A Novel G-Protein-Coupled Receptors Gene from Upland Cotton Enhances Salt Stress Tolerance in Transgenic Arabidopsis

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
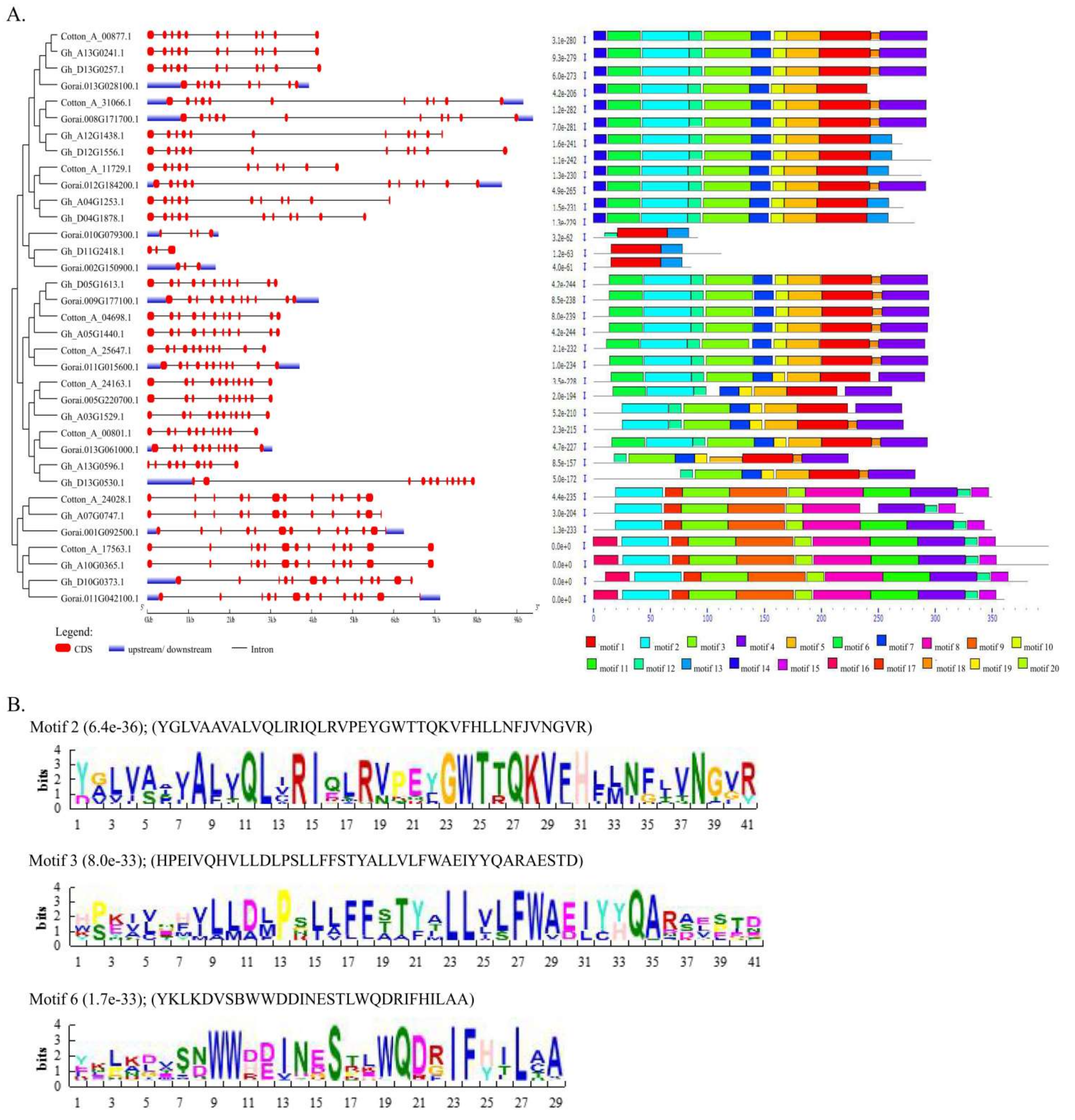
2.1. Identification and Sequence Analysis of TOM Proteins in Cotton
2.2. Chromosomal and Subcellular Localization of the TOM Protein in Cotton
2.3. Phylogenetic Analysis and Functional Classification of TOM Proteins in Cotton
2.4. Analysis of the Expression Patterns of Upland Cotton TOM Genes in Different Tissues, under Salt Stress through RNA Sequencing Data
2.5. Plant Transformation and Screening of Gh_A07G0747 (GhTOM) Gene Arabidopsis Lines
2.6. Subcellular Localization of Gh_A07G0747 (GhTOM) Protein
2.7. Response of Overexpressed Gh_A07G0747 (GhTOM) Transgenic Lines to Salt Stress Tolerance
2.8. Root Elongation and Survival Assays
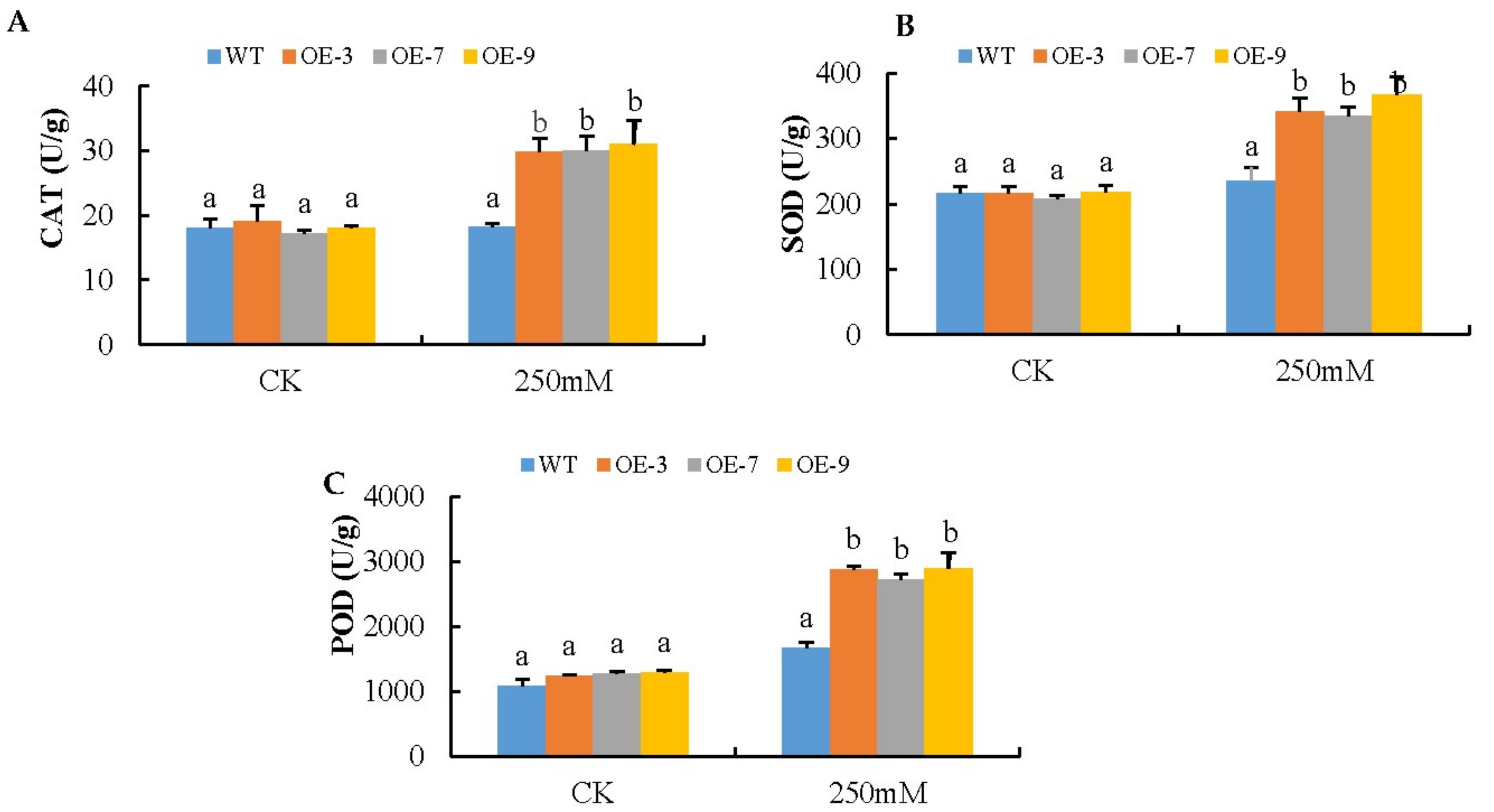
2.9. Catalase Enzymes Extraction and Assay
2.10. Superoxide Dismutase Enzymes Extraction and Assay
2.11. Chlorophyll Determination
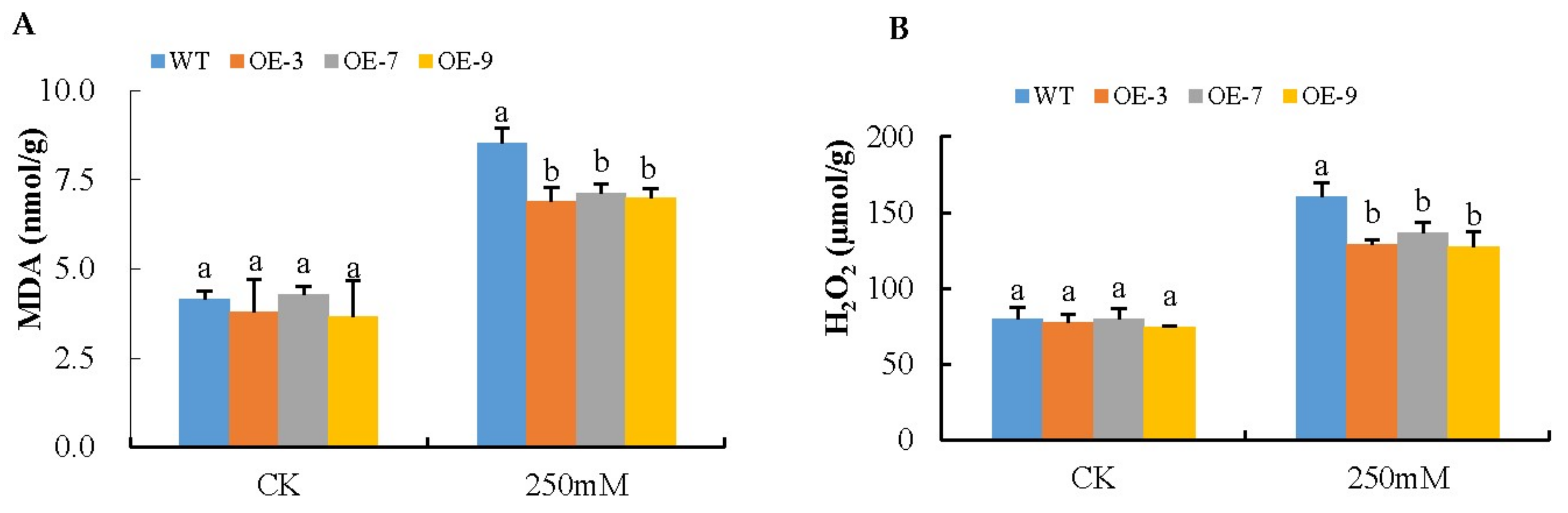
2.12. Malondialdehyde Content
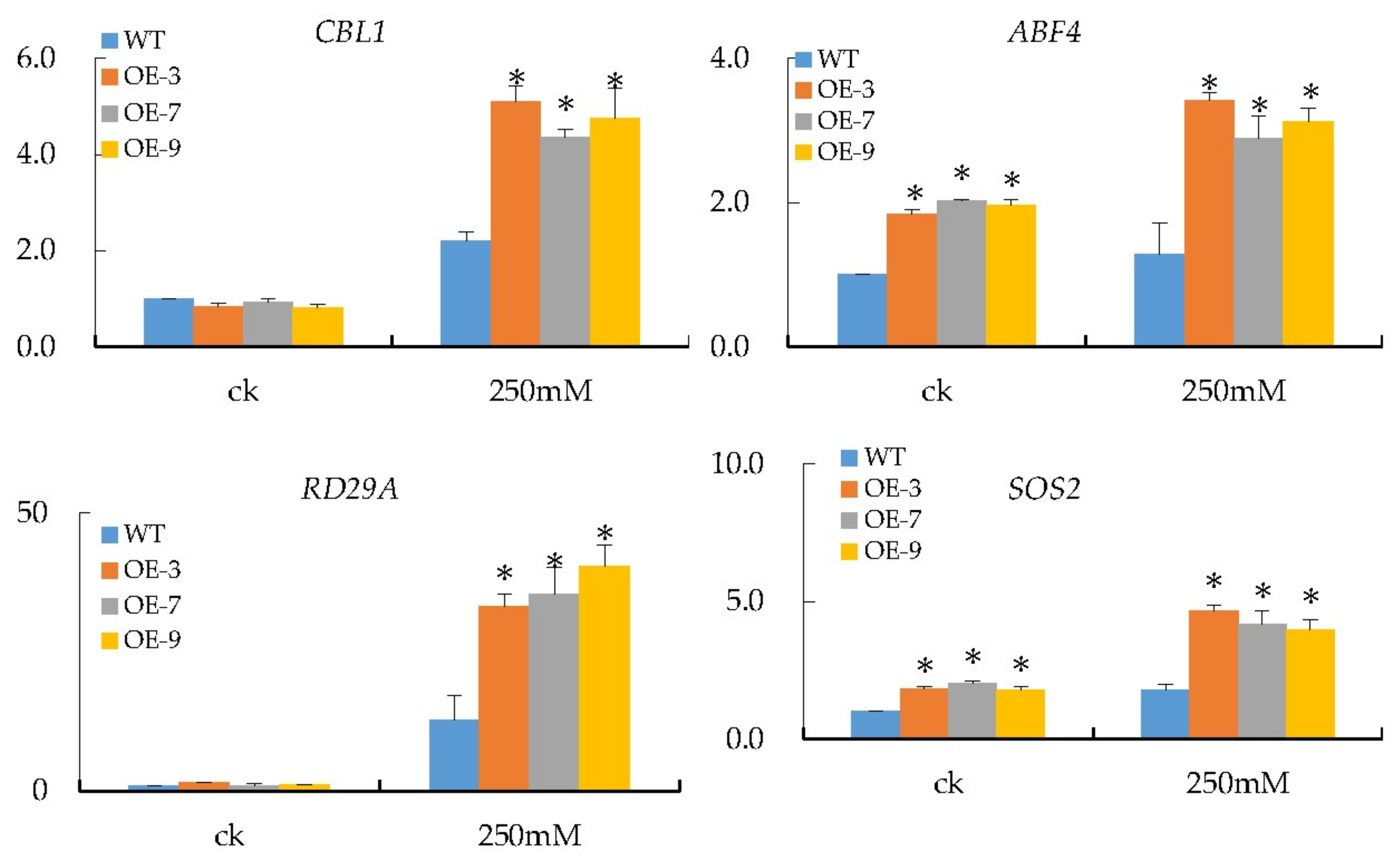
2.13. qRT-PCR Analysis of the Expression of Salt-Responsive Genes in Transgenic Arabidopsis
2.14. RNA Isolation and qRT-PCR Analysis of the Transformed Gene in Cotton Tissues
3. Results
3.1. Identification, Sequence, and Structure Analysis of TOM Candidate Proteins of G-Protein-Coupled Receptors in Cotton
3.2. Phylogenetic Analysis and Functional Classification of TOM Candidate Proteins of G-Protein-Coupled Receptors in Cotton
3.3. Chromosome Mapping and Subcellular Localization of the TOM Candidate Proteins of G-Protein-Coupled Receptors Proteins in Cotton
3.4. RNA Isolation and qRT-PCR Analysis of the Transformed Gene in Cotton Tissues
3.5. Analysis of Oxidant and Antioxidant Content in the Transgenic Lines and Wild-Type under Salt Stress Condition
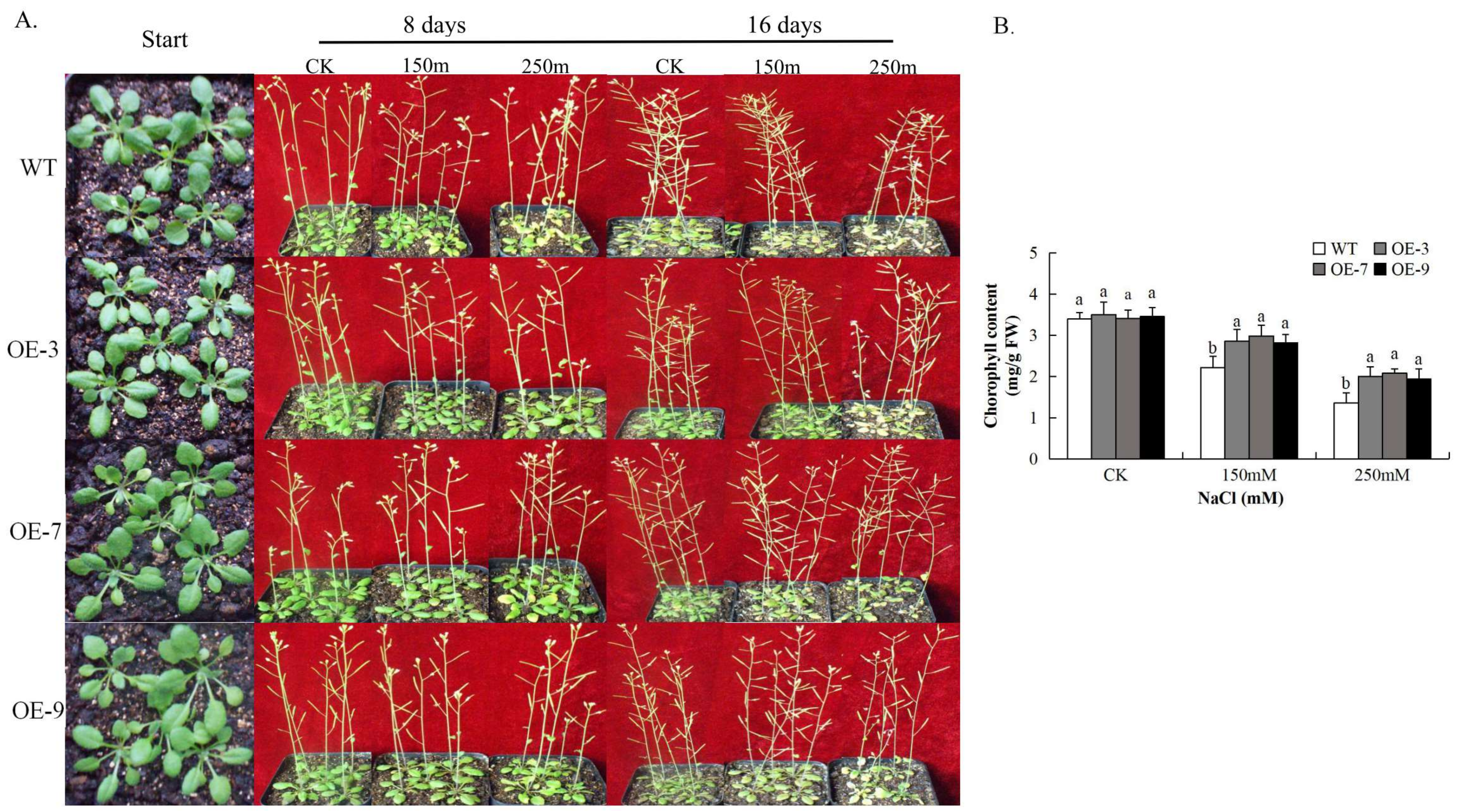
3.6. Over-Expression of Gh_A07G0747 (GhTOM) in Plants Confers Enhanced Salt Tolerance
3.7. Transcripts Investigation of Salt Stress-Responsive Genes
3.8. Analysis of Chlorophyll Content in Gh_A07G0747 (GhTOM) Transgenic Lines
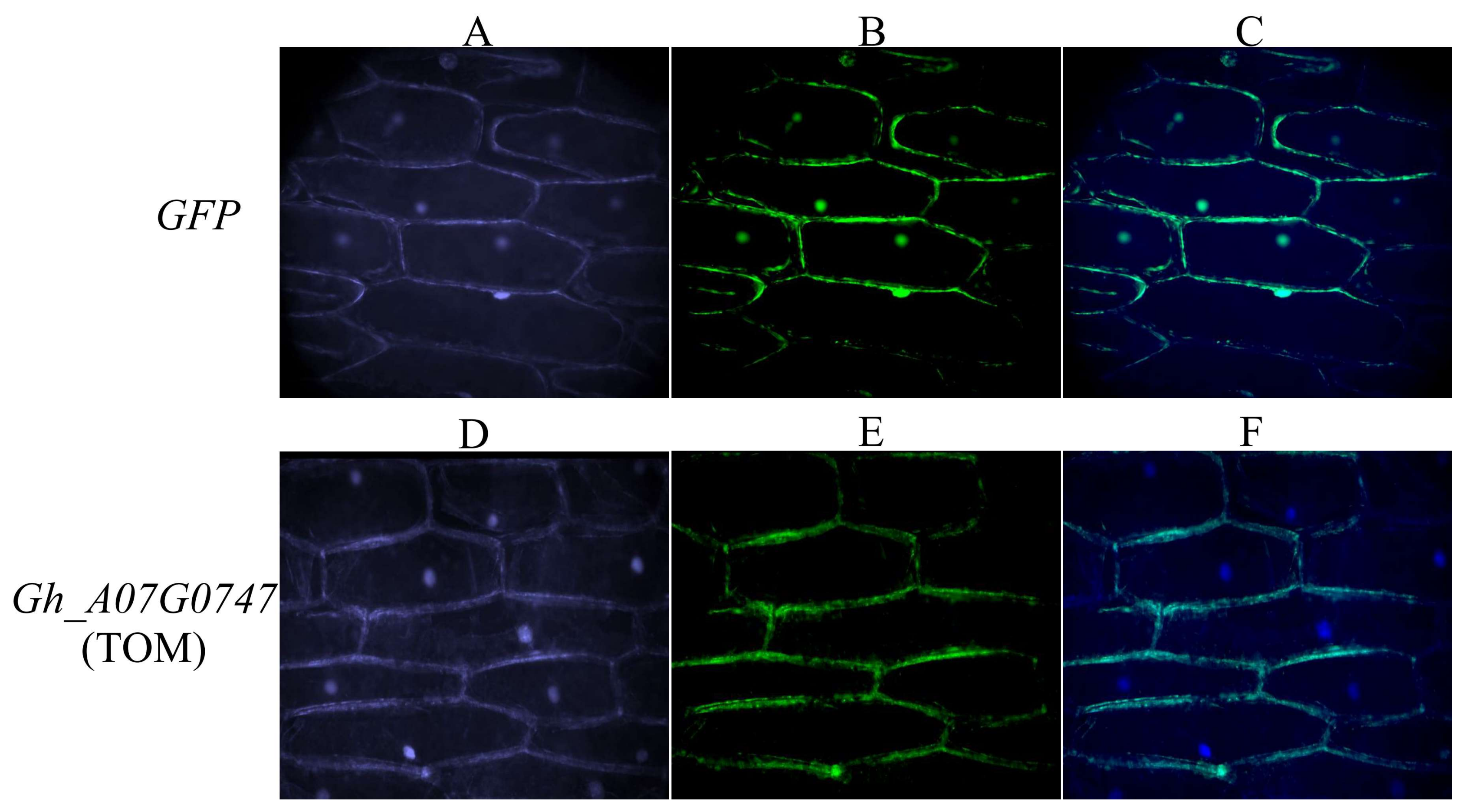
3.9. Subcellular Localization of Gh_A07G0747 (GhTOM) Protein
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Žádníková, P.; Smet, D.; Zhu, Q.; Straeten, D.; Van Der Benková, E. Strategies of seedlings to overcome their sessile nature: Auxin in mobility control. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3544. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, S.; Trewavas, A. Signal processing and transduction in plant cells: The end of the beginning? Nat. Rev. Mol. Cell Biol. 2001, 2, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Mahajan, S.; Tuteja, N.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Dangl, J.L.; Preuss, D.; Schroeder, J.I. Talking through walls: Signaling in plant development. Cell 1995, 83, 1071–1077. [Google Scholar] [CrossRef]
- Xue, C.; Hsueh, Y.P.; Heitman, J. Magnificent seven: Roles of G protein-coupled receptors in extracellular sensing in fungi. FEMS Microbiol. Rev. 2008, 32, 1010–1032. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, E.; Bouhelal, R.; Gerspacher, M.; Seuwen, K. The 7 TM G-protein-coupled receptor target family. ChemMedChem 2006, 1, 760–782. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Ullah, H.; Jones, A.M.; Assmann, S.M. G protein regulation of ion channels and abscisic acid signaling in Arabidopsis guard cells. Science 2001, 292, 2070–2072. [Google Scholar] [CrossRef] [PubMed]
- Wojcikiewicz, R.J.H. Regulated ubiquitination of proteins in GPCR-initiated signaling pathways. Trends Pharmacol. Sci. 2004, 25, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Latijnhouwers, M.; Ligterink, W.; Vleeshouwers, V.G.A.A.; Van West, P.; Govers, F. A Gα subunit controls zoospore motility and virulence in the potato late blight pathogen Phytophthora infestans. Mol. Microbiol. 2004, 51, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Trusov, Y.; Sewelam, N.; Rookes, J.E.; Kunkel, M.; Nowak, E.; Schenk, P.M.; Botella, J.R. Heterotrimeric G proteins-mediated resistance to necrotrophic pathogens includes mechanisms independent of salicylic acid-, jasmonic acid/ethylene- and abscisic acid-mediated defense signaling. Plant J. 2009, 58, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Trusov, Y.; Botella, J.R. Plant G-Proteins come of age: Breaking the bond with animal models. Front. Chem. 2016, 4, 24. [Google Scholar] [CrossRef] [PubMed]
- Urano, D.; Jones, A.M. Heterotrimeric G Protein–coupled signaling in plants. Annu. Rev. Plant Biol. 2014, 65, 365–384. [Google Scholar] [CrossRef] [PubMed]
- Taddese, B.; Upton, G.J.G.; Bailey, G.R.; Jordan, S.R.D.; Abdulla, N.Y.; Reeves, P.J.; Reynolds, C.A. Do plants contain G Protein-coupled receptors? Plant Physiol. 2014, 164, 287–307. [Google Scholar] [CrossRef] [PubMed]
- Josefsson, L.G.; Rask, L. Cloning of a putative G-protein-coupled receptor from Arabidopsis thaliana. Eur. J. Biochem. 1997, 249, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Plakidou-Dymock, S.; Dymock, D.; Hooley, R. A higher plant seven-transmembrane receptor that influences sensitivity to cytokinins. Curr. Biol. 1998, 8, 315–324. [Google Scholar] [CrossRef]
- Gookin, T.E.; Kim, J.; Assmann, S.M. Whole proteome identification of plant candidate G-protein coupled receptors in Arabidopsis, rice, and poplar: Computational prediction and in-vivo protein coupling. Genome Biol. 2008, 9, R120. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, J.M. The probable arrangement of the helices in G protein-coupled receptors. EMBO J. 1993, 12, 1693–1703. [Google Scholar] [PubMed]
- Dong, M.; Koole, C.; Wootten, D.; Sexton, P.M.; Miller, L.J. Structural and functional insights into the juxtamembranous amino-terminal tail and extracellular loop regions of class B GPCRs. Br. J. Pharmacol. 2014, 171, 1085–1101. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, M.; Wootten, D.; Conner, M.T.; Simms, J.; Kendrick, R.; Logan, R.T.; Poyner, D.R.; Barwell, J. Lifting the lid on GPCRs: The role of extracellular loops. Br. J. Pharmacol. 2012, 165, 1688–1703. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, N.; Sharma, P.; Kanyuka, K.; Pathak, R.R.; Choudhury, D.; Hooley, R.; Raghuram, N. G-protein α-subunit (GPA1) regulates stress, nitrate and phosphate response, flavonoid biosynthesis, fruit/seed development and substantially shares GCR1 regulation in A. thaliana. Plant Mol. Biol. 2015, 89, 559–576. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, N.; Singh, N.; Kaur, K.; Raghuram, N. G-protein signaling components GCR1 and GPA1 mediate responses to multiple abiotic stresses in Arabidopsis. Front. Plant Sci. 2015, 6, 1000. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Murat, U.; Md, S.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. WAO J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Gossett, D.R.; Millhollon, E.P.; Lucas, M.C. Antioxidant response to NaCl stress in salt-tolerant and salt-sensitive cultivars of cotton. Crop Sci. 1994, 34, 706–714. [Google Scholar] [CrossRef]
- Vaidyanathan, H.; Sivakumar, P.; Chakrabarty, R.; Thomas, G. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)—Differential response in salt-tolerant and sensitive varieties. Plant Sci. 2003, 165, 1411–1418. [Google Scholar] [CrossRef]
- Radi, A. Physiological and biochemical responses of salt-tolerant and salt-sensitive wheat and bean cultivars to salinity. J. Biol. 2013, 3, B72–B88. [Google Scholar]
- De La Haba, C.; Palacio, J.R.; Martínez, P.; Morros, A. Effect of oxidative stress on plasma membrane fluidity of THP-1 induced macrophages. Biochim. Biophys. Acta Biomembr. 2013, 1828, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Janero, D.R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- Emanuelsson, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef] [PubMed]
- Bodén, M.; Hawkins, J. Prediction of subcellular localization using sequence-biased recurrent networks. Bioinformatics 2005, 21, 2279–2286. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Liang, C.Z.; Meng, Z.G.; Sun, G.Q.; Meng, Z.H.; Guo, S.D.; Zhang, R. CottonFGD: An integrated functional genomics database for cotton. BMC Plant Biol. 2017, 17, 101. [Google Scholar] [CrossRef] [PubMed]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Lu, H.; Wang, X.; Cai, X.; Zhou, Z.; Zhang, Z.; Salih, H.; Wang, K.; et al. Characterization of the late embryogenesis abundant (LEA) proteins family and their role in drought stress tolerance in upland cotton. BMC Genet. 2018, 19, 6. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Bodén, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Moradpour, M.; Abdulah, S.N.A. Evaluation of pEASY-Uni Seamless Cloning and Assembly Kit to clone multiple fragments of Elaeis guineensis DNA. Meta Gene 2017, 14, 134–141. [Google Scholar] [CrossRef]
- Benoit, R.M.; Ostermeier, C.; Geiser, M.; Li, J.S.Z.; Widmer, H.; Auer, M. Seamless insert-plasmid assembly at high efficiency and low cost. PLoS ONE 2016, 11, e0153158. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Cao, Z.; Li, Y.; Zhao, Y.; Zhang, H. A simple and effective method for protein subcellular localization using Agrobacterium-mediated transformation of onion epidermal cells. Biologia (Bratisl.) 2007, 62, 529–532. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Rengel, Z. Drought and salinity differentially influence activities of superoxide dismutases in narrow-leafed lupins. Plant Sci. 1999, 142, 1–11. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: II. purification and quantitative relationship with water-soluble protein in seedlings. Plant Physiol. 1977, 59, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, R.; Kumar, S.; Ahuja, P.S. Induction of a new isozyme of superoxide dismutase at low temperature in Potentilla astrisanguinea Lodd. variety argyrophylla (Wall. ex. Lehm) Griers. J. Plant Physiol. 2001, 158, 1093–1097. [Google Scholar] [CrossRef]
- Kakkar, P.; Das, B.; Viswanathan, P.N. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys. 1984, 21, 130–132. [Google Scholar] [CrossRef] [PubMed]
- Tetley, R.M.; Thimann, K.V. The metabolism of oat leaves during senescence: I. respiration, carbohydrate metabolism, and the action of cytokinins. Plant Physiol. 1974, 54, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Singh, V.K.; Mangalam, A.K.; Dwivedi, S.; Naik, S. Primer premier: Program for design of degenerate primers from a protein sequence. Biotechniques 1998, 24, 318–319. [Google Scholar] [PubMed]
- Tuteja, N. Signaling through G protein coupled receptors. Plant. Signal. Behav. 2009, 4, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Mmadi, M.; Dossa, K.; Wang, L.; Zhou, R.; Wang, Y.; Cisse, N.; Sy, M.; Zhang, X. Functional characterization of the versatile MYB gene family uncovered their important roles in plant development and responses to drought and waterlogging in sesame. Genes (Basel) 2017, 8, 362. [Google Scholar] [CrossRef] [PubMed]
- Revell, L.J.; Harmon, L.J.; Collar, D.C. Phylogenetic signal, evolutionary process, and rate. Syst. Biol. 2008, 57, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Al-aghabary, K.; Zhu, Z.; Shi, Q. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. J. Plant Nutr. 2004, 27, 2101–2115. [Google Scholar] [CrossRef]
- Maggio, A.; Hasegawa, P.M.; Bressan, R.A.; Consiglio, M.F.; Joly, R.J. Unravelling the functional relationship between root anatomy and stress tolerance. Aust. J. Plant Physiol. 2001, 28, 999–1004. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Farooq, S.; Azam, F. The use of cell membrane stability (CMS) technique to screen for salt tolerant wheat varieties. J. Plant Physiol. 2006, 163, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T.; Ma, S.L.; Bai, L.P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Vassilatis, D.K.; Hohmann, J.G.; Zeng, H.; Li, F.; Ranchalis, J.E.; Mortrud, M.T.; Brown, A.; Rodriguez, S.S.; Weller, J.R.; Wright, A.C.; et al. The G protein-coupled receptor repertoires of human and mouse. Proc. Natl. Acad. Sci. USA 2003, 100, 4903–4908. [Google Scholar] [CrossRef] [PubMed]
- Häuser, R.; Pech, M.; Kijek, J.; Yamamoto, H.; Titz, B.; Naeve, F.; Tovchigrechko, A.; Yamamoto, K.; Szaflarski, W.; Takeuchi, N.; et al. RsFA (YbeB) proteins are conserved ribosomal silencing factors. PLoS Genet. 2012, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Guo, S.R.; Sun, J.; Yuan, L.Y. Effects of salt stress on the structure and function of the photosynthetic apparatus in Cucumis sativus and its protection by exogenous putrescine. Physiol. Plant 2012, 146, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Bagag, A.; Jault, J.M.; Sidahmed-Adrar, N.; Réfrégiers, M.; Giuliani, A.; Le Naour, F. Characterization of hydrophobic peptides in the presence of detergent by photoionization mass spectrometry. PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, J.E.; Jang, H.Y.; Kwak, K.J.; Ahn, S.J. CsRCI2A and CsRCI2E genes show opposite salt sensitivity reaction due to membrane potential control. Acta Physiol. Plant. 2016, 38, 1–13. [Google Scholar] [CrossRef]
- Yamanaka, T.; Ohta, T.; Takahashi, M.; Meshi, T.; Schmidt, R.; Dean, C.; Naito, S.; Ishikawa, M. TOM1, an Arabidopsis gene required for efficient multiplication of a tobamovirus, encodes a putative transmembrane protein. Proc. Natl. Acad. Sci. USA 2000, 97, 10107–10112. [Google Scholar] [CrossRef] [PubMed]
- Madhusudhan, K.N.; Srikanta, B.M.; Shylaja, M.D.; Prakash, H.S.; Shetty, H.S. Changes in antioxidant enzymes, hydrogen peroxide, salicylic acid and oxidative stress in compatible and incompatible host-tobamovirus interaction. J. Plant Interact. 2009, 4, 157–166. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Mathur, G.; Koul, S.; Sarin, N.B. Ameliorative effects of proline on salt stress-induced lipid peroxidation in cell lines of groundnut (Arachis hypogaea L.). Plant Cell Rep. 2001, 20, 463–468. [Google Scholar] [CrossRef]
- Chen, X.; Lu, X.; Shu, N.; Wang, D.; Wang, S.; Wang, J.; Guo, L.; Guo, X.; Fan, W.; Lin, Z.; et al. GhSOS1, a plasma membrane Na+/H+ antiporter gene from upland cotton, enhances salt tolerance in transgenic Arabidopsis thaliana. PLoS ONE 2017, 12, e0181450. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2007, 58, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, N.; Sharma, P.; Kanyuka, K.; Pathak, R.R.; Choudhury, D.; Hooley, R.A.; Raghuram, N. Transcriptome analysis of Arabidopsis GCR1 mutant reveals its roles in stress, hormones, secondary metabolism and phosphate starvation. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Assmann, S.M. The Arabidopsis putative G protein-coupled receptor GCR1 interacts with the G protein α subunit GPA1 and regulates abscisic acid signaling. Plant Cell 2004, 16, 1616–1632. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Kishimoto, T.; Mizuno, T.; Shinzato, T.; Uemura, H. Expression of GCR1, the transcriptional activator of glycolytic enzyme genes in the yeast Saccharomyces cerevisiae, is positively autoregulated by Gcr1p. Yeast 2005, 22, 305–319. [Google Scholar] [CrossRef] [PubMed]
- Huie, M.A.; Scott, E.W.; Drazinic, C.M.; Lopez, M.C.; Hornstra, I.K.; Yang, T.P.; Baker, H.V. Characterization of the DNA-binding activity of GCR1: In vivo evidence for two GCR1-binding sites in the upstream activating sequence of TPI of Saccharomyces cerevisiae. Mol. Cell. Biol. 1992, 12, 2690–2700. [Google Scholar] [CrossRef] [PubMed]
- Rögner, M.; Boekema, E.J.; Barber, J. How does photosystem 2 split water? The structural basis of efficient energy conversion. Trends Biochem. Sci. 1996, 21, 44–49. [Google Scholar] [CrossRef]
- Mozafariyan, M.; Saghafi, K.; Esfandiari, A.; Bakhtiari, S. The effects of different sodium Chloride concentrations on the growth and photosynthesis parameters of tomato (Lycopersicum esculentum cv. Foria). Agric. Crop. Sci. 2013, 6, 203–207. [Google Scholar]
- Abdullah, Z.; Ahmad, R. Effect of pre- and post-kinetin treatments on salt tolerance of different potato cultivars growing on saline soils. J. Agron. Crop. Sci. 1990, 165, 94–102. [Google Scholar] [CrossRef]
- Reddy, M.P.; Vora, A.B. Effect of salinity on protein metabolism in bajara (Pennisetum typhoides S and H) leaves. Indian J. Plant. Physiol. 1985, 28, 190–195. [Google Scholar]
- Hashimoto, K.; Eckert, C.; Anschütz, U.; Scholz, M.; Held, K.; Waadt, R.; Reyer, A.; Hippler, M.; Becker, D.; Kudla, J. Phosphorylation of calcineurin B-like (CBL) calcium sensor proteins by their CBL-interacting protein kinases (CIPKs) is required for full activity of CBL–CIPK complexes toward their target proteins. J. Biol. Chem. 2012, 287, 7956–7968. [Google Scholar] [CrossRef] [PubMed]
- Batistič, O.; Kudla, J. Plant calcineurin B-like proteins and their interacting protein kinases. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Gu, D.; Liu, T.; Wang, Z.; Guo, X.; Hou, W.; Bai, Y.; Chen, X.; Wang, G. Overexpression of a putative maize calcineurin B-like protein in Arabidopsis confers salt tolerance. Plant Mol. Biol. 2007, 65, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.K.; Cheong, Y.H.; Kim, K.-N.; Grant, J.J.; Li, L.; Hung, W.; D’Angelo, C.; Weinl, S.; Kudla, J.; Luan, S. The calcium sensor calcineurin B-like 9 modulates abscisic acid sensitivity and biosynthesis in Arabidopsis. Plant Cell 2004, 16, 1912–1924. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.K.; Kanwar, P.; Singh, A.; Steinhorst, L.; Pandey, A.; Yadav, A.K.; Tokas, I.; Sanyal, S.K.; Kim, B.G.; Lee, S.C.; et al. Calcineurin B-like protein-interacting protein kinase CIPK21 regulates osmotic and salt stress responses in Arabidopsis. Plant Physiol. 2015, 169, 780–792. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Park, H.J.; Park, J.H.; Kim, S.; Im, M.Y.; Seo, H.H.; Kim, Y.W.; Hwang, I.; Kim, S.Y. Arabidopsis calcium-dependent protein kinase AtCPK32 interacts with ABF4, a transcriptional regulator of abscisic acid-responsive gene expression, and modulates its activity. Plant Physiol. 2005, 139, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Uno, Y.; Furihata, T.; Abe, H.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef] [PubMed]
- Msanne, J.; Lin, J.; Stone, J.M.; Awada, T. Characterization of abiotic stress-responsive Arabidopsis thaliana RD29A and RD29B genes and evaluation of transgenes. Planta 2011, 234, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Virlouvet, L.; Ding, Y.; Fujii, H.; Avramova, Z.; Fromm, M. ABA signaling is necessary but not sufficient for RD29B transcriptional memory during successive dehydration stresses in Arabidopsis thaliana. Plant J. 2014, 79, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Winicov, I. Alfin1 transcription factor overexpression enhances plant root growth under normal and saline conditions and improves salt tolerance in alfalfa. Planta 2000, 210, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Pasapula, V.; Shen, G.; Kuppu, S.; Paez-Valencia, J.; Mendoza, M.; Hou, P.; Chen, J.; Qiu, X.; Zhu, L.; Zhang, X.; et al. Expression of an Arabidopsis vacuolar H+-pyrophosphatase gene (AVP1) in cotton improves drought- and salt tolerance and increases fibre yield in the field conditions. Plant Biotechnol. J. 2011, 9, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Zelazny, A.M.; Shaish, A.; Pick, U. Plasma membrane sterols are essential for sensing osmotic changes in the halotolerant alga Dunaliella. Plant Physiol. 1995, 109, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Cho, E.J.; Wi, S.G.; Bae, H.; Kim, J.E.; Cho, J.Y.; Lee, S.; Kim, J.H.; Chung, B.Y. Divergences in morphological changes and antioxidant responses in salt-tolerant and salt-sensitive rice seedlings after salt stress. Plant Physiol. Biochem. 2013, 70, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Mitsuya, S.; Takeoka, Y.; Miyake, H. Effects of sodium chloride on foliar ultrastructure of sweet potato (Ipomoea batatas Lam.) plantlets grown under light and dark conditions in vitro. J. Plant Physiol. 2000, 157, 661–667. [Google Scholar] [CrossRef]
- Salman, V.; Amann, R.; Shub, D.A.; Schulz-Vogt, H.N. Multiple self-splicing introns in the 16S rRNA genes of giant sulfur bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 4203–4208. [Google Scholar] [CrossRef] [PubMed]
- Urano, D.; Jones, A.M. “Round up the usual suspects”: A comment on nonexistent plant G protein-coupled receptors. Plant Physiol. 2013, 161, 1097–1102. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Sequence | Reverse Sequence |
|---|---|---|
| AtABF4 | 3′-AACAACTTAGGAGGTGGTGGTCAT-5′ | 3′-TGTAGCAGCTGGCGCAGAAGTCAT-5′ |
| AtSOS2 | 3′-GGCTTGAAGAAAGTGAGTCTCG-5′ | 3′-GCTACATAGTTCGGAGTTCCACA-5′ |
| AtCBL1 | 3′-GAAATGAAACTGGCTGATGAAACCATAGAG-5′ | 3′-CTCGTGGCAATCTACTCGGTCTTAAACC-5′ |
| AtRD29A | 3′-TGAAAGGAGGAGGAGGAATGGTTGG-5′ | 3′-ACAAAACACACATAAACATCCAAAGT-5′ |
| Gene Type | Prediction | Family | Sub Family | Sub Family Type | Gossypium hirsutum | Gossypium arboreum | Gossypium raimondii |
|---|---|---|---|---|---|---|---|
| TOM1 | GPCR | class A | Olfactory | Olfactory II family 10 | 8 | 6 | 6 |
| TOM3 | GPCR | class A | Olfactory | Olfactory II family 5 | 7 | 3 | 4 |
| THH1 | GPCR | class A | Peptide | - | 0 | 0 | 1 |
| Total | 15 | 9 | 11 | ||||
| Gene ID | Gene Name | Chromosome Locations | TargetP | Pprowler | Wolfpsort | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chr. | Start | End | Len | cTP | mTP | SP | other | Loc | ||||
| Gh_A03G1529 | TOM1 | A03 | 95,951,060 | 95,954,035 | 271 | 0.022 | 0.339 | 0.129 | 0.743 | _ | SP | plas |
| Gh_A04G1253 | TOM3 | A04 | 62,591,785 | 62,597,706 | 272 | 0.025 | 0.028 | 0.107 | 0.986 | _ | OTHER | plas |
| Gh_A05G1440 | TOM1 | A05 | 14,950,490 | 14,953,714 | 291 | 0.016 | 0.051 | 0.433 | 0.952 | _ | SP | plas |
| Gh_A07G0747 | TOM1 | A07 | 11,687,587 | 11,693,293 | 325 | 0.003 | 0.052 | 0.924 | 0.044 | S | SP | plas |
| Gh_A10G0365 | TOM1 | A10 | 3,340,797 | 3,347,772 | 396 | 0.003 | 0.056 | 0.751 | 0.59 | S | SP | plas |
| Gh_A12G1438 | TOM3 | A12 | 73,175,985 | 73,183,181 | 268 | 0.019 | 0.035 | 0.176 | 0.986 | _ | OTHER | plas |
| Gh_A13G0241 | TOM3 | A13 | 2,823,366 | 2,827,544 | 289 | 0.006 | 0.075 | 0.218 | 0.981 | _ | OTHER | plas |
| Gh_A13G0596 | TOM1 | A13 | 14,111,783 | 14,113,994 | 224 | 0.013 | 0.153 | 0.191 | 0.98 | _ | SP | plas |
| Gh_D04G1878 | TOM3 | D04 | 51,121,020 | 51,126,349 | 282 | 0.021 | 0.031 | 0.114 | 0.987 | _ | OTHER | plas |
| Gh_D05G1613 | TOM1 | D05 | 14,554,705 | 14,557,876 | 291 | 0.014 | 0.059 | 0.387 | 0.956 | _ | SP | plas |
| Gh_D10G0373 | TOM1 | D10 | 3,299,454 | 3,305,915 | 377 | 0.007 | 0.228 | 0.116 | 0.967 | _ | SP | plas |
| Gh_D11G2418 | TOM3 | D11 | 48,310,923 | 48,311,606 | 112 | 0.017 | 0.406 | 0.232 | 0.41 | _ | SP | plas |
| Gh_D12G1556 | TOM3 | D12 | 46,513,482 | 46,522,251 | 293 | 0.018 | 0.033 | 0.177 | 0.987 | _ | OTHER | plas |
| Gh_D13G0257 | TOM3 | D13 | 2,477,467 | 2,481,698 | 289 | 0.006 | 0.081 | 0.181 | 0.983 | _ | OTHER | plas |
| Gh_D13G0530 | TOM1 | D13 | 6,994,712 | 7,002,690 | 282 | 0.142 | 0.054 | 0.219 | 0.771 | _ | SP | plas |
| Cotton_A_00801 | TOM1 | Chr13 | 71,968,383 | 71,971,076 | 272 | 0.011 | 0.263 | 0.368 | 0.621 | _ | SP | plas |
| Cotton_A_00877 | TOM3 | Chr13 | 73,444,587 | 73,448,764 | 290 | 0.005 | 0.078 | 0.229 | 0.978 | _ | OTHER | plas |
| Cotton_A_04698 | TOM1 | Chr10 | 20,294,407 | 20,297,646 | 292 | 0.014 | 0.059 | 0.387 | 0.956 | _ | SP | plas |
| Cotton_A_11729 | TOM3 | Chr07 | 106,242,296 | 106,246,957 | 288 | 0.025 | 0.028 | 0.117 | 0.985 | _ | OTHER | plas |
| Cotton_A_17563 | TOM1 | Chr09 | 89,683,765 | 89,690,744 | 396 | 0.003 | 0.056 | 0.751 | 0.59 | S | SP | plas |
| Cotton_A_24028 | TOM1 | Chr01 | 107,338,814 | 107,344,313 | 346 | 0.003 | 0.052 | 0.924 | 0.044 | S | SP | plas |
| Cotton_A_24163 | TOM1 | Chr05 | 48,844,703 | 48,847,741 | 291 | 0.01 | 0.07 | 0.264 | 0.931 | _ | SP | plas |
| Cotton_A_25647 | TOM1 | Chr09 | 74,805,749 | 74,808,633 | 291 | 0.016 | 0.096 | 0.34 | 0.953 | _ | SP | plas |
| Cotton_A_31066 | TOM3 | Chr06 | 58,856,809 | 58,865,968 | 289 | 0.017 | 0.037 | 0.169 | 0.987 | _ | OTHER | plas |
| Gorai.001G092500 | TOM1 | Chr01 | 10,292,446 | 10,298,692 | 350 | 0.003 | 0.052 | 0.924 | 0.044 | S | SP | plas |
| Gorai.002G150900 | TOM3 | Chr02 | 29,604,352 | 29,606,009 | 86 | 0.012 | 0.477 | 0.068 | 0.448 | M | mTP | cyto |
| Gorai.005G220700 | TOM1 | Chr05 | 60,350,693 | 60,353,743 | 262 | 0.014 | 0.091 | 0.299 | 0.92 | _ | SP | plas |
| Gorai.008G171700 | TOM3 | Chr08 | 44,664,816 | 44,674,208 | 289 | 0.019 | 0.034 | 0.156 | 0.988 | _ | OTHER | plas |
| Gorai.009G177100 | TOM1 | Chr09 | 13,701,332 | 13,705,507 | 292 | 0.014 | 0.059 | 0.387 | 0.956 | _ | SP | plas |
| Gorai.010G079300 | TOM3 | Chr10 | 11,503,072 | 11,504,804 | 92 | 0.009 | 0.4 | 0.436 | 0.05 | S | SP | extr |
| Gorai.011G015600 | TOM1 | Chr11 | 1,095,851 | 1,099,556 | 291 | 0.018 | 0.061 | 0.407 | 0.907 | _ | SP | plas |
| Gorai.011G042100 | TOM1 | Chr11 | 3,118,826 | 3,125,957 | 357 | 0.003 | 0.056 | 0.751 | 0.59 | S | SP | plas |
| Gorai.012G184200 | TOM3 | Chr12 | 35,143,300 | 35,151,941 | 289 | 0.021 | 0.033 | 0.086 | 0.989 | _ | OTHER | plas |
| Gorai.013G028100 | THH1 | Chr13 | 2,095,047 | 2,098,979 | 243 | 0.007 | 0.079 | 0.173 | 0.984 | _ | OTHER | plas |
| Gorai.013G061000 | TOM1 | Chr13 | 6,622,078 | 6,625,118 | 293 | 0.013 | 0.16 | 0.274 | 0.833 | _ | SP | plas |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, P.; Magwanga, R.O.; Lu, H.; Kirungu, J.N.; Wei, Y.; Dong, Q.; Wang, X.; Cai, X.; Zhou, Z.; Wang, K.; et al. A Novel G-Protein-Coupled Receptors Gene from Upland Cotton Enhances Salt Stress Tolerance in Transgenic Arabidopsis. Genes 2018, 9, 209. https://doi.org/10.3390/genes9040209
Lu P, Magwanga RO, Lu H, Kirungu JN, Wei Y, Dong Q, Wang X, Cai X, Zhou Z, Wang K, et al. A Novel G-Protein-Coupled Receptors Gene from Upland Cotton Enhances Salt Stress Tolerance in Transgenic Arabidopsis. Genes. 2018; 9(4):209. https://doi.org/10.3390/genes9040209
Chicago/Turabian StyleLu, Pu, Richard Odongo Magwanga, Hejun Lu, Joy Nyangasi Kirungu, Yangyang Wei, Qi Dong, Xingxing Wang, Xiaoyan Cai, Zhongli Zhou, Kunbo Wang, and et al. 2018. "A Novel G-Protein-Coupled Receptors Gene from Upland Cotton Enhances Salt Stress Tolerance in Transgenic Arabidopsis" Genes 9, no. 4: 209. https://doi.org/10.3390/genes9040209
APA StyleLu, P., Magwanga, R. O., Lu, H., Kirungu, J. N., Wei, Y., Dong, Q., Wang, X., Cai, X., Zhou, Z., Wang, K., & Liu, F. (2018). A Novel G-Protein-Coupled Receptors Gene from Upland Cotton Enhances Salt Stress Tolerance in Transgenic Arabidopsis. Genes, 9(4), 209. https://doi.org/10.3390/genes9040209