
Association of Cognitive Polygenic Index and Cognitive Performance with Age in Cognitively Healthy Adults

 ,
,
Abstract
1. Introduction
2. Methods
Participants
3. Cognitive Tasks
4. PGI Calculation
5. Brain Measures
6. Statistical Analysis
7. Results
8. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haworth, C.M.; Wright, M.J.; Luciano, M.; Martin, N.G.; de Geus, E.J.; van Beijsterveldt, C.E.; Bartels, M.; Posthuma, D.; Boomsma, D.I.; Davis, O.S.; et al. The heritability of general cognitive ability increases linearly from childhood to young adulthood. Mol. Psychiatry 2010, 15, 1112–1120. [Google Scholar] [CrossRef]
- Fricke-Galindo, I.; Pérez-Aldana, B.E.; Macías-Kauffer, L.R.; González-Arredondo, S.; Dávila-Ortiz de Montellano, D.; Aviña-Cervantes, C.L.; López-López, M.; Rodríguez-Agudelo, Y.; Monroy-Jaramillo, N. Impact of COMT, PRODH and DISC1 Genetic Variants on Cognitive Performance of Patients with Schizophrenia. Arch. Med. Res. 2022, 53, 388–398. [Google Scholar] [CrossRef]
- Mollon, J.; Knowles, E.E.M.; Mathias, S.R.; Gur, R.; Peralta, J.M.; Weiner, D.J.; Robinson, E.B.; Gur, R.E.; Blangero, J.; Almasy, L.; et al. Genetic influence on cognitive development between childhood and adulthood. Mol. Psychiatry 2021, 26, 656–665. [Google Scholar] [CrossRef]
- Pietropaolo, S.; Crusio, W.E. Genes and cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2011, 2, 345–352. [Google Scholar] [CrossRef]
- Riaz, M.; Huq, A.; Ryan, J.; Orchard, S.G.; Tiller, J.; Lockery, J.; Woods, R.L.; Wolfe, R.; Renton, A.E.; Goate, A.M.; et al. Effect of APOE and a polygenic risk score on incident dementia and cognitive decline in a healthy older population. Aging Cell 2021, 20, e13384. [Google Scholar] [CrossRef]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; de Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-wide association studies. Nat. Rev. Methods Prim. 2021, 1, 59. [Google Scholar] [CrossRef]
- Becker, J.; Burik, C.A.P.; Goldman, G.; Wang, N.; Jayashankar, H.; Bennett, M.; Belsky, D.W.; Karlsson Linnér, R.; Ahlskog, R.; Kleinman, A.; et al. Resource profile and user guide of the Polygenic Index Repository. Nat. Hum. Behav. 2021, 5, 1744–1758. [Google Scholar] [CrossRef]
- Tsapanou, A.; Mourtzi, N.; Gu, Y.; Habeck, C.; Belsky, D.; Stern, Y. Polygenic indices for cognition in healthy aging; the role of brain measures. Neuroimage Rep. 2023, 3, 100153. [Google Scholar] [CrossRef]
- Small, S.A.; Stern, Y.; Tang, M.; Mayeux, R. Selective decline in memory function among healthy elderly. Neurology 1999, 52, 1392. [Google Scholar] [CrossRef]
- Shafto, M.A.; Tyler, L.K. Language in the aging brain: The network dynamics of cognitive decline and preservation. Science 2014, 346, 583–587. [Google Scholar] [CrossRef]
- Habeck, C.; Gazes, Y.; Razlighi, Q.; Steffener, J.; Brickman, A.; Barulli, D.; Salthouse, T.; Stern, Y. The Reference Ability Neural Network Study: Life-time stability of reference-ability neural networks derived from task maps of young adults. Neuroimage 2016, 125, 693–704. [Google Scholar] [CrossRef]
- Stern, Y.; Habeck, C.; Steffener, J.; Barulli, D.; Gazes, Y.; Razlighi, Q.; Shaked, D.; Salthouse, T. The Reference Ability Neural Network Study: Motivation, design, and initial feasibility analyses. Neuroimage 2014, 103, 139–151. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, S.; Das, S.; Kretzschmar, W.; Delaneau, O.; Wood, A.R.; Teumer, A.; Kang, H.M.; Fuchsberger, C.; Danecek, P.; Sharp, K.; et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 2016, 48, 1279–1283. [Google Scholar] [CrossRef]
- Choi, S.W.; O’Reilly, P.F. PRSice-2: Polygenic Risk Score software for biobank-scale data. Gigascience 2019, 8, giz082. [Google Scholar] [CrossRef]
- Prive, F.; Vilhjalmsson, B.J.; Aschard, H.; Blum, M.G.B. Making the Most of Clumping and Thresholding for Polygenic Scores. Am. J. Hum. Genet. 2019, 105, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.S.; Segonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 2006, 31, 968–980. [Google Scholar] [CrossRef]
- Fischl, B.; Salat, D.H.; Busa, E.; Albert, M.; Dieterich, M.; Haselgrove, C.; van der Kouwe, A.; Killiany, R.; Kennedy, D.; Klaveness, S.; et al. Whole brain segmentation: Automated labeling of neuroanatomical structures in the human brain. Neuron 2002, 33, 341–355. [Google Scholar] [CrossRef]
- Yoon, Y.B.; Shin, W.G.; Lee, T.Y.; Hur, J.W.; Cho, K.I.K.; Sohn, W.S.; Kim, S.G.; Lee, K.H.; Kwon, J.S. Brain Structural Networks Associated with Intelligence and Visuomotor Ability. Sci. Rep. 2017, 7, 2177. [Google Scholar] [CrossRef]
- Ehrlich, S.; Brauns, S.; Yendiki, A.; Ho, B.C.; Calhoun, V.; Schulz, S.C.; Gollub, R.L.; Sponheim, S.R. Associations of cortical thickness and cognition in patients with schizophrenia and healthy controls. Schizophr. Bull. 2012, 38, 1050–1062. [Google Scholar] [CrossRef]
- Tuladhar, A.M.; Reid, A.T.; Shumskaya, E.; de Laat, K.F.; van Norden, A.G.; van Dijk, E.J.; Norris, D.G.; de Leeuw, F.E. Relationship between white matter hyperintensities, cortical thickness, and cognition. Stroke 2015, 46, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Kloppenborg, R.P.; Nederkoorn, P.J.; Geerlings, M.I.; van den Berg, E. Presence and progression of white matter hyperintensities and cognition: A meta-analysis. Neurology 2014, 82, 2127–2138. [Google Scholar] [CrossRef] [PubMed]
- Fischl, B. FreeSurfer. Neuroimage 2012, 62, 774–781. [Google Scholar] [CrossRef]
- Schmidt, P.; Gaser, C.; Arsic, M.; Buck, D.; Förschler, A.; Berthele, A.; Hoshi, M.; Ilg, R.; Schmid, V.J.; Zimmer, C. An automated tool for detection of FLAIR-hyperintense white-matter lesions in multiple sclerosis. Neuroimage 2012, 59, 3774–3783. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.; Woolrich, M.W.; Smith, S.M. Fsl. Neuroimage 2012, 62, 782–790. [Google Scholar] [CrossRef]
- Moura, A.R.; Lee, S.; Habeck, C.; Razlighi, Q.; Stern, Y. The relationship between white matter hyperintensities and cognitive reference abilities across the life span. Neurobiol. Aging 2019, 83, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Lin, H. Probing Two-way Moderation Effects: A Review of Software to Easily Plot Johnson-Neyman Figures. Struct. Equ. Model. A Multidiscip. J. 2020, 27, 494–502. [Google Scholar] [CrossRef]
- Bauer, D.J.; Curran, P.J. Probing Interactions in Fixed and Multilevel Regression: Inferential and Graphical Techniques. Multivar. Behav. Res. 2005, 40, 373–400. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Ryan, L.H.; Schaie, K.W.; Willis, S.L.; Kolanowski, A. Factors associated with cognition in adults: The Seattle Longitudinal Study. Res. Nurs. Health 2009, 32, 540–550. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yoon, J.A.; Kim, D.Y.; Sohn, M.K.; Lee, J.; Lee, S.G.; Lee, Y.S.; Han, E.Y.; Joo, M.C.; Oh, G.J.; Han, J.; et al. Factors associated with improvement or decline in cognitive function after an ischemic stroke in Korea: The Korean stroke cohort for functioning and rehabilitation (KOSCO) study. BMC Neurol. 2017, 17, 9. [Google Scholar] [CrossRef]
- Walhovd, K.B.; Fjell, A.M.; Espeseth, T. Cognitive decline and brain pathology in aging--need for a dimensional, lifespan and systems vulnerability view. Scand. J. Psychol. 2014, 55, 244–254. [Google Scholar] [CrossRef]
- Pietzuch, M.; King, A.E.; Ward, D.D.; Vickers, J.C. The Influence of Genetic Factors and Cognitive Reserve on Structural and Functional Resting-State Brain Networks in Aging and Alzheimer’s Disease. Front. Aging Neurosci. 2019, 11, 30. [Google Scholar] [CrossRef]
- Savage, J.E.; Jansen, P.R.; Stringer, S.; Watanabe, K.; Bryois, J.; de Leeuw, C.A.; Nagel, M.; Awasthi, S.; Barr, P.B.; Coleman, J.R.I.; et al. Genome-wide association meta-analysis in 269,867 individuals identifies new genetic and functional links to intelligence. Nat. Genet. 2018, 50, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Lupo, G.; Gaetani, S.; Cacci, E.; Biagioni, S.; Negri, R. Molecular Signatures of the Aging Brain: Finding the Links between Genes and Phenotypes. Neurotherapeutics 2019, 16, 543–553. [Google Scholar] [CrossRef]
- Ham, S.; Lee, S.V. Advances in transcriptome analysis of human brain aging. Exp. Mol. Med. 2020, 52, 1787–1797. [Google Scholar] [CrossRef]
- Cross, B.; Turner, R.; Pirmohamed, M. Polygenic risk scores: An overview from bench to bedside for personalised medicine. Front. Genet. 2022, 13, 1000667. [Google Scholar] [CrossRef]
- Gao, Q.; Daunt, P.; Gibson, A.M.; Pither, R.J.; for the Alzheimer’s Disease Neuroimaging Initiative. Utility of Polygenic Risk Scoring to Predict Cognitive Impairment as Measured by Preclinical Alzheimer Cognitive Composite Score. JAR Life 2022, 11, 1–8. [Google Scholar] [CrossRef]


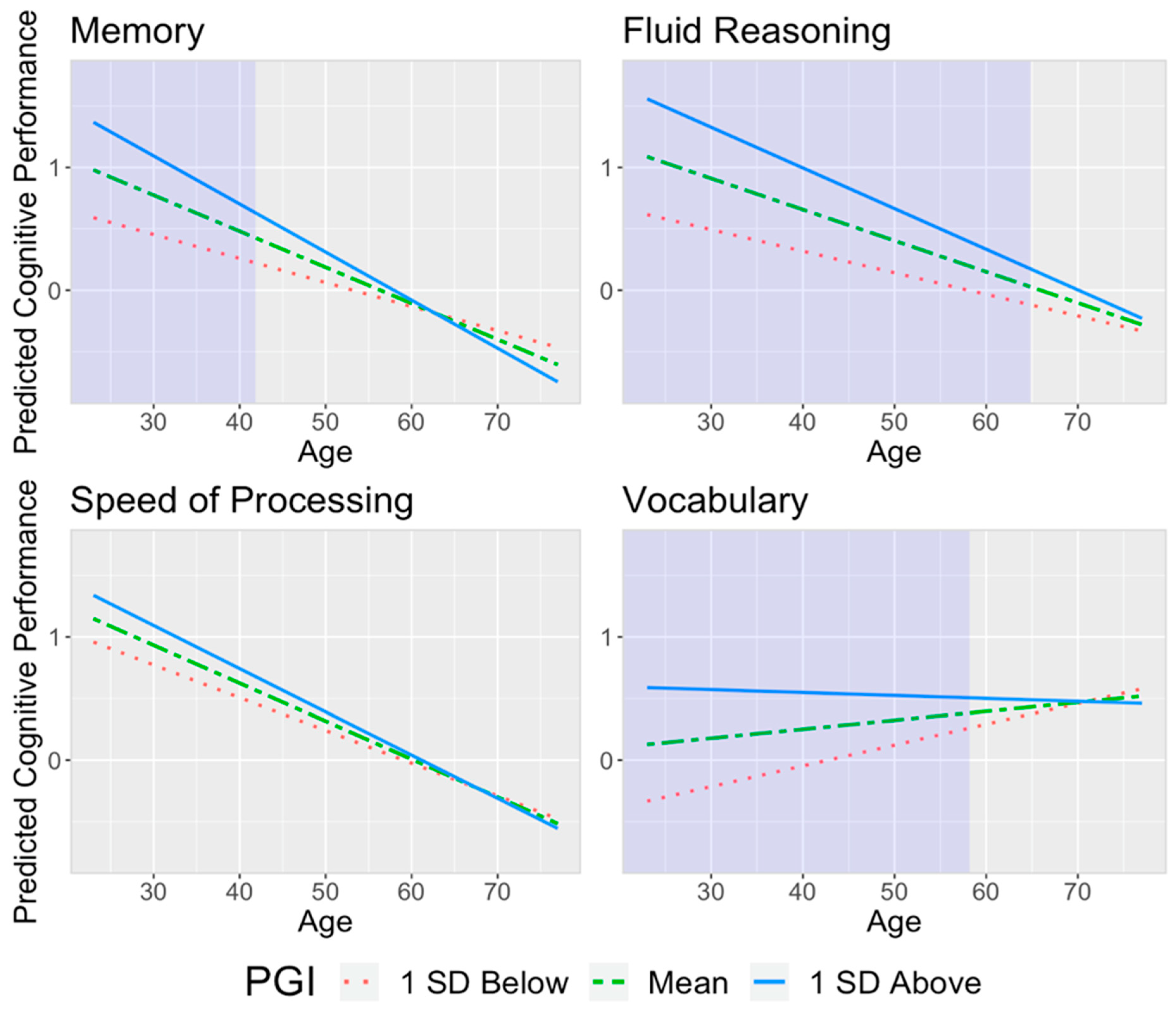{kind=link}
| Characteristics | N | Mean (SD) | Minimum | Maximum |
|---|---|---|---|---|
| Age, years | 168 | 56.90 (15.50) | 23 | 77 |
| Sex, N(%) | 168 | |||
| F | 86 (51.19%) | - | - | |
| M | 82 (48.81%) | - | - | |
| Education, years | 168 | 16.45 (2.27) | 12 | 22 |
| Cognitive Domains (z−scores) | ||||
| Memory | 166 | −0.02 (0.96) | −2.36 | 1.60 |
| Fluid Reasoning | 168 | 0.22 (0.79) | −1.39 | 2.05 |
| Speed of Processing | 168 | 0.10 (0.75) | −1.69 | 2.10 |
| Vocabulary | 165 | 0.36 (0.69) | −2.03 | 1.21 |
| Brain Measures | ||||
| Cortical Thickness | 165 | 2.53 (0.12) | 2.26 | 2.85 |
| Total Gray Matter Volume | 165 | 625,269.41 (59,112.66) | 494,775.94 | 819,078.57 |
| log(White Matter Hyperintensity + 1) | 160 | 4.74 (2.66) | 0 | 9.50 |
| (a) Memory | (b) Fluid Reasoning | |||||
|---|---|---|---|---|---|---|
| Predictors | β | CI | p | β | CI | p |
| Cog PGI | 0.059 | [−0.119, 0.237] | 0.516 | 0.253 | [0.090, 0.415] | 0.003 |
| Age | −0.473 | [−0.608, −0.338] | <0.001 | −0.478 | [−0.601, −0.354] | <0.001 |
| Education | 0.246 | [0.108, 0.384] | 0.001 | 0.142 | [0.017, 0.267] | 0.027 |
| Sex [M] | −0.327 | [−0.593, −0.060] | 0.017 | 0.108 | [−0.138, 0.353] | 0.388 |
| PC 1 | −0.070 | [−0.244, 0.104] | 0.427 | 0.044 | [−0.115, 0.204] | 0.586 |
| Cog PGI x Age | −0.158 | [−0.292, −0.023] | 0.022 | −0.146 | [−0.269, −0.024] | 0.020 |
| (c) Speed of Processing | (d) Vocabulary | |||||
| Predictors | β | CI | p | β | CI | p |
| Cog PGI | 0.061 | [−0.096, 0.219] | 0.444 | 0.173 | [0.017, 0.328] | 0.030 |
| Age | −0.635 | [−0.756, −0.515] | <0.001 | 0.144 | [0.028, 0.260] | 0.015 |
| Education | 0.103 | [−0.019, 0.225] | 0.097 | 0.322 | [0.204, 0.440] | <0.001 |
| Sex [M] | −0.033 | [−0.272, 0.205] | 0.782 | 0.063 | [−0.169, 0.294] | 0.592 |
| PC 1 | 0.110 | [−0.045, 0.266] | 0.161 | 0.029 | [−0.122, 0.181] | 0.701 |
| Cog PGI x Age | −0.088 | [−0.207, 0.031] | 0.148 | −0.191 | [−0.307, −0.076] | 0.001 |
| (a) Memory | (b) Fluid Reasoning | |||||
|---|---|---|---|---|---|---|
| Predictors | β | CI | p | β | CI | p |
| Cog PGI | 0.082 | [−0.105, 0.269] | 0.387 | 0.275 | [0.104, 0.446] | 0.002 |
| Age | −0.476 | [−0.715, −0.236] | <0.001 | −0.503 | [−0.719, −0.287] | <0.001 |
| Education | 0.243 | [0.099, 0.387] | 0.001 | 0.119 | [−0.011, 0.250] | 0.072 |
| Sex [M] | −0.288 | [−0.615, 0.040] | 0.085 | −0.058 | [−0.359, 0.242] | 0.702 |
| PC 1 | −0.096 | [−0.276, 0.083] | 0.291 | 0.023 | [−0.142, 0.188] | 0.784 |
| Cortical Thickness | 0.081 | [−0.120, 0.281] | 0.427 | −0.086 | [−0.268, 0.096] | 0.350 |
| Total Gray Volume | −0.054 | [−0.253, 0.145] | 0.593 | 0.176 | [−0.007, 0.359] | 0.060 |
| log(WMH + 1) | 0.057 | [−0.142, 0.257] | 0.570 | 0.130 | [−0.047, 0.306] | 0.149 |
| Cog PGI x Age | −0.147 | [−0.288, −0.007] | 0.040 | −0.128 | [−0.256, −0.001] | 0.049 |
| (c) Speed of Processing | (d) Vocabulary | |||||
| Predictors | β | CI | p | β | CI | p |
| Cog PGI | 0.068 | [−0.102, 0.239] | 0.429 | 0.202 | [0.041, 0.363] | 0.014 |
| Age | −0.655 | [−0.871, −0.440] | <0.001 | 0.273 | [0.073, 0.472] | 0.008 |
| Education | 0.091 | [−0.039, 0.221] | 0.167 | 0.312 | [0.192, 0.432] | <0.001 |
| Sex [M] | −0.035 | [−0.334, 0.264] | 0.818 | −0.161 | [−0.442, 0.120] | 0.259 |
| PC 1 | 0.099 | [−0.065, 0.264] | 0.235 | −0.015 | [−0.168, 0.138] | 0.848 |
| Cortical Thickness | 0.038 | [−0.143, 0.220] | 0.677 | −0.055 | [−0.229, 0.119] | 0.536 |
| Total Gray Volume | 0.021 | [−0.161, 0.204] | 0.817 | 0.238 | [0.068, 0.408] | 0.006 |
| log(WMH + 1) | 0.070 | [−0.106, 0.246] | 0.436 | −0.004 | [−0.166, 0.158] | 0.959 |
| Cog PGI x Age | −0.096 | [−0.223, 0.031] | 0.138 | −0.162 | [−0.279, −0.044] | 0.007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsapanou, A.; Gacheru, M.; Lee, S.; Mourtzi, N.; Gazes, Y.; Habeck, C.; Belsky, D.W.; Stern, Y. Association of Cognitive Polygenic Index and Cognitive Performance with Age in Cognitively Healthy Adults. Genes 2023, 14, 1814. https://doi.org/10.3390/genes14091814
Tsapanou A, Gacheru M, Lee S, Mourtzi N, Gazes Y, Habeck C, Belsky DW, Stern Y. Association of Cognitive Polygenic Index and Cognitive Performance with Age in Cognitively Healthy Adults. Genes. 2023; 14(9):1814. https://doi.org/10.3390/genes14091814
Chicago/Turabian StyleTsapanou, Angeliki, Margaret Gacheru, Seonjoo Lee, Niki Mourtzi, Yunglin Gazes, Christian Habeck, Daniel W. Belsky, and Yaakov Stern. 2023. "Association of Cognitive Polygenic Index and Cognitive Performance with Age in Cognitively Healthy Adults" Genes 14, no. 9: 1814. https://doi.org/10.3390/genes14091814
APA StyleTsapanou, A., Gacheru, M., Lee, S., Mourtzi, N., Gazes, Y., Habeck, C., Belsky, D. W., & Stern, Y. (2023). Association of Cognitive Polygenic Index and Cognitive Performance with Age in Cognitively Healthy Adults. Genes, 14(9), 1814. https://doi.org/10.3390/genes14091814