Can Genetic Testing Identify Talent for Sport?




{kind=link}
{kind=link}
Abstract
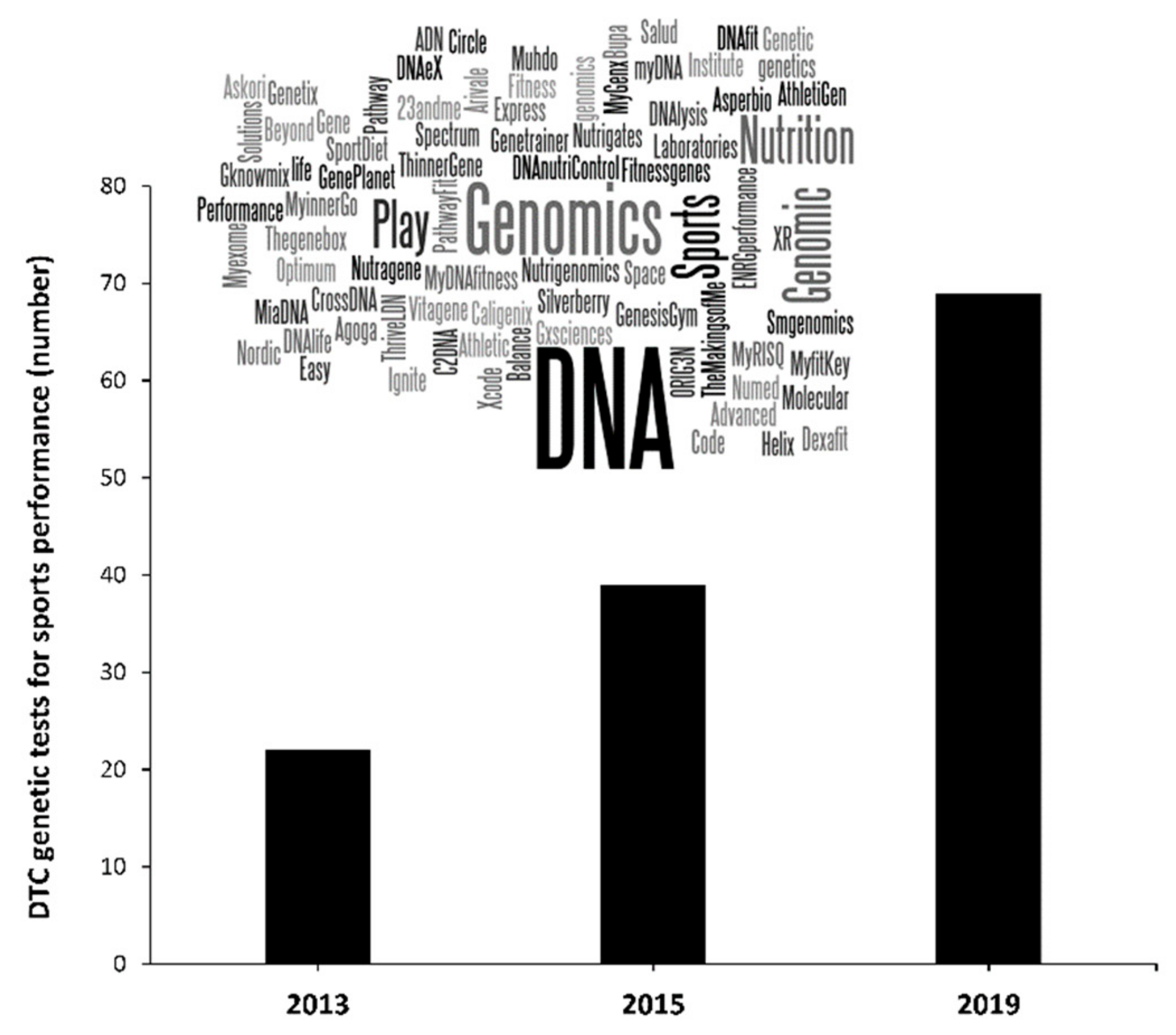
1. Introduction
2. Why We Cannot Currently Use Genetic Information for Talent ID
3. What Further Knowledge Do We Require to Potentially Use Genetic Information for Talent Identification?
3.1. Genome-Wide Association Studies
3.2. Rare Variants
3.3. A Predictive Threshold
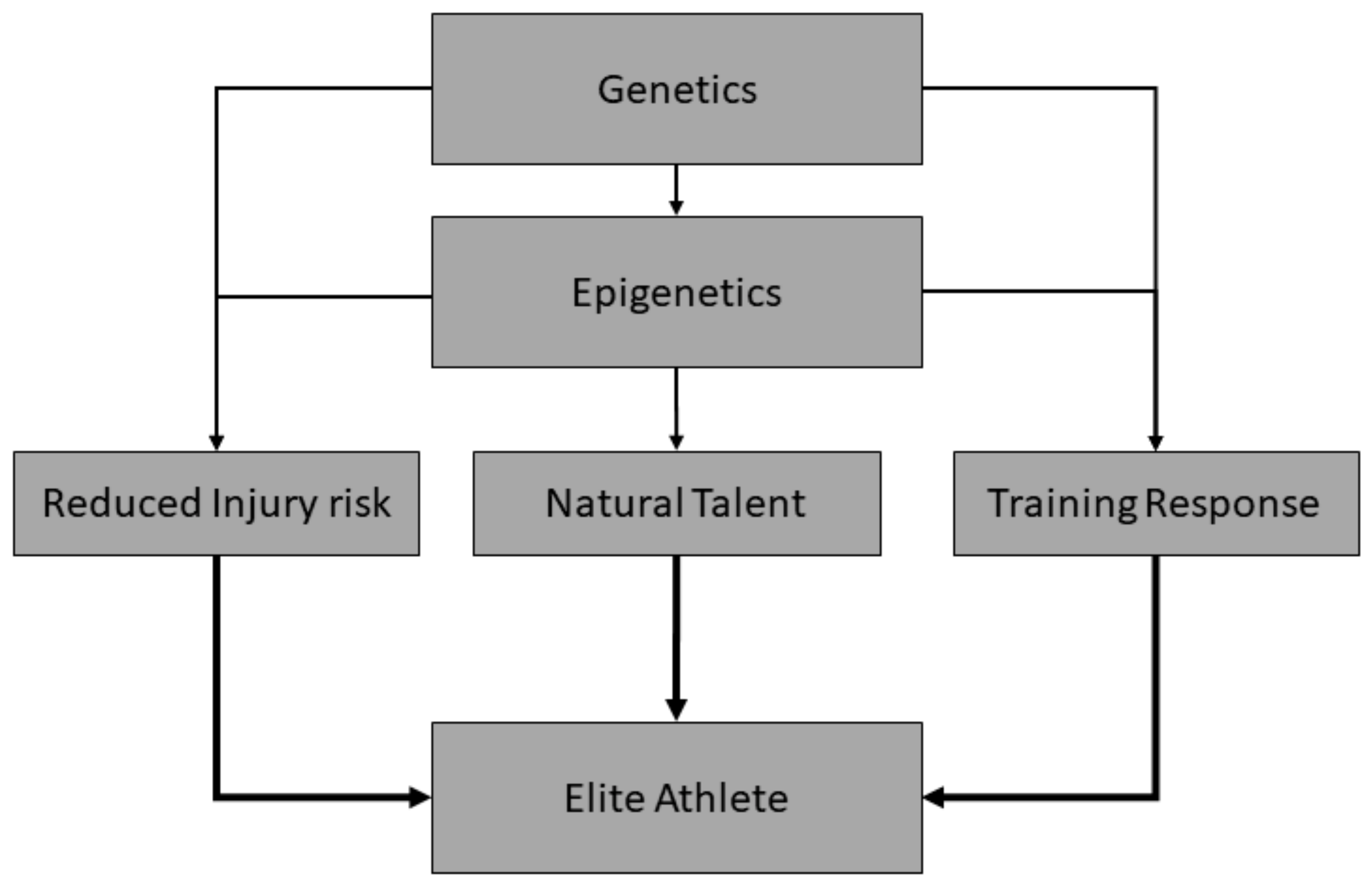
3.4. Epigenetic Modifications
3.5. Lessons from Disease Prediction
4. Even If We Could Test for Talent—Should We?
5. What Could We Potentially Use Genetic Testing for?
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Williams, A.M.; Reilly, T. Talent identification and development in soccer. J. Sports Sci. 2000, 18, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Moran, C.N.; Pitsiladis, Y.P. Tour de France Champions born or made: Where do we take the genetics of performance? J. Sports Sci. 2017, 35, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- De Moor, M.H.; Spector, T.D.; Cherkas, L.F.; Falchi, M.; Hottenga, J.J.; Boomsma, D.I.; De Geus, E.J. Genome-wide linkage scan for athlete status in 700 British female DZ twin pairs. Twin Res. Hum. Genet. 2007, 10, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Houweling, P.J.; Papadimitriou, I.D.; Seto, J.T.; Pérez, L.M.; Coso, J.D.; North, K.N.; Lucia, A.; Eynon, N. Is evolutionary loss our gain? The role of ACTN3 p. Arg577Ter (R577X) genotype in athletic performance, ageing, and disease. Hum. Mutat. 2018, 39, 1774–1787. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Yang, Y.; Li, X.; Zhou, F.; Gao, C.; Li, M.; Gao, L. The association of sport performance with ACE and ACTN3 genetic polymorphisms: A systematic review and meta-analysis. PLoS ONE 2013, 8, e54685. [Google Scholar] [CrossRef]
- Vincent, B.; De Bock, K.; Ramaekers, M.; Van den Eede, E.; Van Leemputte, M.; Hespel, P.; Thomis, M.A. ACTN3 (R577X) genotype is associated with fiber type distribution. Physiol. Genomics 2007, 32, 58–63. [Google Scholar] [CrossRef]
- Yang, N.; MacArthur, D.G.; Gulbin, J.P.; Hahn, A.G.; Beggs, A.H.; Easteal, S.; North, K. ACTN3 genotype is associated with human elite athletic performance. Am. J. Hum. Genet. 2003, 73, 627–631. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Druzhevskaya, A.M.; Lyubaeva, E.V.; Popov, D.V.; Vinogradova, O.L.; Williams, A.G. The dependence of preferred competitive racing distance on muscle fibre type composition and ACTN3 genotype in speed skaters. Exp. Physiol. 2011, 96, 1302–1310. [Google Scholar] [CrossRef]
- Eynon, N.; Sagiv, M.; Meckel, Y.; Duarte, J.A.; Alves, A.J.; Yamin, C.; Sagiv, M.; Goldhammer, E.; Oliveira, J. NRF2 intron 3 A/G polymorphism is associated with endurance athletes’ status. J. Appl. Physiol. 2009, 107, 76–79. [Google Scholar] [CrossRef]
- Scott, R.A.; Irving, R.; Irwin, L.; Morrison, E.; Charlton, V.; Austin, K.; Tladi, D.; Deason, M.; Headley, S.A.; Kolkhorst, F.W.; et al. ACTN3 and ACE genotypes in elite Jamaican and US sprinters. Med. Sci. Sports Exerc. 2010, 42, 107–112. [Google Scholar] [CrossRef]
- Moran, C.N.; Yang, N.; Bailey, M.E.; Tsiokanos, A.; Jamurtas, A.; MacArthur, D.G.; North, K.; Pitsiladis, Y.P.; Wilson, R.H. Association analysis of the ACTN3 R577X polymorphism and complex quantitative body composition and performance phenotypes in adolescent Greeks. Eur. J. Hum. Genet. 2007, 15, 88. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, I.D.; Lucia, A.; Pitsiladis, Y.P.; Pushkarev, V.P.; Dyatlov, D.A.; Orekhov, E.F.; Artioli, G.G.; Guilherme, J.P.; Lancha, A.H.; Ginevičienė, V.; et al. ACTN3 R577X and ACE I/D gene variants influence performance in elite sprinters: A multi-cohort study. BMC Genomics 2016, 17, 285. [Google Scholar] [CrossRef] [PubMed]
- Berman, Y.; North, K.N. A gene for speed: The emerging role of α-actinin-3 in muscle metabolism. Physiology 2010, 25, 250–259. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, D.G.; North, K.N. A gene for speed? The evolution and function of α-actinin-3. Bioessays 2004, 26, 786–795. [Google Scholar] [CrossRef]
- Lee, F.X.; Houweling, P.J.; North, K.N.; Quinlan, K.G. How does α-actinin-3 deficiency alter muscle function? Mechanistic insights into ACTN3, the ‘gene for speed’. Biochim. Biophys. Acta 2016, 1863, 686–693. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Egorova, E.S.; Gabdrakhmanova, L.J.; Fedotovskaya, O.N. Genes and athletic performance: An update. Med. Sports Sci. 2016, 61, 41–54. [Google Scholar]
- Silventoinen, K.; Sammalisto, S.; Perola, M.; Boomsma, D.I.; Cornes, B.K.; Davis, C.; Dunkel, L.; De Lange, M.; Harris, J.R.; Hjelmborg, J.V.; et al. Heritability of adult body height: A comparative study of twin cohorts in eight countries. Twin Res. 2003, 6, 399–408. [Google Scholar] [CrossRef]
- Elks, C.E.; Den Hoed, M.; Zhao, J.H.; Sharp, S.J.; Wareham, N.J.; Loos, R.J.; Ong, K.K. Variability in the heritability of body mass index: A systematic review and meta-regression. Front. Endocrinol. 2012, 3, 29. [Google Scholar] [CrossRef]
- Stein, M.B.; Fallin, M.D.; Schork, N.J.; Gelernter, J. COMT polymorphisms and anxiety-related personality traits. Neuropsychopharmacology 2005, 30, 2092. [Google Scholar] [CrossRef]
- Peplonska, B.; Safranow, K.; Adamczyk, J.; Boguszewski, D.; Szymański, K.; Soltyszewski, I.; Barczak, A.; Siewierski, M.; Ploski, R.; Sozanski, H.; et al. Association of serotoninergic pathway gene variants with elite athletic status in the Polish population. J. Sports Sci. 2019. [Google Scholar] [CrossRef]
- Webborn, N.; Williams, A.; McNamee, M.; Bouchard, C.; Pitsiladis, Y.; Ahmetov, I.; Ashley, E.; Byrne, N.; Camporesi, S.; Collins, M.; et al. Direct-to-consumer genetic testing for predicting sports performance and talent identification: Consensus statement. Br. J. Sports Med. 2015, 49, 1486–1491. [Google Scholar] [CrossRef] [PubMed]
- Camporesi, S.; McNamee, M.J. Ethics, genetic testing, and athletic talent: children’s best interests, and the right to an open (athletic) future. Physiol. Genomics 2016, 48, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.G.; Heffernan, S.M.; Day, S.H. Genetic testing in exercise and sport-have direct-to-consumer genetic tests come of age? Sci. Sport Curr. Trends 2014, 2, 3–9. [Google Scholar]
- Gayagay, G.; Yu, B.; Hambly, B.; Boston, T.; Hahn, A.; Celermajer, D.S.; Trent, R.J. Elite endurance athletes and the ACE I allele—The role of genes in athletic performance. Hum. Genet. 1998, 103, 48–50. [Google Scholar] [CrossRef]
- Lucia, A.; Gómez-Gallego, F.; Barroso, I.; Rabadán, M.; Bandrés, F.; San Juan, A.F.; Chicharro, J.L.; Ekelund, U.; Brage, S.; Earnest, C.P.; et al. PPARGC1 genotype (Gly482Ser) predicts exceptional endurance capacity in European males. J. Appl. Physiol. 2005, 99, 344–348. [Google Scholar] [CrossRef]
- Maciejewska, A.; Sawczuk, M.; Cieszczyk, P.; Mozhayskaya, I.A.; Ahmetov, I.I. The PPARGC1A gene Gly482Ser in Polish and Russian athletes. J. Sports Sci. 2012, 30, 101–113. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Popov, D.V.; Missina, S.S.; Vinogradova, O.L.; Rogozkin, V.A. Association of mitochondrial transcription factor (TFAM) gene polymorphism with physical performance in athletes. Hum. Physiol. 2010, 36, 229–233. [Google Scholar] [CrossRef]
- Williams, A.G.; Wackerhage, H.; Day, S.H. Genetic testing for sports performance, responses to training and injury risk: Practical and ethical considerations. Med. Sport Sci. 2016, 61, 105–119. [Google Scholar]
- Yang, N.; Macarthur, D.G.; Wolde, B.; Onywera, V.O.; Boit, M.; Lau, S.Y.; Wilson, R.H.; Scott, R.A.; Pitsiladis, Y.; North, K.N. The ACTN3 R577X polymorphism in East and West African athletes. Med. Sci. Sports Exerc. 2007, 39, 1985–1988. [Google Scholar] [CrossRef]
- Lucia, A.; Oliván, J.; Gómez-Gallego, F.; Santiago, C.; Montil, M.; Foster, C. Citius and longius (faster and longer) with no α-actinin-3 in skeletal muscles? Br. J. Sports Med. 2007, 41, 616–617. [Google Scholar] [CrossRef]
- Ruiz, J.R.; Gómez-Gallego, F.; Santiago, C.; González-Freire, M.; Verde, Z.; Foster, C.; Lucia, A. Is there an optimum endurance polygenic profile? J. Physiol. 2009, 587, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.R.; Arteta, D.; Buxens, A.; Artieda, M.; Gómez-Gallego, F.; Santiago, C.; Yvert, T.; Morán, M.; Lucia, A. Can we identify a power-oriented polygenic profile? J. Appl. Physiol. 2010, 108, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Santiago, C.; Ruiz, J.R.; Muniesa, C.A.; González-Freire, M.; Gómez-Gallego, F.; Lucia, A. Does the polygenic profile determine the potential for becoming a world-class athlete? Insights from the sport of rowing. Scand. J. Med. Sci. Sports 2010, 20, e188–e194. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.G.; Folland, J.P. Similarity of polygenic profiles limits the potential for elite human physical performance. J. Physiol. 2008, 586, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.C.; Day, S.H.; Ahmetov, I.I.; Williams, A.G. Genetics of muscle strength and power: Polygenic profile similarity limits skeletal muscle performance. J. Sports Sci. 2011, 29, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Rodas, G.; Osaba, L.; Arteta, D.; Pruna, R.; Fernández, D.; Lucia, A. Genomic prediction of tendinopathy risk in elite team sports. Int. J. Sports Physiol. Perform. 2019. [Google Scholar] [CrossRef] [PubMed]
- Rankinen, T.; Fuku, N.; Wolfarth, B.; Wang, G.; Sarzynski, M.A.; Alexeev, D.G.; Ahmetov, I.I.; Boulay, M.R.; Cieszczyk, P.; Eynon, N.; et al. No evidence of a common DNA variant profile specific to world class endurance athletes. PLoS ONE 2016, 11, e0147330. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Kulemin, N.A.; Popov, D.V.; Naumov, V.A.; Akimov, E.B.; Bravy, Y.R.; Egorova, E.S.; Galeeva, A.A.; Generozov, E.V.; Kostryukova, E.S.; et al. Genome-wide association study identifies three novel genetic markers associated with elite endurance performance. Biol. Sport 2015, 32, 3–9. [Google Scholar] [CrossRef]
- Pickering, C.; Suraci, B.; Semenova, E.A.; Boulygina, E.A.; Kostryukova, E.S.; Kulemin, N.A.; Borisov, O.V.; Khabibova, S.A.; Larin, A.K.; Pavlenko, A.V.; et al. A Genome-Wide Association Study of sprint performance in elite youth football players. J. Strength Cond. Res. 2019, 33, 2344–2351. [Google Scholar] [CrossRef]
- Juvonen, E.; Ikkala, E.; Fyhrquist, F.; Ruutu, T. Autosomal dominant erythrocytosis caused by increased sensitivity to erythropoietin. Blood 1991, 78, 3066–3069. [Google Scholar] [CrossRef]
- Waggott, D.; Mattsson, C.M.; Wheeler, M.; Ashley, E.A. The Genomics of Extreme Athletes. The ELITE Study (Exercise at the Limit-Inherited Traits of Endurance) [Abstract]. In Proceedings of the American Society of Human Genetics (ASHG) Annual Meeting, Vancouver, BC, Canada, 18–22 October 2016. [Google Scholar]
- Schuelke, M.; Wagner, K.R.; Stolz, L.E.; Hübner, C.; Riebel, T.; Kömen, W.; Braun, T.; Tobin, J.F.; Lee, S.J. Myostatin mutation associated with gross muscle hypertrophy in a child. N. Engl. J. Med. 2004, 350, 2682–2688. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Shi, L.; Hakenberg, J.; Naughton, B.; Sklar, P.; Zhang, J.; Zhou, H.; Tian, L.; Prakash, O.; Lemire, M.; et al. Analysis of 589,306 genomes identifies individuals resilient to severe Mendelian childhood diseases. Nat. Biotechnol. 2016, 34, 531. [Google Scholar] [CrossRef] [PubMed]
- Ehlert, T.; Simon, P.; Moser, D.A. Epigenetics in sports. Sports Med. 2013, 43, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Voisin, S.; Eynon, N.; Yan, X.; Bishop, D.J. Exercise training and DNA methylation in humans. Acta Physiol. 2015, 213, 39–59. [Google Scholar] [CrossRef]
- Ghosh, A.; Mahajan, P.B. Can genotype determine the sports phenotype? A paradigm shift in sports medicine. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 333–339. [Google Scholar] [CrossRef]
- Luo, W.; Nie, Q.; Zhang, X. MicroRNAs involved in skeletal muscle differentiation. J. Genet. Genomics 2013, 40, 107–116. [Google Scholar] [CrossRef]
- Davidsen, P.K.; Gallagher, I.J.; Hartman, J.W.; Tarnopolsky, M.A.; Dela, F.; Helge, J.W.; Timmons, J.A.; Phillips, S.M. High responders to resistance exercise training demonstrate differential regulation of skeletal muscle microRNA expression. J. Appl. Physiol. 2011, 110, 309–317. [Google Scholar] [CrossRef]
- Richards, E.J. Inherited epigenetic variation—Revisiting soft inheritance. Nat. Rev. Genet. 2006, 7, 395. [Google Scholar] [CrossRef]
- Lokk, K.; Modhukur, V.; Rajashekar, B.; Märtens, K.; Mägi, R.; Kolde, R.; Koltšina, M.; Nilsson, T.K.; Vilo, J.; Salumets, A.; et al. DNA methylome profiling of human tissues identifies global and tissue-specific methylation patterns. Genome Biol. 2014, 15, 324. [Google Scholar] [CrossRef]
- Pickering, C.; Kiely, J. The Development of a Personalised Training Framework: Implementation of Emerging Technologies for Performance. J. Funct. Morphol. Kinesiol. 2019, 4, 25. [Google Scholar] [CrossRef]
- Langie, S.A.; Moisse, M.; Declerck, K.; Koppen, G.; Godderis, L.; Vanden Berghe, W.; Drury, S.; De Boever, P. Salivary DNA methylation profiling: Aspects to consider for biomarker identification. Basic Clin. Pharmacol. Toxicol. 2017, 121, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, N.; Shi, J.; García-Closas, M. Developing and evaluating polygenic risk prediction models for stratified disease prevention. Nat. Rev. Genet. 2016, 17, 392. [Google Scholar] [CrossRef] [PubMed]
- Manolio, T.A.; Collins, F.S.; Cox, N.J.; Goldstein, D.B.; Hindorff, L.A.; Hunter, D.J.; McCarthy, M.I.; Ramos, E.M.; Cardon, L.R.; Chakravarti, A.; et al. Finding the missing heritability of complex diseases. Nature 2009, 461, 747. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Kichaev, G.; Pasaniuc, B. Contrasting the genetic architecture of 30 complex traits from summary association data. Am. J. Hum. Genet. 2016, 99, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Khera, A.V.; Chaffin, M.; Aragam, K.G.; Haas, M.E.; Roselli, C.; Choi, S.H.; Natarajan, P.; Lander, E.S.; Lubitz, S.A.; Ellinor, P.T.; et al. Genome-wide polygenic scores for common diseases identify individuals with risk equivalent to monogenic mutations. Nat. Genet. 2018, 50, 1219. [Google Scholar] [CrossRef]
- Dudbridge, F. Power and predictive accuracy of polygenic risk scores. PLoS Genet. 2013, 9, e1003348. [Google Scholar] [CrossRef]
- Janssens, A.C.; Joyner, M.J. Polygenic risk scores that predict common diseases using millions of single nucleotide polymorphisms: Is more, better? Clin. Chem. 2019. [Google Scholar] [CrossRef]
- Mattsson, C.M.; Wheeler, M.T.; Waggott, D.; Caleshu, C.; Ashley, E.A. Sports genetics moving forward: Lessons learned from medical research. Physiol. Genomics 2016, 48, 175–182. [Google Scholar] [CrossRef]
- Breitbach, S.; Tug, S.; Simon, P. Conventional and genetic talent identification in sports: Will recent developments trace talent? Sports Med. 2014, 44, 1489–1503. [Google Scholar] [CrossRef]
- Pareja-Galeano, H.; Garatachea, N.; Lucia, A. Exercise as a polypill for chronic diseases. Prog. Mol. Biol. Transl. Sci. 2015, 135, 497–526. [Google Scholar]
- Pickering, C.; Kiely, J. Can the ability to adapt to exercise be considered a talent—And if so, can we test for it? Sports Med. Open 2017, 3, 43. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.; Kiely, J.; Suraci, B.; Collins, D.J.; De Lorenzo, D.; Pickering, C.; Grimaldi, K.A. A genetic-based algorithm for personalized resistance training. Biol. Sport 2016, 33, 117. [Google Scholar] [CrossRef] [PubMed]
- Pickering, C.; Kiely, J.; Suraci, B.; Collins, D. The magnitude of Yo-Yo test improvements following an aerobic training intervention are associated with total genotype score. PLoS ONE 2018, 13, e0207597. [Google Scholar] [CrossRef] [PubMed]
- Varley, I.; Hughes, D.C.; Greeves, J.P.; Stellingwerff, T.; Ranson, C.; Fraser, W.D.; Sale, C. The association of novel polymorphisms with stress fracture injury in elite athletes: Further insights from the SFEA cohort. J. Sci. Med. Sport 2018, 21, 564–568. [Google Scholar] [CrossRef] [PubMed]
- Abe, D.; Doi, H.; Asai, T.; Kimura, M.; Wada, T.; Takahashi, Y.; Matsumoto, T.; Shinohara, K. Association between COMT Val158Met polymorphism and competition results of competitive swimmers. J. Sports Sci. 2018, 36, 393–397. [Google Scholar] [CrossRef]
- Van Breda, K.; Collins, M.; Stein, D.J.; Rauch, L. The COMT val158met polymorphism in ultra-endurance athletes. Physiol. Behav. 2015, 151, 279–283. [Google Scholar] [CrossRef]
- Jacob, Y.; Cripps, A.; Evans, T.; Chivers, P.T.; Joyce, C.; Anderton, R.S. Identification of genetic markers for skill and athleticism in sub-elite Australian football players: A pilot study. J. Sports Med. Phys. Fit. 2018, 58, 241–248. [Google Scholar]
- McFie, S.; Abrahams, S.; Patricios, J.; Suter, J.; Posthumus, M.; September, A.V. The association between COMT rs4680 and 5-HTTLPR genotypes and concussion history in South African rugby union players. J. Sports Sci. 2018, 36, 920–933. [Google Scholar] [CrossRef]
- Nielsen, D.E.; El-Sohemy, A. Disclosure of genetic information and change in dietary intake: A randomized controlled trial. PLoS ONE 2014, 9, e112665. [Google Scholar] [CrossRef]
- Vlahovich, N.; Fricker, P.A.; Brown, M.A.; Hughes, D. Ethics of genetic testing and research in sport: A position statement from the Australian Institute of Sport. Br. J. Sports Med. 2017, 51, 5–11. [Google Scholar] [CrossRef]
- Heffernan, S.M.; Kilduff, L.P.; Day, S.H.; Pitsiladis, Y.P.; Williams, A.G. Genomics in rugby union: A review and future prospects. Eur. J. Sport Sci. 2015, 15, 460–468. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pickering, C.; Kiely, J.; Grgic, J.; Lucia, A.; Del Coso, J. Can Genetic Testing Identify Talent for Sport? Genes 2019, 10, 972. https://doi.org/10.3390/genes10120972
Pickering C, Kiely J, Grgic J, Lucia A, Del Coso J. Can Genetic Testing Identify Talent for Sport? Genes. 2019; 10(12):972. https://doi.org/10.3390/genes10120972
Chicago/Turabian StylePickering, Craig, John Kiely, Jozo Grgic, Alejandro Lucia, and Juan Del Coso. 2019. "Can Genetic Testing Identify Talent for Sport?" Genes 10, no. 12: 972. https://doi.org/10.3390/genes10120972
APA StylePickering, C., Kiely, J., Grgic, J., Lucia, A., & Del Coso, J. (2019). Can Genetic Testing Identify Talent for Sport? Genes, 10(12), 972. https://doi.org/10.3390/genes10120972