Beyond Transcription: Fine-Tuning of Circadian Timekeeping by Post-Transcriptional Regulation

 , ,
, , 
Abstract
1. Introduction
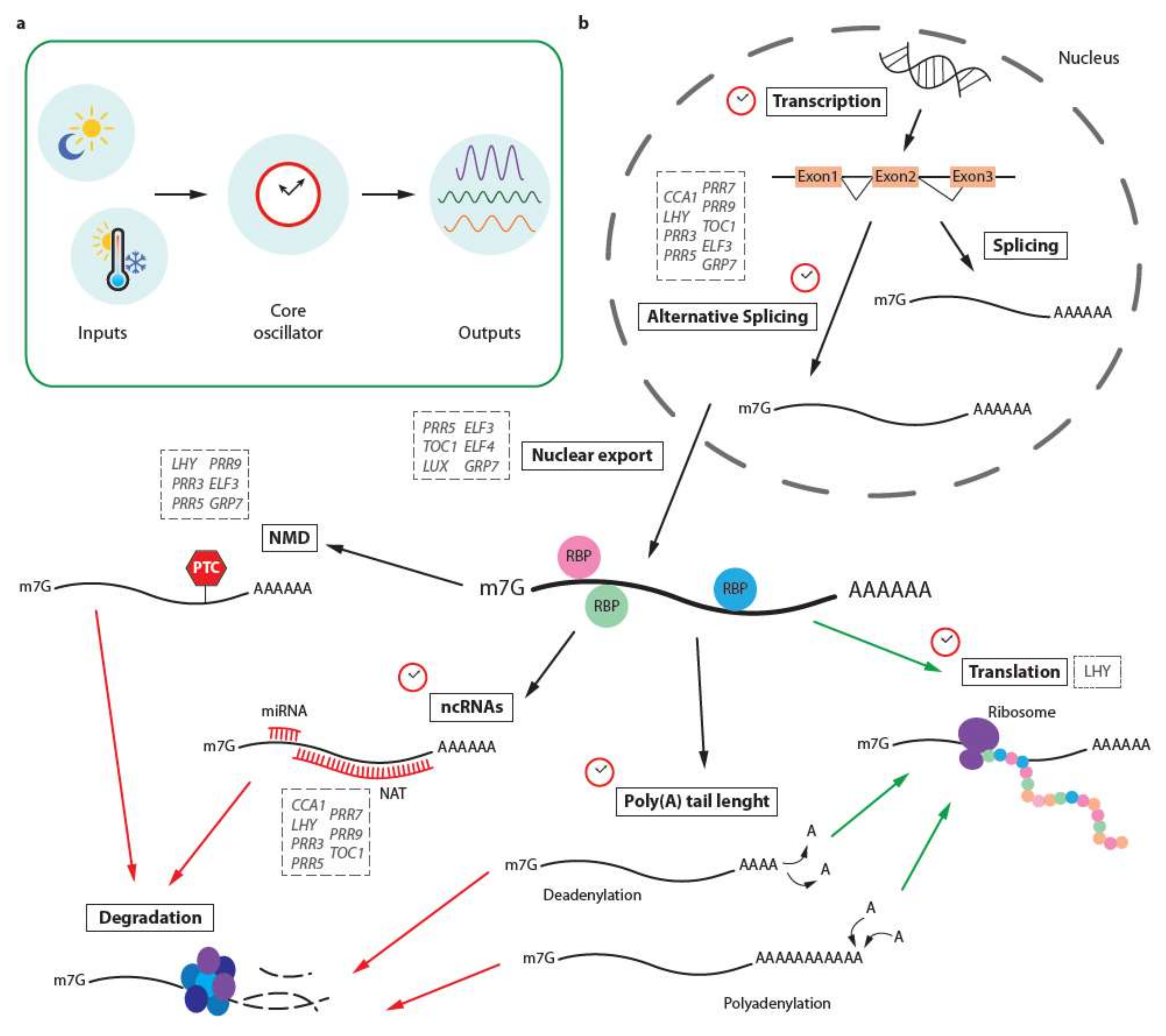
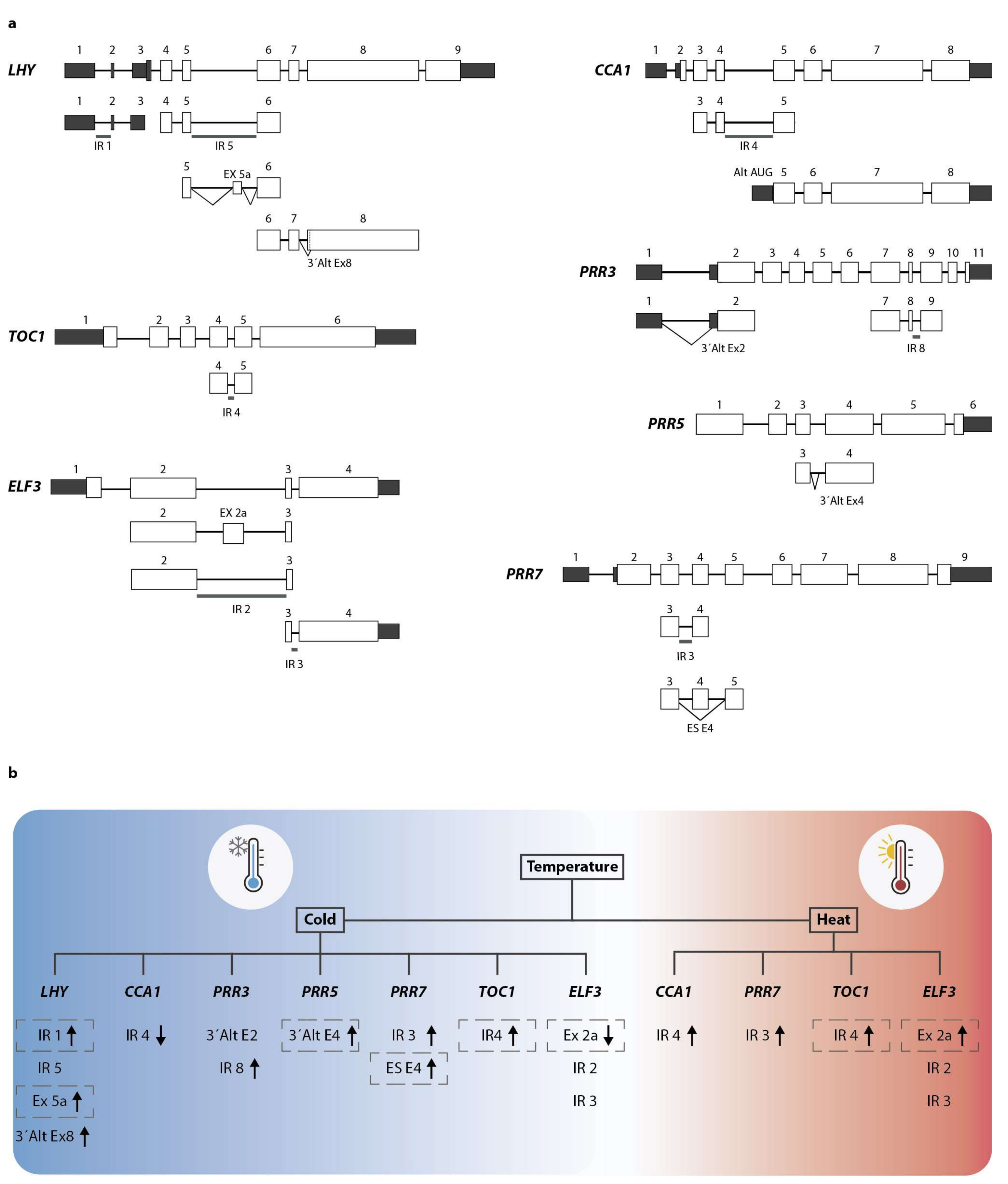
2. Post-Transcriptional Control of the Circadian Clock by Alternative Splicing and Its Connection with Environmental Responses
3. Nonsense-Mediated Decay Fine-Tuning Circadian Rhythms
3.1. Nonsense Mediated Decay in Circadian Control of Transcript Expression
3.2. Nonsense-Mediated Decay Modulating the Levels of Core Clock Genes
4. Nuclear Transport as a Mechanism Controlling Clock Genes
5. Circadian Regulation of Translation
6. Rhythmic Post-Transcriptional Control by Non-Coding RNAs
7. Final Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- McClung, C.R. Plant circadian rhythms. Plant Cell 2006, 18, 792–803. [Google Scholar] [CrossRef] [PubMed]
- Bloch, G.; Bar-Shai, N.; Cytter, Y.; Green, R. Time is honey: Circadian clocks of bees and flowers and how their interactions may influence ecological communities. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef]
- Nohales, M.A.; Kay, S.A. Molecular mechanisms at the core of the plant circadian oscillator. Nat. Struct. Mol. Biol. 2016, 23, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.; Koster, T. On the move through time—A historical review of plant clock research. Plant Biol. (Stuttg.) 2018. [Google Scholar] [CrossRef] [PubMed]
- Danisman, S.; Mateos, J.L.; Staiger, D. Time to Network: The Molecular Blueprint of the Circadian Timing System in Plants. In Rhythms in Plants; Mancuso, S., Shabala, S., Eds.; Springer International Publishing: New York, NY, USA, 2015; pp. 257–278. [Google Scholar]
- Alabadi, D.; Oyama, T.; Yanovsky, M.J.; Harmon, F.G.; Mas, P.; Kay, S.A. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 2001, 293, 880–883. [Google Scholar] [CrossRef] [PubMed]
- Gendron, J.M.; Pruneda-Paz, J.L.; Doherty, C.J.; Gross, A.M.; Kang, S.E.; Kay, S.A. Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proc. Natl. Acad. Sci. USA 2012, 109, 3167–3172. [Google Scholar] [CrossRef]
- Huang, W.; Perez-Garcia, P.; Pokhilko, A.; Millar, A.J.; Antoshechkin, I.; Riechmann, J.L.; Mas, P. Mapping the core of the Arabidopsis circadian clock defines the network structure of the oscillator. Science 2012, 336, 75–79. [Google Scholar] [CrossRef]
- Adams, S.; Manfield, I.; Stockley, P.; Carre, I.A. Revised Morning Loops of the Arabidopsis Circadian Clock Based on Analyses of Direct Regulatory Interactions. PLoS ONE 2015, 10, e0143943. [Google Scholar] [CrossRef]
- Farre, E.M.; Harmer, S.L.; Harmon, F.G.; Yanovsky, M.J.; Kay, S.A. Overlapping and distinct roles of PRR7 and PRR9 in the Arabidopsis circadian clock. Curr. Biol. 2005, 15, 47–54. [Google Scholar] [CrossRef]
- Herrero, E.; Kolmos, E.; Bujdoso, N.; Yuan, Y.; Wang, M.; Berns, M.C.; Uhlworm, H.; Coupland, G.; Saini, R.; Jaskolski, M.; et al. EARLY FLOWERING4 recruitment of EARLY FLOWERING3 in the nucleus sustains the Arabidopsis circadian clock. Plant Cell 2012, 24, 428–443. [Google Scholar] [CrossRef]
- Dixon, L.E.; Knox, K.; Kozma-Bognar, L.; Southern, M.M.; Pokhilko, A.; Millar, A.J. Temporal repression of core circadian genes is mediated through EARLY FLOWERING 3 in Arabidopsis. Curr. Biol. 2011, 21, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Helfer, A.; Nusinow, D.A.; Chow, B.Y.; Gehrke, A.R.; Bulyk, M.L.; Kay, S.A. LUX ARRHYTHMO encodes a nighttime repressor of circadian gene expression in the Arabidopsis core clock. Curr. Biol. 2011, 21, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Mas, P. Circadian clock function in Arabidopsis thaliana: Time beyond transcription. Trends Cell Biol. 2008, 18, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Schoning, J.C.; Staiger, D. At the pulse of time: Protein interactions determine the pace of circadian clocks. FEBS Lett. 2005, 579, 3246–3252. [Google Scholar] [CrossRef] [PubMed]
- Michael, T.P.; McClung, C.R. Enhancer trapping reveals widespread circadian clock transcriptional control in Arabidopsis. Plant Physiol. 2003, 132, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.J.; Kay, S.A. Circadian Control of cab Gene Transcription and mRNA Accumulation in Arabidopsis. Plant Cell 1991, 3, 541–550. [Google Scholar] [CrossRef]
- Michael, T.P.; McClung, C.R. Phase-specific circadian clock regulatory elements in Arabidopsis. Plant Physiol. 2002, 130, 627–638. [Google Scholar] [CrossRef]
- Zhong, H.H.; Resnick, A.S.; Straume, M.; Robertson McClung, C. Effects of synergistic signaling by phytochrome A and cryptochrome1 on circadian clock-regulated catalase expression. Plant Cell 1997, 9, 947–955. [Google Scholar] [CrossRef]
- Pilgrim, M.L.; McClung, C.R. Differential Involvement of the Circadian Clock in the Expression of Genes Required for Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase Synthesis, Assembly, and Activation in Arabidopsis thaliana. Plant Physiol. 1993, 103, 553–564. [Google Scholar] [CrossRef]
- Henriques, R.; Mas, P. Chromatin remodeling and alternative splicing: Pre- and post-transcriptional regulation of the Arabidopsis circadian clock. Semin. Cell Dev. Biol. 2013, 24, 399–406. [Google Scholar] [CrossRef]
- Hernando, C.E.; Romanowski, A.; Yanovsky, M.J. Transcriptional and post-transcriptional control of the plant circadian gene regulatory network. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Nolte, C.; Staiger, D. RNA around the clock—Regulation at the RNA level in biological timing. Front. Plant Sci. 2015, 6, 311. [Google Scholar] [CrossRef] [PubMed]
- Romanowski, A.; Yanovsky, M.J. Circadian rhythms and post-transcriptional regulation in higher plants. Front. Plant Sci. 2015, 6, 437. [Google Scholar] [CrossRef] [PubMed]
- Staiger, D. RNA-binding proteins and circadian rhythms in Arabidopsis thaliana. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 1755–1759. [Google Scholar] [CrossRef]
- Koncz, C.; Dejong, F.; Villacorta, N.; Szakonyi, D.; Koncz, Z. The spliceosome-activating complex: Molecular mechanisms underlying the function of a pleiotropic regulator. Front. Plant Sci. 2012, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Will, C.L.; Luhrmann, R. Spliceosome structure and function. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef]
- Staiger, D. Shaping the Arabidopsis Transcriptome through Alternative Splicing. Adv. Bot. 2015, 2015, 13. [Google Scholar] [CrossRef]
- Staiger, D.; Green, R. RNA-based regulation in the plant circadian clock. Trends Plant Sci. 2011, 16, 517–523. [Google Scholar] [CrossRef]
- Perez-Santangelo, S.; Mancini, E.; Francey, L.J.; Schlaen, R.G.; Chernomoretz, A.; Hogenesch, J.B.; Yanovsky, M.J. Role for LSM genes in the regulation of circadian rhythms. Proc. Natl. Acad. Sci. USA 2014, 111, 15166–15171. [Google Scholar] [CrossRef]
- Hong, S.; Song, H.R.; Lutz, K.; Kerstetter, R.A.; Michael, T.P.; McClung, C.R. Type II protein arginine methyltransferase 5 (PRMT5) is required for circadian period determination in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 21211–21216. [Google Scholar] [CrossRef]
- Sanchez, S.E.; Petrillo, E.; Beckwith, E.J.; Zhang, X.; Rugnone, M.L.; Hernando, C.E.; Cuevas, J.C.; Godoy Herz, M.A.; Depetris-Chauvin, A.; Simpson, C.G.; et al. A methyl transferase links the circadian clock to the regulation of alternative splicing. Nature 2010, 468, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Hernando, C.E.; Sanchez, S.E.; Mancini, E.; Yanovsky, M.J. Genome wide comparative analysis of the effects of PRMT5 and PRMT4/CARM1 arginine methyltransferases on the Arabidopsis thaliana transcriptome. BMC Genom. 2015, 16, 192. [Google Scholar] [CrossRef]
- Deng, X.; Gu, L.; Liu, C.; Lu, T.; Lu, F.; Lu, Z.; Cui, P.; Pei, Y.; Wang, B.; Hu, S.; et al. Arginine methylation mediated by the Arabidopsis homolog of PRMT5 is essential for proper pre-mRNA splicing. Proc. Natl. Acad. Sci. USA 2010, 107, 19114–19119. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Niu, L.; Lu, F.; Liu, C.; Zhai, J.; Kong, X.; Cao, X. Mutations in the Type II protein arginine methyltransferase AtPRMT5 result in pleiotropic developmental defects in Arabidopsis. Plant Physiol. 2007, 144, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Friesen, W.J.; Paushkin, S.; Wyce, A.; Massenet, S.; Pesiridis, G.S.; Van Duyne, G.; Rappsilber, J.; Mann, M.; Dreyfuss, G. The methylosome, a 20S complex containing JBP1 and pICln, produces dimethylarginine-modified Sm proteins. Mol. Cell. Biol. 2001, 21, 8289–8300. [Google Scholar] [CrossRef]
- Brahms, H.; Meheus, L.; de Brabandere, V.; Fischer, U.; Luhrmann, R. Symmetrical dimethylation of arginine residues in spliceosomal Sm protein B/B’ and the Sm-like protein LSm4, and their interaction with the SMN protein. RNA 2001, 7, 1531–1542. [Google Scholar] [CrossRef]
- Wang, X.; Wu, F.; Xie, Q.; Wang, H.; Wang, Y.; Yue, Y.; Gahura, O.; Ma, S.; Liu, L.; Cao, Y.; et al. SKIP is a component of the spliceosome linking alternative splicing and the circadian clock in Arabidopsis. Plant Cell 2012, 24, 3278–3295. [Google Scholar] [CrossRef]
- Jones, M.A.; Williams, B.A.; McNicol, J.; Simpson, C.G.; Brown, J.W.; Harmer, S.L. Mutation of Arabidopsis SPLICEOSOMAL TIMEKEEPER LOCUS1 causes circadian clock defects. Plant Cell 2012, 24, 4066–4082. [Google Scholar] [CrossRef]
- Schlaen, R.G.; Mancini, E.; Sanchez, S.E.; Perez-Santangelo, S.; Rugnone, M.L.; Simpson, C.G.; Brown, J.W.; Zhang, X.; Chernomoretz, A.; Yanovsky, M.J. The spliceosome assembly factor GEMIN2 attenuates the effects of temperature on alternative splicing and circadian rhythms. Proc. Natl. Acad. Sci. USA 2015, 112, 9382–9387. [Google Scholar] [CrossRef]
- Schoning, J.C.; Streitner, C.; Meyer, I.M.; Gao, Y.; Staiger, D. Reciprocal regulation of glycine-rich RNA-binding proteins via an interlocked feedback loop coupling alternative splicing to nonsense-mediated decay in Arabidopsis. Nucl. Acids Res. 2008, 36, 6977–6987. [Google Scholar] [CrossRef]
- Streitner, C.; Hennig, L.; Korneli, C.; Staiger, D. Global transcript profiling of transgenic plants constitutively overexpressing the RNA-binding protein AtGRP7. BMC Plant Biol. 2010, 10, 221. [Google Scholar] [CrossRef] [PubMed]
- James, A.B.; Syed, N.H.; Bordage, S.; Marshall, J.; Nimmo, G.A.; Jenkins, G.I.; Herzyk, P.; Brown, J.W.; Nimmo, H.G. Alternative splicing mediates responses of the Arabidopsis circadian clock to temperature changes. Plant Cell 2012, 24, 961–981. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.J.; Park, M.J.; Kim, S.G.; Baldwin, I.T.; Park, C.M. Alternative splicing and nonsense-mediated decay of circadian clock genes under environmental stress conditions in Arabidopsis. BMC Plant Biol. 2014, 14, 136. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, D.R.; Gould, P.; Foreman, J.; Griffiths, J.; Bird, S.; Page, R.; Stewart, K.; Steel, G.; Young, J.; Paszkiewicz, K.; et al. HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENES1 is required for circadian periodicity through the promotion of nucleo-cytoplasmic mRNA export in Arabidopsis. Plant Cell 2013, 25, 4391–4404. [Google Scholar] [CrossRef]
- Lummer, M.; Humpert, F.; Steuwe, C.; Caesar, K.; Schuttpelz, M.; Sauer, M.; Staiger, D. Reversible photoswitchable DRONPA-s monitors nucleocytoplasmic transport of an RNA-binding protein in transgenic plants. Traffic 2011, 12, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Lummer, M.; Humpert, F.; Wiedenlubbert, M.; Sauer, M.; Schuttpelz, M.; Staiger, D. A new set of reversibly photoswitchable fluorescent proteins for use in transgenic plants. Mol. Plant 2013, 6, 1518–1530. [Google Scholar] [CrossRef]
- Kim, J.Y.; Song, H.R.; Taylor, B.L.; Carre, I.A. Light-regulated translation mediates gated induction of the Arabidopsis clock protein LHY. EMBO J. 2003, 22, 935–944. [Google Scholar] [CrossRef]
- Juntawong, P.; Bailey-Serres, J. Dynamic Light Regulation of Translation Status in Arabidopsis thaliana. Front. Plant Sci. 2012, 3, 66. [Google Scholar] [CrossRef]
- Hazen, S.P.; Naef, F.; Quisel, T.; Gendron, J.M.; Chen, H.; Ecker, J.R.; Borevitz, J.O.; Kay, S.A. Exploring the transcriptional landscape of plant circadian rhythms using genome tiling arrays. Genome Biol. 2009, 10, R17. [Google Scholar] [CrossRef] [PubMed]
- Henriques, R.; Wang, H.; Liu, J.; Boix, M.; Huang, L.F.; Chua, N.H. The antiphasic regulatory module comprising CDF5 and its antisense RNA FLORE links the circadian clock to photoperiodic flowering. New Phytol. 2017, 216, 854–867. [Google Scholar] [CrossRef]
- Jung, J.H.; Seo, Y.H.; Seo, P.J.; Reyes, J.L.; Yun, J.; Chua, N.H.; Park, C.M. The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis. Plant Cell 2007, 19, 2736–2748. [Google Scholar] [CrossRef] [PubMed]
- Sire, C.; Moreno, A.B.; Garcia-Chapa, M.; Lopez-Moya, J.J.; San Segundo, B. Diurnal oscillation in the accumulation of Arabidopsis microRNAs, miR167, miR168, miR171 and miR398. FEBS Lett. 2009, 583, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Lu, F.; Pei, Y.; Liu, C.; Cao, X. Regulation of flowering time by the protein arginine methyltransferase AtPRMT10. EMBO Rep. 2007, 8, 1190–1195. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Y.; Ma, Q.; Zhang, Z.; Xue, Y.; Bao, S.; Chong, K. SKB1-mediated symmetric dimethylation of histone H4R3 controls flowering time in Arabidopsis. EMBO J. 2007, 26, 1934–1941. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Gong, Z.; Rock, C.D.; Subramanian, S.; Guo, Y.; Xu, W.; Galbraith, D.; Zhu, J.K. Modulation of abscisic acid signal transduction and biosynthesis by an Sm-like protein in Arabidopsis. Dev. Cell 2001, 1, 771–781. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, S.; Zhang, Y.; Wang, X.; Li, D.; Li, Q.; Yue, M.; Li, Q.; Zhang, Y.E.; Xu, Y.; et al. Arabidopsis floral initiator SKB1 confers high salt tolerance by regulating transcription and pre-mRNA splicing through altering histone H4R3 and small nuclear ribonucleoprotein LSM4 methylation. Plant Cell 2011, 23, 396–411. [Google Scholar] [CrossRef]
- Filichkin, S.A.; Priest, H.D.; Givan, S.A.; Shen, R.; Bryant, D.W.; Fox, S.E.; Wong, W.K.; Mockler, T.C. Genome-wide mapping of alternative splicing in Arabidopsis thaliana. Genome Res. 2010, 20, 45–58. [Google Scholar] [CrossRef]
- Staiger, D.; Brown, J.W. Alternative splicing at the intersection of biological timing, development, and stress responses. Plant Cell 2013, 25, 3640–3656. [Google Scholar] [CrossRef]
- Covington, M.F.; Panda, S.; Liu, X.L.; Strayer, C.A.; Wagner, D.R.; Kay, S.A. ELF3 modulates resetting of the circadian clock in Arabidopsis. Plant Cell 2001, 13, 1305–1315. [Google Scholar] [CrossRef]
- Yanovsky, M.J.; Mazzella, M.A.; Whitelam, G.C.; Casal, J.J. Resetting of the circadian clock by phytochromes and cryptochromes in Arabidopsis. J. Biol. Rhythm. 2001, 16, 523–530. [Google Scholar] [CrossRef]
- Fankhauser, C.; Staiger, D. Photoreceptors in Arabidopsis thaliana: Light perception, signal transduction and entrainment of the endogenous clock. Planta 2002, 216, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Seo, P.J.; Park, M.J.; Lim, M.H.; Kim, S.G.; Lee, M.; Baldwin, I.T.; Park, C.M. A self-regulatory circuit of CIRCADIAN CLOCK-ASSOCIATED1 underlies the circadian clock regulation of temperature responses in Arabidopsis. Plant Cell 2012, 24, 2427–2442. [Google Scholar] [CrossRef] [PubMed]
- Isken, O.; Maquat, L.E. Quality control of eukaryotic mRNA: Safeguarding cells from abnormal mRNA function. Genes Dev. 2007, 21, 1833–1856. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.F.; Imam, J.S.; Wilkinson, M.F. The nonsense-mediated decay RNA surveillance pathway. Annu. Rev. Biochem. 2007, 76, 51–74. [Google Scholar] [CrossRef] [PubMed]
- Shyu, A.B.; Wilkinson, M.F.; van Hoof, A. Messenger RNA regulation: To translate or to degrade. EMBO J. 2008, 27, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Kerenyi, Z.; Merai, Z.; Hiripi, L.; Benkovics, A.; Gyula, P.; Lacomme, C.; Barta, E.; Nagy, F.; Silhavy, D. Inter-kingdom conservation of mechanism of nonsense-mediated mRNA decay. EMBO J. 2008, 27, 1585–1595. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Watanabe, Y. Context analysis of termination codons in mRNA that are recognized by plant NMD. Plant Cell Physiol. 2007, 48, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kang, J.H.; Hettenhausen, C.; Baldwin, I.T. Nonsense-mediated mRNA decay (NMD) silences the accumulation of aberrant trypsin proteinase inhibitor mRNA in Nicotiana attenuata. Plant J. Cell Mol. Biol. 2007, 51, 693–706. [Google Scholar] [CrossRef]
- Schwartz, A.M.; Komarova, T.V.; Skulachev, M.V.; Zvereva, A.S.; Dorokhov Iu, L.; Atabekov, J.G. Stability of plant mRNAs depends on the length of the 3’-untranslated region. Biochemistry (Mosc.) 2006, 71, 1377–1384. [Google Scholar] [CrossRef]
- Kertesz, S.; Kerenyi, Z.; Merai, Z.; Bartos, I.; Palfy, T.; Barta, E.; Silhavy, D. Both introns and long 3’-UTRs operate as cis-acting elements to trigger nonsense-mediated decay in plants. Nucl. Acids Res. 2006, 34, 6147–6157. [Google Scholar] [CrossRef]
- Arciga-Reyes, L.; Wootton, L.; Kieffer, M.; Davies, B. UPF1 is required for nonsense-mediated mRNA decay (NMD) and RNAi in Arabidopsis. Plant J. Cell Mol. Biol. 2006, 47, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Watanabe, Y. UPF3 suppresses aberrant spliced mRNA in Arabidopsis. Plant J. Cell Mol. Biol. 2005, 43, 530–540. [Google Scholar] [CrossRef]
- Riehs, N.; Akimcheva, S.; Puizina, J.; Bulankova, P.; Idol, R.A.; Siroky, J.; Schleiffer, A.; Schweizer, D.; Shippen, D.E.; Riha, K. Arabidopsis SMG7 protein is required for exit from meiosis. J. Cell Sci. 2008, 121, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Lidder, P.; Gutierrez, R.A.; Salome, P.A.; McClung, C.R.; Green, P.J. Circadian control of messenger RNA stability. Association with a sequence-specific messenger RNA decay pathway. Plant Physiol. 2005, 138, 2374–2385. [Google Scholar] [CrossRef] [PubMed]
- Morgan, L.W.; Feldman, J.F. Isolation and characterization of a temperature-sensitive circadian clock mutant of Neurospora crassa. Genetics 1997, 146, 525–530. [Google Scholar]
- Morgan, L.W.; Feldman, J.F. Epistatic and synergistic interactions between circadian clock mutations in Neurospora crassa. Genetics 2001, 159, 537–543. [Google Scholar]
- Adhvaryu, K.; Firoozi, G.; Motavaze, K.; Lakin-Thomas, P. PRD-1, a Component of the Circadian System of Neurospora crassa, Is a Member of the DEAD-box RNA Helicase Family. J. Biol. Rhythm. 2016, 31, 258–271. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Y.; Sun, Y.; Yu, J.; Wang, P.; Ma, H.; Chen, S.; Ma, L.; Zhang, D.; He, Q.; et al. Up-Fr.ameshift Protein UPF1 Regulates Neurospora crassa Circadian and Diurnal Growth Rhythms. Genetics 2017, 206, 1881–1893. [Google Scholar] [CrossRef]
- Rayson, S.; Ashworth, M.; de Torres Zabala, M.; Grant, M.; Davies, B. The salicylic acid dependent and independent effects of NMD in plants. Plant Signal. Behav. 2012, 7, 1434–1437. [Google Scholar] [CrossRef]
- Riehs-Kearnan, N.; Gloggnitzer, J.; Dekrout, B.; Jonak, C.; Riha, K. Aberrant growth and lethality of Arabidopsis deficient in nonsense-mediated RNA decay factors is caused by autoimmune-like response. Nucl. Acids Res. 2012, 40, 5615–5624. [Google Scholar] [CrossRef]
- Ni, J.Z.; Grate, L.; Donohue, J.P.; Preston, C.; Nobida, N.; O’Brien, G.; Shiue, L.; Clark, T.A.; Blume, J.E.; Ares, M., Jr. Ultraconserved elements are associated with homeostatic control of splicing regulators by alternative splicing and nonsense-mediated decay. Genes Dev. 2007, 21, 708–718. [Google Scholar] [CrossRef]
- Lareau, L.F.; Inada, M.; Green, R.E.; Wengrod, J.C.; Brenner, S.E. Unproductive splicing of SR genes associated with highly conserved and ultraconserved DNA elements. Nature 2007, 446, 926–929. [Google Scholar] [CrossRef]
- Kalyna, M.; Simpson, C.G.; Syed, N.H.; Lewandowska, D.; Marquez, Y.; Kusenda, B.; Marshall, J.; Fuller, J.; Cardle, L.; McNicol, J.; et al. Alternative splicing and nonsense-mediated decay modulate expression of important regulatory genes in Arabidopsis. Nucl. Acids Res. 2012, 40, 2454–2469. [Google Scholar] [CrossRef]
- Carpenter, C.D.; Kreps, J.A.; Simon, A.E. Genes encoding glycine-rich Arabidopsis thaliana proteins with RNA-binding motifs are influenced by cold treatment and an endogenous circadian rhythm. Plant Physiol. 1994, 104, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Staiger, D.; Zecca, L.; Wieczorek Kirk, D.A.; Apel, K.; Eckstein, L. The circadian clock regulated RNA-binding protein AtGRP7 autoregulates its expression by influencing alternative splicing of its own pre-mRNA. Plant J. Cell Mol. Biol. 2003, 33, 361–371. [Google Scholar] [CrossRef]
- Meyer, K.; Koster, T.; Nolte, C.; Weinholdt, C.; Lewinski, M.; Grosse, I.; Staiger, D. Adaptation of iCLIP to plants determines the binding landscape of the clock-regulated RNA-binding protein AtGRP7. Genome Biol. 2017, 18, 204. [Google Scholar] [CrossRef] [PubMed]
- Schmal, C.; Reimann, P.; Staiger, D. A circadian clock-regulated toggle switch explains AtGRP7 and AtGRP8 oscillations in Arabidopsis thaliana. PLoS Comput. Biol. 2013, 9, e1002986. [Google Scholar] [CrossRef]
- Alabadi, D.; Yanovsky, M.J.; Mas, P.; Harmer, S.L.; Kay, S.A. Critical role for CCA1 and LHY in maintaining circadian rhythmicity in Arabidopsis. Curr. Biol. 2002, 12, 757–761. [Google Scholar] [CrossRef]
- Perales, M.; Mas, P. A functional link between rhythmic changes in chromatin structure and the Arabidopsis biological clock. Plant Cell 2007, 19, 2111–2123. [Google Scholar] [CrossRef] [PubMed]
- Gould, P.D.; Locke, J.C.; Larue, C.; Southern, M.M.; Davis, S.J.; Hanano, S.; Moyle, R.; Milich, R.; Putterill, J.; Millar, A.J.; et al. The molecular basis of temperature compensation in the Arabidopsis circadian clock. Plant Cell 2006, 18, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Raices, M.; D’Angelo, M.A. Nuclear pore complex composition: A new regulator of tissue-specific and developmental functions. Nat. Rev. Mol. Cell Biol. 2012, 13, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Fukao, Y.; Iwamoto, M.; Haraguchi, T.; Hara-Nishimura, I. Identification and characterization of nuclear pore complex components in Arabidopsis thaliana. Plant Cell 2010, 22, 4084–4097. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X. A putative nucleoporin 96 is required for both basal defense and constitutive resistance responses mediated by suppressor of npr1-1,constitutive 1. Plant Cell 2005, 17, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.T.; Germain, H.; Wiermer, M.; Bi, D.; Xu, F.; Garcia, A.V.; Wirthmueller, L.; Despres, C.; Parker, J.E.; Zhang, Y.; et al. Nuclear pore complex component MOS7/Nup88 is required for innate immunity and nuclear accumulation of defense regulators in Arabidopsis. Plant Cell 2009, 21, 2503–2516. [Google Scholar] [CrossRef] [PubMed]
- Wiermer, M.; Cheng, Y.T.; Imkampe, J.; Li, M.; Wang, D.; Lipka, V.; Li, X. Putative members of the Arabidopsis Nup107-160 nuclear pore sub-complex contribute to pathogen defense. Plant J. Cell Mol. Biol. 2012, 70, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, N.; Madsen, L.H.; Radutoiu, S.; Frantescu, M.; Quistgaard, E.M.; Miwa, H.; Downie, J.A.; James, E.K.; Felle, H.H.; Haaning, L.L.; et al. A nucleoporin is required for induction of Ca2+ spiking in legume nodule development and essential for rhizobial and fungal symbiosis. Proc. Natl. Acad. Sci. USA 2006, 103, 359–364. [Google Scholar] [CrossRef]
- Saito, K.; Yoshikawa, M.; Yano, K.; Miwa, H.; Uchida, H.; Asamizu, E.; Sato, S.; Tabata, S.; Imaizumi-Anraku, H.; Umehara, Y.; et al. NUCLEOPORIN85 is required for calcium spiking, fungal and bacterial symbioses, and seed production in Lotus japonicus. Plant Cell 2007, 19, 610–624. [Google Scholar] [CrossRef]
- Groth, M.; Takeda, N.; Perry, J.; Uchida, H.; Draxl, S.; Brachmann, A.; Sato, S.; Tabata, S.; Kawaguchi, M.; Wang, T.L.; et al. NENA, a Lotus japonicus homolog of Sec13, is required for rhizodermal infection by arbuscular mycorrhiza fungi and rhizobia but dispensable for cortical endosymbiotic development. Plant Cell 2010, 22, 2509–2526. [Google Scholar] [CrossRef]
- Parry, G.; Ward, S.; Cernac, A.; Dharmasiri, S.; Estelle, M. The Arabidopsis SUPPRESSOR OF AUXIN RESISTANCE proteins are nucleoporins with an important role in hormone signaling and development. Plant Cell 2006, 18, 1590–1603. [Google Scholar] [CrossRef]
- Robles, L.M.; Deslauriers, S.D.; Alvarez, A.A.; Larsen, P.B. A loss-of-function mutation in the nucleoporin AtNUP160 indicates that normal auxin signalling is required for a proper ethylene response in Arabidopsis. J. Exp. Bot. 2012, 63, 2231–2241. [Google Scholar] [CrossRef]
- Lazaro, A.; Valverde, F.; Pineiro, M.; Jarillo, J.A. The Arabidopsis E3 ubiquitin ligase HOS1 negatively regulates CONSTANS abundance in the photoperiodic control of flowering. Plant Cell 2012, 24, 982–999. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.H.; Agarwal, M.; Zhang, Y.; Xie, Q.; Zhu, J.K. The negative regulator of plant cold responses, HOS1, is a RING E3 ligase that mediates the ubiquitination and degradation of ICE1. Proc. Natl. Acad. Sci. USA 2006, 103, 8281–8286. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.H.; Hu, X.; Tang, W.; Zheng, X.; Kim, Y.S.; Lee, B.H.; Zhu, J.K. A putative Arabidopsis nucleoporin, AtNUP160, is critical for RNA export and required for plant tolerance to cold stress. Mol. Cell. Biol. 2006, 26, 9533–9543. [Google Scholar] [CrossRef] [PubMed]
- Tourriere, H.; Chebli, K.; Tazi, J. mRNA degradation machines in eukaryotic cells. Biochimie 2002, 84, 821–837. [Google Scholar] [CrossRef]
- Bologna, N.G.; Iselin, R.; Abriata, L.A.; Sarazin, A.; Pumplin, N.; Jay, F.; Grentzinger, T.; Dal Peraro, M.; Voinnet, O. Nucleo-cytosolic Shuttling of ARGONAUTE1 Prompts a Revised Model of the Plant MicroRNA Pathway. Mol. Cell 2018, 69, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Izaurralde, E.; Jarmolowski, A.; Beisel, C.; Mattaj, I.W.; Dreyfuss, G.; Fischer, U. A role for the M9 transport signal of hnRNP A1 in mRNA nuclear export. J. Cell Biol. 1997, 137, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Ziemienowicz, A.; Haasen, D.; Staiger, D.; Merkle, T. Arabidopsis transportin1 is the nuclear import receptor for the circadian clock-regulated RNA-binding protein AtGRP7. Plant Mol. Biol. 2003, 53, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Hornyik, C.; Duc, C.; Rataj, K.; Terzi, L.C.; Simpson, G.G. Alternative polyadenylation of antisense RNAs and flowering time control. Biochem. Soc. Trans. 2010, 38, 1077–1081. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Manley, J.L. Alternative polyadenylation of mRNA precursors. Nat. Rev. Mol. Cell Biol. 2017, 18, 18–30. [Google Scholar] [CrossRef]
- Robinson, B.G.; Frim, D.M.; Schwartz, W.J.; Majzoub, J.A. Vasopressin mRNA in the suprachiasmatic nuclei: Daily regulation of polyadenylate tail length. Science 1988, 241, 342–344. [Google Scholar] [CrossRef]
- Kojima, S.; Sher-Chen, E.L.; Green, C.B. Circadian control of mRNA polyadenylation dynamics regulates rhythmic protein expression. Genes Dev. 2012, 26, 2724–2736. [Google Scholar] [CrossRef] [PubMed]
- Addepalli, B.; Meeks, L.R.; Forbes, K.P.; Hunt, A.G. Novel alternative splicing of mRNAs encoding poly(A) polymerases in Arabidopsis. Biochim. Biophys. Acta 2004, 1679, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Meeks, L.R.; Addepalli, B.; Hunt, A.G. Characterization of genes encoding poly(A) polymerases in plants: Evidence for duplication and functional specialization. PLoS ONE 2009, 4, e8082. [Google Scholar] [CrossRef] [PubMed]
- Belostotsky, D.A.; Sieburth, L.E. Kill the messenger: MRNA decay and plant development. Curr. Opin. Plant Biol. 2009, 12, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Arae, T.; Green, P.J.; Yamaguchi, J.; Chiba, Y. AtCCR4a and AtCCR4b are Involved in Determining the Poly(A) Length of Granule-bound starch synthase 1 Transcript and Modulating Sucrose and Starch Metabolism in Arabidopsis thaliana. Plant Cell Physiol. 2015, 56, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.J.; Wu, S.H.; Chen, H.M.; Wu, S.H. Widespread translational control contributes to the regulation of Arabidopsis photomorphogenesis. Mol. Syst. Biol. 2012, 8, 566. [Google Scholar] [CrossRef] [PubMed]
- Missra, A.; von Arnim, A.G. Analysis of mRNA translation states in Arabidopsis over the diurnal cycle by polysome microarray. Methods Mol. Biol. 2014, 1158, 157–174. [Google Scholar] [CrossRef]
- Kim, W.Y.; Fujiwara, S.; Suh, S.S.; Kim, J.; Kim, Y.; Han, L.; David, K.; Putterill, J.; Nam, H.G.; Somers, D.E. ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light. Nature 2007, 449, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.K.; Nomura, Y.; Shi, H.; Nakagami, H.; Somers, D.E. Circadian Profiling of the Arabidopsis Proteome Using 2D-DIGE. Front. Plant Sci. 2016, 7, 1007. [Google Scholar] [CrossRef]
- Robles, M.S.; Cox, J.; Mann, M. In-vivo quantitative proteomics reveals a key contribution of post-transcriptional mechanisms to the circadian regulation of liver metabolism. PLoS Genet. 2014, 10, e1004047. [Google Scholar] [CrossRef]
- Reddy, A.B.; Karp, N.A.; Maywood, E.S.; Sage, E.A.; Deery, M.; O’Neill, J.S.; Wong, G.K.; Chesham, J.; Odell, M.; Lilley, K.S.; et al. Circadian orchestration of the hepatic proteome. Curr. Biol. 2006, 16, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Missra, A.; Ernest, B.; Lohoff, T.; Jia, Q.; Satterlee, J.; Ke, K.; von Arnim, A.G. The Circadian Clock Modulates Global Daily Cycles of mRNA Ribosome Loading. Plant Cell 2015, 27, 2582–2599. [Google Scholar] [CrossRef] [PubMed]
- Graf, A.; Coman, D.; Uhrig, R.G.; Walsh, S.; Flis, A.; Stitt, M.; Gruissem, W. Parallel analysis of Arabidopsis circadian clock mutants reveals different scales of transcriptome and proteome regulation. Open Biol. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Kramer, C.; Loros, J.J.; Dunlap, J.C.; Crosthwaite, S.K. Role for antisense RNA in regulating circadian clock function in Neurospora crassa. Nature 2003, 421, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, H.J.; Vu, Q.T.; Jung, S.; McClung, C.R.; Hong, S.; Nam, H.G. Circadian control of ORE1 by PRR9 positively regulates leaf senescence in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, 8448–8453. [Google Scholar] [CrossRef] [PubMed]
- Yanovsky, M.J.; Kay, S.A. Molecular basis of seasonal time measurement in Arabidopsis. Nature 2002, 419, 308–312. [Google Scholar] [CrossRef]
- Yu, Y.; Jia, T.; Chen, X. The ‘how’ and ‘where’ of plant microRNAs. New Phytol. 2017, 216, 1002–1017. [Google Scholar] [CrossRef]
- Kadener, S.; Menet, J.S.; Sugino, K.; Horwich, M.D.; Weissbein, U.; Nawathean, P.; Vagin, V.V.; Zamore, P.D.; Nelson, S.B.; Rosbash, M. A role for microRNAs in the Drosophila circadian clock. Genes Dev. 2009, 23, 2179–2191. [Google Scholar] [CrossRef]
- Koster, T.; Meyer, K.; Weinholdt, C.; Smith, L.M.; Lummer, M.; Speth, C.; Grosse, I.; Weigel, D.; Staiger, D. Regulation of pri-miRNA processing by the hnRNP-like protein AtGRP7 in Arabidopsis. Nucl. Acids Res. 2014, 42, 9925–9936. [Google Scholar] [CrossRef]
- Seo, P.J.; Mas, P. Multiple layers of posttranslational regulation refine circadian clock activity in Arabidopsis. Plant Cell 2014, 26, 79–87. [Google Scholar] [CrossRef]
- Koster, T.; Meyer, K. Plant Ribonomics: Proteins in Search of RNA Partners. Trends Plant Sci. 2018, 23, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Xing, D.; Wang, Y.; Hamilton, M.; Ben-Hur, A.; Reddy, A.S. Transcriptome-Wide Identification of RNA Targets of Arabidopsis SERINE/ARGININE-RICH45 Uncovers the Unexpected Roles of This RNA Binding Protein in RNA Processing. Plant Cell 2015, 27, 3294–3308. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, L.; Hou, Y.; Wang, L.; Deng, X.; Hang, R.; Chen, D.; Zhang, X.; Zhang, Y.; Liu, C.; et al. Integrative genome-wide analysis reveals HLP1, a novel RNA-binding protein, regulates plant flowering by targeting alternative polyadenylation. Cell Res. 2015, 25, 864–876. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.I.A.I. Taking the next step: Building an Arabidopsis information portal. Plant Cell 2012, 24, 2248–2256. [Google Scholar] [CrossRef]
- McHugh, C.A.; Chen, C.K.; Chow, A.; Surka, C.F.; Tran, C.; McDonel, P.; Pandya-Jones, A.; Blanco, M.; Burghard, C.; Moradian, A.; et al. The Xist lncRNA interacts directly with SHARP to silence transcription through HDAC3. Nature 2015, 521, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Rogell, B.; Fischer, B.; Rettel, M.; Krijgsveld, J.; Castello, A.; Hentze, M.W. Specific RNP capture with antisense LNA/DNA mixmers. RNA 2017, 23, 1290–1302. [Google Scholar] [CrossRef]
- Greenham, K.; Lou, P.; Remsen, S.E.; Farid, H.; McClung, C.R. TRiP: Tracking Rhythms in Plants, an automated leaf movement analysis program for circadian period estimation. Plant Methods 2015, 11, 33. [Google Scholar] [CrossRef]
- Wagner, L.; Schmal, C.; Staiger, D.; Danisman, S. The plant leaf movement analyzer (PALMA): A simple tool for the analysis of periodic cotyledon and leaf movement in Arabidopsis thaliana. Plant Methods 2017, 13, 2. [Google Scholar] [CrossRef]
- Dornbusch, T.; Michaud, O.; Xenarios, I.; Fankhauser, C. Differentially phased leaf growth and movements in Arabidopsis depend on coordinated circadian and light regulation. Plant Cell 2014, 26, 3911–3921. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Mechanism | Gene | Function | Clock-Related Observations | References |
|---|---|---|---|---|
| Alternative splicing | PRMT5 | Type II protein arginine methyltransferase of histones, Sm and LSM spliceosomal proteins. | Circadian expression. prmt5: Long period; elevated aberrant PRR9 isoform. | [31,32] |
| SM-like genes | Components of the U6 snRNP. | Circadian expression of LSM5. lsm4and lsm5: Long period; changes in expression and alternative splicing of some core clock genes. | [30] | |
| SKIP | Splicing factor and component of the spliceosome. | Role in mediating light input and maintaining temperature compensation of the clock. skip: Temperature-sensitive long period; increased aberrantly spliced variants of PRR7 and PRR9. | [38] | |
| STIPL1 | RNA-binding protein. | stipl1: Long period. Altered accumulation and alternative splicing of core clock transcripts. | [39] | |
| GEMIN2 | Spliceosomal assembly factor. Attenuates the effects of low temperature on alternative splicing. | Modulates temperature effects on circadian clock. gemin2: Early flowering and short-period phenotype. Alterations in the alternative splicing of core clock genes, similarly affected in wild-type plants at low temperature conditions. Disruption of temperature compensation: Significant period lengthening in response to cold conditions. | [40] | |
| AS-NMD | AtGRP7 AtGRP8 | RNA-binding protein. RNA-binding protein. | AtGRP7 and AtGRP8 are rhythmic transcripts. AtGRP7 binding to its own pre-mRNA causes AS followed by NMD, thus autoregulating its expression. | [41,42] |
| TOC1 * ELF3 * | Core clock gene. Core clock gene. | AS results in PTC and decay. toc1: Short period. elf3: Arrythmic. | [43,44] | |
| Nuclear transport | HOS1 | E3 ubiquitin ligase activity. Attenuates cold responses, prevents precocious flowering, and regulates mRNA export. | hos1: Early flowering and long-period phenotype. Upregulation of circadian-expressed and core clock genes. Reduction in rhythmicity of circadian clock genes. | [45] |
| AtGRP7 | RNA-binding protein. | AtGRP7 shuttles between the nucleus and the cytoplasm. | [46,47] | |
| Polyadenylation | Poly(A)-polymerases and deadenylases | Exhibit circadian expression. | [24] | |
| Regulation of translation | LHY * | Core clock gene. | Translation is promoted by light, whereas transcription repressed at dusk. Both mechanisms contribute to the narrow peak of LHY at dusk. Lhy: Short period. | [48] |
| Light induces translation. | [49] | |||
| nc-RNAs | NATs to clock genes | Natural antisense RNAs to LHY, CCA1, TOC1, PRR3, PRR5, PRR7, and PRR9 exhibit circadian rhythmicity. | [50] | |
| FLORE | CDF5 long-noncoding RNA. Promotes flowering by repressing CDFs. | Antiphasic circadian expression. Reciprocal inhibition contributes to the proper circadian oscillation of both transcripts. Target of PRR7. OxFLORE: Early flowering. | [51] | |
| miR172 | miRNA controlling AP-like transcription factors. Regulates flowering time. | Its biogenesis is controlled by GI. | [52] | |
| MIR157A, MIR158A, MIR160A, MIR167D | miRNA controlling diverse developmental processes in Arabidopsis. | Transcripts show circadian expression. | [50,53] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateos, J.L.; De Leone, M.J.; Torchio, J.; Reichel, M.; Staiger, D. Beyond Transcription: Fine-Tuning of Circadian Timekeeping by Post-Transcriptional Regulation. Genes 2018, 9, 616. https://doi.org/10.3390/genes9120616
Mateos JL, De Leone MJ, Torchio J, Reichel M, Staiger D. Beyond Transcription: Fine-Tuning of Circadian Timekeeping by Post-Transcriptional Regulation. Genes. 2018; 9(12):616. https://doi.org/10.3390/genes9120616
Chicago/Turabian StyleMateos, Julieta Lisa, Maria José De Leone, Jeanette Torchio, Marlene Reichel, and Dorothee Staiger. 2018. "Beyond Transcription: Fine-Tuning of Circadian Timekeeping by Post-Transcriptional Regulation" Genes 9, no. 12: 616. https://doi.org/10.3390/genes9120616
APA StyleMateos, J. L., De Leone, M. J., Torchio, J., Reichel, M., & Staiger, D. (2018). Beyond Transcription: Fine-Tuning of Circadian Timekeeping by Post-Transcriptional Regulation. Genes, 9(12), 616. https://doi.org/10.3390/genes9120616