process. 2011, 25, 1630–1646. [Google Scholar] [CrossRef]
- Wondzell, S.M. The role of the hyporheic zone across stream networks. Hydrol. Process. 2011, 25, 3525–3532. [Google Scholar] [CrossRef]
- Zarnetske, J.P.; Gooseff, M.N.; Brosten, T.R.; Bradford, J.H.; McNamara, J.P.; Bowden, W.B. Transient storage as a function of geomorphology, discharge, and permafrost active layer conditions in Arctic tundra streams. Water Resour. Res. 2007, 43, W07410. [Google Scholar] [CrossRef]
- Hart, D.R.; Mulholland, P.J.; Marzolf, E.R.; DeAngelis, D.L.; Hendricks, S.P. Relationships between hydraulic parameters in a small stream under varying flow and seasonal conditions. Hydrol. Process. 1999, 13, 1497–1510. [Google Scholar] [CrossRef]
- Jin, H.S.; Ward, G.M. Hydraulic characteristics of a small Coastal Plain stream of the southeastern United States: Effects of hydrology and season. Hydrol. Process. 2005, 19, 4147–4160. [Google Scholar] [CrossRef]
- Karwan, D.L.; Saiers, J.E. Influences of seasonal flow regime on the fate and transport of fine particles and a dissolved solute in a New England stream. Water Resour. Res. 2009, 45, W11423. [Google Scholar] [CrossRef]
- Morrice, J.A.; Valett, H.M.; Dahm, C.N.; Campana, M.E. Alluvial characteristics, groundwater-surface water exchange and hydrological retention in headwater streams. Hydrol. Process. 1997, 11, 253–267. [Google Scholar]
- Schmid, B.H. Can Longitudinal Solute Transport Parameters Be Transferred to Different Flow Rates? J. Hydrol. Eng. 2008, 13, 505–509. [Google Scholar]
- Schmid, B.H.; Innocenti, I.; San, U.; Sanfilippo, U. Characterizing solute transport with transient storage across a range of flow rates: The evidence of repeated tracer experiments in Austrian and Italian streams. Adv. Water Resour. 2010, 33, 1340–1346. [Google Scholar]
- Ward, A.S.; Gooseff, M.N.; Singha, K. How does subsurface characterization affect simulations of hyporheic exchange? GroundWater 2013, 51, 14–28. [Google Scholar]
- Wondzell, S.M. Effect of morphology and discharge on hyporheic exchange flows in two small streams in the Cascade Mountains of Oregon, USA. Hydrol. Process. 2006, 20, 267–287. [Google Scholar]
- Jackson, T.R.; Haggerty, R.; Apte, S.V.; O’Connor, B.L. A mean residence time relationship for lateral cavities in gravel-bed rivers and streams: Incorporating streambed roughness and cavity shape. Water Resour. Res. 2013, 49, 3642–3650. [Google Scholar]
- Jackson, T.R.; Haggerty, R.; Apte, S.V. A fluid-mechanics based classification scheme for surface transient storage in riverine environments: Quantitatively separating surface from hyporheic transient storage. Hydrol. Earth Syst. Sci. 2013, 17, 2747–2779. [Google Scholar]
- Jackson, T.R.; Haggerty, R.; Apte, S.V.; Coleman, A.; Drost, K.J. Defining and measuring the mean residence time of lateral surface transient storage zones in small streams. Water Resour. Res. 2012, 48, 1–20. [Google Scholar]
- Briggs, M.A.; Gooseff, M.N.; Arp, C.D.; Baker, M.A. A method for estimating surface transient storage parameters for streams with concurrent hyporheic storage. Water Resour. Res. 2009, 45. [Google Scholar] [CrossRef]
- Pinder, G.F.; Sauer, S.P. Numerical simulation of flood wave modification due to bank storage effects. Water Resour. Res. 1971, 7, 63–70. [Google Scholar]
- Cooper, H.; Rorabaugh, M. Groundwater Movments and Bank Storage Due to Flood Stages in Surface Streams; US Government Printing Office: Washington, DC, USA, 1963.
- Ward, A.S.; Schmadel, N.M.; Wondzell, S.M.; Gooseff, M.N.; Singha, K. Dynamic hyporheic and riparian flow path geometry through base flow recession in two headwater mountain stream corridors. Water Resour. Res. 2017, 53, 3988–4003. [Google Scholar]
- Schmadel, N.M.; Ward, A.S.; Wondzell, S.M. Hydrologic controls on hyporheic exchange in headwater mountain streams. Water Resour. Res. 2017, 53, 6260–6278. [Google Scholar]
- Seitzinger, S.P.; Styles, R.V.; Boyer, E.W.; Alexander, R.B.; Billen, G.; Howarth, R.W.; Mayer, B.; Van Breemen, N. Nitrogen retention in rivers: Model development and application to watersheds in the northeastern USA. Biogeochemistry 2002, 57, 199–237. [Google Scholar]
- Wollheim, W.M.; Vo, C.J.; Vorosmarty, C.J.; Peterson, B.J.; Seitzinger, S.P.; Hopkinson, C.S.; Vörösmarty, C.J. Relationship between river size and nutrient removal. Geophys. Res. Lett. 2006, 33, 4. [Google Scholar]
- Basu, N.B.; Rao, P.S.C.; Thompson, S.E.; Loukinova, N.V.; Donner, S.D.; Ye, S.; Sivapalan, M. Spatiotemporal averaging of in-stream solute removal dynamics. Water Resour. Res. 2011, 47. [Google Scholar] [CrossRef]
- Harvey, J.W.; Gooseff, M.N. River corridor science: Hydrologic exchange and ecological consequences from bedforms to basins. Water Resour. Res. 2015, 51, 6893–6922. [Google Scholar] [CrossRef]
- McConville, M.B.; Cohen, N.M.; Nowicki, S.M.; Lantz, S.R.; Hixson, J.L.; Ward, A.S.; Remucal, C.K. A field analysis of lampricide photodegradation in Great Lakes tributaries. Environ. Sci. Process. Impacts 2017, 19. [Google Scholar] [CrossRef]
- Young, R.G.; Huryn, A.D. Comment: Improvements to the diurnal upstream-downstream dissolved. Can. J. Fish. Aquat. Sci. 1998, 55, 1784–1785. [Google Scholar]
- Hall, R.O.; Yackulic, C.B.; Kennedy, T.A.; Yard, M.D.; Rosi-Marshall, E.J.; Voichick, N.; Behn, K.E. Turbidity, light, temperature, and hydropeaking control primary productivity in the Colorado River, Grand Canyon. Limnol. Oceanogr. 2015, 60, 512–526. [Google Scholar]
- Raymond, P.A.; Saiers, J.E.; Sobczak, W.V. Hydrological and biogeochemical controls on watershed dissolved organic matter transport: Pulse- shunt concept. Ecology 2016, 97, 5–16. [Google Scholar]
- Alexander, R.B.; Smith, R.A.; Schwarz, G.E. Effect of stream channel size on the delivery of nitrogen to the Gulf of Mexico. Nature 2000, 403, 758–761. [Google Scholar] [PubMed]
- Hart, D.D.; Finelli, C.M. Physical-biological coupling in streams: The pervasive effects of flow on benthic organisms. Am. Rev. Ecol. Syst. 1999, 30, 363–395. [Google Scholar]
- Rauter, A.; Weigelhofer, G.; Waringer, J.; Battin, T.J. Transport and metabolic fate of sewage particles in a recipient stream. J. Environ. Qual. 2005, 34, 1591–1599. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Aubeneau, A.F.; Hanrahan, B.; Bolster, D.; Tank, J.L. Biofilm growth in gravel bed streams controls solute residence time distributions. J. Geophys. Res. Biogeosciences 2016, 121, 1840–1850. [Google Scholar]
- Marti, E.; Grimm, N.B.; Fisher, S.G. Pre-and post-flood retention efficiency of nitrogen in a Sonoran Desert stream. J. North Am. Benthol. Soc. 1997, 16, 805–819. [Google Scholar] [CrossRef]
- Uehlinger, U. Resistance and resilience of ecosystem metabolism in a flood-prone river system. Freshw. Biol. 2000, 45, 319–332. [Google Scholar]
- Uehlinger, U.; Kawecka, B.; Robinson, C.T. Effects of experimental floods on periphyton and stream metabolism below a high dam in the Swiss Alps (River Spöl). Aquat. Sci. 2003, 65, 199–209. [Google Scholar]
- Bernal, S.; von Schiller, D.; Sabater, F.; Martí, E. Hydrological extremes modulate nutrient dynamics in mediterranean climate streams across different spatial scales. Hydrobiologia 2013, 719, 31–42. [Google Scholar]
- Guerin, T.F.; Mondido, M.; McClenn, B.; Peasley, B. Application of resazurin for estimating abundance of contaminant-degrading micro-organisms. Lett. Appl. Microbiol. 2001, 32, 340–345. [Google Scholar] [CrossRef]
- O’Brien, J.M.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef]
- Knapp, J.L.A.; González-Pinzón, R.; Haggerty, R. The Resazurin-Resorufin System: Insights From a Decade of “Smart” Tracer Development for Hydrologic Applications. Water Resour. Res. 2018, 54, 6877–6889. [Google Scholar] [CrossRef]
- Haggerty, R.; Argerich, A.; Martí, E.; Marti, E. Development of a “smart” tracer for the assessment of microbiological activity and sediment-water interaction in natural waters: The resazurin-resorufin system. Water Resour. Res. 2008, 44, W00D01. [Google Scholar] [CrossRef]
- Haggerty, R.; Ribot, M.; Singer, G.A.; Marti, E.; Argerich, A.; Agell, G.; Battin, T.J. Ecosystemrespiration increases with biofilm growth and bed forms: Flume measurements with resazurin. J. Geophys. Res. Biogeosciences 2014, 119, 2220–2230. [Google Scholar] [CrossRef]
- Knapp, J.L.A.; González-Pinzón, R.; Drummond, J.D.; Larsen, L.G.; Cirpka, O.A.; Harvey, J.W. Tracer-based characterization of hyporheic exchange and benthic biolayers in streams. Water Resour. Res. 2017, 53, 1575–1594. [Google Scholar] [CrossRef]
- Folegot, S.; Krause, S.; Mons, R.; Hannah, D.M.; Datry, T. Mesocosm experiments reveal the direction of groundwater–surface water exchange alters the hyporheic refuge capacity under warming scenarios. Freshw. Biol. 2018, 63, 165–177. [Google Scholar] [CrossRef]
- Kurz, M.J.; Drummond, J.D.; Mart??, E.; Zarnetske, J.P.; Lee-Cullin, J.; Klaar, M.J.; Folegot, S.; Keller, T.; Ward, A.S.; Fleckenstein, J.H.; et al. Impacts of water level on metabolism and transient storage in vegetated lowland rivers: Insights from a mesocosm study. J. Geophys. Res. Biogeosciences 2017, 122, 628–644. [Google Scholar] [CrossRef]
- Baranov, V.; Lewandowski, J.; Romeijn, P.; Singer, G.; Krause, S. Effects of bioirrigation of non-biting midges (Diptera: Chironomidae) on lake sediment respiration. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Baranov, V.; Lewandowski, J.; Krause, S. Bioturbation enhances the aerobic respiration of lake sediments in warming lakes. Biol. Lett. 2016, 12, 20160448. [Google Scholar] [CrossRef]
- Comer-Warner, S.A.; Romeijn, P.; Gooddy, D.C.; Ullah, S.; Kettridge, N.; Marchant, B.; Hannah, D.M.; Krause, S. Thermal sensitivity of CO2 and CH4 emissions varies with streambed sediment properties. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef]
- Romeijn, P.; Comer-Warner, S.A.; Ullah, S.; Hannah, D.M.; Krause, S. Streambed Organic Matter Controls on Carbon Dioxide and Methane Emissions from Streams. Environ. Sci. Technol. 2019, 53, 2364–2374. [Google Scholar] [CrossRef]
- Blaen, P.J.; Kurz, M.J.; Drummond, J.D.; Knapp, J.L.A.; Mendoza-Lera, C.; Schmadel, N.M.; Klaar, M.J.; Jäger, A.; Folegot, S.; Lee-Cullin, J.; et al. Woody debris is related to reach-scale hotspots of lowland stream ecosystem respiration under baseflow conditions. Ecohydrology 2018, 11, 1–9. [Google Scholar] [CrossRef]
- Argerich, A.; Haggerty, R.; Martí, E.; Sabater, F.; Zarnetske, J.P.; De Barcelona, U.; Haggerty, R.; Sabater, F.; Zarnetske, J.P.; Marti, E.; et al. Quantification of metabolically active transient storage (MATS) in two reaches with contrasting transient storage and ecosystem respiration. J. Geophys. Res. 2011, 116, G03034. [Google Scholar] [CrossRef]
- González-Pinzón, R.; Haggerty, R.; Myrold, D.D. Measuring aerobic respiration in stream ecosystems using the resazurin-resorufin system. J. Geophys. Res. Biogeosciences 2012, 117, 1–10. [Google Scholar] [CrossRef]
- Lemke, D.; Liao, Z.; Wöhling, T.; Osenbrück, K.; Cirpka, O.A. Concurrent conservative and reactive tracer tests in a stream undergoing hyporheic exchange. Water Resour. Res. 2013, 49, 3024–3037. [Google Scholar] [CrossRef]
- Kelleher, C.; Ward, A.S.; Knapp, J.L.A.; Blaen, P.J.; Kurz, M.J.; Drummond, J.D.; Zarnetske, J.P.; Hannah, D.M.; Mendoza-Lera, C.; Schmadel, N.M.; et al. Exploring Tracer Information and Model Framework Trade-Offs to Improve Estimation of Stream Transient Storage Processes. Water Resour. Res. 2019, 55, 3481–3501. [Google Scholar] [CrossRef]
- Kelleher, C.A.; Wagener, T.; McGlynn, B.L.; Ward, A.S.; Gooseff, M.N.; Payn, R.A. Identifiability of transient storage model parameters along a mountain stream. Water Resour. Res. 2013, 49, 5290–5306. [Google Scholar] [CrossRef]
- Ward, A.S.; Kelleher, C.A.; Mason, S.J.K.; Wagener, T. A software tool to assess uncertainty in transient-storage model parameters using Monte Carlo simulations. Freshw. Sci. 2016, 36. [Google Scholar] [CrossRef]
- Wagner, B.J.; Harvey, J.W. Experimental design for estimating parameters of rate-limited mass transfer: Analysis of stream tracer studies. Water Resour. Res. 1997, 33, 1731–1741. [Google Scholar] [CrossRef]
- Wagener, T.; Kollat, J. Numerical and visual evaluation of hydrological and environmental models using the Monte Carlo analysis toolbox. Environ. Model. Softw. 2007, 22, 1021–1033. [Google Scholar] [CrossRef]
- Bencala, K.E.; Walters, R.A. Simulation of solute transport in a mountain pool-and-riffle stream: A transient storage model. Water Resour. Res. 1983, 19, 718–724. [Google Scholar] [CrossRef]
- Haggerty, R.; Reeves, P. STAMMT-L Version 1.0 User’s Manual; Sandia National Laboratories: Albuquerque, NM, USA, 2002; pp. 1–76.
- Runkel, R.L. One-dimensional Transport with Inflow and Storage (OTIS): A Solute Transport Model for Streams and Rivers; Information Services [distributor]; ISBN Water-Resources Investigations Report 98–4018; US Dept. of the Interior, US Geological Survey: Washington, DC, USA, 1998.
- Harman, C.J. Time-variable transit time distributions and transport: Theory and application to storage-dependent transport of chloride in awatershed. Water Resour. Res. 2015, 51, 1–30. [Google Scholar] [CrossRef]
- Harman, C.J.; Ward, A.S.; Ball, A. How does reach-scale stream-hyporheic transport vary with discharge? Insights fromrSAS analysis of sequential tracer injections in a headwater mountain stream. Water Resour. Res. 2016, 52, 7130–7150. [Google Scholar] [CrossRef]
- Ward, A.S.; Wondzell, S.M.; Schmadel, N.M.; Herzog, S.; Zarnetske, J.P.; Baranov, V.; Blaen, P.J.; Brekenfeld, N.; Chu, R.; Derelle, R.; et al. Spatial and temporal variation in river corridor exchange across a 5th order mountain stream network. Hydrol. Earth Syst. Sci. Discuss. 2019, 1–39. [Google Scholar] [CrossRef]
- Dyrness, C.T. Hydrologic properties of soils on three small watersheds in the western Cascades of Oregon. USDA For. Ser. Res. Note PNW 1969, 111, 17. [Google Scholar]
- Swanson, F.J.; Jones, J.A. Geomorphology and hydrology of the HJ Andrews experimental forest, Blue River, Oregon. F. Guid. to Geol. Process. Cascadia 2002, 36, 289–314. [Google Scholar]
- Swanson, F.J.; James, M.E. Geology and Geomorphology of the HJ Andrews Experimental Forest, Western Cascades, Oregon; U.S. Department of Agriculture: Portland, OR, USA, 1975.
- Jefferson, A.; Grant, G.E.; Lewis, S.L. A River Runs Underneath It: Geological Control of Spring and Channel Systems and Management Implications, Cascade Range, Oregon. In Advancing the Fundamental Sciences: Proceedings of the Forest Service National Earth Sciences Conference, San Diego, CA, USA, 18–22 October 2004; U.S. Forest Service, Pacific Northwest Research: Portland, OR, USA, 2004; Volume 1, pp. 18–22. [Google Scholar]
- Deligne, N.I.; Mckay, D.; Conrey, R.M.; Grant, G.E.; Johnson, E.R.; O’Connor, J.; Sweeney, K. Field-trip guide to mafic volcanism of the Cascade Range in Central Oregon—A volcanic, tectonic, hydrologic, and geomorphic journey. Sci. Investig. Rep. 2017, 110. [Google Scholar] [CrossRef]
- Cashman, K.V.; Deligne, N.I.; Gannett, M.W.; Grant, G.E.; Jefferson, A. Fire and water: Volcanology, geomorphology, and hydrogeology of the Cascade Range, central Oregon. F. Guid. 2009, 15, 539–582. [Google Scholar]
- Gooseff, M.N.; Wondzell, S.M.; McGlynn, B.L.; Science, I.G.; Well-being, H.; Northwest, P.; Forestry, O.; Resources, L. On the relationships among temporal patterns of evapo-transpiration, stream flow and riparian water levels in headwater catchments during baseflow. In Proceeding of the 36th IAH Congress: Integrating Groundwater Science and Human Well-Being, Toyama, Japan, 27–31 October 2008; pp. 1–10. [Google Scholar]
- Wondzell, S.M.; Gooseff, M.N.; McGlynn, B.L. Flow velocity and the hydrologic behavior of streams during baseflow. Geophys. Res. Lett. 2007, 34, L24404. [Google Scholar] [CrossRef]
- Cain, M.R.; Ward, A.S.; Hrachowitz, M. Ecohydrologic separation alters interpreted hydrologic stores and fluxes in a headwater mountain catchment. Hydrol. Process. 2019, 33, 2658–2675. [Google Scholar] [CrossRef]
- Barnard, H.R.; Graham, C.B.; Van Verseveld, W.; Brooks, J.; Bond, B.J.; McDonnell, J.J. Mechanistic assessment of hillslope transpiration controls of diel subsurface flow: A steady-state irrigation approach. Ecohydrology 2010, 3, 133–142. [Google Scholar] [CrossRef]
- Perry, T.D.; Jones, J.A. Summer streamflow deficits from regenerating Douglas-fir forest in the Pacific Northwest, USA. Ecohydrology 2017, 10, 1–13. [Google Scholar] [CrossRef]
- Knapp, J.L.A.; Cirpka, O.A. A Critical Assessment of Relating Resazurin–Resorufin Experiments to Reach-Scale Metabolism in Lowland Streams. J. Geophys. Res. Biogeosciences 2018, 123, 3538–3555. [Google Scholar] [CrossRef]
- Danckwerts, P. Continuous flow systems. Distribution of residence times. Chem. Eng. Sci. 1953, 2, 1–13. [Google Scholar] [CrossRef]
- Castro, N.M.; Hornberger, G.M. Surface-subsurface water interactions in an alluviated mountain stream channel. Water Resour. Res. 1991, 27, 1613–1621. [Google Scholar] [CrossRef]
- Kennedy, V.C.; Jackman, A.P.; Zand, S.M.; Zellweger, G.W.; Avanzino, R.J.; Walters, R.A. Transport and concentration controls for chloride, strontium, potassium and lead in Uvas Creek, a small cobble-bed stream in Santa Clara County, California, USA: 2. Mathematical modeling. J. Hydrol. 1984, 75, 67–110. [Google Scholar] [CrossRef]
- Laloy, E.; Vrugt, J.A. High-dimensional posterior exploration of hydrologic models using multiple-try DREAM (ZS) and high-performance computing. Water Resour. Res. 2012, 48, 1–18. [Google Scholar] [CrossRef]
- Vrugt, J.; ter Braak, C.; Diks, C.; Hyman, J.; Robinson, B.; Higdon, D. Accelerating Markov-Chain Monte Carlo simulation by differential evolution with self-adaptive randomized subspace sampling. Int. J. Nonlinear Sci. Numer. Simulatoin 2009, 10, 273–290. [Google Scholar] [CrossRef]
- Ward, A.S.; Zarnetske, J.P.; Baranov, V.; Blaen, P.J.; Brekenfeld, N.; Chu, R.; Derelle, R.; Drummond, J.D.; Fleckenstein, J.; Garayburu-Caruso, V.; et al. Co-located contemporaneous mapping of morphological, hydrological, chemical, and biological conditions in a 5th order mountain stream network, Oregon, USA. Earth Syst. Sci. Data Discuss. 2019, 1–27. [Google Scholar] [CrossRef]
- Argerich, A.; Haggerty, R.; Johnson, S.L.; Wondzell, S.M.; Dosch, N.; Corson-rikert, H.; Ashkenas, L.R.; Pennington, R.; Thomas, C.K. Comprehensive multiyear carbon budget of a temperate headwater stream. J. Geophys. Res. - Biogeosciences 2016, 121, 1306–1315. [Google Scholar] [CrossRef]
- Roche, K.R.; Shogren, A.J.; Aubeneau, A.; Tank, J.L.; Bolster, D. Modeling Benthic Versus Hyporheic Nutrient Uptake in Unshaded Streams With Varying Substrates. J. Geophys. Res. Biogeosciences 2019, 124, 367–383. [Google Scholar] [CrossRef]
- Schaper, J.L.; Posselt, M.; Bouchez, C.; Jaeger, A.; Nuetzmann, G.; Putschew, A.; Singer, G.; Lewandowski, J. Fate of Trace Organic Compounds in the Hyporheic Zone: Influence of Retardation, the Benthic Biolayer, and Organic Carbon. Environ. Sci. Technol. 2019, 53, 4224–4234. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.W.; Wagner, B.J. Quantifying hydrologic interactions between streams and their subsurface hyporheic zones. In Streams and Ground Waters; Jones, J.B., Mulholland, P.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; pp. 3–44. [Google Scholar]
- Briggs, M.A.; Day-Lewis, F.D.; Zarnetske, J.P.; Harvey, J.W. A physical explanation for the development of redox microzones in hyporheic flow. Geophys. Res. Lett. 2015, 42, 4402–4410. [Google Scholar] [CrossRef]
- Rosemond, A.D.; Benstead, J.P.; Bumpers, P.M.; Gulis, V.; Kominoski, J.S.; Manning, D.W.P.; Suberkropp, K.; Wallace, J.B. Experimental nutrient additions accelerate terrestrial carbon loss from stream ecosystems. Science (80-. ). 2015, 347, 1142–1144. [Google Scholar] [CrossRef] [PubMed]
- Vanderbilt, K.L.; Lajtha, K.; Swanson, F.J. Biogeochemistry of unpolluted forested watersheds in the Oregon Cascades: Temporal patterns of precipitation and stream nitrogen fluxe. Biogeochemistry 2003, 62, 87–118. [Google Scholar] [CrossRef]
- Corson-Rikert, H.A.; Wondzell, S.M.; Haggerty, R.; Santelmann, M. V Carbon dynamics in the hyporheic zone of a headwater mountain streamin the Cascade Mountains, Oregon. Water Resour. Res. 2016, 52, 7556–7576. [Google Scholar] [CrossRef]
- Webster, J.R.; Benfield, E.F.; Ehrman, T.P.; Schaeffer, M.A.; Tank, J.E.; Hutchens, J.J.; D’Angelo, D.J. What happens to allochthonous material that falls into streams? A synthesis of new and published information from Coweeta. Freshw. Biol. 1999, 41, 687–705. [Google Scholar] [CrossRef]
- Harvey, J.W.; Wagner, B.J.; Bencala, K.E. By Was S ); Hyporheic Exchange Persisted When Base Flow Was Decreasing By. Water Resour. 1996, 32, 2441–2451. [Google Scholar] [CrossRef]
- Reynolds, K.N.; Loecke, T.D.; Burgin, A.J.; Davis, C.A.; Riveros-Iregui, D.; Thomas, S.A.; St. Clair, M.A.; Ward, A.S. Optimizing Sampling Strategies for Riverine Nitrate Using High-Frequency Data in Agricultural Watersheds. Environ. Sci. Technol. 2016, acs.est.5b05423. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Injection Code | Date and Time | NaCl (g) | Ura (g) | Raz (g) | Q at Release (L s−1) |
|---|---|---|---|---|---|
| Falling | 17:15 29-July-2016 | 899.8 | 2.504 | 8.003 | 0.70 |
| Rising | 05:02 1-August-2016 | 599.0 | 2.504 | 8.005 | 0.59 |
| Peak | 10:39 3-August-2016 | 601.2 | 2.495 | 8.002 | 0.76 |
| Minimum | 23:52 5-August-2016 | 698.8 | 2.501 | 7.999 | 0.24 |
| Discharge at Time of Injection | tpeak (h) | t99 (h) | M1 (h) | CV | Skewness | Apparent Dispersion (×104 m2 h−1) | Holdback |
|---|---|---|---|---|---|---|---|
| Falling | 2.3 | 55.3 | 14.3 | 0.85 | 1.35 | 1.55 | 0.61 |
| Rising | 1.7 | 45.0 | 8.5 | 1.04 | 1.80 | 1.39 | 0.68 |
| Peak | 1.4 | 39.3 | 8.2 | 1.01 | 1.48 | 1.26 | 0.68 |
| Minimum | 7.9 | 53.5 | 15.3 | 0.70 | 1.46 | 1.15 | 0.67 |
| Parameter Values | Parameter Sensitivity | |||
|---|---|---|---|---|
| Parameter | Reference Value | Value at Q10 | Value at Q90 | 1/kX * |
| Storage offset, Smin (L) | 3336 (3288, 3617) | 3728 | 2981 | –1.25 (–1.61, –1.33) |
| Mean of gamma distribution, (L) | 29,063 (28,981, 29,713) | 37,406 | 22,513 | –1.10 (–1.11, –1.02) |
| Gamma distribution shape parameter, (unitless) | 1.63 (1.57, 1.64) | 2.50 | 1.06 | –0.645 (–0.717, –0.632) |
| Underflow discharge, (L s−1) | 0.390 (0.387, 0.405) | 0.954 | 0.158 | –1.03E-3 (–1.06E-3, –1.00E-3) |
| Underflow scale parameter, (L) | 36,305 (35,945, 38,837) | 73,414 | 17,802 | –0.393 (–0.403, –0.358) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ward, A.S.; Kurz, M.J.; Schmadel, N.M.; Knapp, J.L.A.; Blaen, P.J.; Harman, C.J.; Drummond, J.D.; Hannah, D.M.; Krause, S.; Li, A.; et al. Solute Transport and Transformation in an Intermittent, Headwater Mountain Stream with Diurnal Discharge Fluctuations. Water 2019, 11, 2208. https://doi.org/10.3390/w11112208
Ward AS, Kurz MJ, Schmadel NM, Knapp JLA, Blaen PJ, Harman CJ, Drummond JD, Hannah DM, Krause S, Li A, et al. Solute Transport and Transformation in an Intermittent, Headwater Mountain Stream with Diurnal Discharge Fluctuations. Water. 2019; 11(11):2208. https://doi.org/10.3390/w11112208
Chicago/Turabian StyleWard, Adam S., Marie J. Kurz, Noah M. Schmadel, Julia L.A. Knapp, Phillip J. Blaen, Ciaran J. Harman, Jennifer D. Drummond, David M. Hannah, Stefan Krause, Angang Li, and et al. 2019. "Solute Transport and Transformation in an Intermittent, Headwater Mountain Stream with Diurnal Discharge Fluctuations" Water 11, no. 11: 2208. https://doi.org/10.3390/w11112208
APA StyleWard, A. S., Kurz, M. J., Schmadel, N. M., Knapp, J. L. A., Blaen, P. J., Harman, C. J., Drummond, J. D., Hannah, D. M., Krause, S., Li, A., Marti, E., Milner, A., Miller, M., Neil, K., Plont, S., Packman, A. I., Wisnoski, N. I., Wondzell, S. M., & Zarnetske, J. P. (2019). Solute Transport and Transformation in an Intermittent, Headwater Mountain Stream with Diurnal Discharge Fluctuations. Water, 11(11), 2208. https://doi.org/10.3390/w11112208