Novel Brown Coat Color (Cocoa) in French Bulldogs Results from a Nonsense Variant in HPS3

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Selection
2.3. Whole Genome Sequencing
2.4. Variant Calling
2.5. Gene Analysis
2.6. Sanger Sequencing
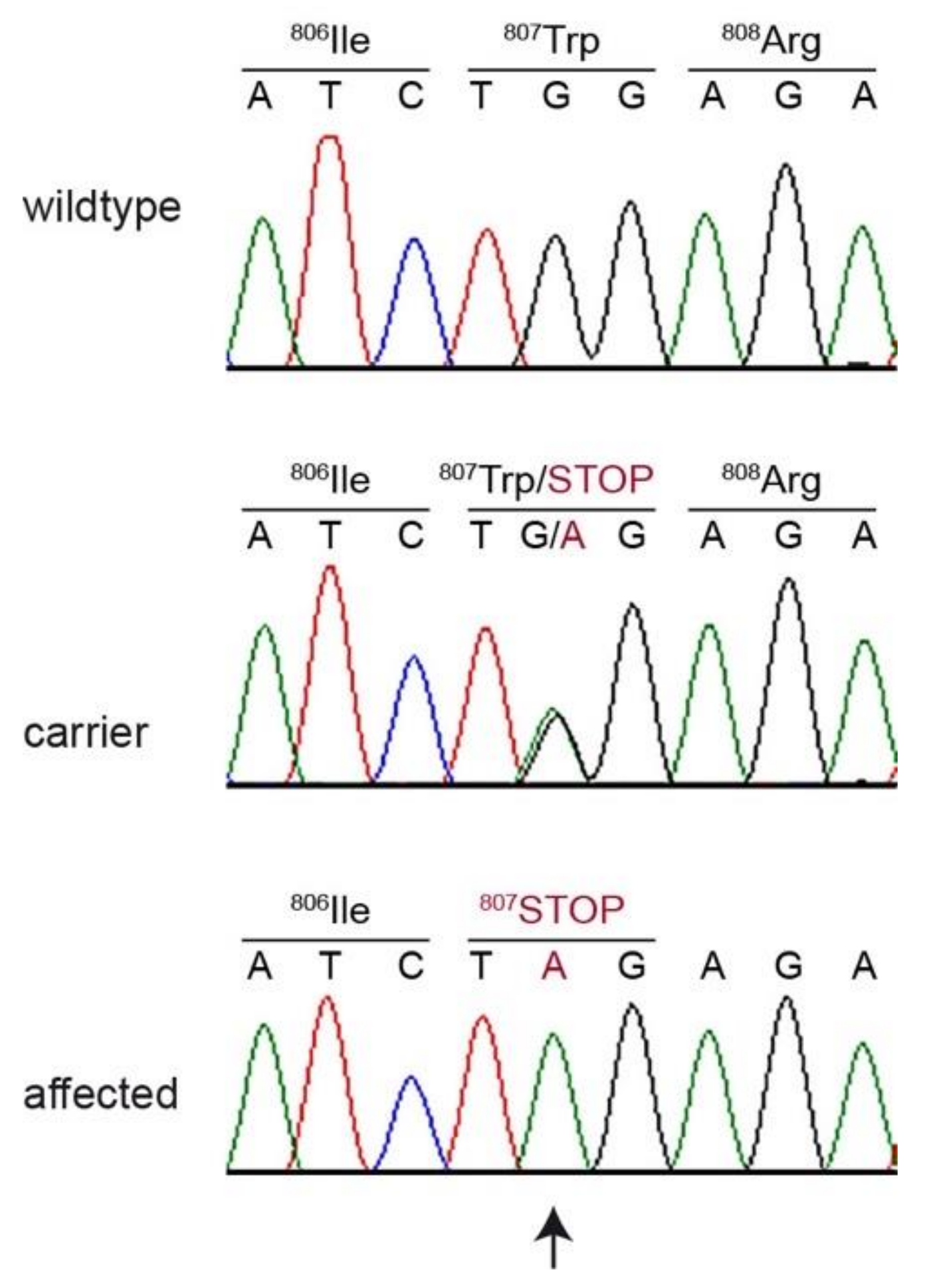
3. Results
3.1. Phenotype Characterization
3.2. Genetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jackson, I.J. A cDNA encoding tyrosinase-related protein maps to the brown locus in mouse. Proc. Natl. Acad. Sci. USA 1988, 85, 4392–4396. [Google Scholar] [CrossRef]
- Zdarsky, E.; Favor, J.; Jackson, I.J. The Molecular Basis of brown, an Old Mouse Mutation, and of an Induced Revertant to Wild. Genetics 1990, 126, 443–449. [Google Scholar]
- Lai, X.; Wichers, H.J.; Soler-Lopez, M.; Dijkstra, B.W. Structure and Function of Human Tyrosinase and Tyrosinase-Related Proteins. Chemistry 2018, 24, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Manga, P.; Kromberg, J.G.R.; Box, N.F.; Sturm, R.A.; Jenkins, T.; Ramsay, M. Rufous oculocutaneous albinism in Southern African blacks is caused by mutations in the TYRP1 gene. Am. J. Hum. Genet. 1997, 61, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Lyons, L.A.; Foe, I.T.; Rah, H.C.; Grahn, R.A. Chocolate coated cats: TYRP1 mutations for brown color in domestic cats. Mamm. Genome 2005, 16, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Berryere, T.G.; Schmutz, S.M.; Schimpf, R.J.; Cowan, C.M.; Potter, J. TYRP1 is associated with dun coat colour in Dexter cattle or how now brown cow? Anim. Genet. 2003, 34, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Bed’hom, B.; Marthey, S.; Valade, M.; Dureux, A.; Moroldo, M.; Péchoux, C.; Coville, J.L.; Gourichon, D.; Vieaud, A.; et al. A missense mutation in TYRP1 causes the chocolate plumage color in chicken and alters melanosome structure. Pigment Cell Melanoma Res. 2019, 32, 381–390. [Google Scholar] [CrossRef]
- Becker, D.; Otto, M.; Ammann, P.; Keller, I.; Drögemüller, C.; Leeb, T. The brown coat colour of Coppernecked goats is associated with a non-synonymous variant at the TYRP1 locus on chromosome 8. Anim. Genet. 2015, 46, 50–54. [Google Scholar] [CrossRef]
- Shibahara, S.; Tomita, Y.; Yoshizawa, M.; Shibata, K.; Tagami, H. Identification of Mutations in the Pigment Cell-Specific Gene Located at the Brown Locus in Mouse. Pigment Cell Res. 2008, 3, 90–95. [Google Scholar] [CrossRef]
- Cirera, S.; Markakis, M.N.; Kristiansen, T.; Vissenberg, K.; Fredholm, M.; Christensen, K.; Anistoroaei, R. A large insertion in intron 2 of the TYRP1 gene associated with American Palomino phenotype in American mink. Mamm. Genome 2016, 27, 135–143. [Google Scholar] [CrossRef]
- Ren, J.; Mao, H.; Zhang, Z.; Xiao, S.; Ding, N.; Huang, L. A 6-bp deletion in the TYRP1 gene causes the brown colouration phenotype in Chinese indigenous pigs. Heredity 2011, 106, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, N.J.; Mundy, N.I.; Gourichon, D.; Minvielle, F. Association of a single-nucleotide substitution in TYRP1 with roux in Japanese quail (Coturnix japonica). Anim. Genet. 2007, 38, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Utzeri, V.J.; Ribani, A.; Fontanesi, L. A premature stop codon in the TYRP1 gene is associated with brown coat colour in the European rabbit (Oryctolagus cuniculus). Anim. Genet. 2014, 45, 600–603. [Google Scholar] [CrossRef] [PubMed]
- Gratten, J.; Beraldi, D.; Lowder, B.V.; McRae, A.F.; Visscher, P.M.; Pemberton, J.M.; Slate, J. Compelling evidence that a single nucleotide substitution in TYRP1 is responsible for coat-colour polymorphism in a free-living population of Soay sheep. Proc. R. Soc. B Biol. Sci. 2007, 274, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, S.M.; Berryere, T.G.; Goldfinch, A.D. TYRP1 and MC1R genotypes and their effects on coat color in dogs. Mamm. Genome 2002, 13, 380–387. [Google Scholar] [CrossRef]
- Hrckova Turnova, E.; Majchrakova, Z.; Bielikova, M.; Soltys, K.; Turna, J.; Dudas, A. A novel mutation in the TYRP1 gene associated with brown coat colour in the Australian Shepherd Dog Breed. Anim. Genet. 2017, 48, 626. [Google Scholar] [CrossRef]
- Wright, H.E.; Schofield, E.; Mellersh, C.S.; Burmeister, L.M. A novel TYRP1 variant is associated with liver and tan coat colour in Lancashire Heelers. Anim. Genet. 2019, 50, 783. [Google Scholar] [CrossRef]
- Schmutz, S.M.; Melekhovets, Y. Coat color DNA testing in dogs: Theory meets practice. Mol. Cell. Probes 2012, 26, 238–242. [Google Scholar] [CrossRef]
- Caduff, M.; Bauer, A.; Jagannathan, V.; Leeb, T. OCA2 splice site variant in German Spitz dogs with oculocutaneous albinism. PLoS ONE 2017, 12, e0185944. [Google Scholar] [CrossRef]
- Bauer, A.; Jagannathan, V.; Högler, S.; Richter, B.; McEwan, N.A.; Thomas, A.; Cadieu, E.; André, C.; Hytönen, M.K.; Lohi, H.; et al. MKLN1 splicing defect in dogs with lethal acrodermatitis. PLoS Genet. 2018, 14, e1007264. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate long-read alignments with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMTOOLS. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Picard Homepage. Available online: https://broadinstitute.github.io/picard/ (accessed on 6 February 2020).
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- Anikster, Y.; Huizing, M.; White, J.; Shevchenko, Y.O.; Fitzpatrick, D.L.; Touchman, J.W.; Compton, J.G.; Bale, S.J.; Swank, R.T.; Gahl, W.A.; et al. Mutation of a new gene causes a unique form of Hermansky-Pudlak syndrome in a genetic isolate of central Puerto Rico. Nat. Genet. 2001, 28, 376–380. [Google Scholar] [CrossRef]
- Hermansky, F.; Pudlak, P. Albinism associated with hemorrhagic diathesis and unusual pigmented reticular cells in the bone marrow: Report of two cases with histochemical studies. Blood 1959, 14, 162–169. [Google Scholar] [CrossRef]
- Suzuki, T.; Li, W.; Zhang, Q.; Novak, E.K.; Sviderskaya, E.; Wilson, A.; Bennett, D.C.; Roe, B.A.; Swank, R.T.; Spritz, R.A. The gene mutated in cocoa mice, carrying a defect of organelle biogenesis, is a homologue of the human hermansky-pudlak syndrome-3 gene. Genomics 2001, 78, 30–37. [Google Scholar] [CrossRef]
- Novak, E.K.; Sweet, H.O.; Prochazka, M.; Parentis, M.; Soble, R.; Reddington, M.; Cairo, A.; Swank, R.T. Cocoa: A new mouse model for platelet storage pool deficiency. Br. J. Haematol. 1988, 69, 371–378. [Google Scholar] [CrossRef]
- Gautam, R.; Chintala, S.; Li, W.; Zhang, Q.; Tan, J.; Novak, E.K.; di Pietro, S.M.; Dell’Angelica, E.C.; Swank, R.T. The Hermansky-Pudlak Syndrome 3 (Cocoa) Protein Is a Component of the Biogenesis of Lysosome-related Organelles Complex-2 (BLOC-2). J. Biol. Chem. 2004, 279, 12935–12942. [Google Scholar] [CrossRef]
- Di Pietro, S.M.; Falcón-Pérez, J.M.; Dell’Angelica, E.C. Characterization of BLOC-2, a Complex Containing the Hermansky-Pudlak Syndrome Proteins HPS3, HPS5 and HPS6. Traffic 2004, 5, 276–283. [Google Scholar] [CrossRef]
- Bultema, J.J.; Ambrosio, A.L.; Burek, C.L.; di Pietro, S.M. BLOC-2, AP-3, and AP-1 proteins function in concert with Rab38 and Rab32 proteins to mediate protein trafficking to lysosome-related organelles. J. Biol. Chem. 2012, 287, 19550–19563. [Google Scholar] [CrossRef]
- Raposo, G.; Marks, M.S. The dark side of lysosome-related organelles: Specialization of the endocytic pathway for melanosome biogenesis. Traffic 2002, 3, 237–248. [Google Scholar] [CrossRef]
- Nguyen, T.; Novak, E.K.; Kermani, M.; Fluhr, J.; Peters, L.L.; Swank, R.T.; Wei, M.L. Melanosome morphologies in murine models of Hermansky-Pudlak syndrome reflect blocks in organelle development. J. Investig. Dermatol. 2002, 119, 1156–1164. [Google Scholar] [CrossRef][Green Version]
- Di Pietro, S.M.; Falcón-Pérez, J.M.; Tenza, D.; Setty, S.R.G.; Marks, M.S.; Raposo, G.; Dell’Angelica, E.C. BLOC-1 interacts with BLOC-2 and the AP-3 complex to facilitate protein trafficking on endosomes. Mol. Biol. Cell 2006, 17, 4027–4038. [Google Scholar] [CrossRef]
- Raposo, G.; Marks, M.S. Melanosomes—Dark organelles enlighten endosomal membrane transport. Nat. Rev. Mol. Cell Biol. 2007, 8, 786–797. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Filtering Step | Homozygous Variants | Heterozygous Variants |
|---|---|---|
| all variants | 2,571,692 | 3,132,757 |
| private variants | 694 | 5483 |
| protein-changing private variants | 2 | 48 |
| Dogs | G/G | G/A | A/A |
|---|---|---|---|
| Cases (brown or lilac French Bulldogs; n = 48) 1 | 1 | 0 | 47 |
| Controls (black or blue French Bulldogs; n = 111) 2 | 69 | 42 | 0 |
| French Bulldogs with other or unknown coat colors (n = 96) 3 | 61 | 24 | 11 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiener, S.; Kehl, A.; Loechel, R.; Langbein-Detsch, I.; Müller, E.; Bannasch, D.; Jagannathan, V.; Leeb, T. Novel Brown Coat Color (Cocoa) in French Bulldogs Results from a Nonsense Variant in HPS3. Genes 2020, 11, 636. https://doi.org/10.3390/genes11060636
Kiener S, Kehl A, Loechel R, Langbein-Detsch I, Müller E, Bannasch D, Jagannathan V, Leeb T. Novel Brown Coat Color (Cocoa) in French Bulldogs Results from a Nonsense Variant in HPS3. Genes. 2020; 11(6):636. https://doi.org/10.3390/genes11060636
Chicago/Turabian StyleKiener, Sarah, Alexandra Kehl, Robert Loechel, Ines Langbein-Detsch, Elisabeth Müller, Danika Bannasch, Vidhya Jagannathan, and Tosso Leeb. 2020. "Novel Brown Coat Color (Cocoa) in French Bulldogs Results from a Nonsense Variant in HPS3" Genes 11, no. 6: 636. https://doi.org/10.3390/genes11060636
APA StyleKiener, S., Kehl, A., Loechel, R., Langbein-Detsch, I., Müller, E., Bannasch, D., Jagannathan, V., & Leeb, T. (2020). Novel Brown Coat Color (Cocoa) in French Bulldogs Results from a Nonsense Variant in HPS3. Genes, 11(6), 636. https://doi.org/10.3390/genes11060636