


Interactive Effects of Elevated CO2, Temperature and Drought on Wheat–Aphid Dynamics
 , , , ,
, , , ,
Abstract
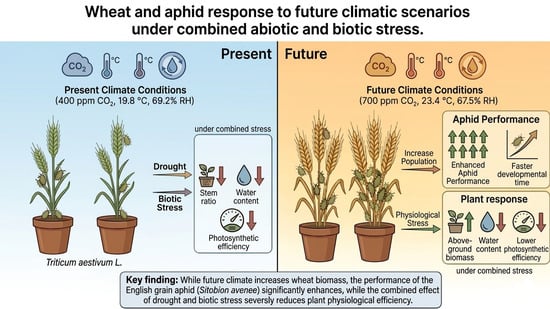
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Data Collection
3. Results
3.1. Aphid Performance
3.2. Plant Physiological Response
3.3. Chlorophyll Florescence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Aphid drought |
| AWS | Automatic weather station |
| GLM | Generalized linear model |
| HSP | Heat shock protein |
| IPCC | Intergovernmental panel on climate change |
| LAR | Leaf area ratio |
| LIST | Luxembourg Institute of Science and Technology |
| PEA | Plant efficiency analyser |
| PPFD | Photosynthetic photon flux density |
| PSII | Photosystem II |
| RH | Relative humidity |
| VWC | Volumetric water content |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SOV | DF | SS | MS | F-Value | Probability |
|---|---|---|---|---|---|
| Treatment (T) | 1 | 2.161 | 2.161 | 1.07 | 0.3069 |
| Climate (C) | 1 | 58.018 | 58.018 | 28.60 | 0.0000 |
| T × C | 1 | 0.875 | 0.875 | 0.43 | 0.514 |
| Error | 52 | 105.50 | 2.029 | ||
| Total | 55 | 166.554 |
| SOV | DF | SS | MS | F-Value | Probability |
|---|---|---|---|---|---|
| Treatment (T) | 1 | 0.446 | 0.446 | 0.22 | 0.6782 |
| Climate (C) | 1 | 50.161 | 50.161 | 24.31 | 0.0000 |
| T × C | 1 | 0.161 | 0.161 | 0.06 | 0.801 |
| Error | 52 | 133.357 | 2.56 | ||
| Total | 55 | 184.125 |
References
- IPCC. Climate Change 2021: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Perkins-Kirkpatrick, S.E.; Lewis, S.C. Increasing Trends in Regional Heatwaves. Nat. Commun. 2020, 11, 3357. [Google Scholar] [CrossRef]
- Bibi, F.; Rahman, A. An Overview of Climate Change Impacts on Agriculture and Their Mitigation Strategies. Agriculture 2023, 13, 1508. [Google Scholar] [CrossRef]
- Malhi, G.; Kaur, M.; Kaushik, P. Impact of Climate Change on Agriculture and Its Mitigation Strategies: A Review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Erenstein, O.; Jaleta, M.; Mottaleb, K.A.; Sonder, K.; Donovan, J.; Braun, H.-J. Global Trends in Wheat Production, Consumption and Trade. In Wheat Improvement; Reynolds, M.P., Braun, H.-J., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 47–66. ISBN 978-3-030-90672-6. [Google Scholar]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature Increase Reduces Global Yields of Major Crops in Four Independent Estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef]
- Zhang, K.-X.; Li, H.-Y.; Quandahor, P.; Gou, Y.-P.; Li, C.-C.; Zhang, Q.-Y.; Haq, I.U.; Ma, Y.; Liu, C.-Z. Responses of Six Wheat Cultivars (Triticum aestivum) to Wheat Aphid (Sitobion avenae) Infestation. Insects 2022, 13, 508. [Google Scholar] [CrossRef]
- Larsson, H. A Crop Loss Model and Economic Thresholds for the Grain Aphid, Sitobion avenae (F.), in Winter Wheat in Southern Sweden. Crop Prot. 2005, 24, 397–405. [Google Scholar] [CrossRef]
- Dampc, J.; Mołoń, M.; Durak, T.; Durak, R. Changes in Aphid—Plant Interactions under Increased Temperature. Biology 2021, 10, 480. [Google Scholar] [CrossRef]
- Milenovic, M.; Eickermann, M.; Junk, J.; Rapisarda, C. Life History Parameters of Bemisia Tabaci MED (Hemiptera: Aleyrodidae) in the Present and Future Climate of Central Europe, Predicted by Physically Realistic Climatic Chamber Simulation. Environ. Entomol. 2023, 52, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Beetge, L.; Krüger, K. Drought and Heat Waves Associated with Climate Change Affect Performance of the Potato Aphid Macrosiphum Euphorbiae. Sci. Rep. 2019, 9, 3645. [Google Scholar] [CrossRef] [PubMed]
- Milenovic, M.; Ripamonti, M.; Eickermann, M.; Rapisarda, C.; Junk, J. Longevity of the Whitefly Parasitoid Eretmocerus Eremicus under Two Different Climate Scenarios. Phytoparasitica 2023, 51, 1041–1046. [Google Scholar] [CrossRef]
- Rao, M.S.; Ranga Rao, G.V.; Venkateswarlu, B. Impact of Climate Change on Insect Pests. In Proceedings of the National Symposium on Climate Change and Rainfed Agriculture, CRIDA, Hyderabad, India, 18–20 February 2010. [Google Scholar]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The Impact of Climate Change on Agricultural Insect Pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The Response of Photosynthesis and Stomatal Conductance to Rising [CO2]: Mechanisms and Environmental Interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What Have We Learned from 15 Years of Free-air CO2 Enrichment (FACE)? A Meta-analytic Review of the Responses of Photosynthesis, Canopy Properties and Plant Production to Rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef]
- Fitzgerald, G.J.; Tausz, M.; O’Leary, G.; Mollah, M.R.; Tausz-Posch, S.; Seneweera, S.; Mock, I.; Löw, M.; Partington, D.L.; McNeil, D.; et al. Elevated Atmospheric [CO2] Can Dramatically Increase Wheat Yields in Semi-arid Environments and Buffer against Heat Waves. Glob. Change Biol. 2016, 22, 2269–2284. [Google Scholar] [CrossRef]
- Sun, Y.; Guo, H.; Ge, F. Plant–Aphid Interactions Under Elevated CO2: Some Cues from Aphid Feeding Behavior. Front. Plant Sci. 2016, 7, 502. [Google Scholar] [CrossRef] [PubMed]
- Huberty, A.F.; Denno, R.F. Plant water stress and its consequences for herbivorous insects: A new synthesis. Ecology 2004, 85, 1383–1398. [Google Scholar] [CrossRef]
- Strock, C. Protocol for Extracting Basic Color Metrics from Images in ImageJ/Fiji 2021. Available online: https://zenodo.org/records/5595203 (accessed on 10 May 2026).
- Radford, P.J. Growth Analysis Formulae: Their Use and Abuse. Crop Sci. 1967, 7, 171–175. [Google Scholar] [CrossRef]
- Reddy, P.P. Impacts on Insect and Mite Pests. In Climate Resilient Agriculture for Ensuring Food Security; Springer: New Delhi, India, 2015; pp. 115–150. ISBN 978-81-322-2198-2. [Google Scholar]
- Moreno-Delafuente, A.; Viñuela, E.; Fereres, A.; Medina, P.; Trębicki, P. Simultaneous Increase in CO2 and Temperature Alters Wheat Growth and Aphid Performance Differently Depending on Virus Infection. Insects 2020, 11, 459. [Google Scholar] [CrossRef]
- Leybourne, D.J.; Preedy, K.F.; Valentine, T.A.; Bos, J.I.B.; Karley, A.J. Drought Has Negative Consequences on Aphid Fitness and Plant Vigor: Insights from a Meta-analysis. Ecol. Evol. 2021, 11, 11915–11929. [Google Scholar] [CrossRef]
- Tabao, N.S.C.; Mendoza, B.C.; Pedro, M.S.; Monsalud, R.G. Mycobiome in Soils from Irrigated, Rice-Based Farming Systems in Apalit, Pampanga and Banaue, Ifugao, Philippines: Diversity and Potential Agro-Biotechnological Applications Determined Using Targeted Metagenomics. Int. J. Agric. Technol. 2023, 19, 781–804. [Google Scholar]
- Xie, H.; Shi, J.; Shi, F.; Xu, H.; He, K.; Wang, Z. Aphid Fecundity and Defenses in Wheat Exposed to a Combination of Heat and Drought Stress. J. Exp. Bot. 2020, 71, 2713–2722. [Google Scholar] [CrossRef] [PubMed]
- Martén-Rodríguez, S.; Cristobal-Pérez, E.J.; De Santiago-Hernández, M.H.; Huerta-Ramos, G.; Clemente-Martínez, L.; Krupnick, G.; Taylor, O.; Lopezaraiza-Mikel, M.; Balvino-Olvera, F.J.; Sentíes-Aguilar, E.M.; et al. Untangling the Complexity of Climate Change Effects on Plant Reproductive Traits and Pollinators: A Systematic Global Synthesis. Glob. Change Biol. 2025, 31, e70081. [Google Scholar] [CrossRef] [PubMed]
- Bierwirth, P.N. Long-term Carbon Dioxide Toxicity and Climate Change: A Critical Unapprehended Risk for Human Health. ResearchGate 2026. [Google Scholar] [CrossRef]
- Wang, J.; Han, Y.; Li, H.; Bai, H.; Liang, H.; Zong, Y.; Zhang, D.; Shi, X.; Li, P.; Hao, X. Elevated CO2 Concentration Extends Reproductive Growth Period and Enhances Carbon Metabolism in Wheat Exposed to Increased Temperature. Plant Cell Environ. 2025, 48, 1452–1467. [Google Scholar] [CrossRef]
- Zhang, D.; Li, A.; Lam, S.K.; Li, P.; Zong, Y.; Gao, Z.; Hao, X. Increased Carbon Uptake under Elevated CO2 Concentration Enhances Water-Use Efficiency of C4 Broomcorn Millet under Drought. Agric. Water Manag. 2021, 245, 106631. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Zohaib, A.; Tanveer, M.; Naeem, M.; Ali, I.; Tabassum, T.; Nazir, U. Growth and Developmental Responses of Crop Plants under Drought Stress: A Review. Zemdirbyste-Agriculture 2017, 104, 267–276. [Google Scholar] [CrossRef]
- Li, C.; Sun, Q.; Gou, Y.; Zhang, K.; Zhang, Q.; Zhou, J.-J.; Liu, C. Long-Term Effect of Elevated CO2 on the Development and Nutrition Contents of the Pea Aphid (Acyrthosiphon pisum). Front. Physiol. 2021, 12, 688220. [Google Scholar] [CrossRef]
- Saha, S.; Das, B.; Chatterjee, D.; Sehgal, V.K.; Chakraborty, D.; Pal, M. Crop Growth Responses Towards Elevated Atmospheric CO2. In Plant Ecophysiology and Adaptation Under Climate Change: Mechanisms and Perspectives I; Hasanuzzaman, M., Ed.; Springer: Singapore, 2020; pp. 147–198. ISBN 978-981-15-2155-3. [Google Scholar]
- Hao, L.K.; Batyrshina, Z.; Goldstein, Y.; Mishra, A.; Köllner, T.G.; Yaakov, B.; Tzin, V. Dual Role of Benzoxazinoids in Plant Response to Combined Drought Stress and Aphid Feeding. Plant Biol. 2026, 28, 45–57. [Google Scholar] [CrossRef]
- Van Der Kooi, C.J.; Reich, M.; Löw, M.; De Kok, L.J.; Tausz, M. Growth and Yield Stimulation under Elevated CO2 and Drought: A Meta-Analysis on Crops. Environ. Exp. Bot. 2016, 122, 150–157. [Google Scholar] [CrossRef]
- Shanker, A.K.; Gunnapaneni, D.; Bhanu, D.; Vanaja, M.; Lakshmi, N.J.; Yadav, S.K.; Prabhakar, M.; Singh, V.K. Elevated CO2 and Water Stress in Combination in Plants: Brothers in Arms or Partners in Crime? Biology 2022, 11, 1330. [Google Scholar] [CrossRef]
- Yang, L.; Duan, J.; Liu, Y.; Hu, W.; Liu, X.; Wang, Y.; Zhou, Z.; Zhao, W. Changes in Carbohydrate Distribution in Cotton Photosynthetic Organs and Increase in Boll Weight Reduce Yield Loss under High Temperature. J. Exp. Bot. 2024, 75, 3483–3499. [Google Scholar] [CrossRef]
- Ben Mariem, S.; Soba, D.; Zhou, B.; Loladze, I.; Morales, F.; Aranjuelo, I. Climate Change, Crop Yields, and Grain Quality of C3 Cereals: A Meta-Analysis of [CO2], Temperature, and Drought Effects. Plants 2021, 10, 1052. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Rostás, M.; Vosteen, I. Drought Aggravates Plant Stress by Favouring Aphids and Weakening Indirect Defense in a Sugar Beet Tritrophic System. J. Pest Sci. 2025, 98, 549–564. [Google Scholar] [CrossRef]
- Wang, X.; Liu, F. Effects of Elevated CO2 and Heat on Wheat Grain Quality. Plants 2021, 10, 1027. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.-T.; Moutinho-Pereira, J.; Correia, C.M. Drought Stress Effects and Olive Tree Acclimation under a Changing Climate. Plants 2019, 8, 232. [Google Scholar] [CrossRef]
- Du, J.-L.; Wu, D.-G.; Li, J.-Q.; Zhan, Q.-W.; Huang, S.-C.; Huang, B.-H.; Wang, X. Effects of Aphid Disoperation on Photosynthetic Performance and Agronomic Traits of Different Sorghum Varieties. Pak. J. Bot. 2021, 53, 2275–2285. [Google Scholar] [CrossRef] [PubMed]
- Sonbol, H.; Korany, S.M.; Nhs, M.; Abdi, I.; Maridueña-Zavala, M.G.; Alsherif, E.A.; Aldailami, D.A.; Elsheikh, S.Y.S. Exploring the Benefits of AMF Colonization for Improving Wheat Growth, Physiology and Metabolism, and Antimicrobial Activity under Biotic Stress from Aphid Infection. BMC Plant Biol. 2025, 25, 198. [Google Scholar] [CrossRef] [PubMed]
- Sommer, S.G.; Han, E.; Li, X.; Rosenqvist, E.; Liu, F. The Chlorophyll Fluorescence Parameter Fv/Fm Correlates with Loss of Grain Yield after Severe Drought in Three Wheat Genotypes Grown at Two CO2 Concentrations. Plants 2023, 12, 436. [Google Scholar] [CrossRef]
- Cao, Y.; Yang, K.; Liu, W.; Feng, G.; Peng, Y.; Li, Z. Adaptive Responses of Common and Hybrid Bermudagrasses to Shade Stress Associated with Changes in Morphology, Photosynthesis, and Secondary Metabolites. Front. Plant Sci. 2022, 13, 817105. [Google Scholar] [CrossRef]
- Chachar, S.; Ahmed, N.; Hu, X. Drought-induced Aesthetic Decline and Ecological Impacts on Ornamentals: Mechanisms of Damage and Innovative Strategies for Mitigation. Plant Biol. 2025, 28, 578–600. [Google Scholar] [CrossRef]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Ashraful Alam, M.; Syed, M.A.; Hossain, J.; Sarkar, S.; Saha, S.; Bhadra, P.; et al. Consequences and Mitigation Strategies of Abiotic Stresses in Wheat (Triticum aestivum L.) under the Changing Climate. Agronomy 2021, 11, 241. [Google Scholar] [CrossRef]






| Trait | Treatment | Climate | Mean | Median | Q1 | Q3 | IQR |
|---|---|---|---|---|---|---|---|
| Pre-reproductive period (days) | Aphid | Present | 13 | 13 | 12 | 14 | 2 |
| Aphid + Drought | 13 | 12.5 | 12 | 14 | 2 | ||
| Aphid | Future | 11 | 10.5 | 10 | 11 | 1 | |
| Aphid + Drought | 10 | 10 | 10 | 11 | 1 | ||
| Reproductive period (days) | Aphid | Present | 10 | 10 | 9 | 11 | 2 |
| Aphid + Drought | 10 | 10.5 | 9 | 11 | 2 | ||
| Aphid | Future | 12 | 12.5 | 12 | 13 | 1 | |
| Aphid + Drought | 12 | 12 | 12 | 13 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Javed, A.; Ahmad, M.N.; Shah, S.; Eickermann, M.; Ripamonti, M.; Seeburger, P.; Junk, J. Interactive Effects of Elevated CO2, Temperature and Drought on Wheat–Aphid Dynamics. Atmosphere 2026, 17, 498. https://doi.org/10.3390/atmos17050498
Javed A, Ahmad MN, Shah S, Eickermann M, Ripamonti M, Seeburger P, Junk J. Interactive Effects of Elevated CO2, Temperature and Drought on Wheat–Aphid Dynamics. Atmosphere. 2026; 17(5):498. https://doi.org/10.3390/atmos17050498
Chicago/Turabian StyleJaved, Amina, Muhammad Nauman Ahmad, Shahen Shah, Michael Eickermann, Matteo Ripamonti, Pauline Seeburger, and Jürgen Junk. 2026. "Interactive Effects of Elevated CO2, Temperature and Drought on Wheat–Aphid Dynamics" Atmosphere 17, no. 5: 498. https://doi.org/10.3390/atmos17050498
APA StyleJaved, A., Ahmad, M. N., Shah, S., Eickermann, M., Ripamonti, M., Seeburger, P., & Junk, J. (2026). Interactive Effects of Elevated CO2, Temperature and Drought on Wheat–Aphid Dynamics. Atmosphere, 17(5), 498. https://doi.org/10.3390/atmos17050498