Engineering CatM, a LysR-Type Transcriptional Regulator, to Respond Synergistically to Two Effectors


Abstract
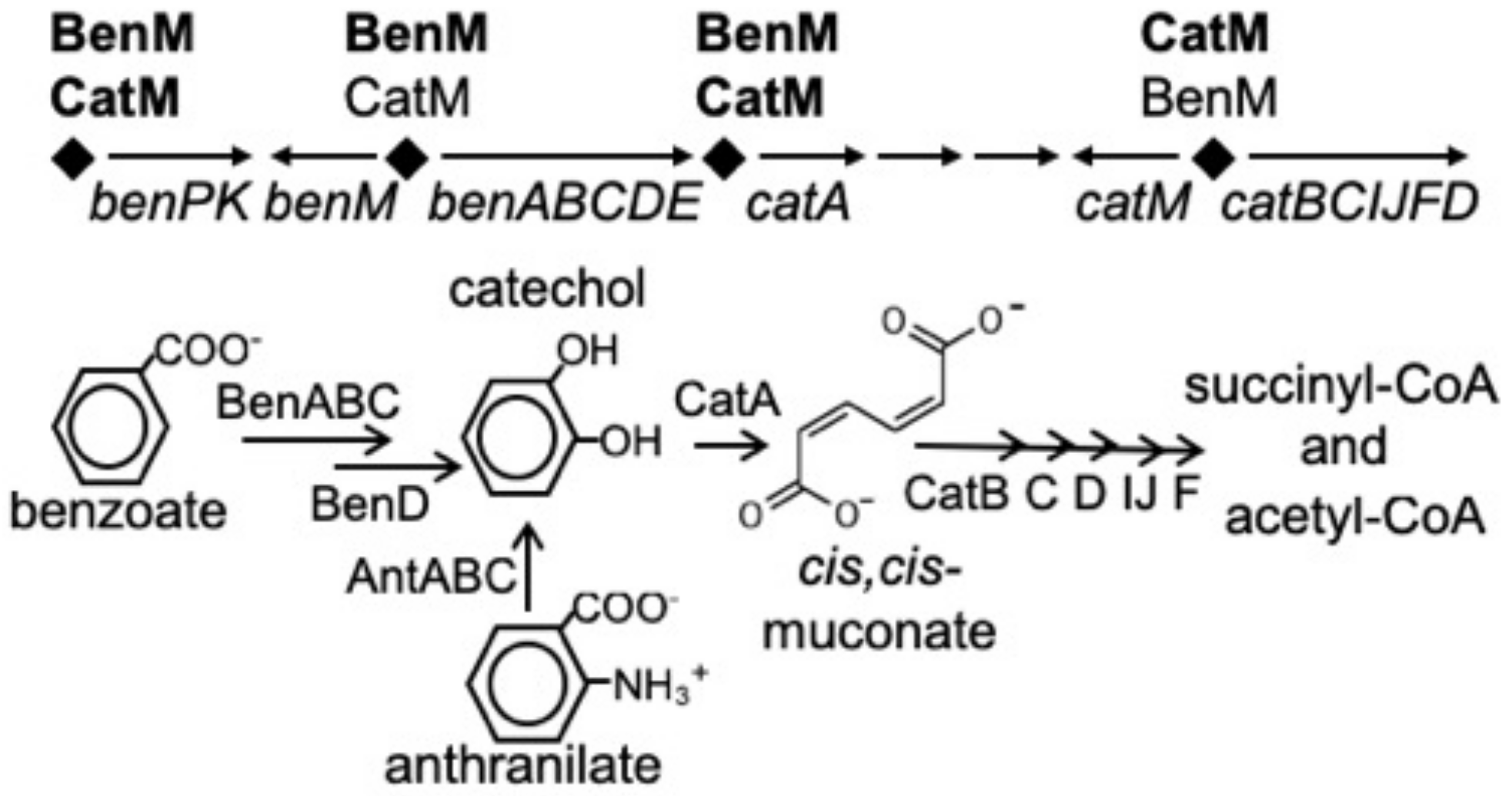
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Site-Directed Mutagenesis and Strain Construction by Allelic Replacement
2.3. Selection for BenM-Independent Growth on Benzoate
2.4. β-Galactosidase (LacZ) Assays
2.5. Purification of BenM and CatM and Variant Proteins
2.6. Electrophoretic Mobility Shift Assays (EMSAs)
2.7. Modeling the Transcription Activation Complex at PbenA and PcatB
3. Results
3.1. Engineered CatM Variants: Amino Acid Replacements at Positions 160 and 293
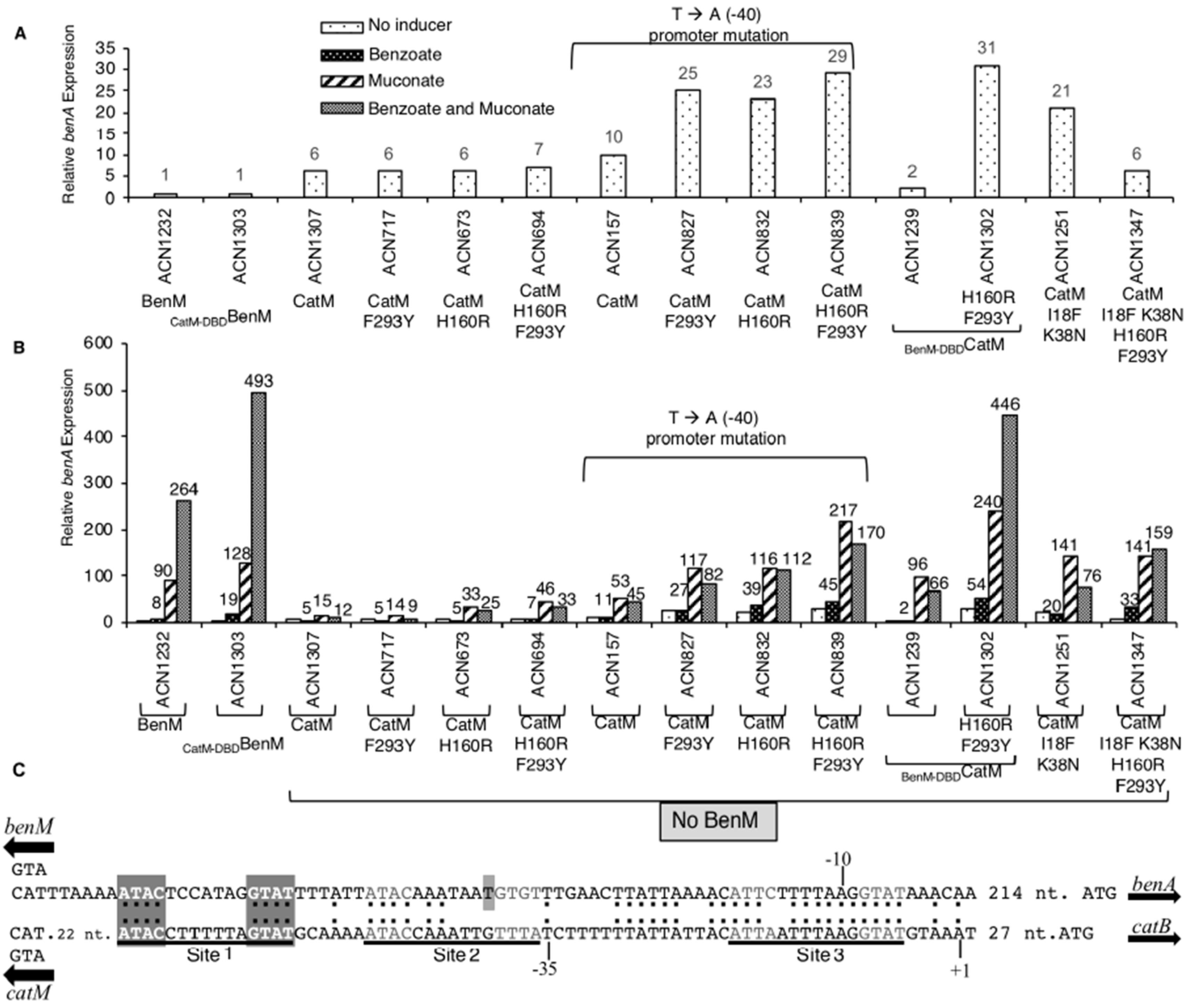
3.2. Transcriptional Regulation of PbenA by CatM Variants
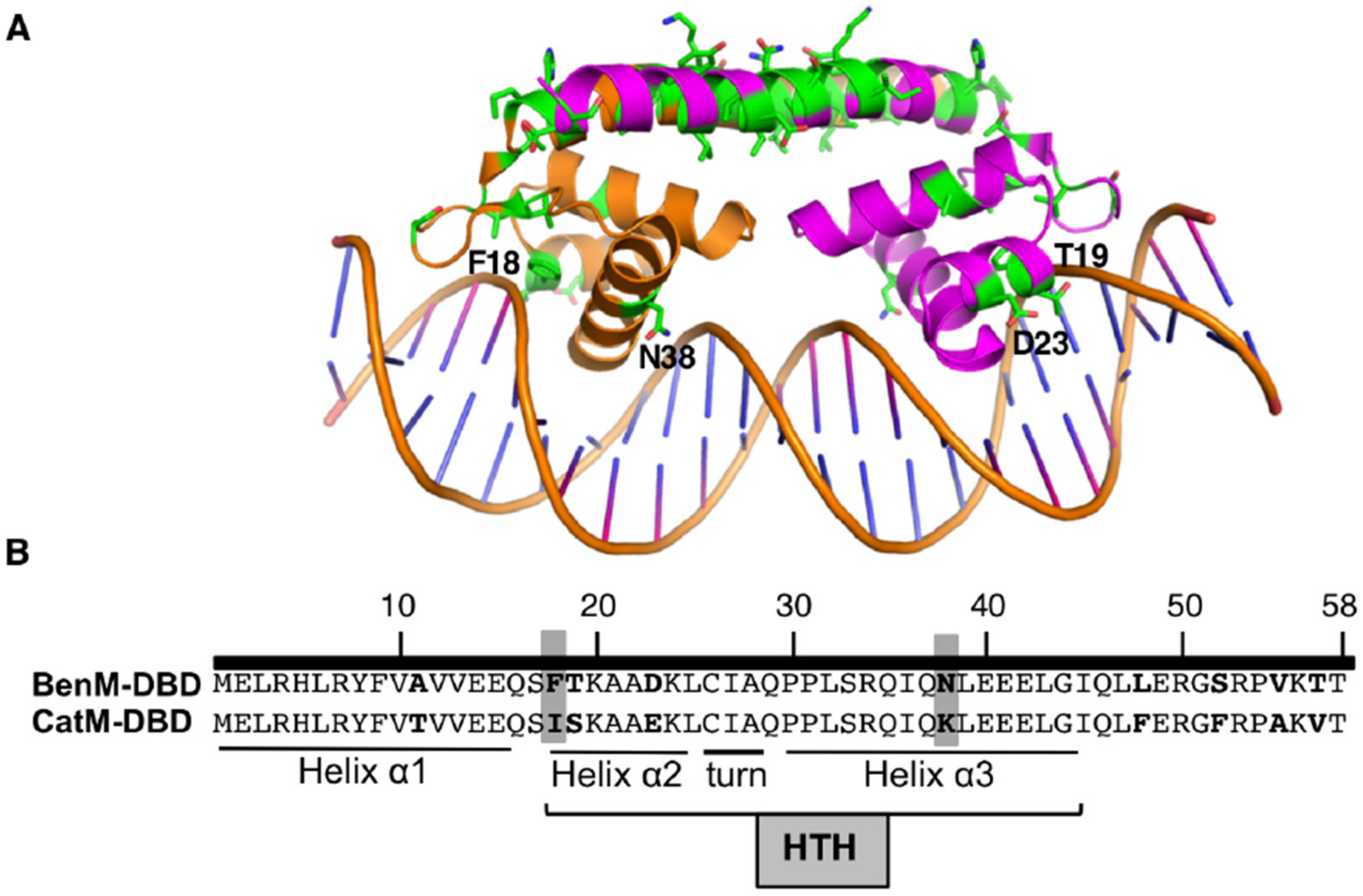
3.3. Spontaneous Mutant with Increased ben-Gene Expression: Changesne in the CatM-DBD
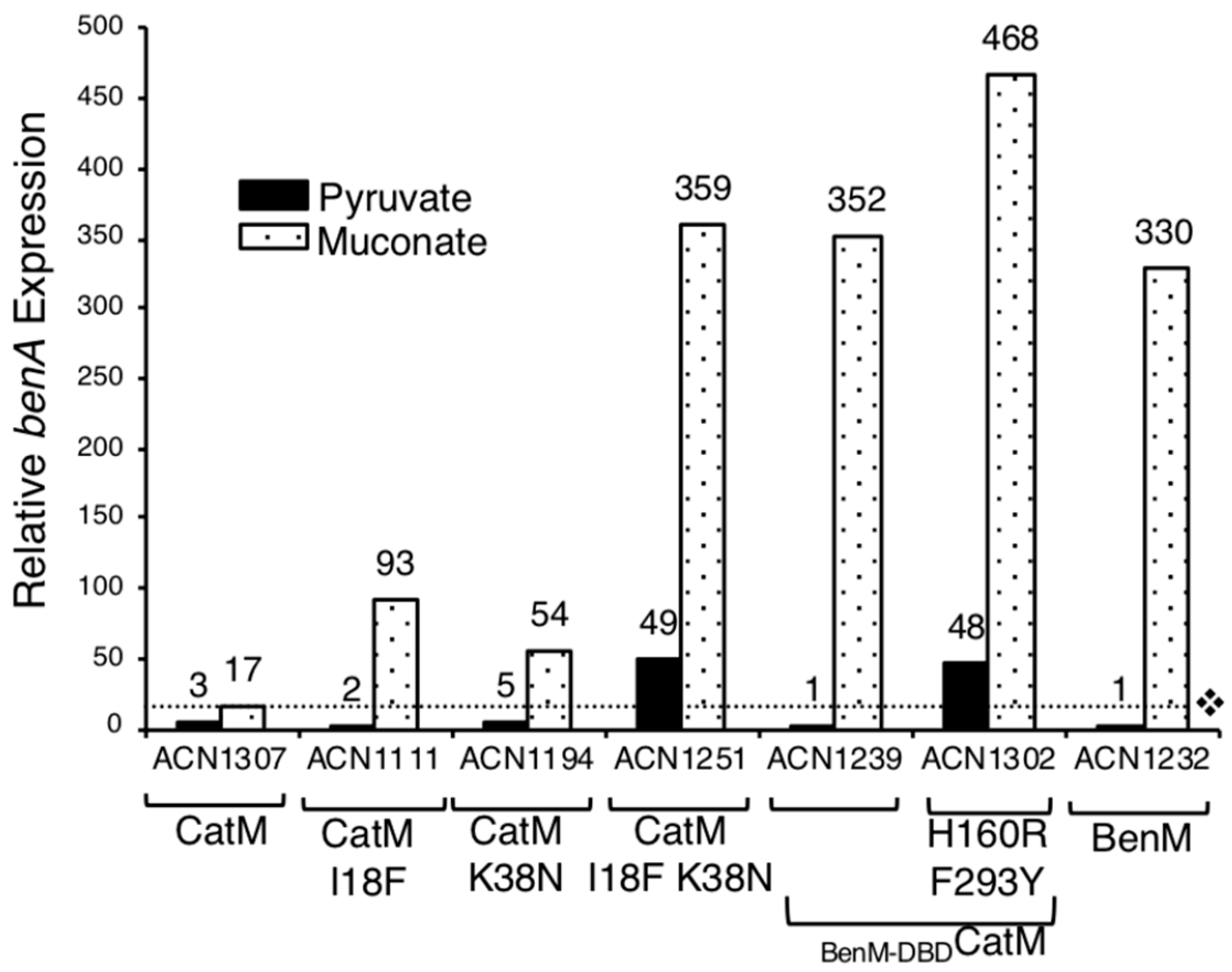
3.4. Further Investigation of DBD Residues in BenM and CatM
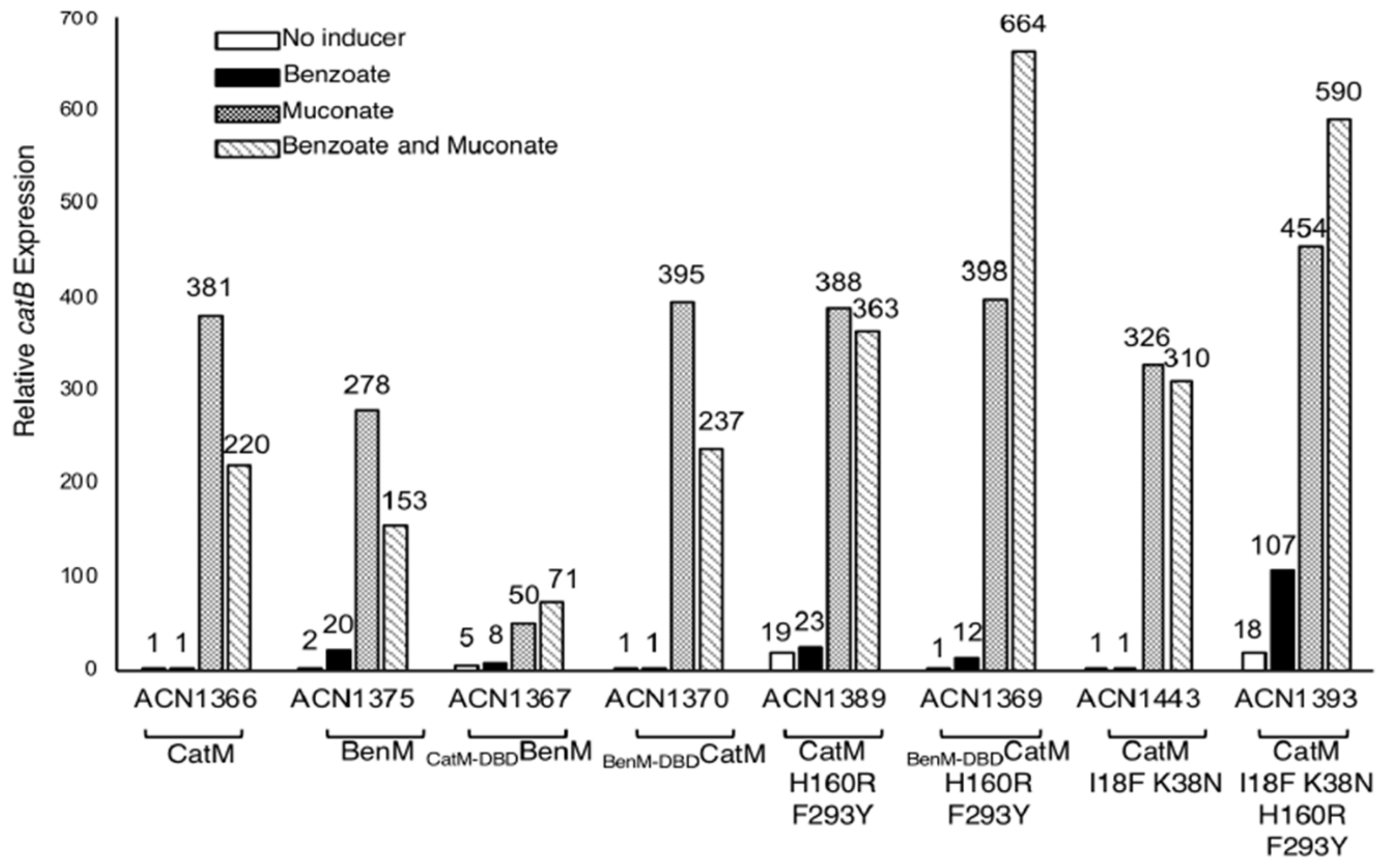
3.5. Benzoate-Responsive CatM Variants Obtained with Combinations of DBD and EBD Changes
3.6. Promoter Specificity and Regulator-DNA Binding Affinities
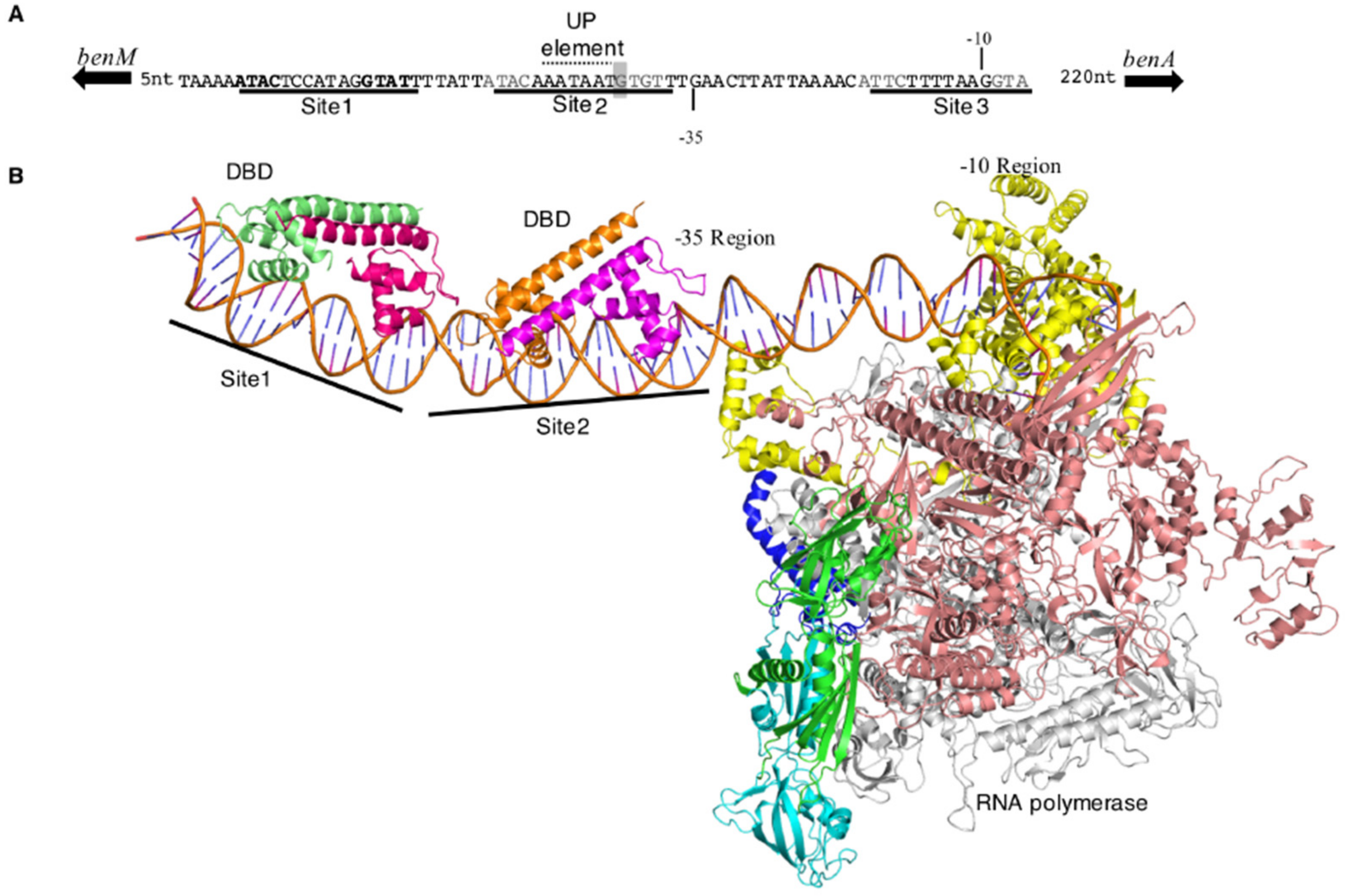
3.7. Regulator-DNA Interactions at PbenA: A Structural Model with RNAP
4. Discussion
4.1. Comparisons of BenM and CatM Provide a Framework to Engineer Effector-Binding Changes in CatM
4.2. Complex Patterns of Regulation
4.3. Interactions Between DBDs and Operator–Promoter Regions of PbenA and PbenA5146
4.4. Interactions Between DBDs and Operator–Promoter Regions of PcatB
4.5. A Model of PbenA and PcatB Promoters with RNAP and the DBDs of BenM and CatM
4.6. Broader Implications and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Craven, S.H.; Ezezika, O.C.; Momany, C.; Neidle, E.L. LysR homologs in Acinetobacter: Insights into a diverse and prevalent family of transcriptional regulators. In Acinetobacter Molecular Biology; Gerischer, U., Ed.; Caister Academic Press: Norfolk, UK, 2008; pp. 163–202. [Google Scholar]
- Bundy, B.M.; Collier, L.S.; Hoover, T.R.; Neidle, E.L. Synergistic transcriptional activation by one regulatory protein in response to two metabolites. Proc. Natl. Acad. Sci. USA 2002, 99, 7693–7698. [Google Scholar] [CrossRef]
- Collier, L.S.; Gaines, G.L., 3rd; Neidle, E.L. Regulation of benzoate degradation in Acinetobacter sp. strain ADP1 by BenM, a LysR-type transcriptional activator. J. Bacteriol. 1998, 180, 2493–2501. [Google Scholar]
- Ezezika, O.C.; Collier-Hyams, L.S.; Dale, H.A.; Burk, A.C.; Neidle, E.L. CatM regulation of the benABCDE operon: functional divergence of two LysR-type paralogs in Acinetobacter baylyi ADP1. Appl. Environ. Microbiol. 2006, 72, 1749–1758. [Google Scholar] [CrossRef]
- Craven, S.H.; Ezezika, O.C.; Haddad, S.; Hall, R.A.; Momany, C.; Neidle, E.L. Inducer responses of BenM, a LysR-type transcriptional regulator from Acinetobacter baylyi ADP1. Mol. Microbiol. 2009, 72, 881–894. [Google Scholar] [CrossRef] [PubMed]
- Ezezika, O.; Haddad, S.; Clark, T.; Neidle, E.; Momany, C. Distinct effector-binding sites enable synergistic transcriptional activation by BenM, a LysR-type regulator. J. Mol. Biol. 2007, 367, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, S.E.; Oyston, P.C. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef]
- Momany, C.; Neidle, E.L. Defying stereotypes: the elusive search for a universal model of LysR-type regulation. Mol. Microbiol. 2012, 83, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Quiocho, F.A.; Ledvina, P.S. Atomic structure and specificity of bacterial periplasmic receptors for active transport and chemotaxis: Variation of common themes. Mol. Microbiol. 1996, 20, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.A. Molecular biology of the LysR family of transcriptional regulators. Annu. Rev. Microbiol. 1993, 47, 597–626. [Google Scholar] [CrossRef]
- Juni, E.; Janik, A. Transformation of Acinetobacter calco-aceticus (Bacterium anitratum). J. Bacteriol. 1969, 98, 281–288. [Google Scholar]
- Vaneechoutte, M.; Young, D.M.; Ornston, L.N.; De Baere, T.; Nemec, A.; Van Der Reijden, T.; Carr, E.; Tjernberg, I.; Dijkshoorn, L. Naturally transformable Acinetobacter sp. strain ADP1 belongs to the newly described species Acinetobacter baylyi. Appl. Environ. Microbiol. 2006, 72, 932–936. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, F.; Maniatis, T. Molecular cloning: A laboratory manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Shanley, M.S.; Neidle, E.L.; Parales, R.E.; Ornston, L.N. Cloning and expression of Acinetobacter calcoaceticus catBCDE genes in Pseudomonas putida and Escherichia coli. J. Bacteriol. 1986, 165, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Bedore, S.R.; Sharma, N.K.; Lee, S.A.; Eiteman, M.A.; Neidle, E.L. Removal of aromatic inhibitors produced from lignocellulosic hydrolysates by Acinetobacter baylyi ADP1 with formation of ethanol by Kluyveromyces marxianus. Biotechnol. Biofuels. 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Horton, R.M.; Cai, Z.L.; Ho, S.N.; Pease, L.R. Gene splicing by overlap extension: tailor-made genes using the polymerase chain reaction. Biotechniques 1990, 8, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Neidle, E.L.; Hartnett, C.; Ornston, L.N. Characterization of Acinetobacter calcoaceticus catM, a repressor gene homologous in sequence to transcriptional activator genes. J. Bacteriol. 1989, 171, 5410–5421. [Google Scholar] [CrossRef] [PubMed]
- Tumen-Velasquez, M.; Johnson, C.W.; Ahmed, A.; Dominick, G.; Fulk, E.M.; Khanna, P.; Lee, S.A.; Schmidt, A.L.; Linger, J.G.; Eiteman, M.A.; et al. Accelerating pathway evolution by increasing the gene dosage of chromosomal segments. Proc. Natl. Acad. Sci. USA 2018, 115, 7105–7110. [Google Scholar] [CrossRef]
- Gregg-Jolly, L.A.; Ornston, L.N. Recovery of DNA from the Acinetobacter calcoaceticus chromosome by gap repair. J. Bacteriol. 1990, 172, 6169–6172. [Google Scholar] [CrossRef] [PubMed]
- Ruangprasert, A.; Craven, S.H.; Neidle, E.L.; Momany, C. Full-length structures of BenM and two variants reveal different oligomerization schemes for LysR-type transcriptional regulators. J. Mol. Biol. 2010, 404, 568–586. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Swillens, S. Interpretation of binding curves obtained with high receptor concentrations: Practical aid for computer analysis. Mol. Pharmacol. 1995, 47, 1197–1203. [Google Scholar]
- Alanazi, A.M.; Neidle, E.L.; Momany, C. The DNA-binding domain of BenM reveals the structural basis for the recognition of a T-N11-A sequence motif by LysR-type transcriptional regulators. Acta Crystallogr. D. Biol. Crystallogr. 2013, 69, 1995–2007. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Steitz, T.A. Crystal structures of the E. coli transcription initiation complexes with a complete bubble. Mol. Cell. 2015, 58, 534–540. [Google Scholar] [CrossRef]
- Lu, X.J.; Olson, W.K. 3DNA: a software package for the analysis, rebuilding and visualization of three-dimensional nucleic acid structures. Nucleic. Acids. Res. 2003, 31, 5108–5121. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. The Pymol User’s Manual. Delano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Cosper, N.J.; Collier, L.S.; Clark, T.J.; Scott, R.A.; Neidle, E.L. Mutations in catB, the gene encoding muconate cycloisomerase, activate transcription of the distal ben genes and contribute to a complex regulatory circuit in Acinetobacter sp. strain ADP1. J. Bacteriol. 2000, 182, 7044–7052. [Google Scholar] [CrossRef] [PubMed]
- Busby, S.; Ebright, R.H. Transcription activation at class II CAP-dependent promoters. Mol. Microbiol. 1997, 23, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, L.T.; Stauffer, G.V. GcvA interacts with both the alpha and sigma subunits of RNA polymerase to activate the Escherichia coli gcvB gene and the gcvTHP operon. FEMS Microbiol. Lett. 2005, 242, 333–338. [Google Scholar] [CrossRef]
- Ross, W.; Gosink, K.K.; Salomon, J.; Igarashi, K.; Zou, C.; Ishihama, A.; Severinov, K.; Gourse, R.L. A third recognition element in bacterial promoters: DNA binding by the alpha subunit of RNA polymerase. Science 1993, 262, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Koentjoro, M.P.; Adachi, N.; Senda, M.; Ogawa, N.; Senda, T. Crystal structure of the DNA-binding domain of the LysR-type transcriptional regulator CbnR in complex with a DNA fragment of the recognition-binding site in the promoter region. FEBS J. 2018, 285, 977–989. [Google Scholar] [CrossRef]
- Punekar, A.S.; Porter, J.; Carr, S.B.; Phillips, S.E. Structural basis for DNA recognition by the transcription regulator MetR. Acta Crystallogr. F. Struct. Biol. Commun. 2016, 72, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Lerche, M.; Dian, C.; Round, A.; Lonneborg, R.; Brzezinski, P.; Leonard, G.A. The solution configurations of inactive and activated DntR have implications for the sliding dimer mechanism of LysR transcription factors. Sci. Rep. 2016, 6, 19988. [Google Scholar] [CrossRef]
- Monferrer, D.; Tralau, T.; Kertesz, M.A.; Dix, I.; Sola, M.; Uson, I. Structural studies on the full-length LysR-type regulator TsaR from Comamonas testosteroni T-2 reveal a novel open conformation of the tetrameric LTTR fold. Mol. Microbiol. 2010, 75, 1199–1214. [Google Scholar] [CrossRef]
- Lonetto, M.A.; Rhodius, V.; Lamberg, K.; Kiley, P.; Busby, S.; Gross, C. Identification of a contact site for different transcription activators in region 4 of the Escherichia coli RNA polymerase sigma70 subunit. J. Mol. Biol. 1998, 284, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.K.; Bingen, J.M.; Johnson, C.W.; Kern, T.L.; Khanna, P.; Trettel, D.S.; Strauss, C.E.; Beckham, G.T.; Dale, T. A protocatechuate biosensor for Pseudomonas putida KT2440 via promoter and protein evolution. Metab. Eng. Commun. 2018, 6, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Juárez, J.F.; Lecube-Azpeitia, B.; Brown, S.L.; Johnston, C.D.; Church, G.M. Biosensor libraries harness large classes of binding domains for construction of allosteric transcriptional regulators. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant Characteristics | Generation Time (min) b | Lag Time (h) c |
|---|---|---|---|
| ADP1 | Wild-type | 70 ± 5 | 4.5 ± 1 |
| ISA36 | No BenM | No growth | No growth |
| ACN682 | No BenM, CatM(F293Y) | No growth | No growth |
| ACN1095 | No BenM, CatM(I18F) | 175 ± 3 | 18 ± 3 |
| ACN1193 | No BenM, CatM(K38N) | 145 ± 6 | 11 ± 1 |
| ACN1249 | No BenM, CatM(I18F,K38N) | 85 ± 2 | 5.5 ± 2 |
| ACN1234 | No BenM, BenM-DBDCatM | 82 ± 3 | 5 ± 1.5 |
| ACN1301 | No BenM, BenM-DBDCatM(F293Y,H160R) | 81 ± 4 | 5 ± 1 |
| ACN1345 | No BenM, CatM(I18F,K38N,H160R,F293Y) | 83 ± 3 | 5 ± 1.5 |
| ACN1294 | CatM-DBDBenM, No CatM | 186 ± 6 | 21 ± 3 |
| DNA | Transcriptional Regulator | Effector Added to Protein-DNA Complex | |||
|---|---|---|---|---|---|
| No Effector Kd (nM) | Benzoate Kd (nM) | Muconate Kd (nM) | Muconate and Benzoate Kd (nM) | ||
| PbenA | Wild-type BenM | 32 ± 3 | 17 ± 2 | 41 ± 3 | 46 ± 3 |
| Wild-type CatM | 70 ± 3 | 71 ± 4 | 67 ± 3 | 71 ± 4 | |
| BenM-DBDCatM | 2 ± 0.5 | 4 ± 1 | 3 ± 0.5 | 9 ± 3 | |
| BenM-DBDCatM(F293Y,H160R) | 1 ± 0.2 | 4 ± 1 | 2 ± 0.5 | 4 ± 2 | |
| CatM(18F,K38N) | 42 ± 2 | 32 ± 3 | 34 ± 1 | 32 ± 4 | |
| PcatB | Wild-type BenM | 26 ± 3 | 30 ± 2 | 16 ± 2 | 19 ± 1 |
| Wild-type CatM | 11 ± 2 | 19 ± 3 | 10 ± 1 | 19 ± 2 | |
| BenM-DBDCatM | 1 ± 0.2 | 3 ± 0.5 | 3 ± 0.2 | 4 ± 1 | |
| BenM-DBDCatM(F293Y,H160R) | 3 ± 0.5 | 3 ± 0.5 | 3 ± 0.5 | 3 ± 0.5 | |
| CatM(18F,K38N) | 28 ± 2 | 13 ± 2 | 8 ± 2 | 14 ± 0.5 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tumen-Velasquez, M.P.; Laniohan, N.S.; Momany, C.; Neidle, E.L. Engineering CatM, a LysR-Type Transcriptional Regulator, to Respond Synergistically to Two Effectors. Genes 2019, 10, 421. https://doi.org/10.3390/genes10060421
Tumen-Velasquez MP, Laniohan NS, Momany C, Neidle EL. Engineering CatM, a LysR-Type Transcriptional Regulator, to Respond Synergistically to Two Effectors. Genes. 2019; 10(6):421. https://doi.org/10.3390/genes10060421
Chicago/Turabian StyleTumen-Velasquez, Melissa P., Nicole S. Laniohan, Cory Momany, and Ellen L. Neidle. 2019. "Engineering CatM, a LysR-Type Transcriptional Regulator, to Respond Synergistically to Two Effectors" Genes 10, no. 6: 421. https://doi.org/10.3390/genes10060421
APA StyleTumen-Velasquez, M. P., Laniohan, N. S., Momany, C., & Neidle, E. L. (2019). Engineering CatM, a LysR-Type Transcriptional Regulator, to Respond Synergistically to Two Effectors. Genes, 10(6), 421. https://doi.org/10.3390/genes10060421