
Evaluation of Time Delay and Cumulative Effects of Meteorological Drought on Net Primary Productivity of Vegetation in the Upper Reaches of the Yellow River, China


Abstract
1. Introduction
2. Data and Methodology
2.1. Study Area Overview
2.2. Data Sources and Preparation Steps
2.2.1. Station Meteorological Data
2.2.2. NDVI Data
2.2.3. Vegetation Type Data
2.2.4. Solar Radiation, Temperature and Precipitation Data
2.2.5. Annual NPP Data
2.3. Research Approach
2.3.1. NPP Calculation
2.3.2. Sen’s Trend Estimation and the Mann–Kendall Statistical Test
2.3.3. Mutation Test Methodology
2.3.4. Delay Effect of Drought on NPP
2.3.5. Cumulative Effect of Drought on NPP
2.4. Data Processing
3. Results
3.1. NPP Precision Assessment
3.2. Temporal and Spatial Variation Characteristics of NPP
3.2.1. Temporal Trends and Spatial Patterns of Annual NPP
3.2.2. Temporal Trends and Spatial Patterns of Seasonal NPP
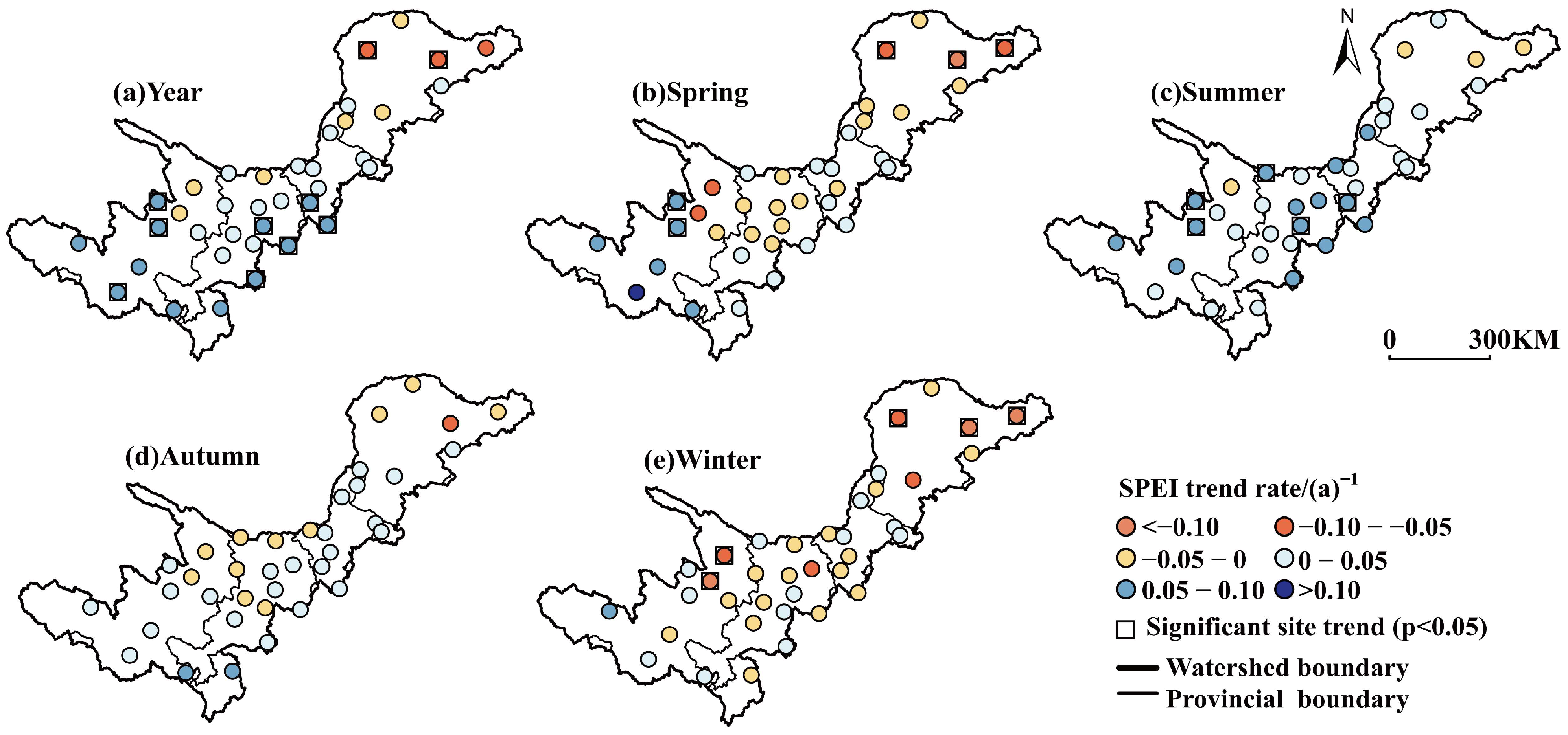
3.3. Temporal and Spatial Distribution of Drought Index
3.4. Delay Effect of Drought on NPP in Growing Season
3.4.1. Response of NPP to Drought at Different Lag Scales
3.4.2. Spatial Distribution of Delay Effect of Drought on NPP
3.4.3. Delay Effect of Drought on NPP Under Different Water Conditions
3.5. Cumulative Effect of Drought on NPP in Growing Season
3.5.1. Response of NPP to Drought at Different Cumulative Scales
3.5.2. Spatial Distribution of the Cumulative Effect of Drought on NPP
3.5.3. Cumulative Effect of Drought on NPP Under Different Water Conditions
4. Discussion
5. Conclusions
- (1)
- From 2001 to 2022, the mean annual NPP rose with an increase of 2.7538 g/(m2·a). Within the study region, NPP increased significantly (47.5%), while NPP decreased significantly (0.85%). The four-season NPP growth slope is summer > autumn > spring > winter. In the past 22 years, NPP in spring, summer and autumn has been significantly improved.
- (2)
- From 2001 to 2022, the interannual variation and regional distribution of seasonal and annual SPEI in the upstream region showed a trend of wetting in summer, autumn and annual scales, except spring and winter.
- (3)
- Drought has a lag effect on NPP in 82.58% of the upstream region during the growing season. Mainly concentrated in 0–2 months, the 1–2 months occupy most areas (36.07% and 17.67%). When drought intensifies in the region, the influence of drought conditions on NPP has only a short time lag.
- (4)
- NPP in 66.14% of the region exhibited a cumulative effect in response to drought, primarily occurring over short time scales, with March and April being the key periods. As drought conditions worsened, the influence of both lag and cumulative effects on NPP became more pronounced.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, X.; Liao, X.; Di, D.; Shi, W. A Review of Drought Disturbance on Socioeconomic Development. Water 2023, 15, 3912. [Google Scholar] [CrossRef]
- Mani, Z.A.; Khorram-Manesh, A.; Goniewicz, K. Global health emergencies of extreme drought events: Historical impacts and future preparedness. Atmosphere 2024, 15, 1137. [Google Scholar] [CrossRef]
- Dai, R.; Huang, J.; Chen, Z.; Zhou, J.; Havea, P.H. Impacts of anthropogenic climate change on meteorological drought in China. Front. Earth Sci. 2024, 12, 1369523. [Google Scholar] [CrossRef]
- Zhou, T. New physical science behind climate change: What does IPCC AR6 tell us? Innovation 2021, 2, 100173. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Z.; Yang, H.; Zhao, Y.; Zhang, Z.; Li, Z.; Hussain, Z. Copula-based drought analysis using Standardized Precipitation Evapotranspiration Index: A case study in the Yellow river basin, China. Water 2019, 11, 1298. [Google Scholar] [CrossRef]
- Gajendiran, K.; Kandasamy, S.; Narayanan, M. Influences of wildfire on the forest ecosystem and climate change: A comprehensive study. Environ. Res. 2024, 240, 117537. [Google Scholar] [CrossRef]
- Crocetti, L.; Forkel, M.; Fischer, M.; Jurečka, F.; Grlj, A.V.S.; Salentinig, A.; Trnka, M.; Anderson, M.; Ng, W.; Kokalj, Ž.; et al. Earth Observation for agricultural drought monitoring in the Pannonian Basin (southeastern Europe): Current state and future directions. Reg. Environ. Change 2020, 20, 123. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, Z.; Zhu, M.; Wan, J.; Wu, X.; Liu, R.; Zheng, J. Study on the Response of Net Primary Productivity to Vegetation Phenology and Its Influencing Factors in Karst Ecologically Fragile Regions. Atmosphere 2024, 15, 1464. [Google Scholar] [CrossRef]
- Lin, Z.; Liu, Y.; Wen, Z.; Chen, X.; Han, P.; Zheng, C.; Yao, H.; Wang, Z.; Shi, H. Spatial--temporal variation characteristics and driving factors of net primary production in the Yellow River Basin over multiple time scales. Remote Sens. 2023, 15, 5273. [Google Scholar] [CrossRef]
- Xue, H.; Chen, Y.; Dong, G.; Li, J. Quantitative analysis of spatiotemporal changes and driving forces of vegetation net primary productivity (NPP) in the Qimeng region of Inner Mongolia. Ecol. Indic. 2023, 154, 110610. [Google Scholar] [CrossRef]
- PIAO, S.; FANG, J.; GUO, Q. Application of CASA model to the estimation of Chinese terrestrial net primary productivity. Chin. J. Plant Ecol. 2001, 25, 603. [Google Scholar]
- Yang, A.; Zhang, H.; Yang, X.; Zhang, X. Quantitative analysis of the impact of climate change and human activities on vegetation NPP in the Qilian Mountain. Hum. Ecol. Risk Assess. Int. J. 2023, 29, 202–221. [Google Scholar] [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef]
- Ma, R.; Xia, C.; Liu, Y.; Wang, Y.; Zhang, J.; Shen, X.; Lu, X.; Jiang, M. Spatiotemporal change of net primary productivity and its response to climate change in temperate grasslands of China. Front. Plant Sci. 2022, 13, 899800. [Google Scholar] [CrossRef]
- Svoboda, M.; Hayes, M.; Wood, D. Standardized Precipitation Index: User Guide; United States National Drought Mitigation Center: Lincoln, Nebraska, 2012; pp. 3–15. [Google Scholar]
- Alley, W.M. The Palmer drought severity index: Limitations and assumptions. J. Clim. Appl. Meteorol. 1984, 23, 1100–1109. [Google Scholar] [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef]
- Liu, C.; Yang, C.; Yang, Q.; Wang, J. Spatiotemporal drought analysis by the standardized precipitation index (SPI) and standardized precipitation evapotranspiration index (SPEI) in Sichuan Province, China. Sci. Rep. 2021, 11, 1280. [Google Scholar] [CrossRef]
- Dikshit, A.; Pradhan, B.; Huete, A. An improved SPEI drought forecasting approach using the long short-term memory neural network. J. Environ. Manag. 2021, 283, 111979. [Google Scholar] [CrossRef]
- Politi, N.; Vlachogiannis, D.; Sfetsos, A.; Nastos, P.T.; Dalezios, N.R. High resolution future projections of drought characteristics in Greece based on SPI and SPEI indices. Atmosphere 2022, 13, 1468. [Google Scholar] [CrossRef]
- Xu, C.; McDowell, N.G.; Fisher, R.A.; Wei, L.; Sevanto, S.; Christoffersen, B.O.; Weng, E.; Middleton, R.S. Increasing impacts of extreme droughts on vegetation productivity under climate change. Nat. Clim. Change 2019, 9, 948–953. [Google Scholar] [CrossRef]
- Sun, M.; Li, X.; Xu, H.; Wang, K.; Anniwaer, N.; Hong, S. Drought thresholds that impact vegetation reveal the divergent responses of vegetation growth to drought across China. Glob. Change Biol. 2024, 30, e16998. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.I.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef]
- Huang, W.; Wang, W.; Cao, M.; Fu, G.; Xia, J.; Wang, Z.; Li, J. Local climate and biodiversity affect the stability of China’s grasslands in response to drought. Sci. Total Environ. 2021, 768, 145482. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, R.; Bento, V.I.L.A.; Leng, S.; Qi, J.; Zeng, J.; Wang, Q. The effect of drought on vegetation gross primary productivity under different vegetation types across China from 2001 to 2020. Remote Sens. 2022, 14, 4658. [Google Scholar] [CrossRef]
- Geng, G.; Yang, R.; Liu, L. Downscaled solar-induced chlorophyll fluorescence has great potential for monitoring the response of vegetation to drought in the Yellow River Basin, China: Insights from an extreme event. Ecol. Indic. 2022, 138, 108801. [Google Scholar] [CrossRef]
- Kong, D.; Miao, C.; Wu, J.; Zheng, H.; Wu, S. Time lag of vegetation growth on the Loess Plateau in response to climate factors: Estimation, distribution, and influence. Sci. Total Environ. 2020, 744, 140726. [Google Scholar] [CrossRef]
- Lu, J.; Peng, J.; Li, G.; Guan, J.; Han, W.; Liu, L.; Wang, R.; Zheng, J. Assessment of time-lag and cumulative effects of drought on gross primary productivity of grassland in Central Asia from 1982 to 2018. Acta Ecol. Sin. 2023, 43, 9745–9757. [Google Scholar]
- Liu, L.; Guan, J.; Zheng, J.; Wang, Y.; Han, W.; Liu, Y. Cumulative effects of drought have an impact on net primary productivity stability in Central Asian grasslands. J. Environ. Manag. 2023, 344, 118734. [Google Scholar] [CrossRef]
- Wei, X.; He, W.; Zhou, Y.; Ju, W.; Xiao, J.; Li, X.; Liu, Y.; Xu, S.; Bi, W.; Zhang, X.; et al. Global assessment of lagged and cumulative effects of drought on grassland gross primary production. Ecol. Indic. 2022, 136, 108646. [Google Scholar] [CrossRef]
- Qin, Y.; Jin, X.; Du, K.; Jin, Y. Changes in river morphology and influencing factors in the upper Yellow River over the past 25 years. Geomorphology 2024, 465, 109397. [Google Scholar] [CrossRef]
- Jiang, W.; Yuan, L.; Wang, W.; Cao, R.; Zhang, Y.; Shen, W. Spatio-temporal analysis of vegetation variation in the Yellow River Basin. Ecol. Indic. 2015, 51, 117–126. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, J.; Liu, H.; Zhu, Y.; He, G.; Li, W.; Huang, H. Coordinated Development of Water, Food, and Energy in Upper and Middle Reaches of the Yellow River. Strateg. Study Chin. Acad. Eng. 2023, 25, 180–190. [Google Scholar] [CrossRef]
- Cui, Y.; Li, H.; Zheng, L.; Wu, M. Study of ecological environmental quality changes in the upper Yellow River basin based on remote sensing ecological index. J. Desert Res. 2023, 43, 107. [Google Scholar]
- Liu, C.; Zhang, X.; Wang, T.; Chen, G.; Zhu, K.; Wang, Q.; Wang, J. Detection of vegetation coverage changes in the Yellow River Basin from 2003 to 2020. Ecol. Indic. 2022, 138, 108818. [Google Scholar] [CrossRef]
- Mngadi, M.; Odindi, J.; Mutanga, O.; Sibanda, M. Estimating aboveground net primary productivity of reforested trees in an urban landscape using biophysical variables and remotely sensed data. Sci. Total Environ. 2022, 802, 149958. [Google Scholar] [CrossRef]
- Liu, X.; Wang, P.; Song, H.; Zeng, X. Determinants of net primary productivity: Low-carbon development from the perspective of carbon sequestration. Technol. Forecast. Soc. Change 2021, 172, 121006. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, Y.; Li, M.; Wang, Z.; Wu, J.; Zhang, X.; Cao, Y. Impacts of human appropriation of net primary production on ecosystem regulating services in Tibet. Ecosyst. Serv. 2021, 47, 101231. [Google Scholar] [CrossRef]
- Yang, J.; Xie, B.; Zhang, D. Spatial-temporal heterogeneity of ecosystem services trade-off synergy in the Yellow River Basin. J. Desert Res. 2021, 41, 78. [Google Scholar]
- Wu, L.; Zhang, Y.; Luo, G.; Chen, D.; Yang, D.; Yang, Y.; Tian, F. Characteristics of vegetation carbon sink carrying capacity and restoration potential of China in recent 40 years. Front. For. Glob. Change 2023, 6, 1266688. [Google Scholar] [CrossRef]
- Omer, A.; Zhuguo, M.; Yuan, X.; Zheng, Z.; Saleem, F. A hydrological perspective on drought risk-assessment in the Yellow River Basin under future anthropogenic activities. J. Environ. Manag. 2021, 289, 112429. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xie, B.; Tao, W.; Zhang, D. Ecosystem services assessment, trade-off, and bundles in the Yellow River Basin, China. Diversity 2021, 13, 308. [Google Scholar] [CrossRef]
- Yong, X.U.; Chuansheng, W. Ecological protection and high-quality development in the Yellow River Basin: Framework, path, and countermeasure. Bull. Chin. Acad. Sci. 2020, 35, 875–883. [Google Scholar]
- Xie, H.; Tong, X.; Li, J.; Zhang, J.; Liu, P.; Yu, P. Changes of NDVI and EVI and their responses to climatic variables in the Yellow River Basin during the growing season of 2000–2018. Acta Ecol. Sin. 2022, 42, 4536–4549. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration-Guidelines for computing crop water requirements-FAO Irrigation and drainage paper 56. Fao Rome 1998, 300, D05109. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Lu, G.Y.; Wong, D.W. An adaptive inverse-distance weighting spatial interpolation technique. Comput. Geosci. 2008, 34, 1044–1055. [Google Scholar] [CrossRef]
- Zhang, H.; Yin, Y.; An, H.; Lei, J.; Li, M.; Song, J.; Han, W. Surface urban heat island and its relationship with land cover change in five urban agglomerations in China based on GEE. Environ. Sci. Pollut. Res. 2022, 29, 82271–82285. [Google Scholar] [CrossRef]
- Di, Y.; Zhang, G.; You, N.; Yang, T.; Zhang, Q.; Liu, R.; Doughty, R.B.; Zhang, Y. Mapping croplands in the granary of the tibetan plateau using all available landsat imagery, a phenology-based approach, and google earth engine. Remote Sens. 2021, 13, 2289. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 170191. [Google Scholar] [CrossRef]
- Wang, J.; He, H.; Dong, J.; Guo, B. Spatio-temporal distribution of vegetation net primary productivity in the yellow river basin in 2000-2019 and its natural driving factors. J. Desert Res. 2021, 41, 213. [Google Scholar]
- Mao, S.; Shangguan, Z. Evolution of spatiotemporal patterns in vegetation net primary productivity and the driving forces on the Loess Plateau. Front. Environ. Sci. 2023, 11, 1134917. [Google Scholar] [CrossRef]
- Shi, S.; Zhu, L.; Luo, Z.; Qiu, H. Quantitative analysis of the contributions of climatic and anthropogenic factors to the variation in net primary productivity, China. Remote Sens. 2023, 15, 789. [Google Scholar] [CrossRef]
- Zhu, W.; Pan, Y.; Zhang, J. Estimation of net primary productivity of Chinese terrestrial vegetation based on remote sensing. Chin. J. Plant Ecol. 2007, 31, 413. [Google Scholar]
- Mann, H.B. Nonparametric tests against trend. Econom. J. Econom. Soc. 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Xiao, F.; Liu, Q.; Xu, Y. Estimation of terrestrial net primary productivity in the Yellow River Basin of China using light use efficiency model. Sustainability 2022, 14, 7399. [Google Scholar] [CrossRef]
- Liu, S.; Shao, Q.; Ning, J.; Niu, L.; Zhang, X.; Liu, G.; Huang, H. Remote-sensing-based assessment of the ecological restoration degree and restoration potential of ecosystems in the upper yellow river over the past 20 years. Remote Sens. 2022, 14, 3550. [Google Scholar] [CrossRef]
- Zhang, F.; Hu, X.; Zhang, J.; Li, C.; Zhang, Y.; Li, X. Change in Alpine Grassland NPP in response to climate variation and human activities in the Yellow River Source Zone from 2000 to 2020. Sustainability 2022, 14, 8790. [Google Scholar] [CrossRef]
- Ahmed, K.; Shahid, S.; Nawaz, N. Impacts of climate variability and change on seasonal drought characteristics of Pakistan. Atmos. Res. 2018, 214, 364–374. [Google Scholar] [CrossRef]
- Laimighofer, J.; Laaha, G. How standard are standardized drought indices? Uncertainty components for the SPI \& SPEI case. J. Hydrol. 2022, 613, 128385. [Google Scholar]
- Gao, B.L.; Gong, J.; Li, Y.; Jin, T. Analysis of multiscalar characteristics of dry and wet conditions in the Yellow River Basin based on SPEI. AridZone Research 2022, 39, 723–733. [Google Scholar]
- Li, X.; Yu, K.; Xu, G.; Li, P.; Li, Z.; Shi, P. Spatial Characteristics and Driving Factors of Net Primary Productivity of Vegetation in the Upper and Middle Yellow River Basin. Environ. Sci. 2024, 45, 6448–6457. [Google Scholar]
- Wu, X.; Yan, Z.; Yang, H.; Wang, S.; Zhang, H.; Shen, Y.; Song, S.; Liu, Y.; Guo, Y.; Yang, D.; et al. Ecological restoration in the Yellow River Basin enhances hydropower potential. Nat. Commun. 2025, 16, 2566. [Google Scholar] [CrossRef]
- Cao, Y.; Li, H.; Liu, Y.; Zhang, Y.; Jiang, Y.; Dai, W.; Shen, M.; Guo, X.; Qi, W.; Li, L.; et al. Regional contribution and attribution of the interannual variation of net primary production in the yellow River Basin, China. Remote Sens. 2023, 15, 5212. [Google Scholar] [CrossRef]
- Wang, W.; Sun, L.; Luo, Y. Changes in vegetation greenness in the upper and middle reaches of the Yellow River Basin over 2000–2015. Sustainability 2019, 11, 2176. [Google Scholar] [CrossRef]
- Huang, M.; Wang, X.; Keenan, T.F.; Piao, S. Drought timing influences the legacy of tree growth recovery. Glob. Change Biol. 2018, 24, 3546–3559. [Google Scholar] [CrossRef]
- He, W.; Ju, W.; Schwalm, C.R.; Sippel, S.; Wu, X.; He, Q.; Song, L.; Zhang, C.; Li, J.; Sitch, S.; et al. Large-scale droughts responsible for dramatic reductions of terrestrial net carbon uptake over North America in 2011 and 2012. J. Geophys. Res. Biogeosci. 2018, 123, 2053–2071. [Google Scholar] [CrossRef]
- Zhan, C.; Liang, C.; Zhao, L.; Jiang, S.; Niu, K.; Zhang, Y. Drought-related cumulative and time-lag effects on vegetation dynamics across the Yellow River Basin, China. Ecol. Indic. 2022, 143, 109409. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Li, J.; Yan, X.; Yan, X.; Guo, W.; Wang, K.; Qiao, J. Temporal and spatial variation study of vegetation coverage in the Yellow River Basin based on GEE cloud platform. J. China Coal Soc. 2021, 46, 1439–1450. [Google Scholar]
- Wu, C.; Chen, K.; E, C.; You, X.; He, D.; Hu, L.; Liu, B.; Wang, R.; Shi, Y.; Li, C.; et al. Improved CASA model based on satellite remote sensing data: Simulating net primary productivity of Qinghai Lake basin alpine grassland. Geosci. Model Dev. Discuss. 2022, 2022, 1–24. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Moisture Level | SPEI Value |
|---|---|---|
| 1 | Extreme drought (Ed) | ≤−2.0 |
| 2 | Moderate drought (Mod) | (−2.0, −1.0] |
| 3 | Mild drought (Mid) | (−1.0, −0.5] |
| 4 | Normal (N) | (−0.5, 0.5] |
| 5 | Mildly wet (Miw) | (0.5, 1.0] |
| 6 | Moderately wet (Mow) | (1.0, 2.0] |
| 7 | Extremely wet (Ew) | >2.0 |
| Land Use Types | /(gC·MJ−1) |
|---|---|
| Evergreen needleleaf forest | 0.389 |
| Deciduous needleleaf forest | 0.485 |
| Deciduous broadleaf forest | 0.692 |
| Mixed forest | 0.629 |
| Shrubland | 0.429 |
| Grasslands | 0.542 |
| Cultivated land | 0.542 |
| Trend Type | ||
|---|---|---|
| > 0 | 1.96 < | Significantly Increase |
| 1.96 ≥ | Insignificant Increase | |
| = 0 | - | No Trend |
| < 0 | 1.96 < | Significantly Decrease |
| 1.96 ≥ | Insignificant Decrease |
| Trend | Spring | Summer | Autumn | Winter |
|---|---|---|---|---|
| Significantly Decrease (SD) | 2.59% | 0.98% | 0.97% | 0.29% |
| Insignificant Decrease (ID) | 6.48% | 12.22% | 9.94% | 7.90% |
| No Trend (NT) | 6.95% | 5.67% | 7.08% | 53.51% |
| Insignificantly Increase (II) | 36.02% | 48.35% | 42.20% | 19.36% |
| Significantly Increase (SI) | 47.96% | 32.78% | 39.81% | 18.93% |
| % | SPEI1 | SPEI2 | SPEI3 | SPEI4 | SPEI5 | SPEI6 | SPEI7 | SPEI8 | SPEI9 | SPEI10 | SPEI11 | SPEI12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mod (Mow) | 6.8 (7.6) | 7.6 (5.7) | 4.5 (5.3) | 4.9 (4.5) | 2.7 (5.3) | 2.7 (5.7) | 1.5 (4.2) | 1.1 (4.5) | 0.8 (3.8) | 0.8 (4.9) | 0.0 (4.5) | 0.8 (4.5) |
| Ed (Ew) | 0.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| N | 54.9 | 59.5 | 54.9 | 57.6 | 61.4 | 63.3 | 65.9 | 66.7 | 66.3 | 66.3 | 66.3 | 64.0 |
| Mid (Miw) | 17.0 (13.3) | 15.5 (11.7) | 21.6 (13.6) | 18.9 (14.0) | 19.7 (11.0) | 17.0 (11.4) | 16.7 (11.7) | 15.5 (12.1) | 17.4 (11.7) | 15.9 (12.1) | 17.4 (11.7) | 17.8 (12.9) |
| Seasons | Hysteresis Characteristics | Lag Months |
|---|---|---|
| Spring | Average lag time/month | 6 |
| Main lag time/month | 0, 8 | |
| Summer | Average lag time/month | 4 |
| Main lag time/month | 0, 1 | |
| Autumn | Average lag time/month | 5 |
| Main lag time/month | 0, 3 | |
| Winter | Average lag time/month | 6 |
| Main lag time/month | 6, 8 | |
| Growing Season | Average lag time/month | 4 |
| Main lag time/month | 2, 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, H.; Li, Z.; Dong, G.; Wang, H. Evaluation of Time Delay and Cumulative Effects of Meteorological Drought on Net Primary Productivity of Vegetation in the Upper Reaches of the Yellow River, China. Atmosphere 2025, 16, 602. https://doi.org/10.3390/atmos16050602
Xue H, Li Z, Dong G, Wang H. Evaluation of Time Delay and Cumulative Effects of Meteorological Drought on Net Primary Productivity of Vegetation in the Upper Reaches of the Yellow River, China. Atmosphere. 2025; 16(5):602. https://doi.org/10.3390/atmos16050602
Chicago/Turabian StyleXue, Huazhu, Zhi Li, Guotao Dong, and Hao Wang. 2025. "Evaluation of Time Delay and Cumulative Effects of Meteorological Drought on Net Primary Productivity of Vegetation in the Upper Reaches of the Yellow River, China" Atmosphere 16, no. 5: 602. https://doi.org/10.3390/atmos16050602
APA StyleXue, H., Li, Z., Dong, G., & Wang, H. (2025). Evaluation of Time Delay and Cumulative Effects of Meteorological Drought on Net Primary Productivity of Vegetation in the Upper Reaches of the Yellow River, China. Atmosphere, 16(5), 602. https://doi.org/10.3390/atmos16050602