Production of Plant Secondary Metabolites: Examples, Tips and Suggestions for Biotechnologists

 ,
, 

 , , ,
, , ,  ,
, 
Abstract
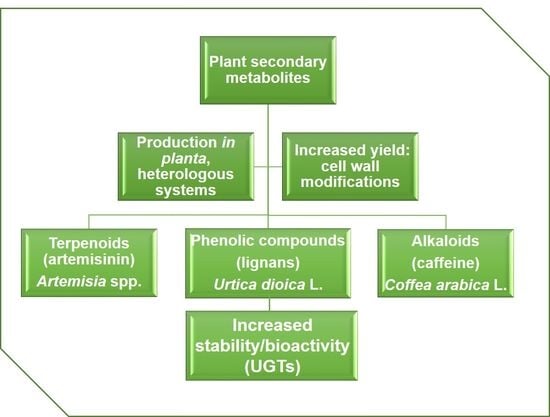
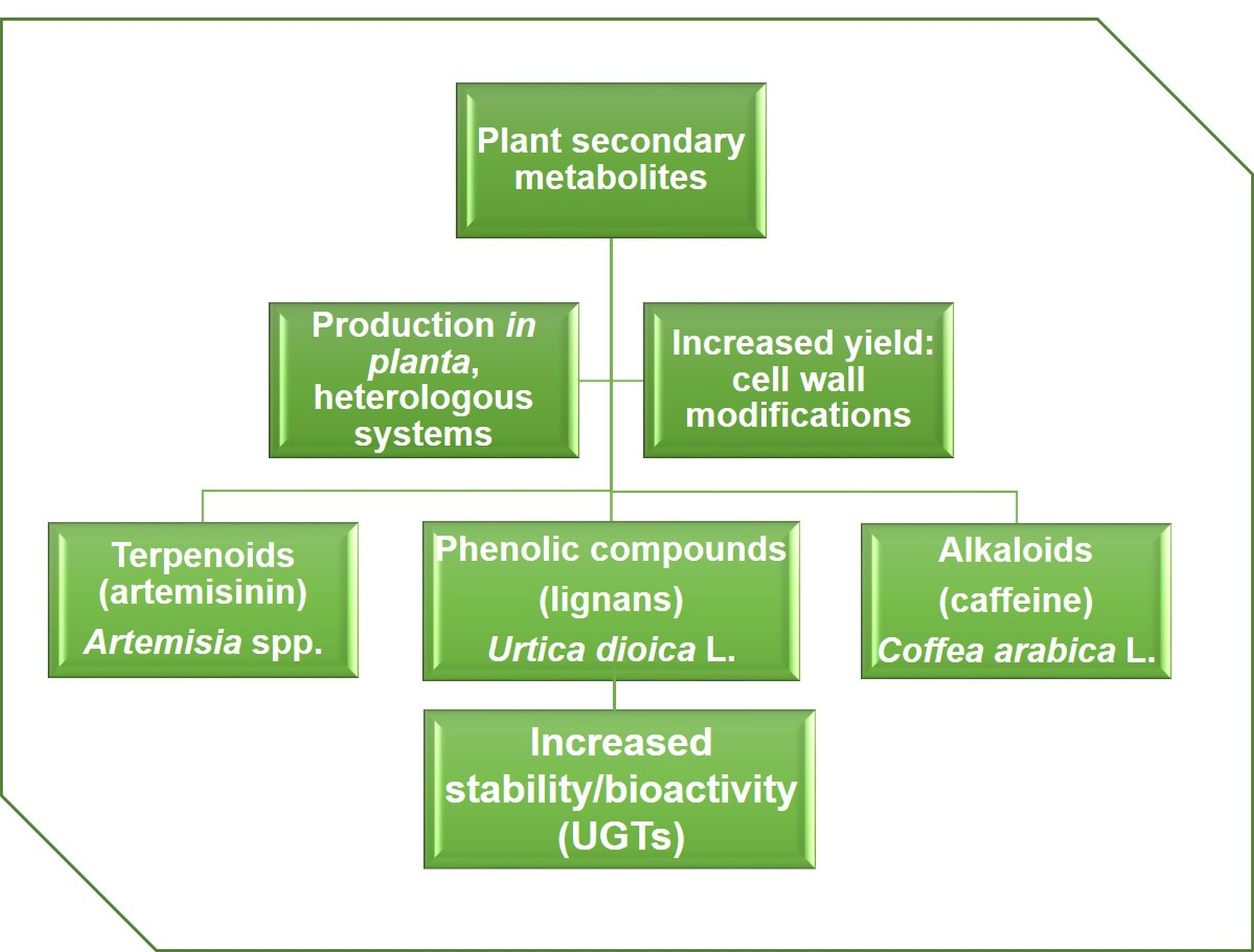
1. Introduction
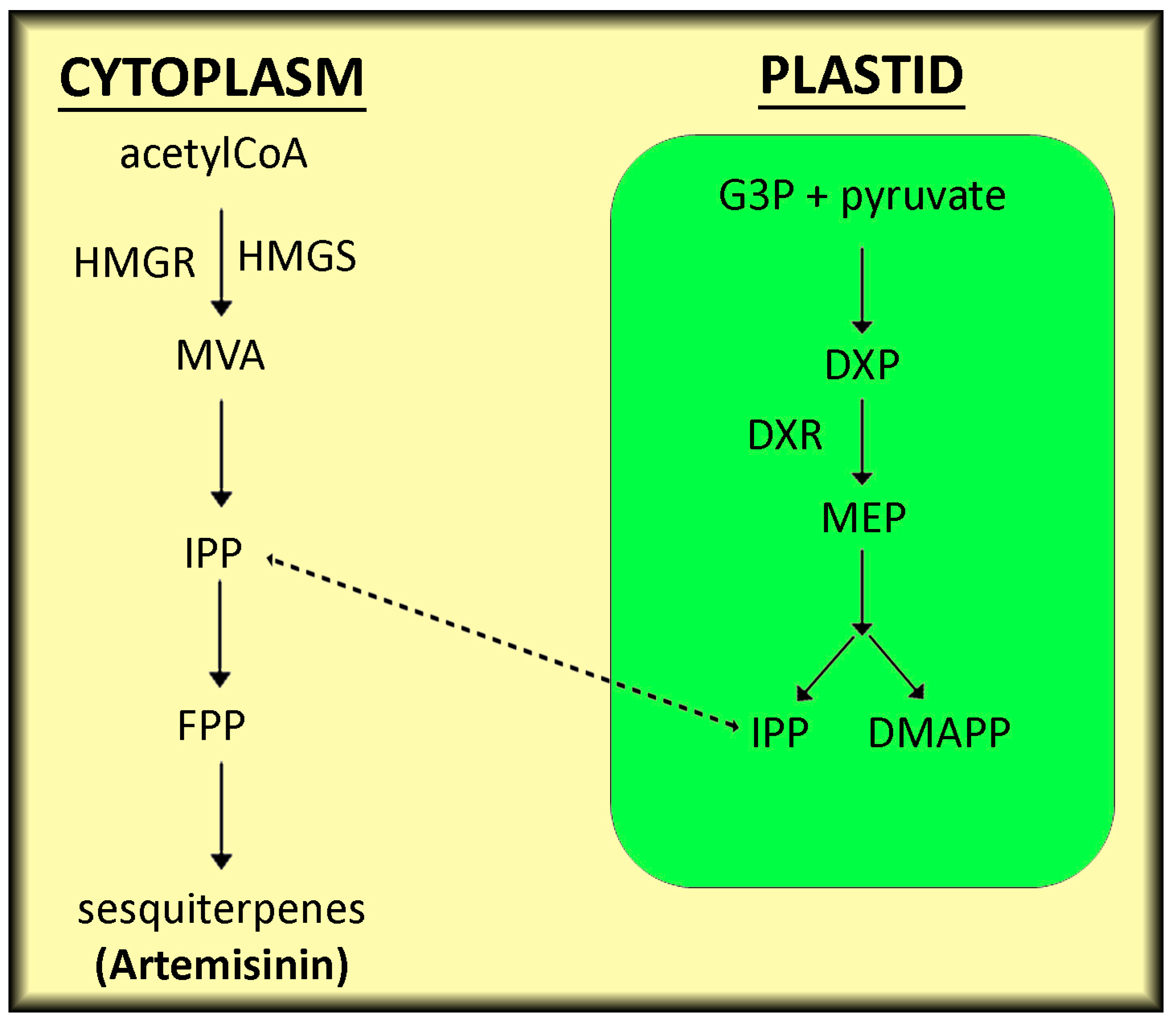
2. Production of the Sesquiterpene Artemisinin in Artemisia and Heterologous Hosts
3. Valorizing Stinging Nettle for the Production of Phenolic Compounds
4. Production of Caffeine in Coffee Cell Cultures and Heterologous Hosts
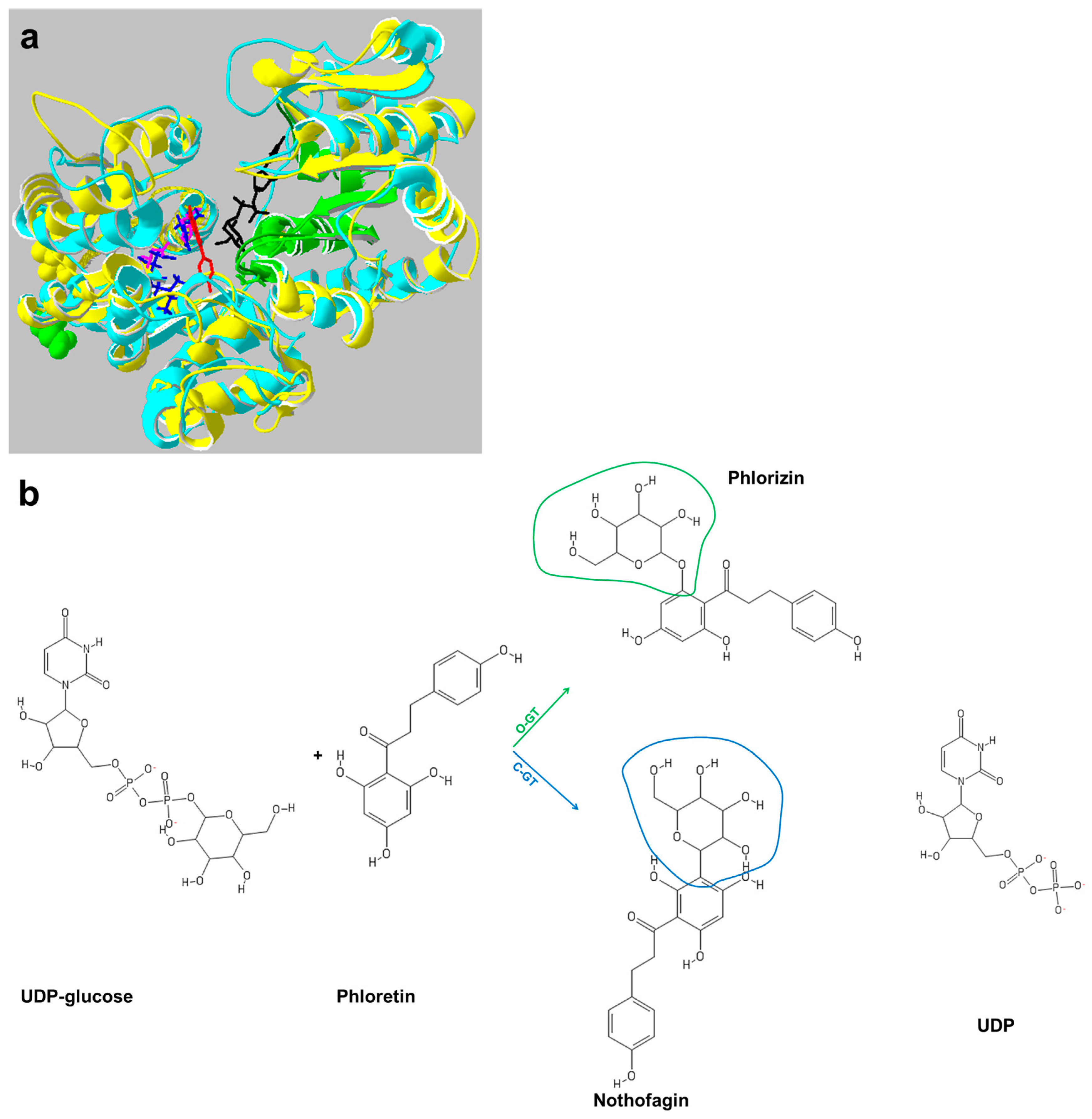
5. Plant Glycosyltransferases: Versatile Players in Biotechnology
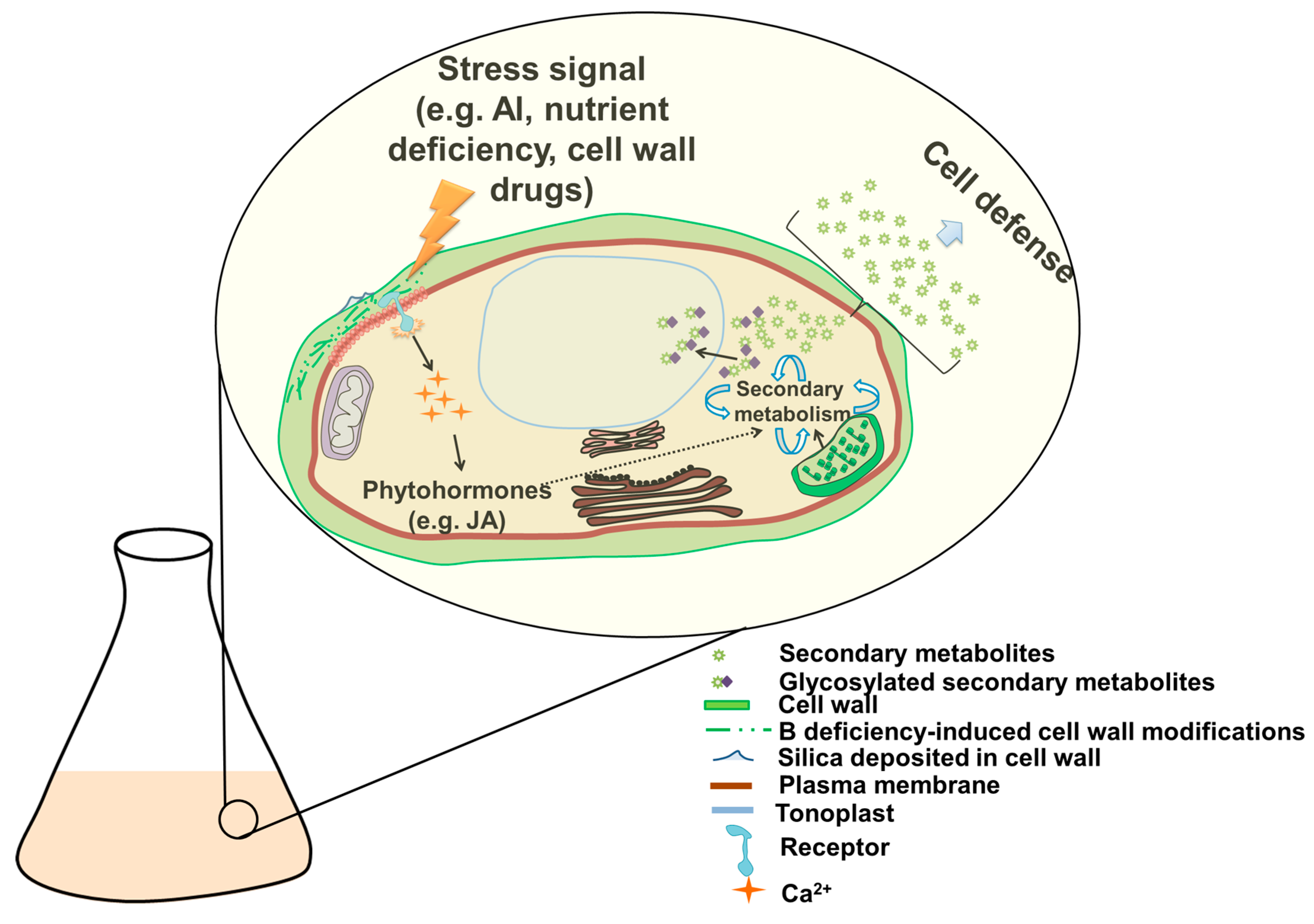
6. How to Boost Secondary Metabolite Production in Plant Cell Cultures?
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guerriero, G.; Hausman, J.-F.; Strauss, J.; Ertan, H.; Siddiqui, K.S. Lignocellulosic biomass: Biosynthesis, degradation, and industrial utilization. Eng. Life Sci. 2016, 16, 1–16. [Google Scholar] [CrossRef]
- Ncube, B.; Van Staden, J. Tilting plant metabolism for improved metabolite biosynthesis and enhanced human benefit. Molecules 2015, 20, 12698–12731. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, D.; Sanchez, R.; Lalaleo, L.; Bonfill, M.; Corchete, P.; Palazon, J. Biotechnological production of pharmaceuticals and biopharmaceuticals in plant cell and organ cultures. Curr. Med. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Shasmita, N.; Rai, M.K.; Naik, S.K. Exploring plant tissue culture in Withania somnifera (L.) Dunal: In vitro propagation and secondary metabolite production. Crit. Rev. Biotechnol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis sativa: The plant of the thousand and one molecules. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Mousa, A.A.; Syamasundar, K.V.; Alkhathlan, H.Z. Determination of chemical constituents of leaf and stem essential oils of Artemisia monosperma from central Saudi Arabia. Nat. Prod. Commun. 2012, 7, 1079–1082. [Google Scholar] [PubMed]
- Tripathi, A.K.; Prajapati, V.; Aggarwal, K.K.; Kumar, S. Toxicity, feeding deterrence, and effect of activity of 1,8-cineole from Artemisia annua on progeny production of Tribolium castanaeum (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2001, 94, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Ghahraman, A.; Nourbakhsh, N.; Mehdi, G.K.; Atar, F. Pollen morphology of Artemisia L. (Asteraceae) in Iran. Acta Bot. Bras. 2007. [Google Scholar] [CrossRef]
- Vallès, J.; Torrell, M.; Garnatje, T.; Garcia-Jacas, N.; Vilatersana, R.; Susanna, A. The genus Artemisia and its allies: Phylogeny of the subtribe artemisiinae (Asteraceae, Anthemideae) based on nucleotide sequences of nuclear ribosomal DNA Internal Transcribed Spacers (ITS). Plant Biol. 2003, 5, 274–284. [Google Scholar] [CrossRef]
- Pellicer Moscardó, J.; Garnatje i Roca, T.; Vallès Xirau, J. Artemisia (Asteraceae): Understanding its evolution using cytogenetic and molecular systematic tools, with emphasis on subgenus Dracunculus. Recent Adv. Pharm. Sci. 2011, 37, 199–222. [Google Scholar]
- Wang, W.-M. On the origin and development of Artemisia (Asteraceae) in the geological past. Bot. J. Linn. Soc. 2004, 145, 331–336. [Google Scholar] [CrossRef]
- Hunter, R.F.; Grant, S.A. The effect of altitude on grass growth in east Scotland. J. Appl. Ecol. 1971, 8, 1–19. [Google Scholar] [CrossRef]
- Pareto, G. Artemisie: Ricerca ed applicazione. In Quaderno Agricolo Supplemento; Federagrario: Torino, Italy, 1985; pp. 1–30. [Google Scholar]
- Tan, R.X.; Zheng, W.F.; Tang, H.Q. Biologically active substances from the genus Artemisia. Planta Med. 1998, 64, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.A.; Rahbari, S.; Rassouli, A.; Moslemi, M.H.; Khosravirad, F. Determination of artemisinin in Artemisia sieberi and anticoccidial effects of the plant extract in broiler chickens. Trop. Anim. Health Prod. 2006, 38, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Rustaiyan, A.; Nahrevanian, H.; Kazemi, M. A new antimalarial agent; effect of extracts of Artemisia diffusa against Plasmodium berghei. Pharmacogn. Mag. 2009, 5, 1–7. [Google Scholar] [CrossRef]
- Sun, Y.; Li, Y.-H.; Wu, X.-X.; Zheng, W.; Guo, Z.-H.; Li, Y.; Chen, T.; Hua, Z.-C.; Xu, Q. Ethanol extract from Artemisia vestita, a traditional Tibetan medicine, exerts anti-sepsis action through down-regulating the MAPK and NF-kappaB pathways. Int. J. Mol. Med. 2006, 17, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Juteau, F.; Masotti, V.; Bessière, J.M.; Dherbomez, M.; Viano, J. Antibacterial and antioxidant activities of Artemisia annua essential oil. Fitoterapia 2002, 73, 532–535. [Google Scholar] [CrossRef]
- Sayyah, M.; Nadjafnia, L.; Kamalinejad, M. Anticonvulsant activity and chemical composition of Artemisia dracunculus L. essential oil. J. Ethnopharmacol. 2004, 94, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Mirzaee, H.; Sharafi, A.; Hashemi Sohi, H. In vitro regeneration and transient expression of recombinant sesquiterpene cyclase (SQC) in Artemisia annua L. S. Afr. J. Bot. 2016, 104, 225–231. [Google Scholar] [CrossRef]
- Hailu, T.; Abera, B.; Mariam, G. In vitro mass propagation of Artemisia (Artemisia annua L.) cv: Anamed. Plant Tissue Cult. Biotechnol. 2014, 23, 165–176. [Google Scholar] [CrossRef]
- Janarthanam, B.; Rashmi, P.; Sumathi, E. Rapid and efficient plant regeneration from nodal explants of Artemisia annua L. Plant Tissue Cult. Biotechnol. 2012, 22, 33–39. [Google Scholar] [CrossRef]
- Dangash, A.; Ram, M.; Niranjan, R.; Bharillya, A.; Misra, H.; Pandya, N.; Jain, C.D. In vitro selection and hormonal regulation in cell culture of Artemisia annua L. Plant JSM Cell Dev. Biol. 2015, 3, 1013–1019. [Google Scholar]
- Sharafi, A.; Hashemi Sohi, H.; Sharafi, A.A.; Azadi, P.; Mousavi, A. Tissue culture and regeneration of an antimalarial plant, Artemisia sieberi Besser. Res. J. Pharmacogn. 2014, 1, 15–20. [Google Scholar]
- Sujatha, G.; Kumari, B.D.R. Micropropagation, encapsulation and growth of Artemisia vulgaris node explants for germplasm preservation. S. Afr. J. Bot. 2008, 74, 93–100. [Google Scholar] [CrossRef]
- Sudarshana, M.S.; Rajashekar, N.; Niranjan, M.H.; Borzabad, R.K. In vitro regeneration of multiple shoots from encapsulated somatic embryos of Artemisia vulgaris, L. IOSR J. Pharm. Biol. Sci. 2013, 6, 11–15. [Google Scholar] [CrossRef]
- Shinde, S.; Katewal, P.R.; Shanbhag, D.D.; Joseph, K.S.; Murthy, H.N. In vitro propagation of Artemisia japonica. J. Herbs Spices Med. Plants 2017, 23, 36–43. [Google Scholar] [CrossRef]
- Shinde, S.; Sebastian, J.K.; Jain, J.R.; Hanamanthagouda, M.S.; Murthy, H.N. Efficient in vitro propagation of Artemisia nilagirica var. nilagirica (Indian wormwood) and assessment of genetic fidelity of micropropagated plants. Physiol. Mol. Biol. Plants 2016, 22, 595–603. [Google Scholar] [PubMed]
- Mannan, A.; Syed, T.N.; Yameen, M.A.; Ullah, N.; Ismail, T.; Hussain, I.; Mirza, B. Effect of growth regulators on in vitro germination of Artemisia absinthium. Sci. Res. Essays 2012, 7, 1501–1507. [Google Scholar] [CrossRef]
- Kour, B.; Kour, G.; Kaul, S.; Dhar, M.K. In vitro mass multiplication and assessment of genetic stability of in vitro raised Artemisia absinthium L. plants using ISSR and SSAP molecular markers. Adv. Bot. 2014. [Google Scholar] [CrossRef]
- Bolyard, M. In vitro regeneration of Artemisia abrotanum L. by means of somatic organogenesis. Vitr. Cell. Dev. Biol. Plant 2018, 54, 127–130. [Google Scholar] [CrossRef]
- Mubashir, K.; Ganai, B.A.; Kamili, A.N.; Mustafa, K.G. In vitro plant regeneration of critically endangered medicinal plant Artemisia amygdalina D. Int. Res. J. Pharm. 2014, 5, 115–118. [Google Scholar]
- Dilshad, E.; Ismail, H.; Kayani, W.K.; Mirza, B. Optimization of conditions for genetic transformation and in vitro propagation of Artemisia carvifolia Buch. Curr. Synth. Syst. Biol. 2016, 4, 1–5. [Google Scholar]
- Sharafi, A.; Sohi, H.H.; Mirzaee, H.; Azadi, P. In vitro regeneration and Agrobacterium mediated genetic transformation of Artemisia aucheri Boiss. Physiol. Mol. Biol. Plants 2014, 20, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Aslam, N.; Zia, M.; Chaudhary, M.F. Callogenesis and direct organogenesis of Artemisia scoparia. Pak. J. Biol. Sci. 2006, 9, 1783–1786. [Google Scholar]
- Liu, C.Z.; Murch, S.J.; EL-Demerdash, M.; Saxena, P.K. Artemisia judaica L.: Micropropagation and antioxidant activity. J. Biotechnol. 2004, 110, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Shekhawat, M.S.; Manokari, M. Efficient in vitro propagation by ex vitro rooting methods of Artemisia absinthium L., an ethnobotanically important plant. Chin. J. Biol. 2015, 2015. [Google Scholar] [CrossRef]
- Hristova, L.; Damyanova, E.; Doichinova, Z.; Kapchina-Toteva, V. Effect of 6-benzylaminopurine on micropropagation of Artemisia chamaemelifolia Vill. (Asteraceae). Bulg. J. Agric. Sci. 2013, 19, 57–60. [Google Scholar]
- Nathar, V.N.; Yatoo, G.M. Micropropagation of an antidiabetic medicinal plant, Artemisia pallens. Turk. J. Bot. 2014, 38, 491–498. [Google Scholar] [CrossRef]
- Alok, A.; Shukla, V.; Pala, Z.; Kumar, J.; Kudale, S.; Desai, N. In vitro regeneration and optimization of factors affecting Agrobacterium mediated transformation in Artemisia pallens, an important medicinal plant. Physiol. Mol. Biol. Plants 2016, 22, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Kjær, A.; Grevsen, K.; Jensen, M. Effect of external stress on density and size of glandular trichomes in full-grown Artemisia annua, the source of anti-malarial artemisinin. AoB Plants 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Ikram, N.K.B.K.; Simonsen, H.T. A review of biotechnological artemisinin production in plants. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Baldi, A.; Dixit, V.K. Yield enhancement strategies for artemisinin production by suspension cultures of Artemisia annua. Explor. Horiz. Biotechnol. Glob. Ventur. 2008, 99, 4609–4614. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhao, C.; Huang, Y.; Yang, R.-Y.; Zeng, Q.-P. Abiotic stress-induced expression of artemisinin biosynthesis genes in Artemisia annua L. Chin. J. Appl. Environ. Biol. 2008, 14, 1–5. [Google Scholar]
- Pandey, N.; Pandey-Rai, S. Short term UV-B radiation-mediated transcriptional responses and altered secondary metabolism of in vitro propagated plantlets of Artemisia annua L. Plant Cell Tissue Organ Cult. 2014, 116, 371–385. [Google Scholar] [CrossRef]
- Xiang, L.; Zhu, S.; Zhao, T.; Zhang, M.; Liu, W.; Chen, M.; Lan, X.; Liao, Z. Enhancement of artemisinin content and relative expression of genes of artemisinin biosynthesis in Artemisia annua by exogenous MeJA treatment. Plant Growth Regul. 2015, 75, 435–441. [Google Scholar] [CrossRef]
- Patra, N.; Srivastava, A.K. Mass production of artemisinin using hairy root cultivation of Artemisia annua in bioreactor. In Bioprocessing of Plant In vitro Systems; Pavlov, A., Bley, T., Eds.; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Ting, H.M.; Wang, B.; Rydén, A.M.; Woittiez, L.; van Herpen, T.; Verstappen, F.W.; Ruyter-Spira, C.; Beekwilder, J.; Bouwmeester, H.J.; van der Krol, A. The metabolite chemotype of Nicotiana benthamiana transiently expressing artemisinin biosynthetic pathway genes is a function of CYP71AV1 type and relative gene dosage. New Phytol. 2013, 199, 352–366. [Google Scholar] [CrossRef] [PubMed]
- Farhi, M.; Marhevka, E.; Ben-Ari, J.; Algamas-Dimantov, A.; Liang, Z.; Zeevi, V.; Edelbaum, O.; Spitzer-Rimon, B.; Abeliovich, H.; Schwartz, B.; et al. Generation of the potent anti-malarial drug artemisinin in tobacco. Nat. Biotechnol. 2011, 29, 1072–1074. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Nowak, G.; Reed, D.W.; Covello, P.S. The production of artemisinin precursors in tobacco. Plant Biotechnol. J. 2011, 9, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, K.; Subramaniyan, M.; Rawat, K.; Kalamuddin, M.; Qureshi, M.I.; Malhotra, P.; Mohmmed, A.; Cornish, K.; Daniell, H.; Kumar, S. Compartmentalized metabolic engineering for artemisinin biosynthesis and effective malaria treatment by oral delivery of plant cells. Mol. Plant 2016, 9, 1464–1477. [Google Scholar] [CrossRef] [PubMed]
- Reski, R.; Parsons, J.; Decker, E.L. Moss-made pharmaceuticals: From bench to bedside. Plant Biotechnol. J. 2015, 13, 1191–1198. [Google Scholar] [CrossRef] [PubMed]
- Salehi, M.; Karimzadeh, G.; Naghavi, M.R.; Naghdi Badi, H.; Rashidi Monfared, S. Expression of artemisinin biosynthesis and trichome formation genes in five Artemisia species. Ind. Crop. Prod. 2018, 112, 130–140. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Janick, J. Floral morphology of Artemisia annua with special reference to trichomes. Int. J. Plant Sci. 1995, 156, 807–815. [Google Scholar]
- Liu, S.; Tian, N.; Li, J.; Huang, J.; Liu, Z. Isolation and identification of novel genes involved in artemisinin production from flowers of Artemisia annua using suppression subtractive hybridization and metabolite analysis. Planta Med. 2009, 75, 1542–1547. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Xiao, L.; Gao, S.; Li, Q.; Chen, J.; Xiao, Y.; Ji, Q.; Chen, R.; Chen, W.; Zhang, L. Trichome and artemisinin regulator 1 is required for trichome development and artemisinin biosynthesis in Artemisia annua. Mol. Plant 2015, 8, 1396–1411. [Google Scholar] [CrossRef] [PubMed]
- Matías-Hernández, L.; Jiang, W.; Yang, K.; Tang, K.; Brodelius, P.E.; Pelaz, S. AaMYB1 and its orthologue AtMYB61 affect terpene metabolism and trichome development in Artemisia annua and Arabidopsis thaliana. Plant J. 2017, 90, 520–534. [Google Scholar] [CrossRef] [PubMed]
- Towler, M.J.; Weathers, P.J. Evidence of artemisinin production from IPP stemming from both the mevalonate and the nonmevalonate pathways. Plant Cell Rep. 2007, 26, 2129–2136. [Google Scholar] [CrossRef] [PubMed]
- Graham, I.A.; Besser, K.; Blumer, S.; Branigan, C.A.; Czechowski, T.; Elias, L.; Guterman, I.; Harvey, D.; Isaac, P.G.; Khan, A.M.; et al. The genetic map of Artemisia annua L. identifies loci affecting yield of the antimalarial drug artemisinin. Science 2010, 327, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Zeng, L.; Yuan, Y.; Chen, M.; Wang, F.; Liu, X.; Zeng, L.; Lan, X.; Liao, Z. Enhancement of artemisinin biosynthesis by overexpressing dxr, cyp71av1 and cpr in the plants of Artemisia annua L. Plant Omics 2012, 5, 503–507. [Google Scholar]
- Wen, W.; Yu, R. Artemisinin biosynthesis and its regulatory enzymes: Progress and perspective. Pharmacogn. Rev. 2011, 5, 189–194. [Google Scholar] [PubMed]
- Ram, M.; Khan, M.A.; Jha, P.; Khan, S.; Kiran, U.; Ahmad, M.M.; Javed, S.; Abdin, M.Z. HMG-CoA reductase limits artemisinin biosynthesis and accumulation in Artemisia annua L. plants. Acta Physiol. Plant. 2010, 32, 859–866. [Google Scholar] [CrossRef]
- Covello, P.S.; Teoh, K.H.; Polichuk, D.R.; Reed, D.W.; Nowak, G. Functional genomics and the biosynthesis of artemisinin. Phytochem. Soc. N. Am. 2007, 68, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Teoh Keat, H.; Polichuk Devin, R.; Reed Darwin, W.; Nowak, G.; Covello Patrick, S. Artemisia annua L. (Asteraceae) trichome-specific cDNAs reveal CYP71AV1, a cytochrome P450 with a key role in the biosynthesis of the antimalarial sesquiterpene lactone artemisinin. FEBS Lett. 2006, 580, 1411–1416. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.-P.; Zeng, X.-M.; Yin, L.-L.; Yang, R.-Y.; Feng, L.-L.; Yang, X.-Q. Quantification of three key enzymes involved in artemisinin biogenesis in Artemisia annua by polyclonal antisera-based ELISA. Plant Mol. Biol. Rep. 2009, 27, 50–57. [Google Scholar] [CrossRef]
- Ro, D.-K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Ćavar, S.; Maksimović, M.; Vidic, D.; Parić, A. Chemical composition and antioxidant and antimicrobial activity of essential oil of Artemisia annua L. from Bosnia. Ind. Crop. Prod. 2012, 37, 479–485. [Google Scholar] [CrossRef]
- Radulović, N.S.; Randjelović, P.J.; Stojanović, N.M.; Blagojević, P.D.; Stojanović-Radić, Z.Z.; Ilić, I.R.; Djordjević, V.B. Toxic essential oils. Part II: Chemical, toxicological, pharmacological and microbiological profiles of Artemisia annua L. volatiles. Food Chem. Toxicol. 2013, 58, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, M. Essential oil of the aerial parts of Artemisia annua (Asteraceae) from Iran. J. Essent. Oil Bear. Plants 2015, 18, 1003–1005. [Google Scholar] [CrossRef]
- Marinas Ioana, C.; Oprea, E.; Chifiriuc Mariana, C.; Badea Irinel, A.; Buleandra, M. Lazar veronica chemical composition and antipathogenic activity of Artemisia annua essential oil from Romania. Chem. Biodivers. 2015, 12, 1554–1564. [Google Scholar] [CrossRef] [PubMed]
- Asgharpour, N.; Honarvar, M. Identification and comparison of essential oil composition of Artemisia sieberi and Artemisia aucheri cultivated in the south of Iran. J. Essent. Oil Bear. Plants 2016, 19, 756–761. [Google Scholar] [CrossRef]
- Mohammadhosseini, M.; Akbarzadeh, A.; Hashemi-Moghaddam, H.; Nafchi, A.M.; Mashayekhi, H.A.; Aryanpour, A. Chemical composition of the essential oils from the aerial parts of Artemisia sieberi by using conventional hydrodistillation and microwave assisted hydrodistillation: A comparative study. J. Essent. Oil Bear. Plants 2016, 19, 32–45. [Google Scholar] [CrossRef]
- Guetat, A.; Al-Ghamdi, F.A.; Osman, A.K. The genus Artemisia L. in the northern region of Saudi Arabia: Essential oil variability and antibacterial activities. Nat. Prod. Res. 2017, 31, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Belhattab, R.; Amor, L.; Barroso, J.G.; Pedro, L.G.; Cristina Figueiredo, A. Essential oil from Artemisia herba-alba Asso grown wild in Algeria: Variability assessment and comparison with an updated literature survey. Arab. J. Chem. 2014, 7, 243–251. [Google Scholar] [CrossRef]
- Al-Shuneigat, J.; Al-Sarayreh, S.; Al-Qudah, M.; Al-Tarawneh, I.; Al-Saraireh, Y.; Al-Qtaitat, A. GC-MS analysis and antibacterial activity of the essential oil isolated from wild Artemisia herba-alba grown in South Jordan. Br. J. Med. Med. Res. 2015, 5. [Google Scholar] [CrossRef]
- Dahmani-Hamzaoui, N.; Baaliouamer, A. Volatile constituents of Algerian Artemisia herba-alba essential oils. J. Essent. Oil Res. 2015, 27, 437–446. [Google Scholar] [CrossRef]
- Abu-Darwish, M.S.; Cabral, C.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Efferth, T.; Salgueiro, L. Artemisia herba-alba essential oil from Buseirah (South Jordan): Chemical characterization and assessment of safe antifungal and anti-inflammatory doses. J. Ethnopharmacol. 2015, 174, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Younsi, F.; Trimech, R.; Boulila, A.; Ezzine, O.; Dhahri, S.; Boussaid, M.; Messaoud, C. Essential oil and phenolic compounds of Artemisia herba-alba (Asso.): Composition, antioxidant, antiacetylcholinesterase, and antibacterial activities. Int. J. Food Prop. 2016, 19, 1425–1438. [Google Scholar] [CrossRef]
- Bellili, S.; Jazi, S.; Hrira, M.Y.; Lamari, A.; Dhifi, W.; Diouani, M.F.; Araújo, M.E.; Cioni, P.L.; Flamini, G.; Cherif, A. Phytochemical identification of volatile fraction, essential oil and screening of antioxidant, antibacterial, allelopathic and insecticidal potential from Artemisia herba-alba leaves. Main Group Chem. 2017, 16, 95–109. [Google Scholar] [CrossRef]
- Han, J.-L.; Liu, B.-Y.; Ye, H.-C.; Wang, H.; Li, Z.-Q.; Li, G.-F. Effects of overexpression of the endogenous farnesyl diphosphate synthase on the artemisinin content in Artemisia annua L. J. Integr. Plant Biol. 2006, 48, 482–487. [Google Scholar] [CrossRef]
- Aquil, S.; Husaini, A.M.; Abdin, M.Z.; Rather, G.M. Overexpression of the HMG-CoA reductase gene leads to enhanced artemisinin biosynthesis in transgenic Artemisia annua plants. Planta Med. 2009, 75, 1453–1458. [Google Scholar] [CrossRef] [PubMed]
- Banyai, W.; Kirdmanee, C.; Mii, M.; Supaibulwatana, K. Overexpression of farnesyl pyrophosphate synthase (FPS) gene affected artemisinin content and growth of Artemisia annua L. Plant Cell Tissue Organ Cult. 2010, 103, 255–265. [Google Scholar] [CrossRef]
- Nafis, T.; Akmal, M.; Ram, M.; Alam, P.; Ahlawat, S.; Mohd, A.; Abdin, M.Z. Enhancement of artemisinin content by constitutive expression of the HMG-CoA reductase gene in high-yielding strain of Artemisia annua L. Plant Biotechnol. Rep. 2011, 5, 53–60. [Google Scholar] [CrossRef]
- Ma, D.; Li, G.; Alejos-Gonzalez, F.; Zhu, Y.; Xue, Z.; Wang, A.; Zhang, H.; Li, X.; Ye, H.; Wang, H.; et al. Overexpression of a type-I isopentenyl pyrophosphate isomerase of Artemisia annua in the cytosol leads to high arteannuin B production and artemisinin increase. Plant J. 2017, 91, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Fu, X.; Liu, M.; Shen, Q.; Jiang, W.; Li, L.; Sun, X.; Tang, K. Promotion of artemisinin content in Artemisia annua by overexpression of multiple artemisinin biosynthetic pathway genes. Plant Cell Tissue Organ Cult. 2017, 129, 251–259. [Google Scholar] [CrossRef]
- Chen, J.-L.; Fang, H.-M.; Ji, Y.-P.; Pu, G.-B.; Guo, Y.-W.; Huang, L.-L.; Du, Z.-G.; Liu, B.-Y.; Ye, H.-C.; Li, G.-F.; et al. Artemisinin biosynthesis enhancement in transgenic Artemisia annua plants by downregulation of the beta-caryophyllene synthase gene. Planta Med. 2011, 77, 1759–1765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jing, F.; Li, F.; Li, M.; Wang, Y.; Wang, G.; Sun, X.; Tang, K. Development of transgenic Artemisia annua (Chinese wormwood) plants with an enhanced content of artemisinin, an effective anti-malarial drug, by hairpin-RNA-mediated gene silencing. Biotechnol. Appl. Biochem. 2010, 52, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Amanullah, B.M.; Rizvi, Z.F.; Zia, M. Production of artemisinin and its derivatives in hairy roots of Artemisia dubia induced by rola gene transformation. Pak. J. Bot. 2016, 48, 699–706. [Google Scholar]
- Dilshad, E.; Cusido, R.M.; Estrada, K.R.; Bonfill, M.; Mirza, B. Genetic transformation of Artemisia carvifolia buch with rol genes enhances artemisinin accumulation. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Dilshad, E.; Cusido, R.M.; Palazon, J.; Estrada, K.R.; Bonfill, M.; Mirza, B. Enhanced artemisinin yield by expression of rol genes in Artemisia annua. Malar. J. 2015, 14. [Google Scholar] [CrossRef]
- Shen, Q.; Yan, T.; Fu, X.; Tang, K. Transcriptional regulation of artemisinin biosynthesis in Artemisia annua L. Sci. Bull. 2016, 61, 18–25. [Google Scholar]
- Guerriero, G.; Behr, M.; Backes, A.; Faleri, C.; Hausman, J.-F.; Lutts, S.; Cai, G. Bast fibre formation: Insights from Next-Generation Sequencing. Procedia Eng. 2017, 200, 229–235. [Google Scholar] [CrossRef]
- Bacci, L.; Baronti, S.; Predieri, S.; di Virgilio, N. Fiber yield and quality of fiber nettle (Urtica dioica L.) cultivated in Italy. Ind. Crop. Prod. 2009, 29, 480–484. [Google Scholar] [CrossRef]
- Farag, M.A.; Weigend, M.; Luebert, F.; Brokamp, G.; Wessjohann, L.A. Phytochemical, phylogenetic, and anti-inflammatory evaluation of 43 Urtica accessions (stinging nettle) based on UPLC-Q-TOF-MS metabolomic profiles. Phytochemistry 2013, 96, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Viktorova, J.; Jandova, Z.; Madlenakova, M.; Prouzova, P.; Bartunek, V.; Vrchotova, B.; Lovecka, P.; Musilova, L.; Macek, T. Native phytoremediation potential of Urtica dioica for removal of PCBs and heavy metals can be improved by genetic manipulations using constitutive CaMV 35S promoter. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Angeles, G.; Berrio-Sierra, J.; Joseleau, J.-P.; Lorimier, P.; Lefèbvre, A.; Ruel, K. Preparative laser capture microdissection and single-pot cell wall material preparation: A novel method for tissue-specific analysis. Planta 2006, 224, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Pinelli, P.; Ieri, F.; Vignolini, P.; Bacci, L.; Baronti, S.; Romani, A. Extraction and HPLC analysis of phenolic compounds in leaves, stalks, and textile fibers of Urtica dioica L. J. Agric. Food Chem. 2017, 56, 9127–9132. [Google Scholar] [CrossRef] [PubMed]
- Modarresi-Chahardehi, A.; Ibrahim, D.; Fariza-Sulaiman, S.; Mousavi, L. Screening antimicrobial activity of various extracts of Urtica dioica. Rev. Biol. Trop. 2012, 60, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Asgarpanah, J.; Mohajerani, R. Phytochemistry and pharmacologic properties of Urtica dioica L. J. Med. Plants Res. 2012, 6, 5714–5719. [Google Scholar]
- Fraser, C.M.; Chapple, C. The phenylpropanoid pathway in Arabidopsis. Arab. Book Am. Soc. Plant Biol. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Schöttner, M.; Gansser, D.; Spiteller, G. Lignans from the roots of Urtica dioica and their metabolites bind to human sex hormone binding globulin (SHBG). Planta Med. 1997, 63, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Murata, J.; Matsumoto, E.; Morimoto, K.; Koyama, T.; Satake, H. Generation of triple-transgenic Forsythia cell cultures as a platform for the efficient, stable, and sustainable production of lignans. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Kokotkiewicz, A.; Marzec-Wróblewska, U.; Bucinski, A.; Luczkiewicz, M.; Ekiert, H. Accumulation of dibenzocyclooctadiene lignans in agar cultures and in stationary and agitated liquid cultures of Schisandra chinensis (Turcz.) Baill. Appl. Microbiol. Biotechnol. 2016, 100, 3965–3977. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian Chashmi, N.; Sharifi, M.; Behmanesh, M. Lignan enhancement in hairy root cultures of Linum album using coniferaldehyde and methylenedioxycinnamic acid. Prep. Biochem. Biotechnol. 2016, 46, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Li, Q.; Tan, H.; Chen, J.; Xiao, Y.; Ma, R.; Gao, S.; Zerbe, P.; Chen, W.; Zhang, L. Gene-to-metabolite network for biosynthesis of lignans in MeJA-elicited Isatis indigotica hairy root cultures. Front. Plant Sci. 2015, 6, 952. [Google Scholar] [CrossRef] [PubMed]
- Gabr, A.M.M.; Mabrok, H.B.; Abdel-Rahim, E.A.; El-Bahr, M.K.; Smetanska, I. Determination of lignans, phenolic acids and antioxidant capacity in transformed hairy root culture of Linum usitatissimum. Nat. Prod. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Van Summeren-Wesenhagen, P.V.; Marienhagen, J. Putting bugs to the blush. Bioengineered 2013, 4, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Satake, H.; Koyama, T.; Bahabadi, S.E.; Matsumoto, E.; Ono, E.; Murata, J. Essences in Metabolic Engineering of Lignan Biosynthesis. Metabolites 2015, 5, 270–290. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef] [PubMed]
- Ruelland, E.; Kravets, V.; Derevyanchuk, M.; Martinec, J.; Zachowski, A.; Pokotylo, I. Role of phospholipid signalling in plant environmental responses. Environ. Exp. Bot. 2015, 114, 129–143. [Google Scholar] [CrossRef]
- Gopalakrishnakone, P.; Carlini, C.R.; Ligabue-Braun, R. Toxinology; Springer: Dordrecht, The Netherlands, 2017. [Google Scholar]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [PubMed]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. Acad. J. 2009, 3, 1222–1239. [Google Scholar]
- Toledo Paulo, R.A.B.; Pezza, L.; Pezza Helena, R.; Toci Aline, T. Relationship between the different aspects related to coffee quality and their volatile compounds. Compr. Rev. Food Sci. Food Saf. 2016, 15, 705–719. [Google Scholar] [CrossRef]
- Tolessa, K.; Rademaker, M.; De Baets, B.; Boeckx, P. Prediction of specialty coffee cup quality based on near infrared spectra of green coffee beans. Talanta 2016, 150, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Estévez, M.; Muñoz-Sánchez, J.; Loyola-Vargas, V.; Hernández-Sotomayor, S. Modification of the culture medium to produce aluminum toxicity in cell suspensions of coffee (Coffea arabica L.). Plant Cell Rep. 2001, 20, 469–474. [Google Scholar] [CrossRef]
- Arroyo-Serralta, G.A.; Kú-González, A.; Hernández-Sotomayor, S.M.; Zúñiga, J.A. Exposure to toxic concentrations of aluminum activates a MAPK-like protein in cell suspension cultures of Coffea arabica. Plant Physiol. Biochem. 2005, 43, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Poot-Poot, W.; Rodas-Junco, B.A.; Muñoz-Sánchez, J.A.; Hernández-Sotomayor, S.M.T. Protoplasts: A friendly tool to study aluminum toxicity and coffee cell viability. SpringerPlus 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Pech-Kú, R.; Muñoz-Sánchez, J.A.; Monforte-González, M.; Vázquez-Flota, F.; Rodas-Junco, B.A.; González-Mendoza, V.M.; Hernández-Sotomayor, S.M.T. Relationship between aluminum stress and caffeine biosynthesis in suspension cells of Coffea arabica L. J. Inorg. Biochem. 2018, 181, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Ostergaard, S.; Olsson, L.; Nielsen, J. Metabolic Engineering of Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2000, 64, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Algharrawi, K.H.R.; Summers, R.M.; Gopishetty, S.; Subramanian, M. Direct conversion of theophylline to 3-methylxanthine by metabolically engineered E. coli. Microb. Cell Factories 2015, 14. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Bhuiya, M.W.; Li, M.; Liu, X.; Han, J.; Deng, W.; Wang, M.; Yu, O.; Zhang, Z. Metabolic engineering of Saccharomyces cerevisiae for caffeine and theobromine production. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- McKeague, M.; Wang, Y.-H.; Cravens, A.; Win, M.N.; Smolke, C.D. Engineering a microbial platform for de novo biosynthesis of diverse methylxanthines. Metab. Eng. 2016, 38, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Gokulakrishnan, S.; Chandraraj, K.; Gummadi, S.N. Microbial and enzymatic methods for the removal of caffeine. Enzym. Microb. Technol. 2005, 37, 225–232. [Google Scholar] [CrossRef]
- Gachon, C.M.M.; Langlois-Meurinne, M.; Saindrenan, P. Plant secondary metabolism glycosyltransferases: The emerging functional analysis. Trends Plant Sci. 2005, 10, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Le Roy, J.; Huss, B.; Creach, A.; Hawkins, S.; Neutelings, G. Glycosylation is a major regulator of phenylpropanoid availability and biological activity in plants. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wei, G.; Zhou, H.; Gu, C.; Vimolmangkang, S.; Liao, L.; Han, Y. Unraveling the mechanism underlying the glycosylation and methylation of anthocyanins in peach. Plant Physiol. 2014, 166, 1044–1058. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucl. Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Bowles, D.; Lim, E.-K.; Poppenberger, B.; Vaistij, F.E. Glycosyltransferases of lipophilic small molecules. Annu. Rev. Plant Biol. 2006, 57, 567–597. [Google Scholar] [CrossRef] [PubMed]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyltransferases: Structures, functions, and mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Hughes, M.A. Multiple secondary plant product UDP-glucose glucosyltransferase genes expressed in cassava (Manihot esculenta Crantz) cotyledons. DNA Seq. J. DNA Seq. Mapp. 1994, 5, 41–49. [Google Scholar] [CrossRef]
- Osmani, S.A.; Bak, S.; Møller, B.L. Substrate specificity of plant UDP-dependent glycosyltransferases predicted from crystal structures and homology modeling. Phytochemistry 2009, 70, 325–347. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T.; Jones, P. Glycosyltransferases in plant natural product synthesis: Characterization of a supergene family. Trends Plant Sci. 2000, 5, 380–386. [Google Scholar] [CrossRef]
- Masada, S.; Terasaka, K.; Mizukami, H. A single amino acid in the PSPG-box plays an important role in the catalytic function of CaUGT2 (Curcumin glucosyltransferase), a Group D Family 1 glucosyltransferase from Catharanthus roseus. FEBS Lett. 2007, 581, 2605–2610. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kim, B.-G.; Sung, S.; Kim, M.; Mok, H.; Chong, Y.; Ahn, J.-H. Engineering flavonoid glycosyltransferases for enhanced catalytic efficiency and extended sugar-donor selectivity. Planta 2013, 238, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Hansen, E.H.; Osmani, S.A.; Kristensen, C.; Møller, B.L.; Hansen, J. Substrate specificities of family 1 UGTs gained by domain swapping. Phytochemistry 2009, 70, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Osmani, S.A.; Bak, S.; Imberty, A.; Olsen, C.E.; Møller, B.L. Catalytic key amino acids and UDP-sugar donor specificity of a plant glucuronosyltransferase, UGT94B1: Molecular modeling substantiated by site-specific mutagenesis and biochemical analyses. Plant Physiol. 2008, 148, 1295–1308. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-Y.; Li, X. Identification of a residue responsible for UDP-sugar donor selectivity of a dihydroxybenzoic acid glycosyltransferase from Arabidopsis natural accessions. Plant J. Cell Mol. Biol. 2017, 89, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S.; Shemsi, A.M.; Guerriero, G.; Najnin, T.; Ertan, H.; Taha. Biotechnological improvements of cold-adapted enzymes: Commercialization via an integrated approach. In Psychrophiles: From Biodiversity to Biotechnology; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Karim, M.R.; Hashinaga, F. Screenig and some properties of limonoid glucosyltransferase. Pak. J. Biol. Sci. 2001, 4, 483–486. [Google Scholar]
- Vaghari, H.; Jafarizadeh-Malmiri, H.; Mohammadlou, M.; Berenjian, A.; Anarjan, N.; Jafari, N.; Nasiri, S. Application of magnetic nanoparticles in smart enzyme immobilization. Biotechnol. Lett. 2016, 38, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Brazier-Hicks, M.; Evans, K.M.; Gershater, M.C.; Puschmann, H.; Steel, P.G.; Edwards, R. The c-glycosylation of flavonoids in cereals. J. Biol. Chem. 2009, 284, 17926–17934. [Google Scholar] [CrossRef] [PubMed]
- Falcone Ferreyra, M.L.; Rodriguez, E.; Casas, M.I.; Labadie, G.; Grotewold, E.; Casati, P. Identification of a bifunctional maize C- and O-glucosyltransferase. J. Biol. Chem. 2013, 288, 31678–31688. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Capanoglu, E.; Jassbi, A.R.; Miron, A. Advance on the flavonoid c-glycosides and health benefits. Crit. Rev. Food Sci. Nutr. 2016, 56. [Google Scholar] [CrossRef] [PubMed]
- Gutmann, A.; Nidetzky, B. Enzymatic C-glycosylation: Insights from the study of a complementary pair of plant O- and C-glucosyltransferases. Pure Appl. Chem. 2013, 85, 1865–1877. [Google Scholar] [CrossRef]
- Soda, N.; Verma, L.; Giri, J. CRISPR-Cas9-based plant genome editing: Significance, opportunities and recent advances. Plant Physiol. Biochem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Křen, V.; Thiem, J. Glycosylation employing bio-systems: From enzymes to whole cells. Chem. Soc. Rev. 1997, 26, 463–473. [Google Scholar] [CrossRef]
- McArthur, J.B.; Chen, X. Glycosyltransferase engineering for carbohydrate synthesis. Biochem. Soc. Trans. 2016, 44, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Liu, L.; Shin, H.; Chen, R.R.; Li, J.; Du, G.; Chen, J. Iterative saturation mutagenesis of −6 subsite residues in cyclodextrin glycosyltransferase from Paenibacillus macerans to improve maltodextrin specificity for 2-O-d-glucopyranosyl-l-ascorbic acid synthesis. Appl. Environ. Microbiol. 2013, 79, 7562–7568. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Hamann, T. Plant cell wall integrity maintenance as an essential component of biotic stress response mechanisms. Front. Plant Sci. 2012, 3, 77. [Google Scholar] [CrossRef] [PubMed]
- Hamann, T. The plant cell wall integrity maintenance mechanism—A case study of a cell wall plasma membrane signaling network. Phytochemistry 2015, 112, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Gall, H.L.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.E. Cell wall integrity signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 262–291. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Silvestrini, L.; Legay, S.; Maixner, F.; Sulyok, M.; Hausman, J.F.; Strauss, J. Deletion of the celA gene in Aspergillus nidulans triggers overexpression of secondary metabolite biosynthetic genes. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Blanco, C.; Feng, D.X.; Hu, J.; Sánchez-Vallet, A.; Deslandes, L.; Llorente, F.; Berrocal-Lobo, M.; Keller, H.; Barlet, X.; Sánchez-Rodríguez, C.; et al. Impairment of cellulose synthases required for Arabidopsis secondary cell wall formation enhances disease resistance. Plant Cell 2007, 19, 890–903. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.; Karafyllidis, I.; Wasternack, C.; Turner, J.G. The Arabidopsis mutant cev1 links cell wall signaling to jasmonate and ethylene responses. Plant Cell 2002, 14, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Tateno, M.; Brabham, C.; DeBolt, S. Cellulose biosynthesis inhibitors—A multifunctional toolbox. J. Exp. Bot. 2016, 67, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Bacete, L.; Mélida, H.; Miedes, E.; Molina, A. Plant cell wall-mediated immunity: Cell wall changes trigger disease resistance responses. Plant J. 2018, 93, 614–636. [Google Scholar] [CrossRef] [PubMed]
- Mélida, H.; García-Angulo, P.; Alonso-Simón, A.; Alvarez, J.M.; Acebes, J.L.; Encina, A. The phenolic profile of maize primary cell wall changes in cellulose-deficient cell cultures. Phytochemistry 2010, 71, 1684–1689. [Google Scholar] [CrossRef] [PubMed]
- Mélida, H.; Encina, A.; Alvarez, J.; Acebes, J.L.; Caparrós-Ruiz, D. Unraveling the biochemical and molecular networks involved in maize cell habituation to the cellulose biosynthesis inhibitor dichlobenil. Mol. Plant 2010, 3, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Mélida, H.; Álvarez, J.; Acebes, J.L.; Encina, A.; Fry, S.C. Changes in cinnamic acid derivatives associated with the habituation of maize cells to dichlobenil. Mol. Plant 2011, 4, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Elfahmi, N.; Batterman, S.; Koulman, A.; Hackl, T.; Bos, R.; Kayser, O.; Woerdenbag, H.J.; Quax, W.J. Lignans from cell suspension cultures of Phyllanthus niruri, an Indonesian medicinal plant. J. Nat. Prod. 2006, 69, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Cristóbal, J.J.; Lunar, L.; Lafont, F.; Baumert, A.; González-Fontes, A. Boron deficiency causes accumulation of chlorogenic acid and caffeoyl polyamine conjugates in tobacco leaves. J. Plant Physiol. 2004, 161, 879–881. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulos, G.; Karabourniotis, G. Boron deficiency and concentrations and composition of phenolic compounds in Olea europaea leaves: A combined growth chamber and field study. Tree Physiol. 2005, 25, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Hausman, J.-F.; Legay, S. Silicon and the plant extracellular matrix. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, M.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Silicon and plants: Current knowledge and technological perspectives. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Van Bockhaven, J.; De Vleesschauwer, D.; Höfte, M. Towards establishing broad-spectrum disease resistance in plants: Silicon leads the way. J. Exp. Bot. 2013, 64, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Rostkowska, C.; Mota, C.M.; Oliveira, T.C.; Santiago, F.M.; Oliveira, L.A.; Korndörfer, G.H.; Lana, R.M.Q.; Rossi, M.L.; Nogueira, N.L.; Simonnet, X.; et al. Si-accumulation in Artemisia annua glandular trichomes increases artemisinin concentration, but does not interfere in the impairment of Toxoplasma gondii growth. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Sivanesan, I.; Park, S.W. The role of silicon in plant tissue culture. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, L.; Liu, J.; Liu, X.; Li, X.; Ma, J.; Lin, Y.; Xu, F. Evidence for “silicon” within the cell walls of suspension-cultured rice cells. New Phytol. 2013, 200, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Landi, S.; Hausman, J.-F.; Guerriero, G.; Esposito, S. Poaceae vs. abiotic stress: Focus on drought and salt stress, recent insights and perspectives. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Chemical Constituents (%) | References |
|---|---|---|
| A. annua | Camphor (44), germacrene D (16), trans-pinocarveol (11), β-selinene (9), β-caryophyllene (9), artemisia ketone (3) | [18] |
| Artemisia ketone (30.7), camphor (15.8) | [67] | |
| Artemisia ketone (35.7), α-pinene (16.5), 1,8-cineole (5.5) | [68] | |
| α-Pinene (7.33), camphene (5.68), sabinene (4.78), β-myrcene (22.41), 1,8-cineole (17.17), camphor (20.41) | [69] | |
| Camphor (17.74), α-pinene (9.66), germacrene D (7.55), 1,8-cineole (7.24), β-caryophyllene (7.02), artemisia ketone (6.26) | [70] | |
| A. sieberi | Camphor (29.5), cis-thujone (22.58), 1,8-cineole (12.91), trans-thujone (10.60), camphene (5.05) | [71] |
| Camphor (22.0), 1,8-cineole (19.3), cis-davanone (15.0), camphene (4.6), terpinene-4-ol (3.2) | [72] | |
| Spathulenol (30.42) | [73] | |
| A. monosperma | β -Pinene (50.3), α-terpinolene (10.0), limonone (5.4), α-pinene (4.6) | [6] |
| Butanoic acid (17.87) | [73] | |
| A. herba-alba | Camphor (39.1), chrysanthenone (15.0), cis-thujone (7.8) | [6] |
| Camphor (17–33), α-thujone (7–28), chrysanthenone (4–19) | [74] | |
| Cis-chrysanthenol (13.83), 1, 8-cineole (12.84), cis-limonene (12.57), α-terpinenol (6.97), γ-muurolene (4.50) | [75] | |
| α-Thujone (trace-47.1), camphor (5.6–30.0), chrysanthenone (trace-13.5), β -thujone (trace-9.2), 1,8-cineole (4.1–11.4) | [76] | |
| 1,8-cineole (20.1), α-thujone (25.1), β -thujone (22.9), camphor (10.5) | [77] | |
| β -thujone (41.9), α-thujone (18.4), camphor (13.2) | [78] | |
| α-thujone (37.9), germacrene D (16.5), 1,8-cineole (8.4), β-thujone (7.8) | [79] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerriero, G.; Berni, R.; Muñoz-Sanchez, J.A.; Apone, F.; Abdel-Salam, E.M.; Qahtan, A.A.; Alatar, A.A.; Cantini, C.; Cai, G.; Hausman, J.-F.; et al. Production of Plant Secondary Metabolites: Examples, Tips and Suggestions for Biotechnologists. Genes 2018, 9, 309. https://doi.org/10.3390/genes9060309
Guerriero G, Berni R, Muñoz-Sanchez JA, Apone F, Abdel-Salam EM, Qahtan AA, Alatar AA, Cantini C, Cai G, Hausman J-F, et al. Production of Plant Secondary Metabolites: Examples, Tips and Suggestions for Biotechnologists. Genes. 2018; 9(6):309. https://doi.org/10.3390/genes9060309
Chicago/Turabian StyleGuerriero, Gea, Roberto Berni, J. Armando Muñoz-Sanchez, Fabio Apone, Eslam M. Abdel-Salam, Ahmad A. Qahtan, Abdulrahman A. Alatar, Claudio Cantini, Giampiero Cai, Jean-Francois Hausman, and et al. 2018. "Production of Plant Secondary Metabolites: Examples, Tips and Suggestions for Biotechnologists" Genes 9, no. 6: 309. https://doi.org/10.3390/genes9060309
APA StyleGuerriero, G., Berni, R., Muñoz-Sanchez, J. A., Apone, F., Abdel-Salam, E. M., Qahtan, A. A., Alatar, A. A., Cantini, C., Cai, G., Hausman, J.-F., Siddiqui, K. S., Hernández-Sotomayor, S. M. T., & Faisal, M. (2018). Production of Plant Secondary Metabolites: Examples, Tips and Suggestions for Biotechnologists. Genes, 9(6), 309. https://doi.org/10.3390/genes9060309