The Impacts of Dam Construction and Removal on the Genetics of Recovering Steelhead (Oncorhynchus mykiss) Populations across the Elwha River Watershed

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
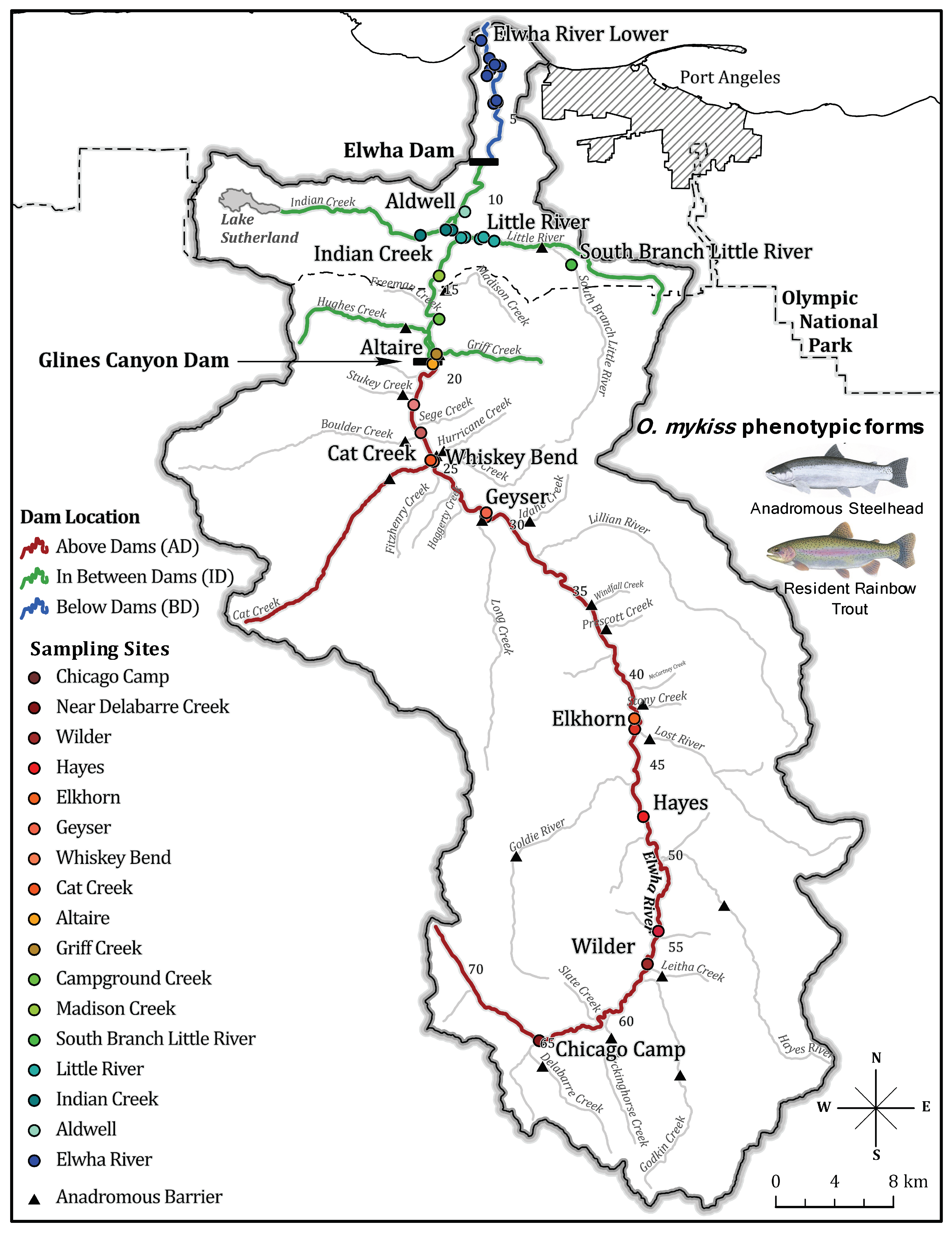
2.1. Sample Collection
2.2. Library Preparation and DNA Sequencing
2.3. Read Alignment and Filtering
2.4. Principal Component Analysis (PCA)
2.5. Population Genetic Structure Analysis
2.6. Population Genetics Analysis
2.7. Overview of Genetic Stock Identification (GSI) Analyses
2.8. Evaluation of Candidate Loci for Involvement in Migration Phenotypes
2.8.1. Candidate Genetic Variants for Migration and Run Timing
2.8.2. Testing for Correlations of Putatively Adaptive Alleles and Migration Phenotypes
2.8.3. Investigating the Geographic Structure of Adaptive Genetic Variants Involved in Migration
3. Results
3.1. RAD-Sequence Filtering and Quality Control
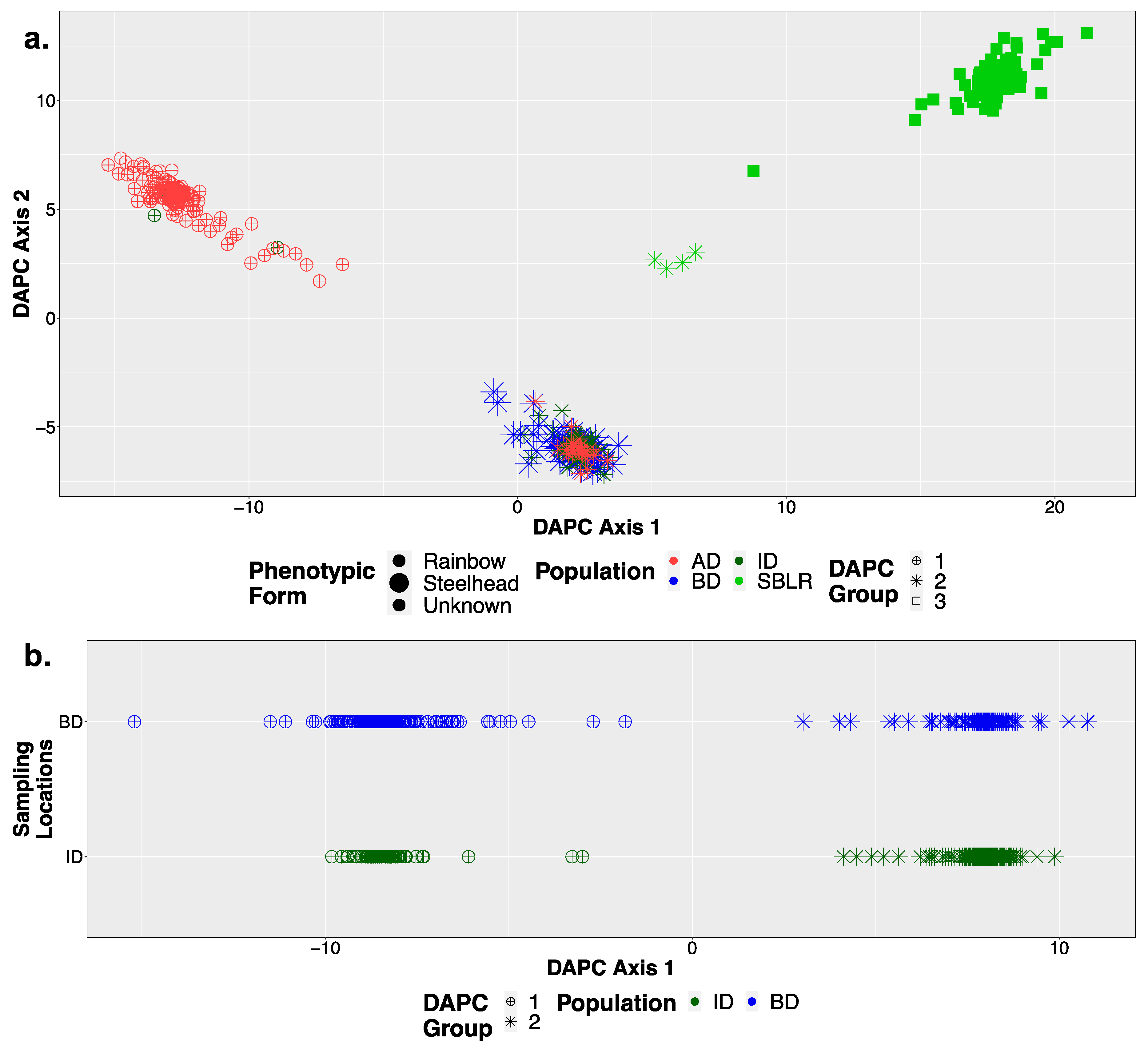
3.2. Principal Components Analysis (PCA)
3.3. Population Structural Analyses
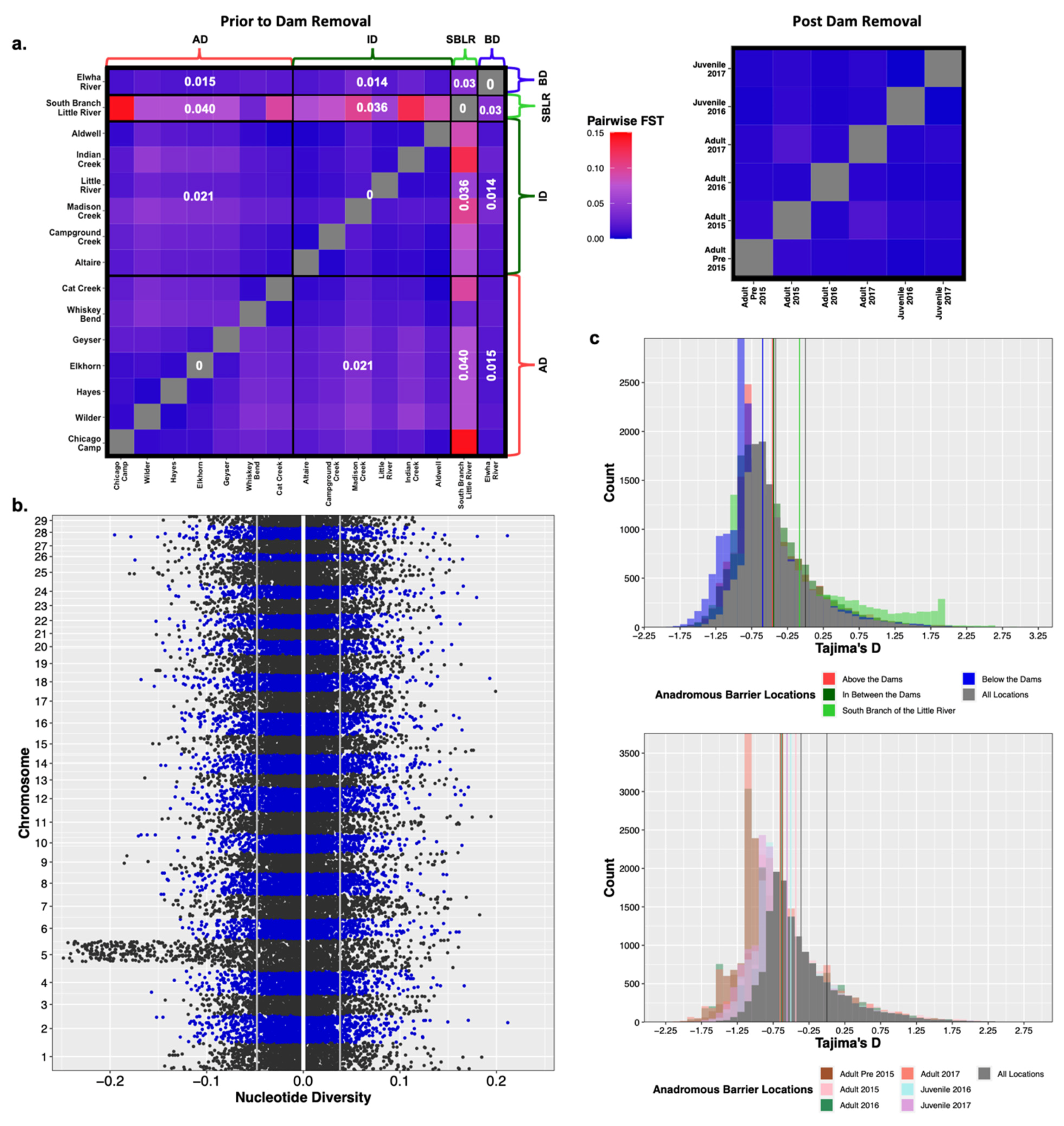
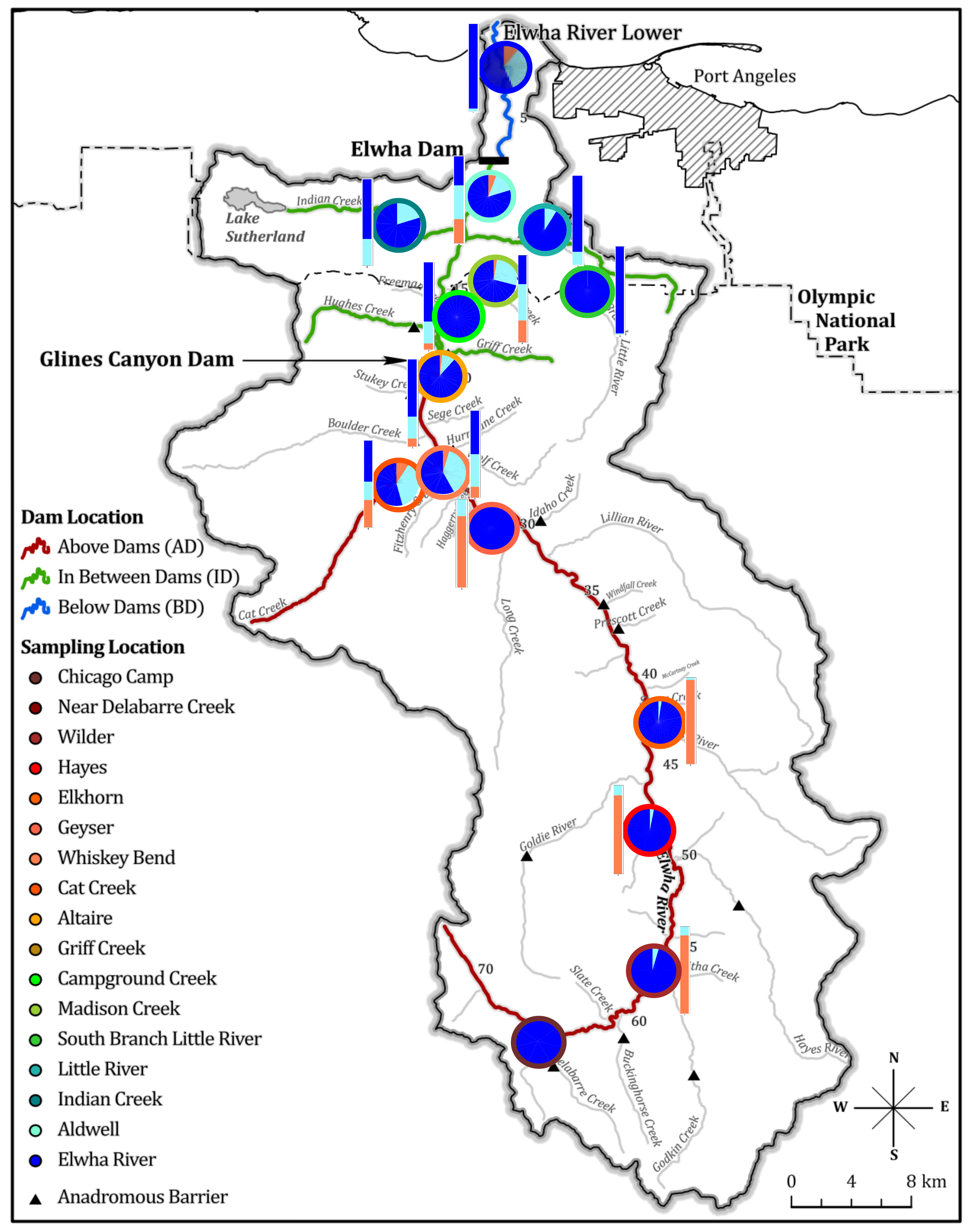
3.4. Population Genetic Diversity on the Elwha River
3.4.1. Natural Barriers and Dams Impacted Patterns of Genetic Diversity Pre-Dam Removal
3.4.2. No Observed Shift in Population Genetic Diversity Post-Dam Removal
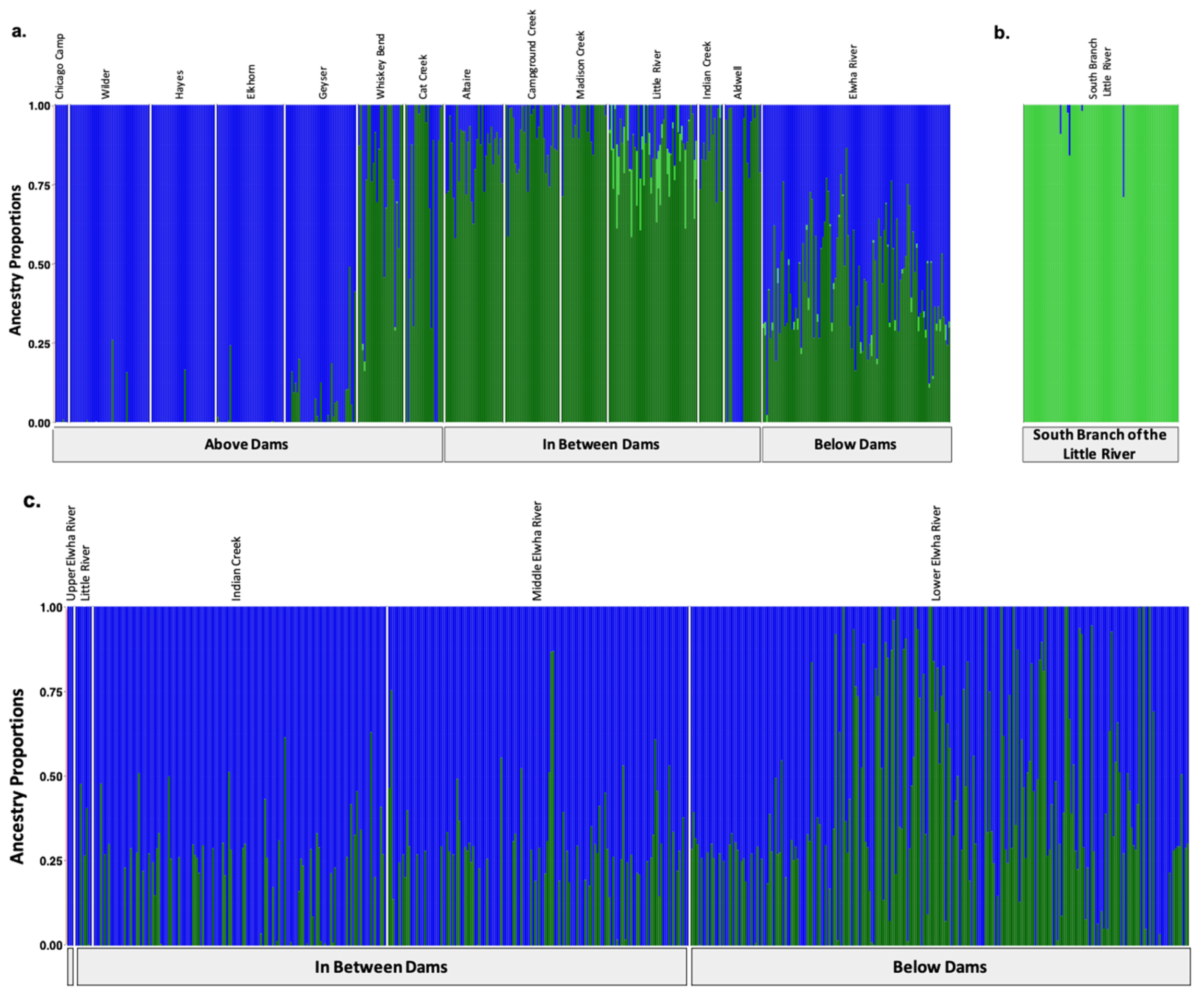
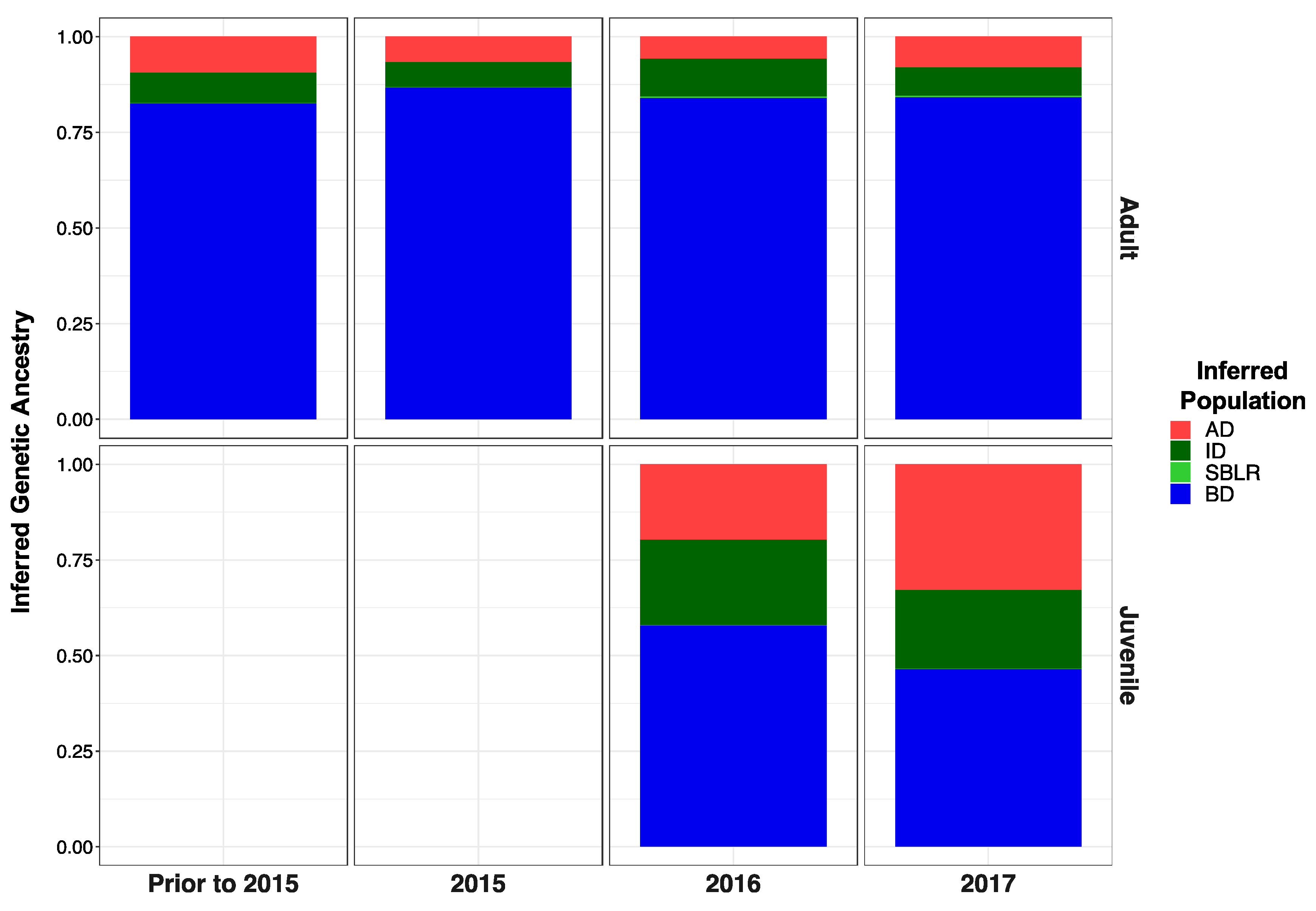
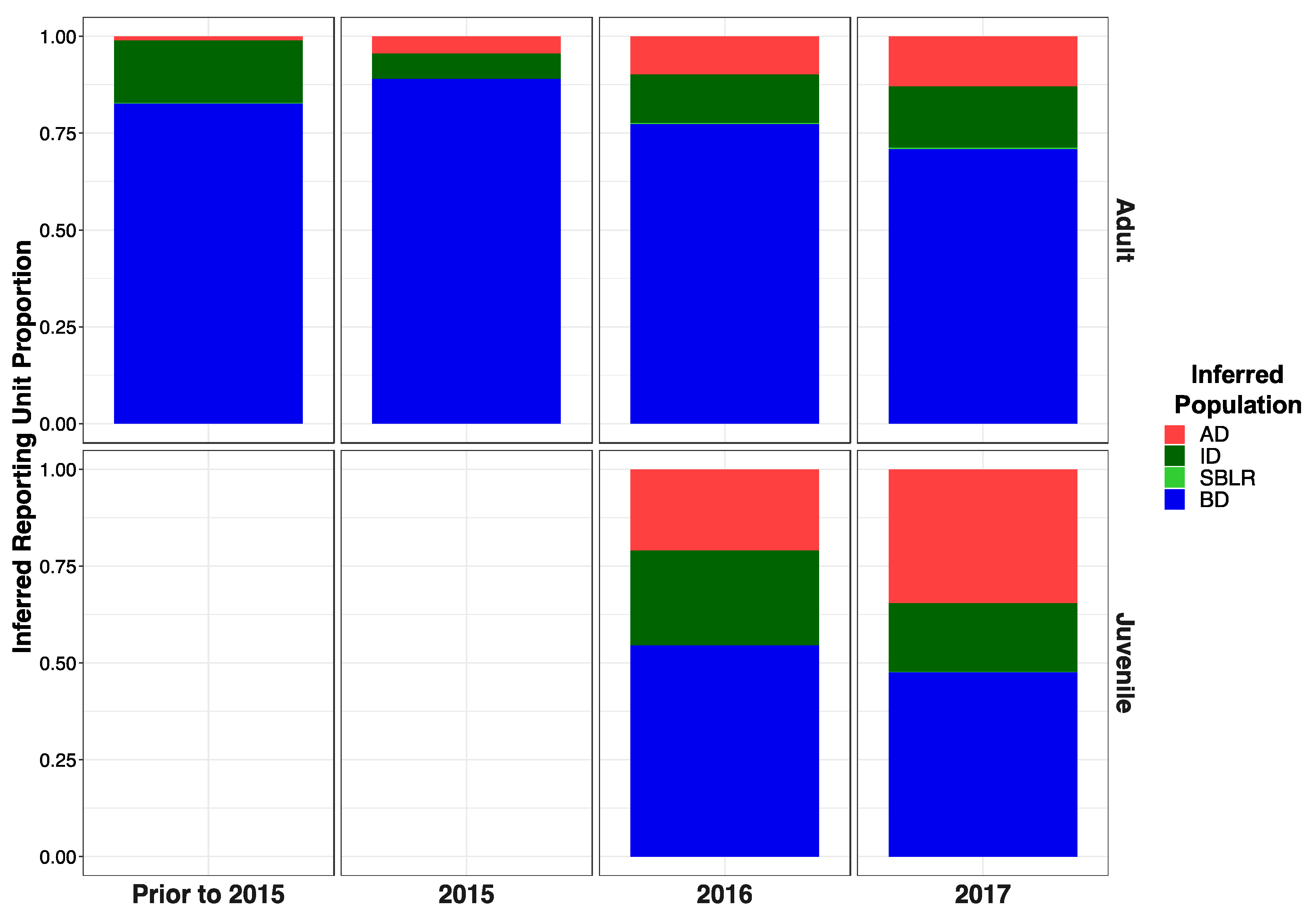
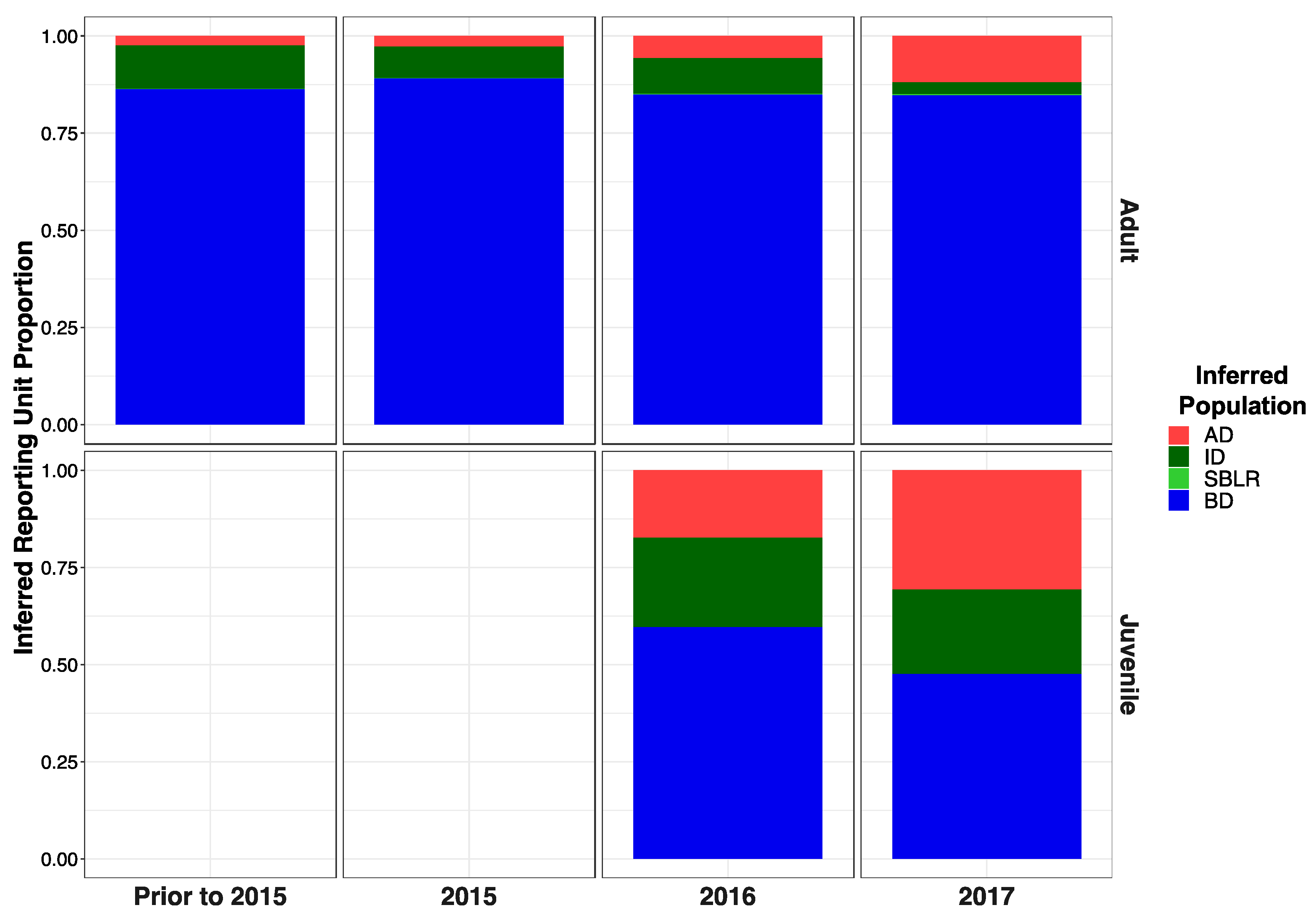
3.5. Genetic Stock Identification
3.5.1. Assessment of the Accuracy of Our Reference Sampling Sites (Collections) and Populations (Reporting Units)
3.5.2. Few Individuals Were Identified as of Potential Non-Elwha Origin
3.5.3. Recolonizing Steelhead Cohorts Appeared to Be Derived from All across the River
3.5.4. Recolonizing Steelhead Cohorts Appeared to Be Derived from All across the River
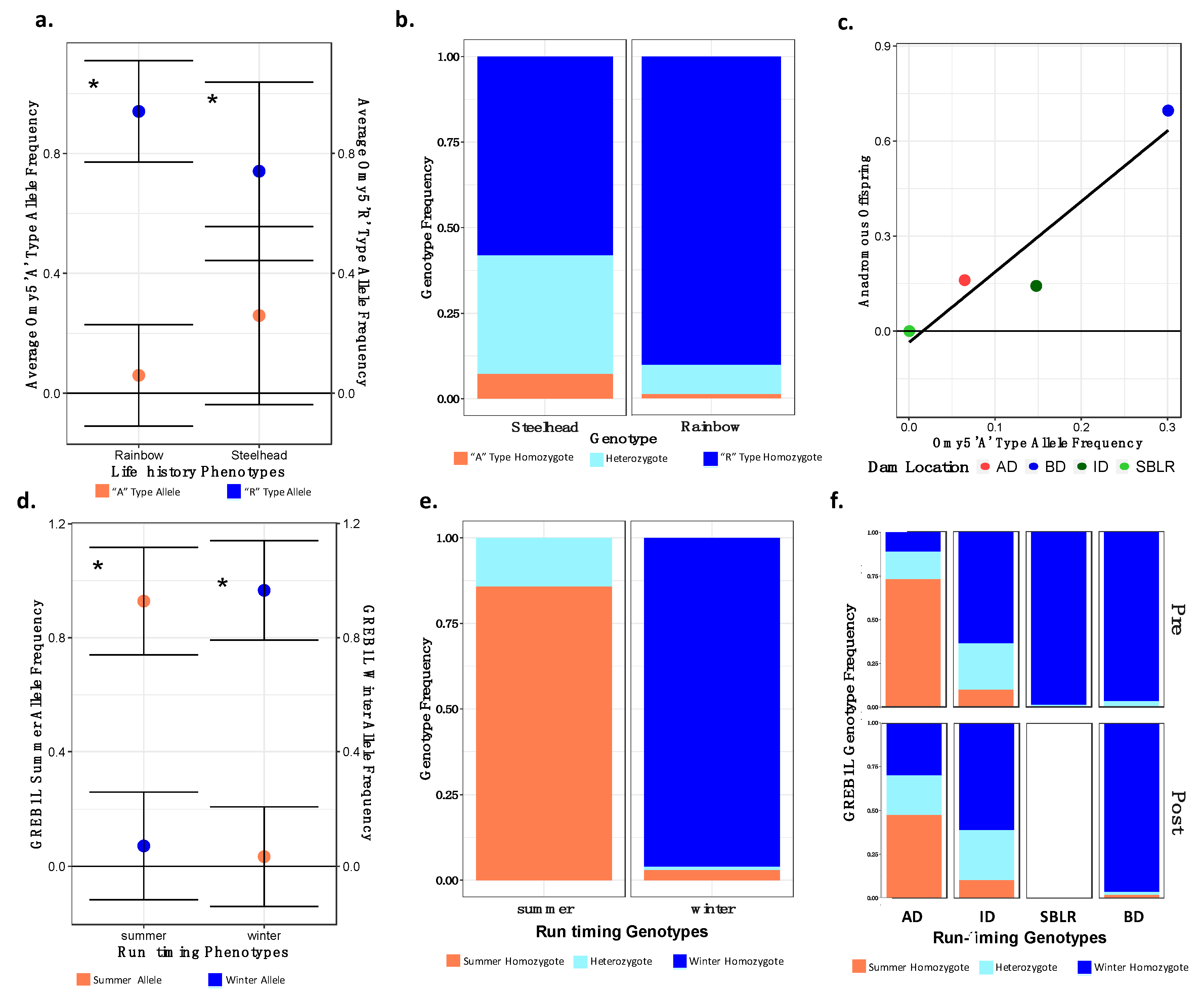
3.6. Allelic Variation in Candidate Loci for Migratory Life History Adaptations
3.6.1. Temporal and Geographic Variation in the Distribution of Adaptive Migratory Potential
3.6.2. Variation in the GREB1L Candidate for the Run Timing Phenotype
4. Discussion
4.1. Natural Barriers Had a Greater Effect on Patterns of Genetic Variation Than Dams
4.2. Population Genetic Structure Eroded Rapidly Following Removal of the Dams
4.3. Recolonizing Steelhead Appear to Primarily Be Derived from the Elwha River Watershed
4.4. Patterns of Genetic Diversity Did Not Change Following Dam Construction or Removal
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Reference and Mixed Sample Set Assignment
Appendix A.2. Selection of Loci to Be Used in GSI
- GSI analysis (i) Sample set one + SNP set one;
- GSI analysis (ii) Sample set two + SNP set two;
- GSI analysis (iii) Sample set two + SNP set one.
Appendix A.3. Self-Assignment Accuracy Assessment
Appendix A.4. GSI Mixture Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Reporting Unit | Inferred Reporting Unit | |||
|---|---|---|---|---|---|
| AD | ID | SBLR | BD | ||
| Chicago Camp | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Wilder | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Hayes | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Elkhorn | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Geyser | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Whiskey Bend | AD | 0.717 | 0.283 | 0.000 | 0.000 |
| Cat Creek | AD | 0.565 | 0.387 | 0.000 | 0.048 |
| Altaire | ID | 0.000 | 1.000 | 0.000 | 0.000 |
| Campground Creek | ID | 0.030 | 0.966 | 0.000 | 0.004 |
| Madison Creek | ID | 0.000 | 1.000 | 0.000 | 0.000 |
| Little River | ID | 0.001 | 0.996 | 0.000 | 0.003 |
| Indian Creek | ID | 0.000 | 1.000 | 0.000 | 0.000 |
| Aldwell | ID | 0.613 | 0.387 | 0.000 | 0.000 |
| South Branch Little River | SBLR | 0.000 | 0.012 | 0.942 | 0.046 |
| Elwha River | BD | 0.005 | 0.058 | 0.000 | 0.937 |
| Sampling Site | Reporting Unit | Inferred Reporting Unit | |||
|---|---|---|---|---|---|
| AD | ID | SBLR | BD | ||
| Chicago Camp | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Wilder | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Hayes | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Elkhorn | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Geyser | AD | 0.950 | 0.050 | 0.000 | 0.000 |
| Whiskey Bend | AD | 0.349 | 0.531 | 0.000 | 0.120 |
| Cat Creek | AD | 0.452 | 0.534 | 0.000 | 0.014 |
| Altaire | ID | 0.170 | 0.830 | 0.000 | 0.000 |
| Campground Creek | ID | 0.094 | 0.906 | 0.000 | 0.000 |
| Madison Creek | ID | 0.001 | 0.999 | 0.000 | 0.000 |
| Little River | ID | 0.020 | 0.980 | 0.000 | 0.000 |
| Indian Creek | ID | 0.000 | 1.000 | 0.000 | 0.000 |
| Aldwell | ID | 0.451 | 0.549 | 0.000 | 0.000 |
| South Branch Little River | SBLR | 0.004 | 0.031 | 0.943 | 0.022 |
| Elwha River | BD | 0.034 | 0.031 | 0.000 | 0.934 |


References
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, J.; Xu, J.; Zeng, G.; Sang, L.; Liu, Q.; Yin, Z.; Dai, J.; Yin, D.; Liang, J.; et al. Effects of dam construction on biodiversity: A review. J. Clean. Prod. 2019, 221, 480–489. [Google Scholar] [CrossRef]
- Liermann, C.R.; Nilsson, C.; Robertson, J.; Ng, R.Y. Implications of Dam Obstruction for Global Freshwater Fish Diversity. Bioscience 2012, 62, 539–548. [Google Scholar] [CrossRef]
- Gosset, C.; Rives, J.; LaBonne, J. Effect of habitat fragmentation on spawning migration of brown trout (Salmo trutta L.). Ecol. Freshw. Fish 2006, 15, 247–254. [Google Scholar] [CrossRef]
- Leclerc, E.; Mailhot, Y.; Mingelbier, M.; Bernatchez, L. The landscape genetics of yellow perch (Perca flavescens) in a large fluvial ecosystem. Mol. Ecol. 2008, 17, 1702–1717. [Google Scholar] [CrossRef]
- Winans, G.A.; Baker, J.; McHenry, M.; Ward, L.; Myers, J. Genetic Characterization of Oncorhynchus mykiss Prior to Dam Removal with Implications for Recolonization of the Elwha River Watershed, Washington. Trans. Am. Fish. Soc. 2017, 146, 160–172. [Google Scholar] [CrossRef]
- Winans, G.A.; Allen, M.B.; Baker, J.; Lesko, E.; Shrier, F.; Strobel, B.; Myers, J. Dam trout: Genetic variability in Oncorhynchus mykiss above and below barriers in three Columbia River systems prior to restoring migrational access. PLoS ONE 2018, 13, e0197571. [Google Scholar] [CrossRef]
- Poff, N.L.; Olden, J.D.; Merritt, D.M.; Pepin, D.M. Homogenization of regional river dynamics by dams and global biodiversity implications. Proc. Natl. Acad. Sci. USA 2007, 104, 5732–5737. [Google Scholar] [CrossRef]
- Beechie, T.; Moir, H.; Pess, G. Hierarchical Physical Controls on Salmonid Spawning Location and Timing. In Salmonid Spawning Habitat in Rivers: Physical Controls, Biological Responses, and Approaches to Remediation; American Fisheries Society: Bethesda, MD, USA, 2008; Volume 65, pp. 83–102. [Google Scholar]
- Pearse, D.E.; Hayes, S.A.; Bond, M.H.; Hanson, C.V.; Anderson, E.C.; Macfarlane, R.B.; Garza, J.C. Over the Falls? Rapid Evolution of Ecotypic Differentiation in Steelhead/Rainbow Trout (Oncorhynchus mykiss). J. Hered. 2009, 100, 515–525. [Google Scholar] [CrossRef]
- Nehlsen, W.; Williams, J.E.; Lichatowich, J.A. Pacific Salmon at the Crossroads—Stocks at Risk from California, Oregon, Idaho, and Washington. Fisheries 1991, 16, 4–21. [Google Scholar] [CrossRef]
- Ackiss, A.S.; Dang, B.T.; Bird, C.E.; Biesack, E.E.; Chheng, P.; Phounvisouk, L.; Vu, Q.H.; Uy, S.; Carpenter, K.E. Cryptic Lineages and a Population Dammed to Incipient Extinction? Insights into the Genetic Structure of a Mekong River Catfish. J. Hered. 2019, 110, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Holecek, D.E.; Scarnecchia, D.L.; Miller, S.E. Smoltification in an Impounded, Adfluvial Redband Trout Population Upstream from an Impassable Dam: Does It Persist? Trans. Am. Fish. Soc. 2012, 141, 68–75. [Google Scholar] [CrossRef]
- McClure, M.M.; Carlson, S.M.; Beechie, T.J.; Pess, G.R.; Jorgensen, J.C.; Sogard, S.M.; Sultan, S.E.; Holzer, D.M.; Travis, J.; Sanderson, B.L.; et al. Evolutionary consequences of habitat loss for Pacific anadromous salmonids. Evol. Appl. 2008, 1, 300–318. [Google Scholar] [CrossRef]
- Quinn, T.P. The Behaviour and Ecology of Pacific Salmon and Trout; University of Washington Press: Seattle, WA, USA, 2005. [Google Scholar]
- Behnke, R.J. Trout and Salmon of North America; The Free Press: New York, NY, USA, 2002. [Google Scholar]
- Moyle, P.B. Inland Fishes of California: Revised and Expanded; University of California Press: Berkeley, CA, USA, 2002. [Google Scholar]
- Waples, R.S.; Zabel, R.W.; Scheuerell, M.D.; Sanderson, B.L. Evolutionary responses by native species to major anthropogenic changes to their ecosystems: Pacific salmon in the Columbia River hydropower system. Mol. Ecol. 2008, 17, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.B.; Wuttig, K.; Fleming, D.; Kretschmer, E.J.; Wenburg, J.K. Evidence of partial anadromy and resident-form dispersal bias on a fine scale in populations of Oncorhynchus mykiss. Conserv. Genet. 2006, 7, 613–619. [Google Scholar] [CrossRef]
- Fraser, D.J.; Weir, L.K.; Bernatchez, L.; Hansen, M.M.; Taylor, E.B. Extent and scale of local adaptation in salmonid fishes: Review and meta-analysis. Heredity 2011, 106, 404–420. [Google Scholar] [CrossRef]
- Katz, J.; Moyle, P.B.; Quiñones, R.M.; Israel, J.; Purdy, S. Impending extinction of salmon, steelhead, and trout (Salmonidae) in California. Environ. Biol. Fishes 2012, 96, 1169–1186. [Google Scholar] [CrossRef]
- Kendall, N.W.; McMillan, J.R.; Sloat, M.R.; Buehrens, T.W.; Quinn, T.P.; Pess, G.R.; Kuzishchin, K.V.; McClure, M.M.; Zabel, R.W.; Bradford, M. Anadromy and residency in steelhead and rainbow trout (Oncorhynchus mykiss): A review of the processes and patterns. Can. J. Fish. Aquat. Sci. 2015, 72, 319–342. [Google Scholar] [CrossRef]
- Hoar, W.S. The Physiology of Smolting Salmonids. In Fish Physiology; Academic Press: Cambridge, MA, USA, 1988; Volume 11, pp. 275–343. [Google Scholar]
- Nichols, K.M.; Edo, A.F.; Wheeler, P.A.; Thorgaard, G.H. The Genetic Basis of Smoltification-Related Traits in Oncorhynchus mykiss. Genetics 2008, 179, 1559–1575. [Google Scholar] [CrossRef]
- Thrower, F.P.; Hard, J.J.; Joyce, J.E. Genetic architecture of growth and early life history transitions in anadromous and derived freshwater populations of steelhead. J. Fish Biol. 2004, 65, 286–307. [Google Scholar] [CrossRef]
- Hecht, B.C.; Thrower, F.P.; Hale, M.C.; Miller, M.R.; Nichols, K.M. Genetic Architecture of Migration-Related Traits in Rainbow and Steelhead Trout, Oncorhynchus mykiss. G3 Genes Genomes Genet. 2012, 2, 1113–1127. [Google Scholar] [CrossRef] [PubMed]
- Narum, S.R.; Zendt, J.S.; Graves, D.; Sharp, W.R. Influence of landscape on resident and anadromous life history types of Oncorhynchus mykiss. Can. J. Fish. Aquat. Sci. 2008, 65, 1013–1023. [Google Scholar] [CrossRef]
- McMillan, J.R.; Dunham, J.B.; Reeves, G.H.; Mills, J.S.; Jordan, C.E. Individual condition and stream temperature influence early maturation of rainbow and steelhead trout, Oncorhynchus mykiss. Environ. Biol. Fishes 2011, 93, 343–355. [Google Scholar] [CrossRef]
- Hecht, B.C.; Valle, M.E.; Thrower, F.P.; Nichols, K.M. Divergence in Expression of Candidate Genes for the Smoltification Process Between Juvenile Resident Rainbow and Anadromous Steelhead Trout. Mar. Biotechnol. 2014, 16, 638–656. [Google Scholar] [CrossRef] [PubMed]
- McKinney, G.J.; Hale, M.C.; Goetz, G.; Gribskov, M.; Thrower, F.P.; Nichols, K.M. Ontogenetic changes in embryonic and brain gene expression in progeny produced from migratory and resident Oncorhynchus mykiss. Mol. Ecol. 2015, 24, 1792–1809. [Google Scholar] [CrossRef]
- Narum, S.R.; Contor, C.; Powell, M.S. Genetic divergence of sympatric resident and anadromous forms of Oncorhynchus mykiss in the Walla Walla River, U.S.A. J. Fish Biol. 2004, 65, 471–488. [Google Scholar] [CrossRef]
- Heath, A.C.; Martin, N.G.; Montgomery, G.W.; Goddard, M.E.; Visscher, P.M. Common SNPs explain a large proportion of the heritability for human height. Nat. Genet. 2010, 42, 565–569. [Google Scholar]
- Clemento, A.J.; Anderson, E.C.; Boughton, D.; Girman, D.; Garza, J.C. Population genetic structure and ancestry of Oncorhynchus mykiss populations above and below dams in south-central California. Conserv. Genet. 2009, 10, 1321–1336. [Google Scholar] [CrossRef]
- Docker, M.F.; Dale, A.; Heath, D.D. Erosion of interspecific reproductive barriers resulting from hatchery supplementation of rainbow trout sympatric with cutthroat trout. Mol. Ecol. 2003, 12, 3515–3521. [Google Scholar] [CrossRef]
- Deiner, K.; Garza, J.C.; Coey, R.; Girman, D. Population structure and genetic diversity of trout (Oncorhynchus mykiss) above and below natural and man-made barriers in the Russian River, California. Conserv. Genet. 2007, 8, 437–454. [Google Scholar] [CrossRef]
- Pearse, D.E.; Miller, M.R.; Abadía-Cardoso, A.; Garza, J.C. Rapid parallel evolution of standing variation in a single, complex, genomic region is associated with life history in steelhead/rainbow trout. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140012. [Google Scholar] [CrossRef] [PubMed]
- Leitwein, M.; Garza, J.C.; Pearse, D.E. Ancestry and adaptive evolution of anadromous, resident, and adfluvial rainbow trout (Oncorhynchus mykiss) in the San Francisco bay area: Application of adaptive genomic variation to conservation in a highly impacted landscape. Evol. Appl. 2017, 10, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Micheletti, S.J.; Hess, J.E.; Zendt, J.S.; Narum, S.R. Selection at a genomic region of major effect is responsible for evolution of complex life histories in anadromous steelhead. BMC Evol. Biol. 2018, 18, 140. [Google Scholar] [CrossRef] [PubMed]
- Pearse, D.E.; Barson, N.J.; Nome, T.; Gao, G.; Campbell, M.A.; Abadía-Cardoso, A.; Anderson, E.C.; Rundio, D.E.; Williams, T.H.; Naish, K.A.; et al. Sex-dependent dominance maintains migration supergene in rainbow trout. Nat. Ecol. Evol. 2019, 3, 1731–1742. [Google Scholar] [CrossRef]
- Hecht, B.C.; Campbell, N.R.; Holecek, D.E.; Narum, S.R. Genome-wide association reveals genetic basis for the propensity to migrate in wild populations of rainbow and steelhead trout. Mol. Ecol. 2013, 22, 3061–3076. [Google Scholar] [CrossRef] [PubMed]
- Arostegui, M.C.; Quinn, T.P.; Seeb, L.W.; Seeb, J.E.; McKinney, G.J. Retention of a chromosomal inversion from an anadromous ancestor provides the genetic basis for alternative freshwater ecotypes in rainbow trout. Mol. Ecol. 2019, 28, 1412–1427. [Google Scholar] [CrossRef]
- Weinstein, S.Y.; Thrower, F.P.; Nichols, K.M.; Hale, M.C. A large-scale chromosomal inversion is not associated with life history development in rainbow trout from Southeast Alaska. PLoS ONE 2019, 14, e0223018. [Google Scholar] [CrossRef]
- Dodson, J.J.; Aubin-Horth, N.; Theriault, V.; Paez, D.J. The evolutionary ecology of alternative migratory tactics in salmonid fishes. Biol. Rev. 2013, 88, 602–625. [Google Scholar] [CrossRef]
- Hess, J.E.; Zendt, J.S.; Matala, A.R.; Narum, S.R. Genetic basis of adult migration timing in anadromous steelhead discovered through multivariate association testing. Proc. R. Soc. B Biol. Sci. 2016, 283, 20153064. [Google Scholar] [CrossRef]
- Prince, D.J.; O’Rourke, S.M.; Thompson, T.Q.; Ali, O.A.; Lyman, H.S.; Saglam, I.K.; Hotaling, T.J.; Spidle, A.P.; Miller, M.R. The evolutionary basis of premature migration in Pacific salmon highlights the utility of genomics for informing conservation. Sci. Adv. 2017, 3, e1603198. [Google Scholar] [CrossRef]
- Micheletti, S.J.; Matala, A.R.; Matala, A.P.; Narum, S.R. Landscape features along migratory routes influence adaptive genomic variation in anadromous steelhead (Oncorhynchus mykiss). Mol. Ecol. 2018, 27, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Thompson, T.Q.; Bellinger, M.R.; O’Rourke, S.M.; Prince, D.J.; Stevenson, A.E.; Rodrigues, A.T.; Sloat, M.R.; Speller, C.F.; Yang, D.Y.; Butler, V.L.; et al. Anthropogenic habitat alteration leads to rapid loss of adaptive variation and restoration potential in wild salmon populations. Proc. Natl. Acad. Sci. USA 2019, 116, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Thompson, N.F.; Anderson, E.C.; Clemento, A.J.; Campbell, M.A.; Pearse, D.E.; Hearsey, J.W.; Kinziger, A.P.; Garza, J.C. A complex phenotype in salmon controlled by a simple change in migratory timing. Science 2020, 370, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Willis, S.C.; Hess, J.E.; Fryer, J.K.; Whiteaker, J.M.; Brun, C.; Gerstenberger, R.; Narum, S.R. Steelhead (Oncorhynchus mykiss) lineages and sexes show variable patterns of association of adult migration timing and age-at-maturity traits with two genomic regions. Evol. Appl. 2020, 13, 2836–2856. [Google Scholar] [CrossRef] [PubMed]
- Collins, E.E.; Hargrove, J.S.; Delomas, T.A.; Narum, S.R. Distribution of genetic variation underlying adult migration timing in steelhead of the Columbia River basin. Ecol. Evol. 2020, 10, 9486–9502. [Google Scholar] [CrossRef]
- Lindley, S.T.; Schick, R.S.; Agrawal, A.; Goslin, M.; Pearson, T.E.; Mora, E.; Anderson, J.J.; May, B.; Greene, S.; Hanson, C.; et al. Historical Population Structure of Central Valley Steelhead and its Alteration by Dams. San Franc. Estuary Watershed Sci. 2006, 4. [Google Scholar] [CrossRef]
- Blanchet, S.; Prunier, J.G.; Paz-Vinas, I.; Saint-Pé, K.; Rey, O.; Raffard, A.; Mathieu-Bégné, E.; Loot, G.; Fourtune, L.; Dubut, V. A river runs through it: The causes, consequences, and management of intraspecific diversity in river networks. Evol. Appl. 2020, 13, 1195–1213. [Google Scholar] [CrossRef]
- Loomis, J.B. Measuring the Economic Benefits of Removing Dams and Restoring the Elwha River: Results of a Contingent Valuation Survey. Water Resour. Res. 1996, 32, 441–447. [Google Scholar] [CrossRef]
- Wunderlich, R.C.; Winter, B.D.; Meyer, J.H. Restoration of the Elwha River Ecosystem. Fisheries 1994, 19, 11–19. [Google Scholar] [CrossRef]
- Duda, J.J.; Freilich, J.E.; Schreiner, E.G. Baseline Studies in the Elwha River Ecosystem Prior to Dam Removal: Introduction to the Special Issue. Northwest Sci. 2008, 82, 1–12. [Google Scholar] [CrossRef]
- Winans, G.A.; McHenry, M.L.; Baker, J.; Elz, A.; Goodbla, A.; Iwamoto, E.; Kuligowski, D.; Miller, K.M.; Small, M.P.; Spruell, P.; et al. Genetic Inventory of Anadromous Pacific Salmonids of the Elwha River Prior to Dam Removal. Northwest Sci. 2008, 82, 128–141. [Google Scholar] [CrossRef]
- Pess, G.R.; McHenry, M.L.; Beechie, T.J.; Davies, J. Biological Impacts of the Elwha River Dams and Potential Salmonid Responses to Dam Removal. Northwest Sci. 2008, 82, 72–90. [Google Scholar] [CrossRef]
- Brenkman, S.J.; Pess, G.R.; Torgersen, C.E.; Kloehn, K.K.; Duda, J.J.; Corbett, S.C. Predicting Recolonization Patterns and Interactions Between Potamodromous and Anadromous Salmonids in Response to Dam Removal in the Elwha River, Washington State, USA. Northwest Sci. 2008, 82, 91–106. [Google Scholar] [CrossRef]
- Brenkman, S.J.; Mumford, S.L.; House, M.; Patterson, C. Establishing Baseline Information on the Geographic Distribution of Fish Pathogens Endemic in Pacific Salmonids Prior to Dam Removal and Subsequent Recolonization by Anadromous Fish in the Elwha River, Washington. Northwest Sci. 2008, 82, 142–152. [Google Scholar] [CrossRef]
- Brenkman, S.J.; Duda, J.J.; Torgersen, C.E.; Welty, E.; Pess, G.R.; Peters, R.; McHenry, M.L. A riverscape perspective of Pacific salmonids and aquatic habitats prior to large-scale dam removal in the Elwha River, Washington, USA. Fish. Manag. Ecol. 2012, 19, 36–53. [Google Scholar] [CrossRef]
- Pess, G.R.; McHenry, M.L.; Denton, K.; Anderson, J.H.; Liermann, M.C.; Peters, R.J.; Brenkman, S.; Bennett, T.R. Initial response of Chinook salmon (Oncorhynchus tshawytscha) and steelhead (Oncorhynchus mykiss) to removal of two dams on the Elwha River, Washington State, U.S.A. Can. J. Fish. Aquat. Sci. 2020. in review. [Google Scholar]
- Ali, O.A.; O’Rourke, S.M.; Amish, S.J.; Meek, M.H.; Luikart, G.; Jeffres, C.; Miller, M.R. RAD Capture (Rapture): Flexible and Efficient Sequence-Based Genotyping. Genetics 2016, 202, 389–400. [Google Scholar] [CrossRef]
- Paris, J.R.; Stevens, J.R.; Catchen, J.M. Lost in parameter space: A road map for stacks. Methods Ecol. Evol. 2017, 10, 1360–1373. [Google Scholar] [CrossRef]
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An analysis tool set for population genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Making the Leap: Maq to BWA. Mass Genom. 2009, 25, 1754–1760. [Google Scholar]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; Depristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Glasauer, S.M.; Neuhauss, S.C. Whole-genome duplication in teleost fishes and its evolutionary consequences. Mol. Genet. Genom. 2014, 289, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- McKinney, G.J.; Waples, R.K.; Seeb, L.W.; Seeb, J.E. Paralogs are revealed by proportion of heterozygotes and deviations in read ratios in genotyping-by-sequencing data from natural populations. Mol. Ecol. Resour. 2017, 17, 656–669. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 11, 1403–1405. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.; Stephens, M.; Pritchard, J.K. fastSTRUCTURE: Variational Inference of Population Structure in Large SNP Data Sets. Genetics 2014, 197, 573–589. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Tajima, F. Statistical Method for Testing the Neutral Mutation Hypothesis by DNA Polymorphism. Genetics 1989, 123, 585–595. [Google Scholar]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator v2: Re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour. 2014, 14, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Puth, M.T.; Neuhäuser, M.; Ruxton, G.D. On the variety of methods for calculating confidence intervals by bootstrapping. J. Anim. Ecol. 2015, 84, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Moran, B.M.; Anderson, E.C. Bayesian inference from the conditional genetic stock identification model. Can. J. Fish. Aquat. Sci. 2019, 76, 551–560. [Google Scholar] [CrossRef]
- Anderson, E.C.; Waples, R.S.; Kalinowski, S.T. An improved method for predicting the accuracy of genetic stock identification. Can. J. Fish. Aquat. Sci. 2008, 65, 1475–1486. [Google Scholar] [CrossRef]
- Miller, M.R.; Brunelli, J.P.; Wheeler, P.A.; Liu, S.; Rexroad, C.E., 3rd; Palti, Y.; Doe, C.Q.; Thorgaard, G.H. A conserved haplotype controls parallel adaptation in geographically distant salmonid populations. Mol. Ecol. 2012, 21, 237–249. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Rousset, F. genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Abadía-Cardoso, A.; Pearse, D.E.; Jacobson, S.; Marshall, J.; Dalrymple, D.; Kawasaki, F.; Ruiz-Campos, G.; Garza, J.C. Population genetic structure and ancestry of steelhead/rainbow trout (Oncorhynchus mykiss) at the extreme southern edge of their range in North America. Conserv. Genet. 2016, 17, 675–689. [Google Scholar] [CrossRef]
- Phelps, S.R.; Hiss, J.M.; Peters, R.J. Genetic Relationships of Elwha River Oncorhynchus mykiss to Hatchery-Origin Rainbow Trout and Washington Steelhead; Washington Department of Fish and Wildlife: Olympia, WA, USA, 1999.
- Robison, E.G.; Mirati, A.; Allen, M. Oregon Road/Stream Crossing Restoration Guide: Spring 1999; Advanced Fish Passage Training Version; Oregon Department of Fish and Wildlife: Bend, OR, USA, 2000.
- Samarasin, P.; Shuter, B.J.; Rodd, F.H. After 100 years: Hydroelectric dam-induced life-history divergence and population genetic changes in sockeye salmon (Oncorhynchus nerka). Conserv. Genet. 2017, 18, 1449–1462. [Google Scholar] [CrossRef]
- Limborg, M.T.; Blankenship, S.M.; Young, S.F.; Utter, F.M.; Seeb, L.W.; Hansen, M.H.; Seeb, J.E. Signatures of natural selection among lineages and habitats inOncorhynchus mykiss. Ecol. Evol. 2012, 2, 1–18. [Google Scholar] [CrossRef]
- Busby, P.J.; Wainright, T.C.; Bryant, G.J.; Lierheimer, L.J.; Waples, R.S.; Waknitz, F.W.; Lagomarsino, I.V. Status Review of West Coast Steelhead from Washington, Idaho, Oregon, and California; US Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Northwest Fisheries Science Center, Coastal Zone and Estuarine Studies Division; National Technical Information Service, U.S. Department of Commerce: Springfield, VA, USA, 1996.
- Peacock, M.M.; Gustin, M.S.; Kirchoff, V.S.; Robinson, M.L.; Hekkala, E.; Pizzarro-Barraza, C.; Loux, T. Native fishes in the Truckee River: Are in-stream structures and patterns of population genetic structure related? Sci. Total Environ. 2016, 563, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Kelson, S.J.; Miller, M.R.; Thompson, T.Q.; O’Rourke, S.M.; Carlson, S.M. Temporal dynamics of migration-linked genetic variation are driven by streamflows and riverscape permeability. Mol. Ecol. 2020, 29, 870–885. [Google Scholar] [CrossRef] [PubMed]
- Hiss, J.M.; Wunderlich, R.C. Salmonid Availability and Migration in the Middle Elwha River System. Miscellaneous Report; Western Washington Fishery Resource Office, U.S. Fish and Wildlife Service: Olympia, WA, USA, 1994.
- Keefer, M.L.; Caudill, C.C. Homing and straying by anadromous salmonids: A review of mechanisms and rates. Rev. Fish Biol. Fish. 2014, 24, 333–368. [Google Scholar] [CrossRef]
- Hendry, A.P.; Castric, V.; Kinnison, M.T.; Quinn, T.P. The evolution of philopatry and dispersal. In Evolution Illuminated: Salmon and Their Relatives; Hendry, A., Stearns, S., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 52–91. [Google Scholar]
- Ostberg, C.O.; Slatton, S.L.; Rodriguez, R.J. Spatial partitioning and asymmetric hybridization among sympatric coastal steelhead trout (Oncorhynchus mykiss irideus), coastal cutthroat trout (O. clarki clarki) and interspecific hybrids. Mol. Ecol. 2004, 13, 2773–2788. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 12, 1225–1241. [Google Scholar] [CrossRef]
- Davis, C.D.; Epps, C.W.; Flitcroft, R.L.; Banks, M.A. Refining and defining riverscape genetics: How rivers influence population genetic structure. Wiley Interdiscip. Rev. Water 2018, 5, e1269. [Google Scholar] [CrossRef]
- Brannon, E.L.; Powell, M.S.; Quinn, T.P.; Talbot, A. Population Structure of Columbia River Basin Chinook Salmon and Steelhead Trout. Rev. Fish. Sci. 2010, 12, 99–232. [Google Scholar] [CrossRef]
- Luikart, G.; Ryman, N.; Tallmon, D.A.; Schwartz, M.K.; Allendorf, F.W. Estimation of census and effective population sizes: The increasing usefulness of DNA-based approaches. Conserv. Genet. 2010, 11, 355–373. [Google Scholar] [CrossRef]







| Relative Dam Location Sampling Site | Prior to Dam Removal | Post Dam Removal |
|---|---|---|
| Above the Dams (AD) | 208 | 3 |
| Cat Creek | 21 | |
| Chicago Camp | 7 | |
| Elkhorn | 37 | |
| Elwha River | 3 | |
| Geyser | 39 | |
| Hayes | 35 | |
| Whiskey Bend | 25 | |
| Wilder | 44 | |
| In between the Dams (ID) | 169 | 304 |
| Aldwell | 20 | |
| Altaire | 32 | |
| Campground Creek | 30 | |
| Elwha River | 150 | |
| Indian Creek | 13 | 146 |
| Little River | 49 | 8 |
| Madison Creek | 25 | |
| South Branch of the Little River (SBLR) | 86 | 0 |
| Below the Dams (BD) | 104 | 249 |
| Total | 567 | 556 |
| Sampling Site | Population | Inferred Population | |||
|---|---|---|---|---|---|
| AD | ID | SBLR | BD | ||
| Chicago Camp | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Wilder | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Hayes | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Elkhorn | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Geyser | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Cat Creek | AD | 1.000 | 0.000 | 0.000 | 0.000 |
| Altaire | ID | 0.004 | 0.996 | 0.000 | 0.000 |
| Campground Creek | ID | 0.105 | 0.895 | 0.000 | 0.000 |
| Madison Creek | ID | 0.000 | 1.000 | 0.000 | 0.000 |
| Little River | ID | 0.000 | 1.000 | 0.000 | 0.000 |
| Indian Creek | ID | 0.000 | 1.000 | 0.000 | 0.000 |
| Aldwell | ID | 0.000 | 1.000 | 0.000 | 0.000 |
| SBLR | SBLR | 0.012 | 0.000 | 0.976 | 0.012 |
| Elwha River | BD | 0.148 | 0.020 | 0.000 | 0.833 |
| Mixture Collection | Mean Posterior Probability | Mean Posterior Probability Credible Interval | ||||||
|---|---|---|---|---|---|---|---|---|
| AD | ID | SBLR | BD | AD | ID | SBLR | BD | |
| Adults Pre 2015 | 0.094 | 0.0789 | 0.001 | 0.825 | 0.035–0.183 | 0.024–0.165 | 0–0.012 | 0.716–0.913 |
| Adults 2015 | 0.067 | 0.065 | 0.000 | 0.868 | 0.034–0.109 | 0.032–0.105 | 0–0.005 | 0.813–0.916 |
| Adults 2016 | 0.057 | 0.100 | 0.003 | 0.840 | 0.004–0.175 | 0.016–0.239 | 0–0.035 | 0.673–0.956 |
| Adults 2017 | 0.080 | 0.074 | 0.003 | 0.842 | 0.006–0.246 | 0.005–0.223 | 0–0.035 | 0.644–0.969 |
| Juveniles 2016 | 0.198 | 0.223 | 0.000 | 0.578 | 0.141–0.257 | 0.164–0.288 | 0–0.004 | 0.502–0.654 |
| Juveniles 2017 | 0.330 | 0.206 | 0.001 | 0.464 | 0.243–0.414 | 0.138–0.282 | 0–0.006 | 0.374–0.562 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraik, A.K.; McMillan, J.R.; Liermann, M.; Bennett, T.; McHenry, M.L.; McKinney, G.J.; Wells, A.H.; Winans, G.; Kelley, J.L.; Pess, G.R.; et al. The Impacts of Dam Construction and Removal on the Genetics of Recovering Steelhead (Oncorhynchus mykiss) Populations across the Elwha River Watershed. Genes 2021, 12, 89. https://doi.org/10.3390/genes12010089
Fraik AK, McMillan JR, Liermann M, Bennett T, McHenry ML, McKinney GJ, Wells AH, Winans G, Kelley JL, Pess GR, et al. The Impacts of Dam Construction and Removal on the Genetics of Recovering Steelhead (Oncorhynchus mykiss) Populations across the Elwha River Watershed. Genes. 2021; 12(1):89. https://doi.org/10.3390/genes12010089
Chicago/Turabian StyleFraik, Alexandra K., John R. McMillan, Martin Liermann, Todd Bennett, Michael L. McHenry, Garrett J. McKinney, Abigail H. Wells, Gary Winans, Joanna L. Kelley, George R. Pess, and et al. 2021. "The Impacts of Dam Construction and Removal on the Genetics of Recovering Steelhead (Oncorhynchus mykiss) Populations across the Elwha River Watershed" Genes 12, no. 1: 89. https://doi.org/10.3390/genes12010089
APA StyleFraik, A. K., McMillan, J. R., Liermann, M., Bennett, T., McHenry, M. L., McKinney, G. J., Wells, A. H., Winans, G., Kelley, J. L., Pess, G. R., & Nichols, K. M. (2021). The Impacts of Dam Construction and Removal on the Genetics of Recovering Steelhead (Oncorhynchus mykiss) Populations across the Elwha River Watershed. Genes, 12(1), 89. https://doi.org/10.3390/genes12010089