Identification of Long Non-Coding RNA-Associated Competing Endogenous RNA Network in the Differentiation of Chicken Preadipocytes

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Culture and Induction
2.3. RNA Extraction and Library Construction
2.4. LncRNA Identification
2.5. miRNA Identification
2.6. Identification of Differentially Expressed Genes
2.7. Construction of LncRNAs-miRNAs-mRNAs ceRNA Regulatory Network
2.8. Functional Enrichment Analysis
2.9. Quantitative Reverse Transcription Polymerase Chain Reaction
2.10. Statistical Analysis
3. Results
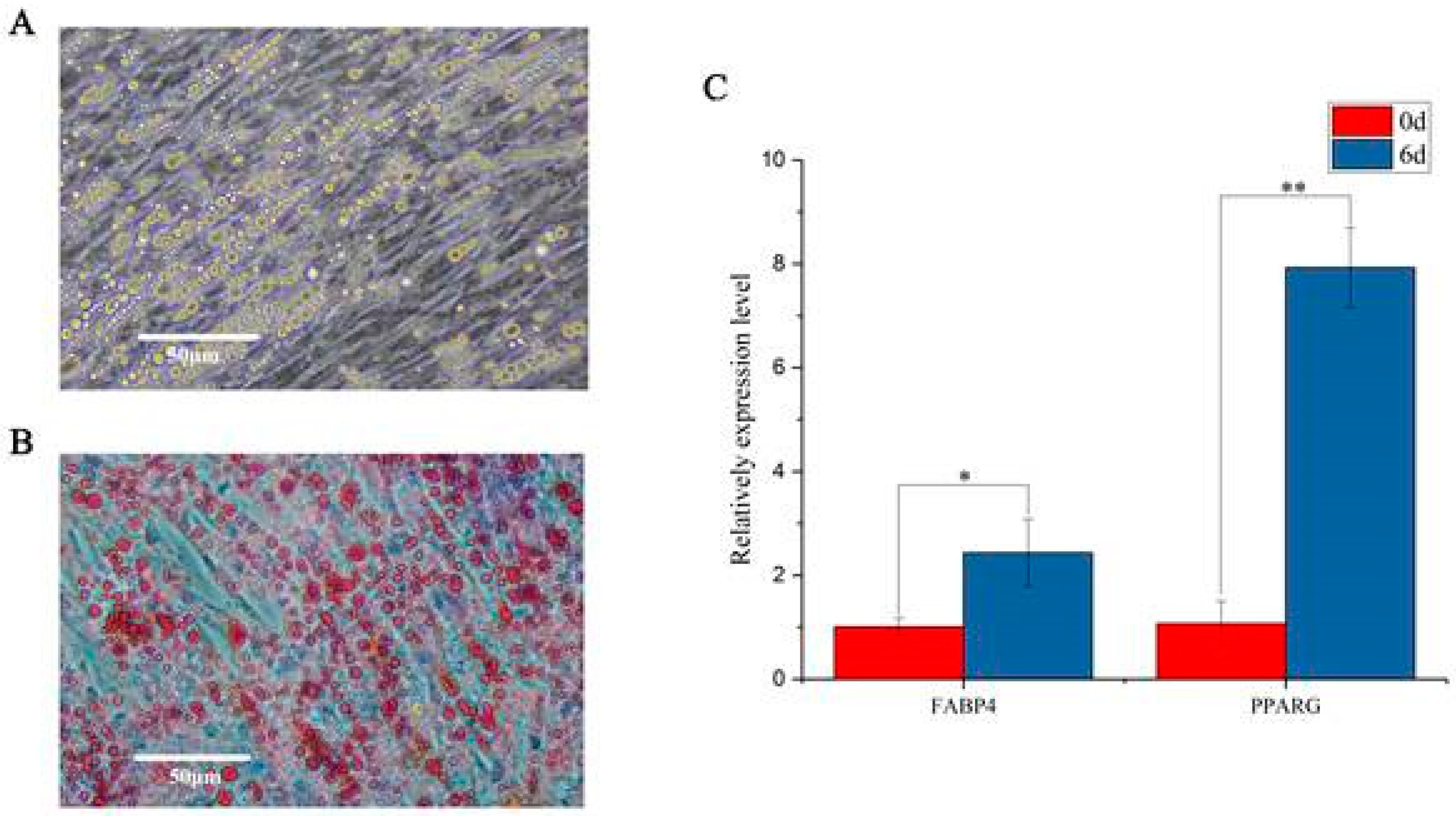
3.1. Induction of Chicken Preadipocytes
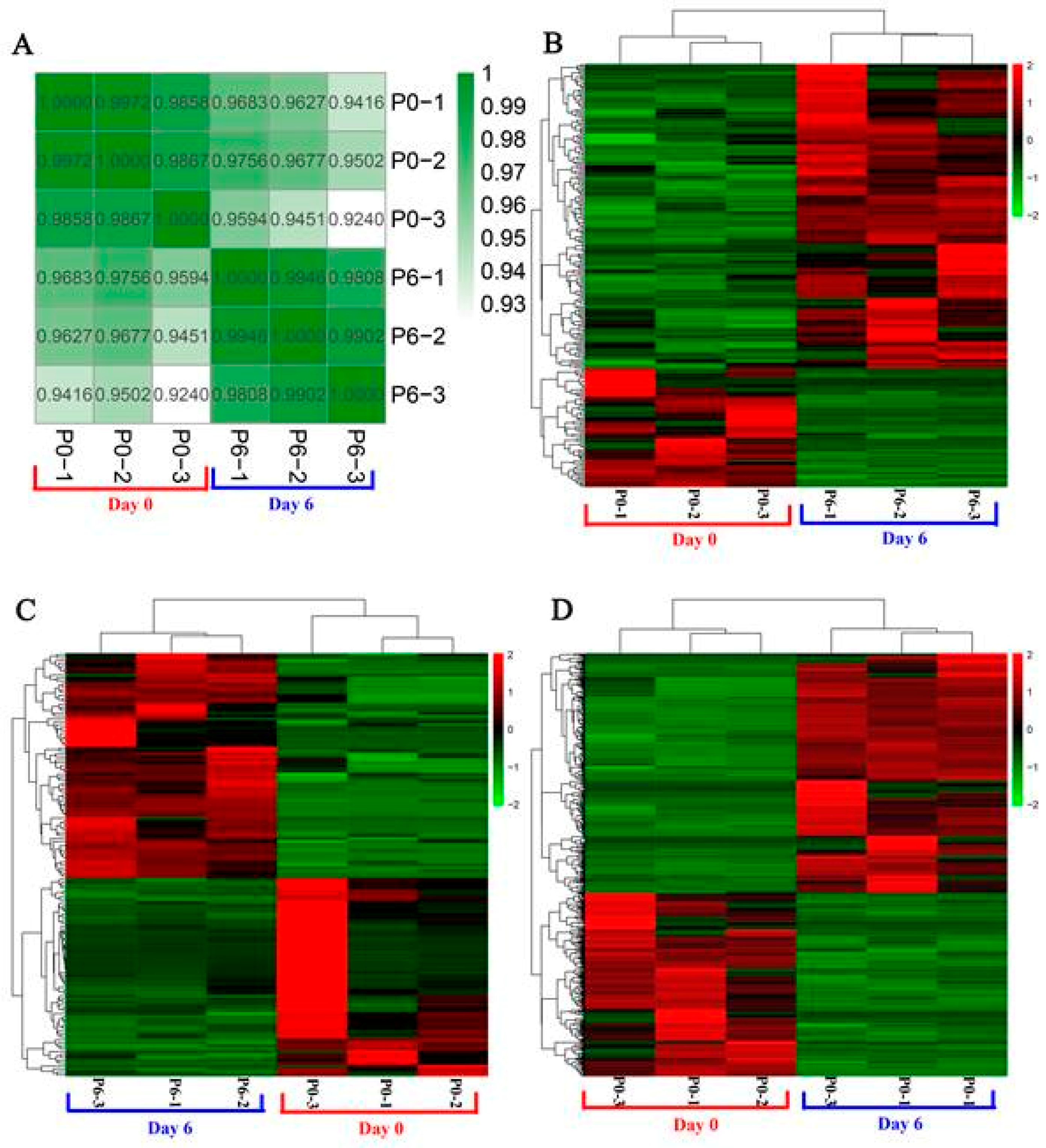
3.2. Identification of Differentially Expressed lncRNAs, miRNAs, and mRNAs
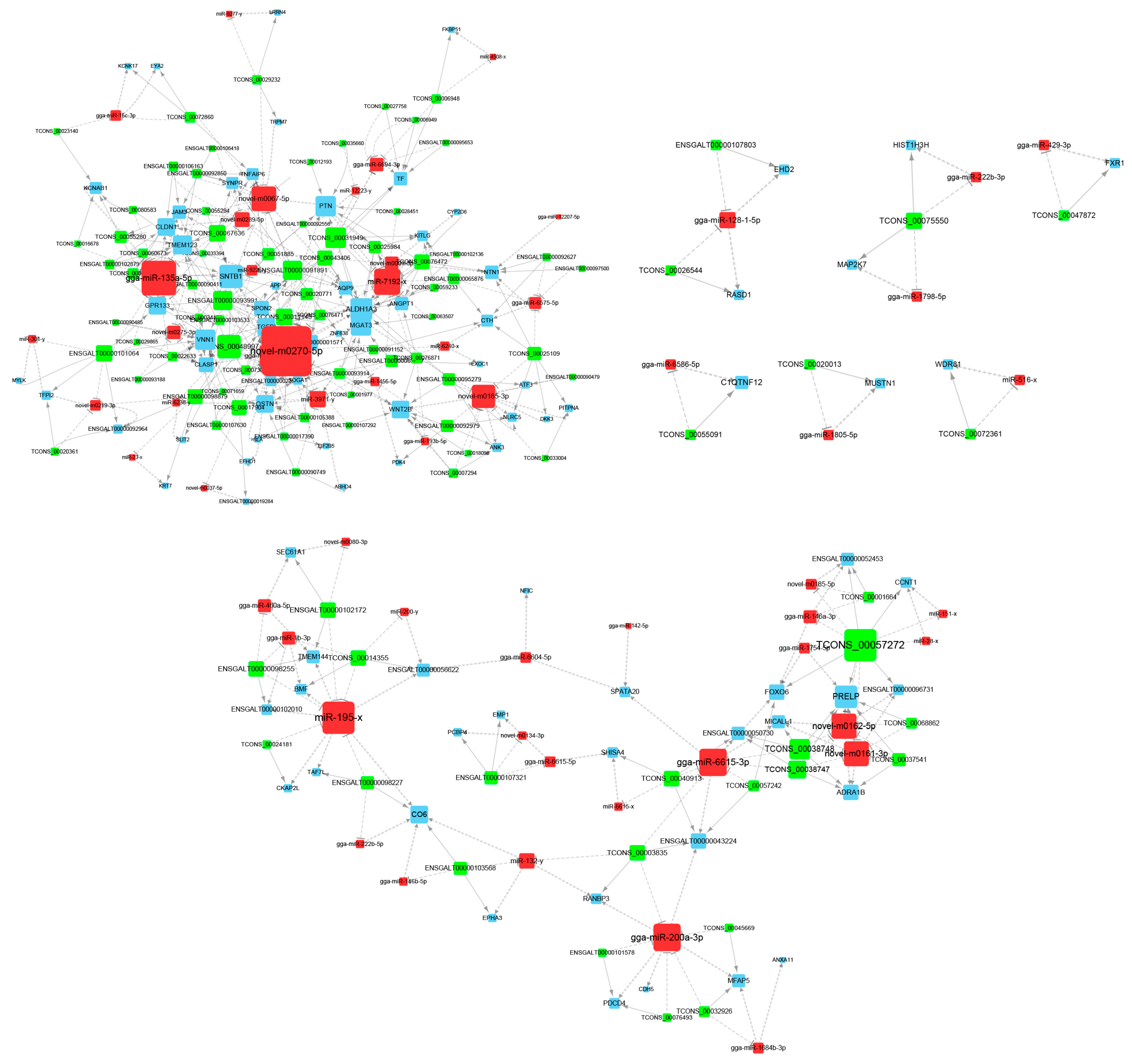
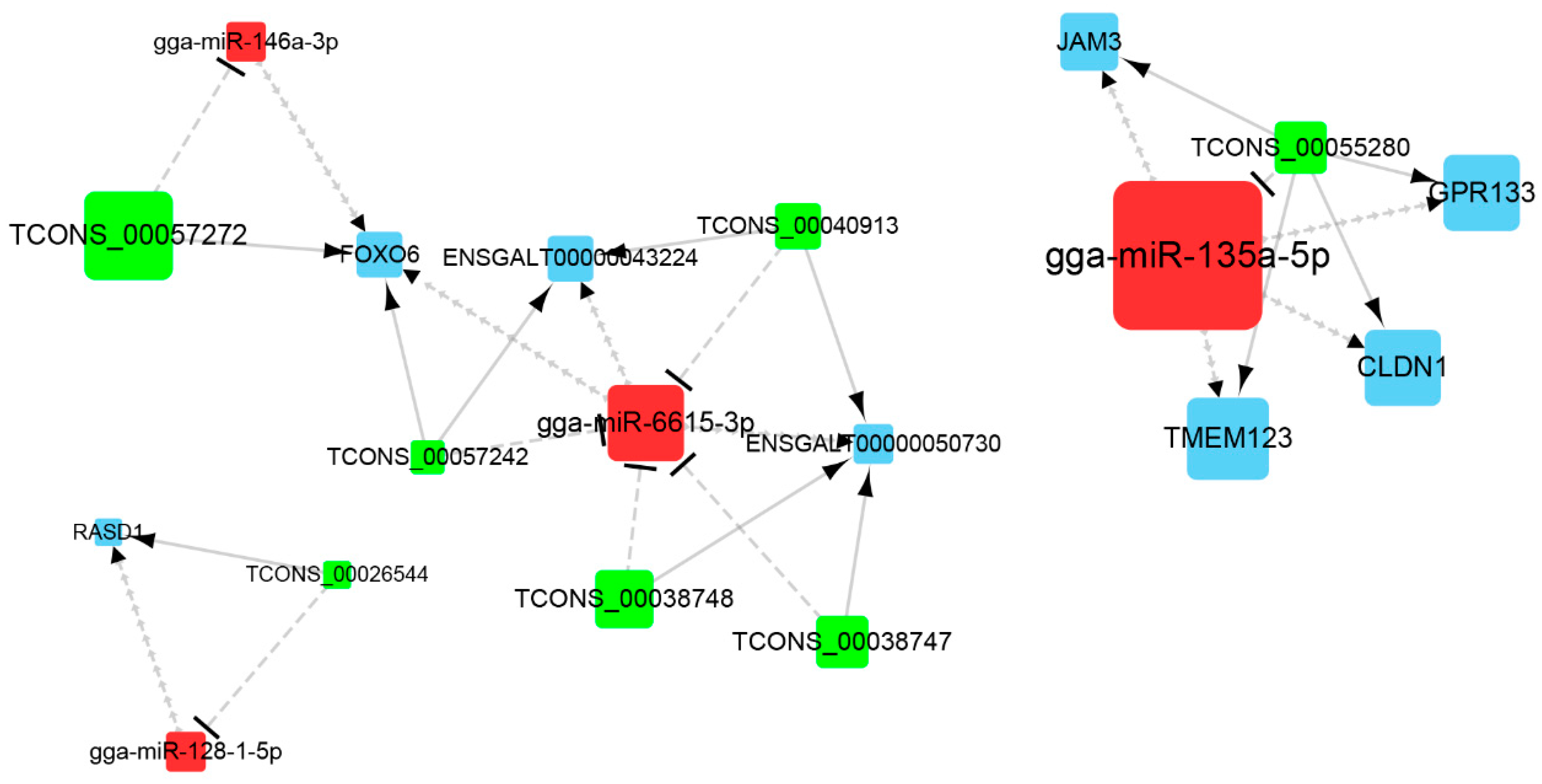
3.3. Construction of ceRNA Network Related to Preadipocytes Differentiation
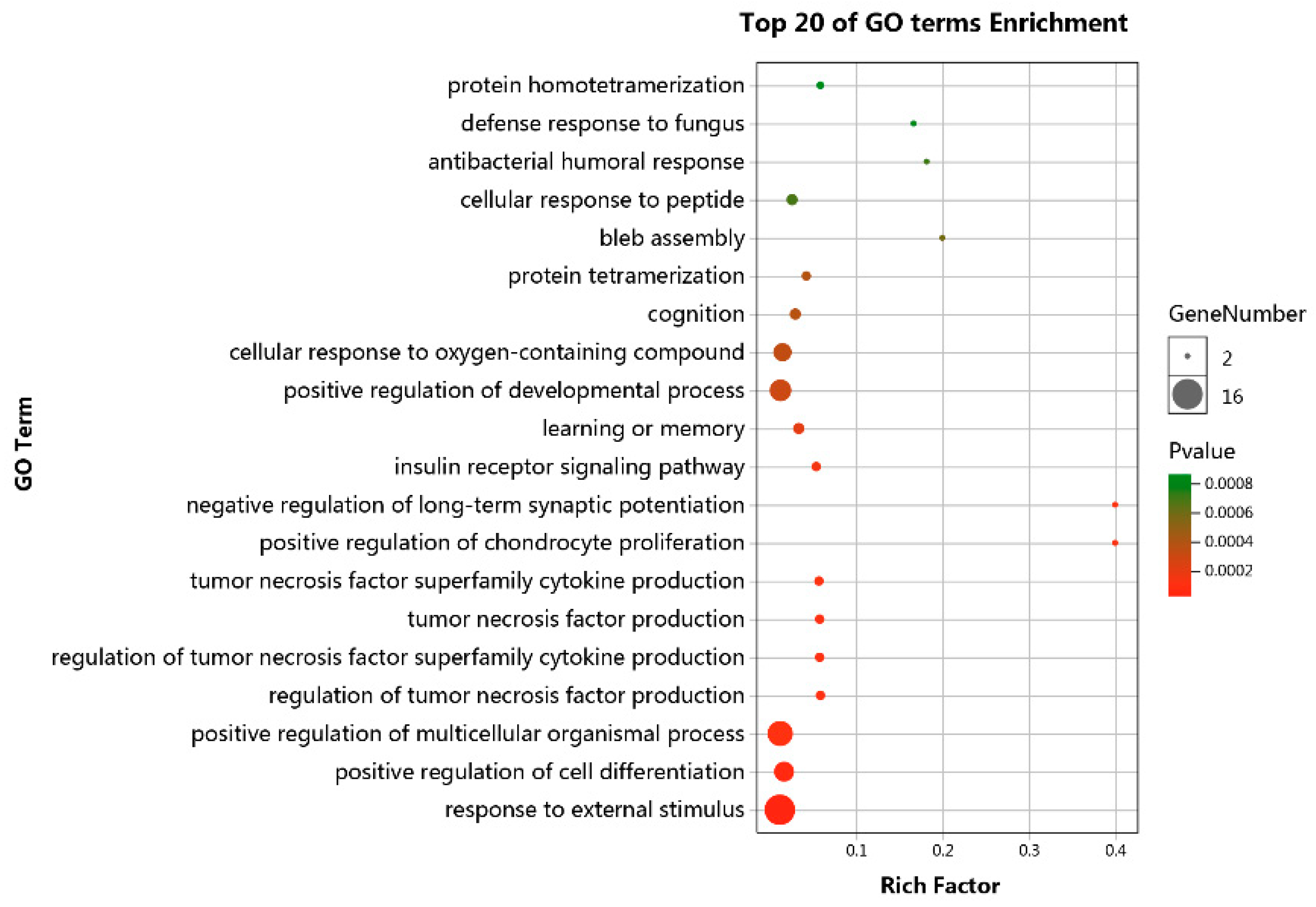
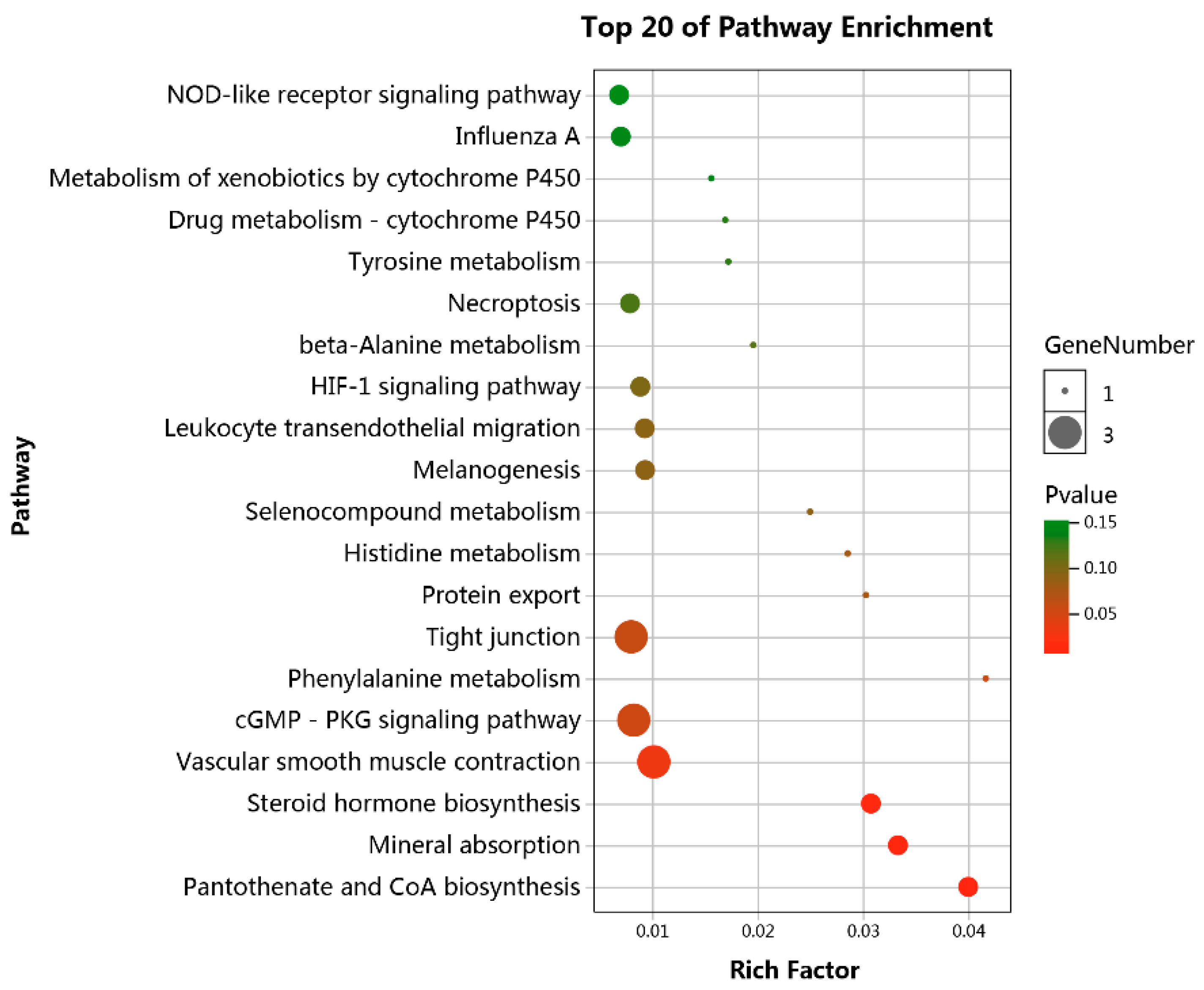
3.4. Functional Enrichment Analyses
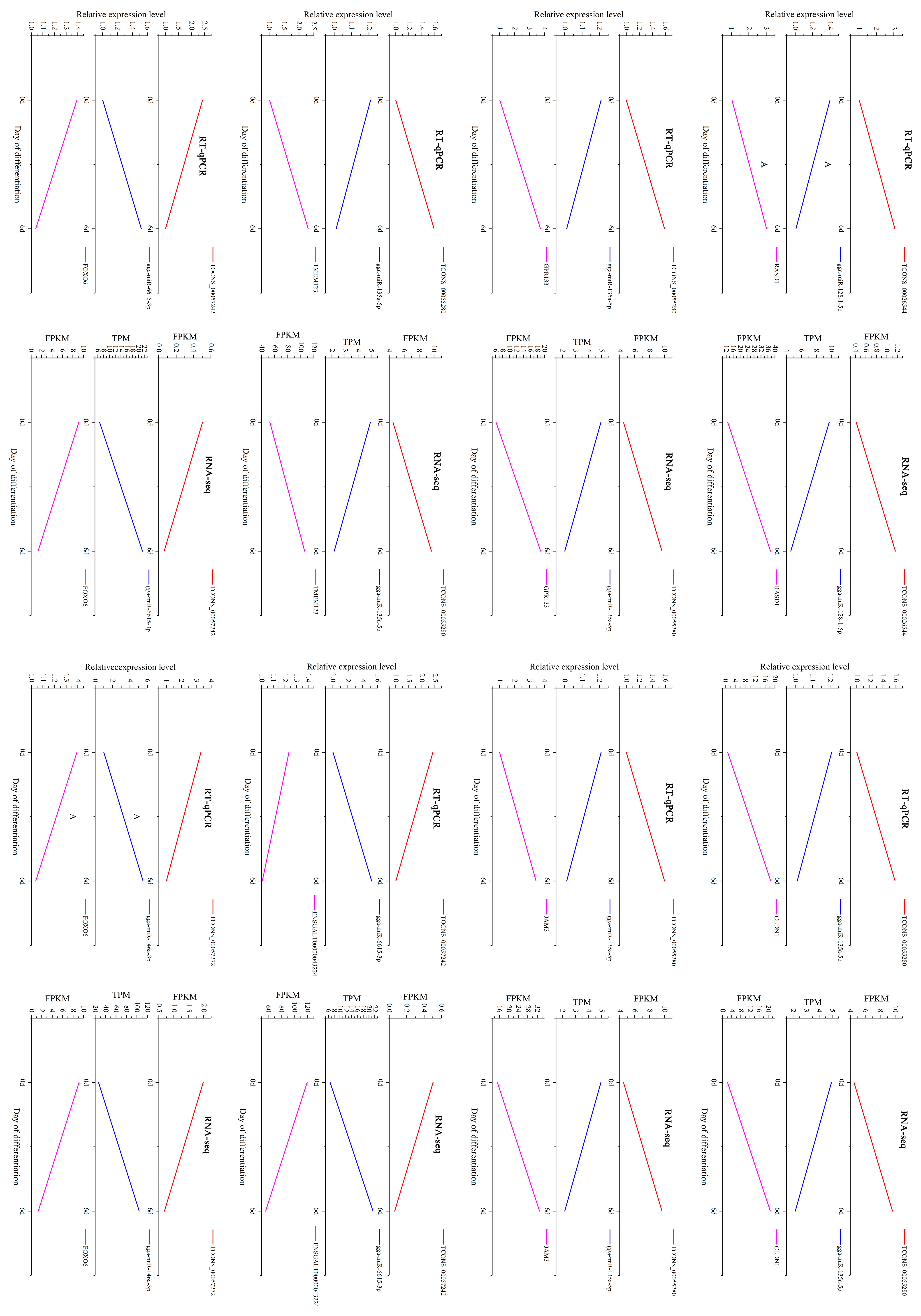
3.5. Identification and RT-qPCR Validation of Crucial Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Knowles, T.G.; Kestin, S.C.; Haslam, S.M.; Brown, S.N.; Green, L.E.; Butterworth, A.; Pope, S.J.; Pfeiffer, D.; Nicol, C.J. Leg disorders in broiler chickens: Prevalence, risk factors and prevention. PLoS ONE 2008, 3, e1545. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Deeb, N.; Evock-Clover, C.M.; Ashwell, C.M.; Lamont, S.J. Genome-wide linkage analysis to identify chromosomal regions affecting phenotypic traits in the chicken. II. Body composition. Poult. Sci. 2006, 85, 1712–1721. [Google Scholar] [CrossRef] [PubMed]
- Sahraei, M. Feed restriction in broiler chickens production. Biotechnol. Anim. Husb. 2012, 28, 333–352. [Google Scholar] [CrossRef]
- Braun, E.J.; Sweazea, K.L. Glucose regulation in birds. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 151, 1–9. [Google Scholar] [CrossRef]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. MicroRNAs in metabolism. Acta Physiol. 2017, 219, 346–361. [Google Scholar] [CrossRef]
- Bhaskaran, M.; Mohan, M. MicroRNAs: History, biogenesis, and their evolving role in animal development and disease. Vet. Pathol. 2014, 51, 759–774. [Google Scholar] [CrossRef]
- Wei, S.; Du, M.; Jiang, Z.; Hausman, G.J.; Zhang, L.; Dodson, M.V. Long noncoding RNAs in regulating adipogenesis: New RNAs shed lights on obesity. Cell. Mol. Life Sci. 2016, 73, 2079–2087. [Google Scholar] [CrossRef]
- Tang, H.; Chen, Y.; Zhou, H. [Research progress of long noncoding RNA in regulating adipogenesis]. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2018, 43, 912–919. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Z.; Wu, N.; Guo, H. Integrative Analysis of Dysregulated lncRNA-Associated ceRNA Network Reveals Functional lncRNAs in Gastric Cancer. Genes 2018, 9, 303. [Google Scholar] [CrossRef]
- Pilyugin, M.; Irminger-Finger, I. Long non-coding RNA and microRNAs might act in regulating the expression of BARD1 mRNAs. Int. J. Biochem. Cell B. 2014, 54, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ma, C.; Yang, B.; Yin, C.; Zhang, B.; Xiao, Y. LncRNA Gm15290 sponges miR-27b to promote PPARγ-induced fat deposition and contribute to body weight gain in mice. Biochem. Biophys. Res. Commun. 2017, 493, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xie, Z.; Wang, P.; Li, J.; Liu, W.; Tang, S.; Liu, Z.; Wu, X.; Wu, Y.; Shen, H. The long noncoding RNA GAS5 negatively regulates the adipogenic differentiation of MSCs by modulating the miR-18a/CTGF axis as a ceRNA. Cell Death Dis. 2018, 9, 554. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, H.; Jin, L.; Li, G.; Hu, S.; Ning, C.; Guo, J.; Shuai, S.; Li, X.; Li, M. Long Noncoding RNA GAS5 Suppresses 3T3-L1 Cells Adipogenesis Through miR-21a-5p/PTEN Signal Pathway. DNA Cell Biol. 2018, 37, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Sun, X.; Cai, H.; Sun, Y.; Plath, M.; Li, C.; Lan, X.; Lei, C.; Lin, F.; Bai, Y.; et al. Long non-coding RNA ADNCR suppresses adipogenic differentiation by targeting miR-204. Biochim. Biophys. Acta 2016, 1859, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Guo, L.; Wang, N.; Shi, H.; Wang, Y.; Li, H. Oleate promotes differentiation of chicken primary preadipocytes in vitro. Biosci. Rep. 2014, 34, e00093. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Bairoch, A.; Apweiler, R. The SWISS-PROT protein sequence database and its supplement TrEMBL in 2000. Nucleic Acids Res. 2000, 28, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Karsch-Mizrachi, I.; Clark, K.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2012, 40, D48–D53. [Google Scholar] [CrossRef] [PubMed]
- Kalvari, I.; Argasinska, J.; Quinones-Olvera, N.; Nawrocki, E.P.; Rivas, E.; Eddy, S.R.; Bateman, A.; Finn, R.D.; Petrov, A.I. Rfam 13.0: Shifting to a genome-centric resource for non-coding RNA families. Nucleic Acids Res. 2018, 46, D335–D342. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.; Wang, X. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015, 43, D146–D152. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Xu, X.W.; Zhou, X.H.; Wang, R.R.; Peng, W.L.; An, Y.; Chen, L.L. Functional analysis of long intergenic non-coding RNAs in phosphate-starved rice using competing endogenous RNA network. Sci. Rep. 2016, 6, 20715. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Chen, C.; Deng, Y.; Hu, X.; Ren, H.; Zhu, J.; Fu, S.; Xie, J.; Peng, Y. miR-128-3p regulates 3T3-L1 adipogenesis and lipolysis by targeting Pparg and Sertad2. J. Physiol. Biochem. 2018, 74, 381–393. [Google Scholar] [CrossRef]
- Ye, X.; Wang, X.B.; Wang, J.; Ming, J. Effect of MicroRNA-146a on Differentiation Potential of Human Bone Marrow Mesenchymal Stem Cells. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2016, 24, 596–601. [Google Scholar] [CrossRef]
- Chen, C.; Peng, Y.; Peng, Y.; Peng, J.; Jiang, S. miR-135a-5p inhibits 3T3-L1 adipogenesis through activation of canonical Wnt/β-catenin signaling. J. Mol. Endocrinol. 2014, 52, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Kim, W.K.; Cline, M.A.; Gilbert, E.R. Factors affecting adipose tissue development in chickens: A review. Poul. Sci. 2017, 96, 3687–3699. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Hao, Q.; Prasanth, K.V. Nuclear Long Noncoding RNAs: Key Regulators of Gene Expression. Trends Genet. 2018, 34, 142–157. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, J.; Guo, X.; Chen, Y.; Xing, C.; Gao, A. Benzene and its metabolite decreases cell proliferation via LncRNA-OBFC2A-mediated anti-proliferation effect involving NOTCH1 and KLF15. Oncotarget 2017, 8, 40857–40871. [Google Scholar] [CrossRef]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef]
- Chen, L.; Yang, W.; Guo, Y.; Chen, W.; Zheng, P.; Zeng, J.; Tong, W. Exosomal lncRNA GAS5 regulates the apoptosis of macrophages and vascular endothelial cells in atherosclerosis. PLoS ONE 2017, 12, e0185406. [Google Scholar] [CrossRef]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Y.; Lu, S.; Yin, L.; Zong, C.; Cui, S.; Qin, D.; Yang, Y.; Guan, Q.; Li, X.; et al. The role and possible mechanism of lncRNA U90926 in modulating 3T3-L1 preadipocyte differentiation. Int. J. Obes. 2017, 41, 299–308. [Google Scholar] [CrossRef]
- Shipp, S.L.; Cline, M.A.; Gilbert, E.R. Promotion of adipogenesis by neuropeptide Y during the later stages of chicken preadipocyte differentiation. Physiol. Rep. 2016, 4, e13006. [Google Scholar] [CrossRef]
- Wang, W.; Cheng, M.; Qiao, S.; Wang, Y.; Li, H.; Wang, N. Gga-miR-21 inhibits chicken pre-adipocyte proliferation in part by down-regulating Kruppel-like factor 5. Poul. Sci. 2017, 96, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, F.; Sun, J.W.; Li, D.H.; Li, W.T.; Jiang, R.R.; Li, Z.J.; Liu, X.J.; Han, R.L.; Li, G.X.; et al. LncRNA IMFNCR Promotes Intramuscular Adipocyte Differentiation by Sponging miR-128-3p and miR-27b-3p. Front. Genet. 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, D.; Zhang, M.; Sun, J.; Li, W.; Jiang, R.; Han, R.; Wang, Y.; Tian, Y.; Kang, X.; et al. miRNA-223 targets the GPAM gene and regulates the differentiation of intramuscular adipocytes. Gene 2019, 685, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Hu, G.; Zhang, X.; Yu, T.; Yang, J. Long non-coding RNA DSCAM-AS1 accelerates the progression of hepatocellular carcinoma via sponging miR-338-3p. Am. J. Transl. Res. 2019, 11, 4290–4302. [Google Scholar] [PubMed]
- Chen, K.; Xie, S.; Jin, W. Crucial lncRNAs associated with adipocyte differentiation from human adipose-derived stem cells based on co-expression and ceRNA network analyses. PeerJ 2019, 7, e7544. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Cao, Y. An lncRNAmiRNAmRNA ceRNA network for adipocyte differentiation from human adiposederived stem cells. Mol. Med. Rep. 2019, 19, 4271–4287. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Liu, Y.; Cui, J.; Zhao, Z.; Cao, H.; Fu, Z.; Liu, B. Long noncoding RNA H19 mediates LCoR to impact the osteogenic and adipogenic differentiation of mBMSCs in mice through sponging miR-188. J. Cell. Physiol. 2018, 233, 7435–7446. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer sequences |
|---|---|
| TOCNS_00055280 | GCAGAACGGATGGTGCCTTTGT |
| GGCTGTCCTGGCTGAAGATGGA | |
| TOCNS_00057242 | CCCTTCTCCGCTCACAGTCCTT |
| CGCTTCGGCTGTAAACGTCCAT | |
| TOCNS_00057272 | GGCTACCCGTCTCCTCCAAGAT |
| GTGCCTCGTTACGCCTGATTGA | |
| TOCNS_00026544 | ACGGAGATGCTGCGGTTATGC |
| AACAAACCCAACCCGTTCCCAG | |
| ENSGALT00000043224 | TCCAAGAAGGCGGTCACCAAGA |
| GCACCTGCTTCAGCACCTTGT | |
| FOXO6 | CAGCAGACCTGGACCTGGACAT |
| CGCCGAGTCGAAGTTGAAGTCC | |
| RASD1 | CCTCGTGTTCAGCCTGGACAAC |
| CGGCACCTCGATGTTCTCCTTG | |
| GPR133 | GTGAGCACCATCCGCAACCAA |
| AAGGCACCGTTCCAGGACTGAA | |
| JAM3 | GGAAGTCCTCCTCGCAGCAGTT |
| AACAAGCCAGGTGCCCACTCT | |
| CLDN1 | GCTGATTGCTTCCAACCAGGCT |
| GCACACGGCTCTCCTTGTCTAC | |
| TMEM123 | CGTACCACTCGAGAAGAGGC |
| CAGCTGTGACAGGATGGGTT | |
| gga-miR-6615-3p | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTGTGGA |
| CGCGTGGCACTGATGTGTTC | |
| gga-miR-146a-3p | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTGAAG |
| CGACCCATGGGGCTCAGTT | |
| gga-miR-128-1-5p | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCTCAG |
| GCGGGGCCGTAACACTGT | |
| gga-miR-135a-5p | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCACAT |
| CGCGTATGGCTTTTTATTCCT |
| lncRNA | miRNA | mRNA |
|---|---|---|
| TCONS_00026544 | gga-miR-128-1-5p | RASD1 |
| TCONS_00057272 | gga-miR-146a-3p | FOXO6 |
| TCONS_00057242 | gga-miR-6615-3p | FOXO6 |
| TCONS_00057242 | gga-miR-6615-3p | ENSGALT00000043224 |
| TCONS_00040913 | gga-miR-6615-3p | ENSGALT00000043224 |
| TCONS_00040913 | gga-miR-6615-3p | ENSGALT00000050730 |
| TCONS_00038747 | gga-miR-6615-3p | ENSGALT00000050730 |
| TCONS_00038748 | gga-miR-6615-3p | ENSGALT00000050730 |
| TCONS_00055280 | gga-miR-135a-5p | JAM3 |
| TCONS_00055280 | gga-miR-135a-5p | GPR133 |
| TCONS_00055280 | gga-miR-135a-5p | CLDN1 |
| TCONS_00055280 | gga-miR-135a-5p | TMEM123 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Zhang, T.; Zhang, S.; Huang, J.; Zhang, G.; Xie, K.; Wang, J.; Wu, H.; Dai, G. Identification of Long Non-Coding RNA-Associated Competing Endogenous RNA Network in the Differentiation of Chicken Preadipocytes. Genes 2019, 10, 795. https://doi.org/10.3390/genes10100795
Chen L, Zhang T, Zhang S, Huang J, Zhang G, Xie K, Wang J, Wu H, Dai G. Identification of Long Non-Coding RNA-Associated Competing Endogenous RNA Network in the Differentiation of Chicken Preadipocytes. Genes. 2019; 10(10):795. https://doi.org/10.3390/genes10100795
Chicago/Turabian StyleChen, Lan, Tao Zhang, Shanshan Zhang, Jinxing Huang, Genxi Zhang, Kaizhou Xie, Jinyu Wang, Haiqing Wu, and Guojun Dai. 2019. "Identification of Long Non-Coding RNA-Associated Competing Endogenous RNA Network in the Differentiation of Chicken Preadipocytes" Genes 10, no. 10: 795. https://doi.org/10.3390/genes10100795
APA StyleChen, L., Zhang, T., Zhang, S., Huang, J., Zhang, G., Xie, K., Wang, J., Wu, H., & Dai, G. (2019). Identification of Long Non-Coding RNA-Associated Competing Endogenous RNA Network in the Differentiation of Chicken Preadipocytes. Genes, 10(10), 795. https://doi.org/10.3390/genes10100795