Astroglial Isopotentiality and Calcium-Associated Biomagnetic Field Effects on Cortical Neuronal Coupling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
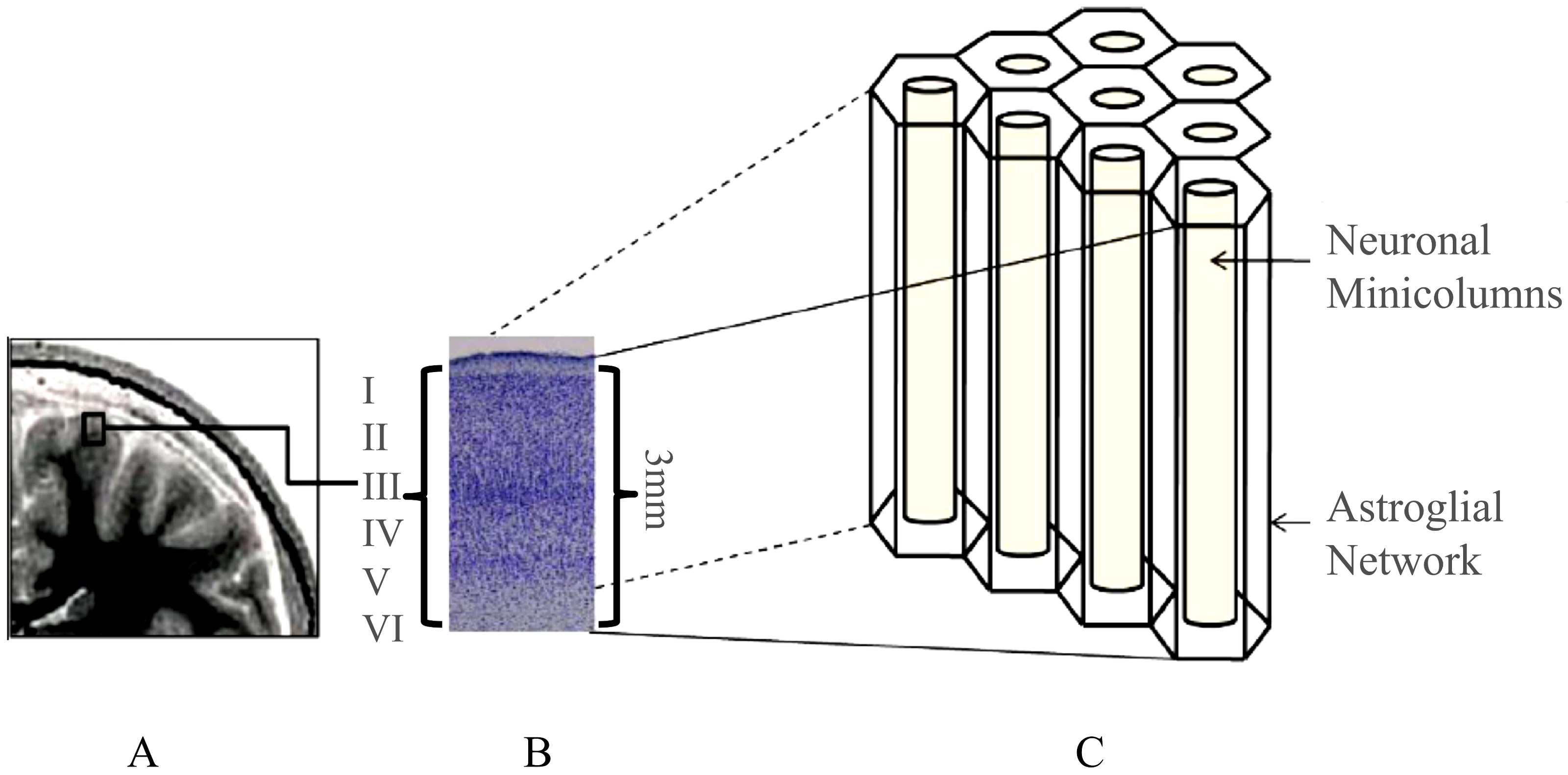
2. The Complexity of the Neocortex
Structural–Functional Relationship in the Neocortex
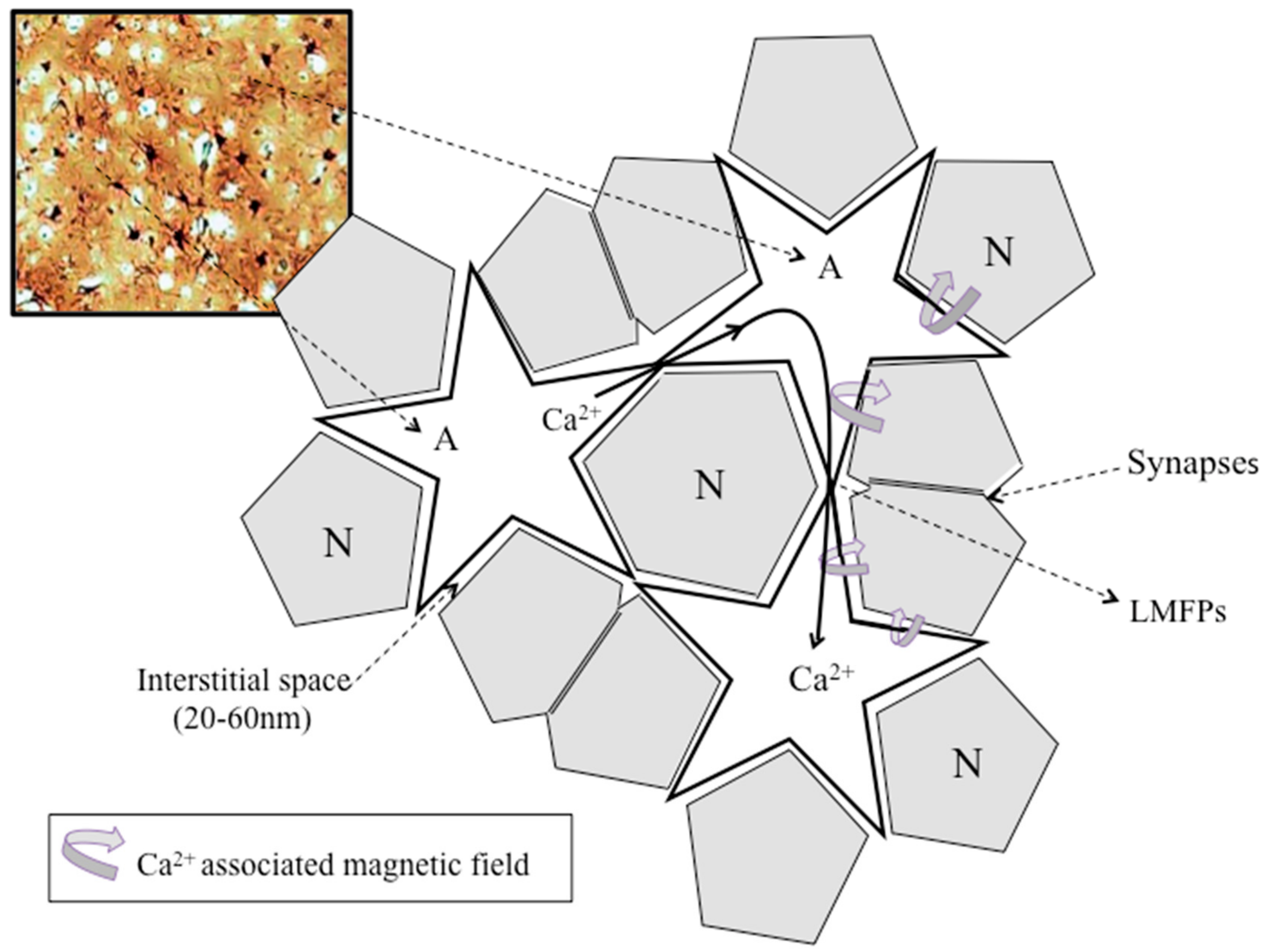
3. Ionic Neuron–Astroglial Crosstalk in the Neocortex
3.1. Potassium
3.2. Calcium
3.3. LFPs in Neocortical Synchronization
3.4. LFP and Ca2+ Transients in Astrocytes
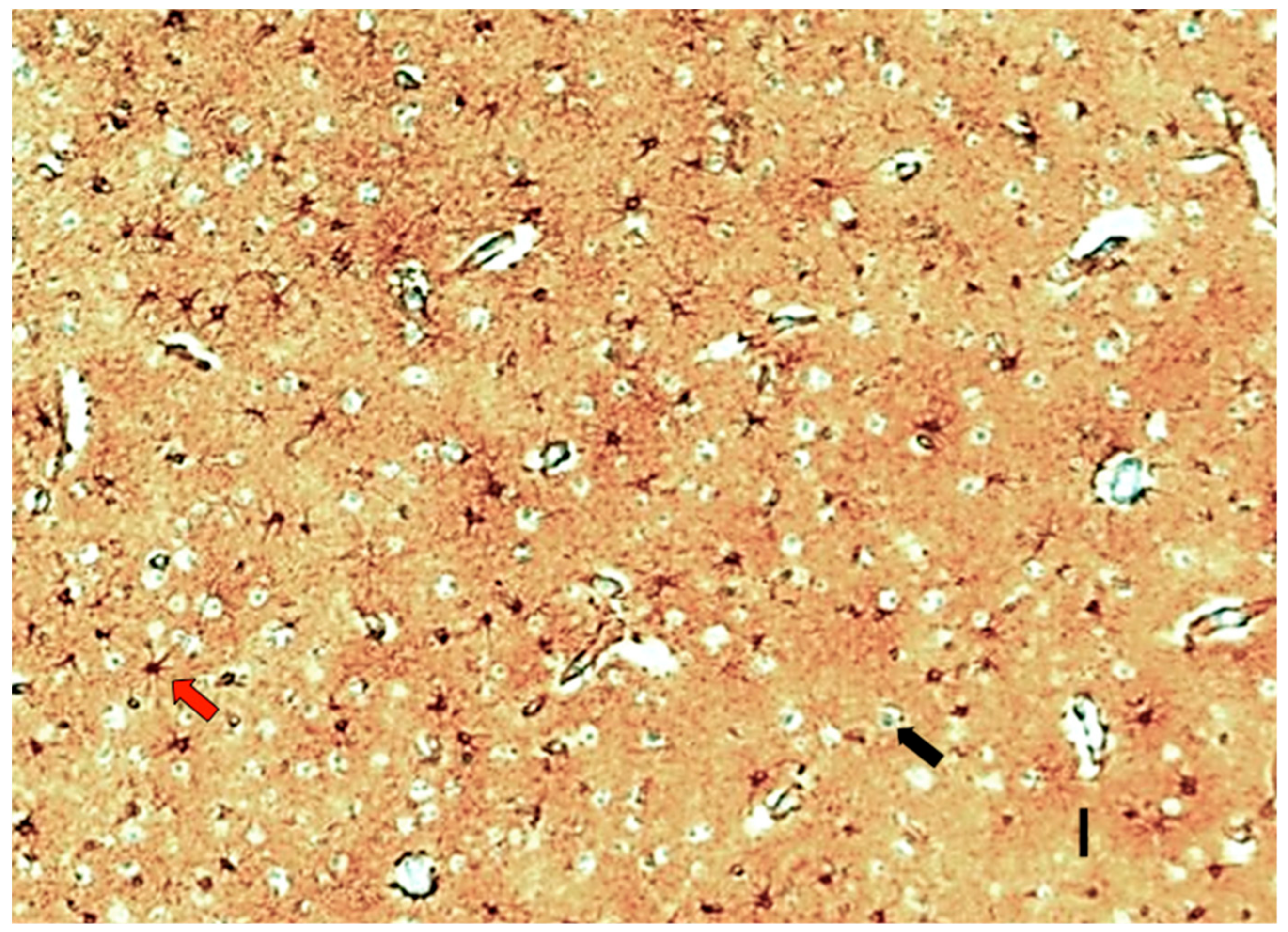
3.5. Neocortical Astroglial Isopotentiality
3.6. Mechanisms of Synchronization in the Neocortex
4. Bioelectromagnetic Fields in the Neocortex
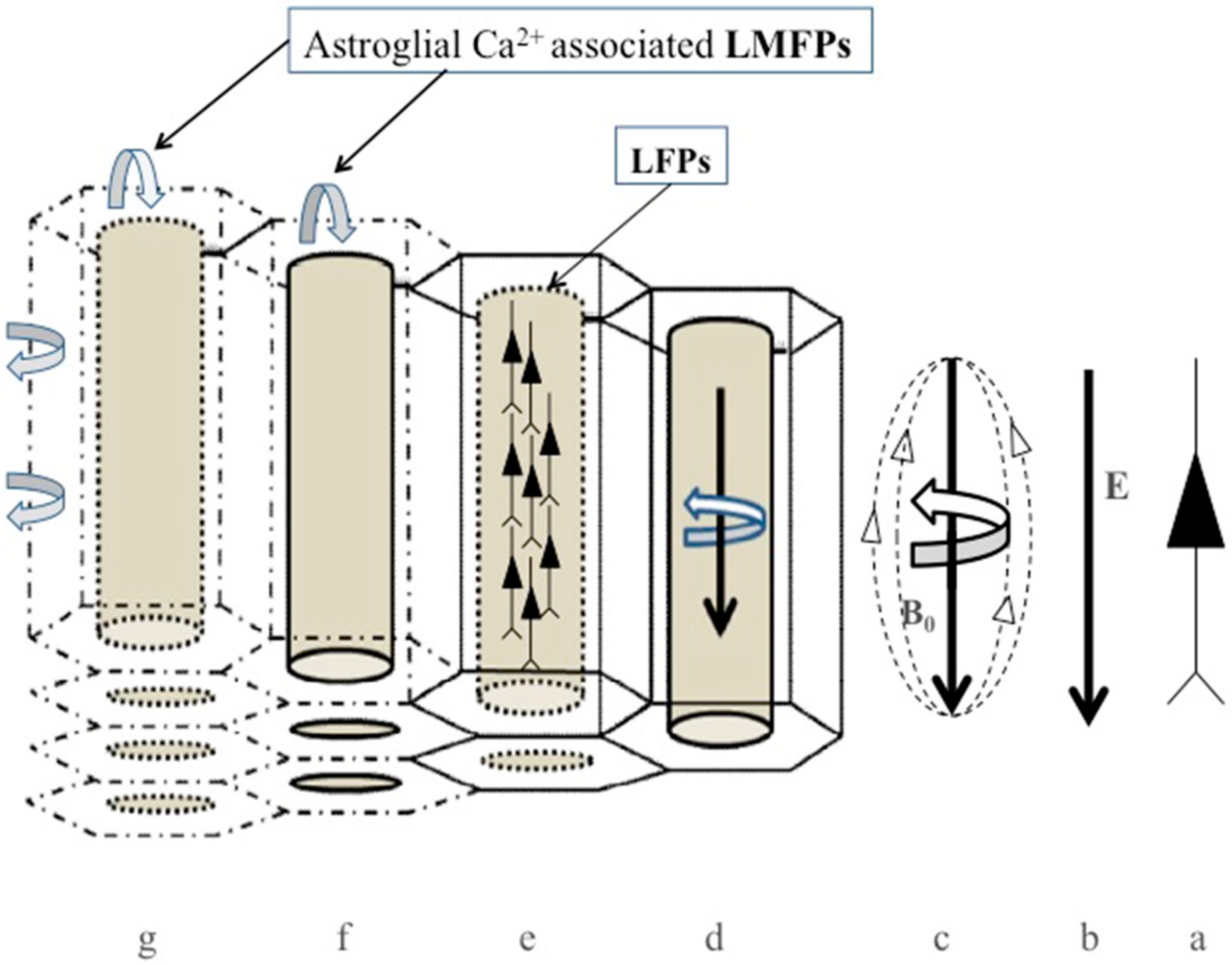
4.1. LFP, LMFP, and Ca2+-Associated Astroglial Biomagnetic Field
4.2. Transcranial Electromagnetic Stimulation: Role of Astroglial Isopotentiality
5. Ongoing Challenges
6. Conclusions
Funding
Conflicts of Interest
References
- Katz, B.; Schmitt, O.H. Electric interaction between two adjacent nerve fibers. J. Physiol. 1940, 97, 471–488. [Google Scholar] [CrossRef]
- Arvanitaki, A. Effects evoked in an axon by the activity of a contiguous one. J. Neurophysiol. 1942, 5, 89–108. [Google Scholar] [CrossRef]
- Rosen, A.S.; Andrew, R.D. Osmotic effects upon excitability in rat neocortical slices. Neuroscience 1990, 38, 579–590. [Google Scholar] [CrossRef]
- Syková, E. Extrasynaptic volumen transmission and diffusion parameters of the extracelular space. Neuroscience 2004, 129, 861–876. [Google Scholar] [CrossRef]
- Durand, D.M.; Park, E.-H.; Jensen, A.L. Potassium diffusive coupling in neural networks. Philos. Trans. R Soc. Lond. B Biol. Sci. 2010, 365, 2347–2362. [Google Scholar] [CrossRef] [PubMed]
- Faber, D.S.; Pereda, A.E. Two Forms of Electrical Transmission between Neurons. Front. Mol. Neurosci. 2018, 11, 427. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G.; Anastassiou, C.A.; Koch, C. The origin of extracellular fields and currents- EEG, ECoG, LFP and spikes. Nat. Rev. Neurosci. 2012, 13, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Reimann, M.W.; Anastassiou, C.A.; Perin, R.; Hill, S.L.; Markram, H.; Koch, C. A biophysically detailed model of neocortical local field potentials predicts the critical role of active membrane currents. Neuron 2013, 79, 375–390. [Google Scholar] [CrossRef]
- Jefferys, J.G.; Haas, H.L. Synchronized bursting of CA1 hippocampal pyramidal cells in the absence of synaptic transmission. Nature 1982, 300, 448–450. [Google Scholar] [CrossRef]
- Jefferys, J.G. Nonsynaptic modulation of neuronal activity in the brain: Electric currents and extracellular ions. Physiol. Rev. 1985, 75, 689–723. [Google Scholar] [CrossRef]
- Ray, S. Challenges in the quantification and interpretation of spike-LFP relationships. Curr. Opin. Neurobiol. 2015, 31, 111–118. [Google Scholar] [CrossRef]
- Creutzfeldt, O.D.; Houchin, J. Neuronal basis of EEG waves. In Handbook of Electroencephalography and Clinical Neurophysiology; Remond, A., Ed.; Elsevier: Amsterdam, The Nederlands, 1974; Volume 2, pp. 5–55. [Google Scholar]
- Mitzdorf, U. Current source-density method and application in cat cerebral cortex: Investigation of evoked potentials and EEG phenomena. Physiol. Rev. 1985, 65, 37–100. [Google Scholar] [CrossRef] [PubMed]
- Peterchev, A.V.; Wagner, T.A.; Miranda, P.C.; Nitsche, M.A.; Paulus, W.; Lisanby, S.H.; Pascual-Leone, A.; Bikson, M. Fundamentals of transcranial electric and magnetic stimulation dose: Definition, selection, and reporting practices. Brain Stimul. 2012, 5, 435–453. [Google Scholar] [CrossRef] [PubMed]
- Ramón y Cajal, S. Histology of the nervous system of man and vertebrates (1897); Reprint [Swanson, N.; Swanson, L.W., translators]; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Markram, H.; Muller, E.; Ramaswamy, S.; Reimann, M.W.; Abdellah, M.; Sanchez, C.A.; Ailamaki, A.; Alonso-Nanclares, L.; Antille, N.; Arsever, S.; et al. Reconstruction and simulation of neocortical microcircuitry. Cell 2015, 163, 456–492. [Google Scholar] [CrossRef] [PubMed]
- Lorente de Nó, R. Cerebral cortex: Architectonics, intracortical connections. In Physiology of the Nervous System; Fulton, J.F., Ed.; Oxford University Press: EEUU, New York, NY, USA, 1949; pp. 274–301. [Google Scholar]
- Mountcastle, V.B. Modality and topographic properties of single neurons of cat’s somatic sensory cortex. J. Neurophysiol. 1957, 20, 408–434. [Google Scholar] [CrossRef] [PubMed]
- Marin-Padilla, M. Ontogenesis of the pyramidal cell of the mammalian neocortex and developmental cytoarchitectonic: A unifying theory. J. Comp. Neurol. 1992, 321, 223–240. [Google Scholar] [CrossRef]
- Buxhoeveden, D.P.; Casanova, M.F. The minicolumn hypothesis in neuroscience. Brain 2002, 125, 935–951. [Google Scholar] [CrossRef]
- DeFelipe, J.; Alonso-Nanclares, L.; Arellano, J.I. Microstructure of the neocortex: Comparative aspects. J. Neurocytol. 2002, 31, 299–316. [Google Scholar] [CrossRef]
- Mugniani, E. Cell junctions of astrocytes, ependyma and related cells in the mammalian central nervous system, with emphasis on the hypothesis of a generalized functional syncytium of supporting cells. In Astrocytes: Development, Morphology, and Regional Specialization of Astrocytes; Ferodoff, S., Vernadakis, A., Eds.; Academic Press: London, UK, 1986; pp. 329–371. [Google Scholar]
- Martinez-Banaclocha, M. Neuromagnetic dialogue between neuronal minicolumns and astroglial network: A new approach for memory and cerebral computation. Brain Res. Bull. 2007, 73, 21–27. [Google Scholar] [CrossRef]
- Martinez-Banaclocha, M. Ephaptic coupling of cortical neurons: Possible contribution of astroglial magnetic fields? Neuroscience 2018, 370, 37–45. [Google Scholar] [CrossRef]
- Sun, W.; Cornwell, A.; Li, J.; Peng, S.; Osorio, M.J.; Aalling, N.; Wang, S.; Benraiss, A.; Lou, N.; Goldman, S.A.; et al. SOX9 is an astrocyte-specific nuclear marker in the adult brain outside the neurogenic regions. J. Neurosci. 2017, 37, 4493–4507. [Google Scholar] [CrossRef] [PubMed]
- Santello, M.; Volterra, A. Astrocytes as aide-mémoires. Nature 2010, 463, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Haydon, P.G. Glia: Listening and talking to the synapse. Nat. Neurosci. 2001, 2, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, M.; Ransom, B.; Goldman, S.A. New roles for astrocytes: Redefining the functional architecture of the brain. Trends Neurosci. 2003, 26, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Oberheim, N.A.; Takano, T.; Han, X.; He, W.; Lin, J.H.; Wang, F.; Xu, Q.; Wyatt, J.D.; Pilcher, W.; Ojemann, J.G.; et al. Uniquely hominid features of adult human astrocytes. J. Neurosci. 2009, 29, 3276–3287. [Google Scholar] [CrossRef] [PubMed]
- Marcoli, M.; Agnati, L.; Benedetti, F.; Genedani, S.; Guidolin, D.; Ferraro, L.; Maura, G.; Fuxe, K. On the role of the extracellular space on the holistic behavior of the brain. Rev. Neurosci. 2015, 26, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Lanjakornsiripan, D.; Pior, B.J.; Kawaguchi, D.; Furutachi, S.; Tahara, T.; Katsuyama, Y.; Suzuki, Y.; Fukazawa, Y.; Gotoh, Y. Layer-specific morphological and molecular differences in neocortical astrocytes and their dependence on neuronal layers. Nat. Commun. 2018, 9, 1623. [Google Scholar] [CrossRef]
- Reisin, H.D.; Colombo, J.A. Considerations on the astroglial architecture and the columnar organization of the cerebral cortex. Cell Mol. Neurobiol. 2002, 22, 634–644. [Google Scholar] [CrossRef]
- Eilam, R.; Aharoni, R.; Arnon, R.; Malach, R. Astrocyte morphology is confined by cortical functional boundaries in mammals ranging from mice to human. eLife 2016, 5, e15915. [Google Scholar] [CrossRef]
- Roux, L.; Benchenane, K.; Rothstein, J.D.; Bonvento, G.; Giaume, C. Plasticity of astroglial networks in olfactory glomeruli. Proc. Natl. Acad. Sci. USA 2011, 108, 18442–18446. [Google Scholar] [CrossRef]
- Roux, L.; Madar, A.; Lacroix, M.M.; Yi, C.; Benchenane, K.; Giaume, C. Astroglial connexin 43 hemichannels modulate olfactory bulb slow oscillations. J. Neurosci. 2015, 35, 15339–15352. [Google Scholar] [CrossRef] [PubMed]
- Perez-Alvarez, A.; Navarrete, M.; Covelo, A.; Martin, E.D.; Araque, A. Structural and functional plasticity of astrocyte processes and dendritic spine interactions. J. Neurosci. 2014, 34, 12738–12744, Erratum in: J. Neurosci. 2014, 34, 14163. [Google Scholar] [CrossRef] [PubMed]
- Mederos, S.; González-Arias, C.; Perea, G. Astrocyte-neuron networks: A multilane highway of signaling for homeostatic brain function. Front. Synaptic Neurosci. 2018, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Kang, U.; Chang, H.; Cho, K.-H. The hidden control architecture of complex brain networks. iScience 2019, 13, 154–162. [Google Scholar] [CrossRef]
- Hahn, G.; Skeide, M.A.; Mantini, D.; Ganzetti, M.; Destexhe, A.; Friederici, A.D.; Deco, G. A new computational approach to estimate whole-brain effective connectivity from functional and structural MRI, applied to language development. Sci. Rep. 2019, 9, 8479. [Google Scholar] [CrossRef]
- Reimann, M.W.; Nolte, M.; Scolamiero, M.; Turner, K.; Perin, R.; Chindemi, G.; Dłotko, P.; Levi, R.; Hess, K.; Markram, H. Cliques of neurons bound into cavities provide a missing link between structure and function. Front. Comput. Neurosci. 2017, 11, 48. [Google Scholar] [CrossRef]
- Luczak, A.; McNaughton, B.; Harris, K. Packet-based communication in the cortex. Nat. Rev. Neurosci. 2015, 16, 745–755. [Google Scholar] [CrossRef]
- Demertzi, A.; Tagliazucchi, E.; Dehaene, S.; Deco, G.; Barttfeld, P.; Raimondo, F.; Martial, C.; Fernández-Espejo, D.; Rohaut, B.; Voss, H.U.; et al. Human consciousness is supported by dynamic complex patterns of brain signal coordination. Sci. Adv. 2019, 5, eaat7603. [Google Scholar] [CrossRef]
- Demertzi, A.; Soddu, A.; Laureys, S. Consciousness supporting networks. Curr. Opin. Neurobiol. 2013, 23, 239–244. [Google Scholar] [CrossRef]
- Hutchison, R.M.; Womelsdorf, T.; Allen, E.A.; Bandettini, P.A.; Calhoun, V.D.; Corbetta, M.; Della Penna, S.; Duyn, J.H.; Glover, G.H.; Gonzalez-Castillo, J.; et al. Dynamic functional connectivity: Promise, issues, and interpretations. Neuroimage 2013, 80, 360–378. [Google Scholar] [CrossRef]
- Hahn, G.; Bujan, A.F.; Frégnac, Y.; Aertsen, A.; Kumar, A. Communication through resonance in spiking neuronal networks. Plos Comput. Biol. 2014, 10, e1003811. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Banaclocha, M.; Martinez-Banaclocha, H. Spontaneous neocortical activity and cognitive functions: A neuron-astroglial bio-magnetic and self-organized process. NeuroQuantology 2010, 2010, 191–199. [Google Scholar]
- Martinez-Banaclocha, M. Architectural organization of neuronal activity-associated magnetic fields: A hypothesis for memory. Med. Hypotheses 2004, 63, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Banaclocha, M. Magnetic storage of information in the human cerebral cortex: A hypothesis for memory. Int. J. Neurosci. 2005, 115, 329–337. [Google Scholar]
- Amzica, F.; Steriade, M. Neuronal and glial membrane potentials during sleep and paroxismal oscillations in the neocortex. J. Neurosci. 2000, 20, 6648–6665. [Google Scholar] [CrossRef]
- Araque, A.; Carmignoto, G.; Haydon, P.G. Dynamic signaling between astrocytes and neurons. Ann. Rev. Physiol. 2001, 63, 795–813. [Google Scholar] [CrossRef]
- Amzica, F.; Massimini, M. Glial and neuronal interactions during slow wave and paroxysmal activities in the neocortex. Cereb. Cortex 2002, 12, 1101–1113. [Google Scholar] [CrossRef]
- Seigneur, J.; Kroeger, D.; Nita, D.A.; Amzica, F. Cholinergic action on cortical glial cells in vivo. Cereb. Cortex 2006, 16, 655–668. [Google Scholar] [CrossRef]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.; Robitaille, R.; Volterra, A. Gliotransmitters travel in time and space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef]
- Tian, G.-F.; Azmi, H.; Takano, T.; Xu, Q.; Peng, W.; Lin, J.; Oberheim, N.A.; Lou, N.; Wang, X.; Zielke, H.R.; et al. An astrocytic basis of epilepsy. Nat. Med. 2005, 11, 973–981. [Google Scholar] [CrossRef]
- Witcher, M.R.; Ellis, T.L. Astroglial networks and implications for therapeutic neuromodulation of epilepsy. Front. Comput. Neurosci. 2012, 6, 61. [Google Scholar] [CrossRef]
- Bellot-Saez, A.; Cohen, G.; van Schaik, A.; Ooi, L.; Morley, J.W.; Buskila, Y. Astrocytic modulation of cortical oscillations. Sci. Rep. 2018, 11565. [Google Scholar] [CrossRef]
- Halnes, G.; Mäki-Marttunen, T.; Keller, D.; Pettersen, K.H.; Andreassen, O.A.; Einevoll, G.T. Effect of ionic diffusion on extracellular potentials in neural tissue. Plos Comput. Biol. 2016, 12, e1005193. [Google Scholar] [CrossRef]
- Di Castro, M.; Chuquet, J.; Liaudet, N.; Bhaukaurally, K.; Santello, M.; Bouvier, D.; Tiret, P.; Volterra, A. Local Ca2+ detection and modulation of synaptic release by astrocytes. Nat. Neurosci. 2011, 14, 1276–1284. [Google Scholar] [CrossRef]
- Nett, W.J.; Oloff, S.H.; McCarthy, K.D. Hippocampal astrocytes in situ exhibit calcium oscillations that occur independent of neuronal activity. J. Neurophysiol. 2002, 87, 528–537. [Google Scholar] [CrossRef]
- López-Hidalgo, M.; Kellner, V.; Schummers, J. Astrocyte calcium responses to sensory input: Influence of circuit organization and experimental factors. Front. Neural Circuits 2017, 11, 16. [Google Scholar] [CrossRef]
- Reynolds, J.P.; Zheng, K.; Rusakov, D.A. Multiplexed calcium imaging of single-synapse activity and astroglial responses in the intact brain. Neurosci. Lett. 2019, 689, 26–32. [Google Scholar] [CrossRef]
- López-Hidalgo, M.; Kellner, V.; Schummers, J. Astrocyte subdomains respond independently in vivo. bioRxiv 2019, 675769. [Google Scholar] [CrossRef]
- Wang, M.; He, Y.; Sejnowski, T.J.; Yu, X. Brain-state dependent astrocytic Ca2+ signals are coupled to both positive and negative BOLD-fMRI signals. Proc. Natl. Acad. Sci. USA 2018, 115, E1647–E1656. [Google Scholar] [CrossRef]
- Vöröslakos, M.; Takeuchi, Y.; Brinyiczki, K.; Zombori, T.; Oliva, A.; Fernández-Ruiz, A.; Kozák, G.; Kincses, Z.T.; Iványi, B.; Buzsáki, G.; et al. Direct effects of transcranial electric stimulation on brain circuits in rats and humans. Nat. Commun. 2018, 9, 483. [Google Scholar] [CrossRef]
- Weiss, S.A.; Faber, D.S. Field effects in the CNS play functional roles. Front. Neural Circuit 2010, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, F.; McCormick, D.A. Endogenous electric fields may guide neocortical network activity. Neuron 2010, 67, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Anastassiou, C.A.; Perin, R.; Markram, H.; Koch, C. Ephaptic coupling of cortical neurons. Nat. Neurosci. 2011, 14, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Han, K.S.; Guo, C.; Chen, C.H.; Witter, L.; Osorno, T.; Regehr, W.G. Ephaptic coupling promotes synchronous firing of cerebellar Purkinje cells. Neuron 2018, 100, 564–578. [Google Scholar] [CrossRef] [PubMed]
- Blot, A.; Barbour, B. Ultra-rapid axon-axon ephaptic inhibition of cerebellar Purkinje cells by the pinceau. Nat. Neurosci. 2014, 17, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.-C.; Shivacharan, R.S.; Wei, X.; Gonzalez-Reyes, L.E.; Durand, D.M. Slow periodic activity in the longitudinal hippocampal slice can self-propagate non-synaptically by a mechanism consistent with ephaptic coupling. J. Physiol. 2019, 597, 249–269. [Google Scholar] [CrossRef]
- Lubenov, E.V.; Siapas, A.G. Hippocampal theta oscillations are travelling waves. Nature 2009, 459, 534–539. [Google Scholar] [CrossRef]
- Savtchenko, L.P.; Bard, L.; Jensen, T.P.; Reynolds, J.P.; Kraev, I.; Medvedev, N.; Stewart, M.G.; Henneberger, C.; Rusakov, D.A. Disentangling astroglial physiology with a realistic cell model in silico. Nat. Commun. 2018, 9, 3554. [Google Scholar] [CrossRef]
- Polykretis, I.E.; Ivanov, V.A.; Michmizos, K.P. Computational astrocyence: Astrocytes encode inhibitory activity into the frequency and spatial extent of their calcium elevations. IEEE EMBS Int. Conf. Biomed. Health Inform. (BHI) 2019, 1–4. [Google Scholar] [CrossRef]
- Semyanov, S. Spatiotemporal pattern of calcium activity in astrocytic network. Cell Calcium 2019, 78, 15–25. [Google Scholar] [CrossRef]
- Bar El, Y.; Kanner, S.; Barzilai, A.; Hanein, Y. Activity changes in neuron-astrocyte networks in culture under the effect of norepinephrine. PLoS ONE 2018, 13, e0203761. [Google Scholar] [CrossRef] [PubMed]
- Orkand, R.K.; Nicholls, J.G.; Kuffler, S.W. Effect of nerve impulses on the membrane potential of glial cells in the central nervous system of amphibia. J. Neurophysiol. 1966, 29, 788–806. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Buckalew, R.; Du, Y.; Kiyoshi, C.M.; Alford, C.C.; Wang, W.; McTigue, D.M.; Enyeart, J.J.; Terman, D.; Zhou, M. Gap junction coupling confers isopotentiality on astrocyte syncytium. Glia 2016, 64, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Kiyoshi, C.M.; Du, Y.; Zhong, S.; Wang, W.; Taylor, A.T.; Xiong, B.; Ma, B.; Terman, D.; Zhou, M. Syncytial isopotentiality: A system-wide electrical feature of astrocytic networks in the brain. Glia 2018, 66, 2756–2769. [Google Scholar] [CrossRef]
- Majhi, S.; Ghosha, D. Alternating chimeras in networks of ephaptically coupled bursting neurons. Chaos 2018, 28, 083113. [Google Scholar] [CrossRef]
- Cuffin, B.N.; Cohen, D. Comparison of the magnetoencephalogram and electroencephalogram. Electroencephalogr. Clin. Neurophysiol. 1979, 47, 132–146. [Google Scholar] [CrossRef]
- Gallen, C.C.; Hirschkoff, E.C.; Buchanan, D.S. Magnetoencephalography and magnetic source imaging: Capabilities and limitations. Neuroimaging Clin. N. Am. 1995, 5, 227–249. [Google Scholar]
- Nunez, P.L. The brain’s magnetic field: Some effects of multiple sources on localization methods. Electroencephalogr. Clin. Neurophysiol. 1986, 63, 7–82. [Google Scholar] [CrossRef]
- Hamalainen, M.; Hari, R.; Ilmoniemi, R.J.; Knuutila, J.; Lounasmaa, O.V. Magnetoencephalography: Theory, instrumentation, and applications to noninvasive studies of the working human brain. Rev. Mod. Phys. 1993, 65, 413–497. [Google Scholar] [CrossRef]
- Hall, L.T.; Beart, G.C.G.; Thomas, E.A.; Simpson, D.A.; McGuinness, L.P.; Cole, J.H.; Hollenberg, L.C.L. High spatial and temporal resolution wide-field imaging of neuron activity using quantum NV-diamond. Sci. Rep. 2012, 2, 401. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo. J. Physiol. 1952, 116, 449–472. [Google Scholar] [CrossRef] [PubMed]
- Cragg, B.G. Ultrastructural features of human cerebral cortex. J. Anat. 1976, 121, 331–362. [Google Scholar] [PubMed]
- Thorne, R.G.; Nicholson, C. In vivo diffusion analysis with quantum dots and dextrans predicts the width of brain extracellular space. Proc. Natl. Acad. Sci. USA 2006, 103, 5567–5572. [Google Scholar] [CrossRef]
- Nicholson, C.; Kamali-Zare, P.; Tao, L. Brain extracellular space as a diffusion barrier. Comput. Vis. Sci. 2011, 14, 309–325. [Google Scholar] [CrossRef]
- Lei, Y.; Han, H.; Yuan, F.; Javeed, A.; Zhao, Y. The brain interstitial system: Anatomy, modeling, in vivo measurement, and applications. Prog. Neurobiol. 2017, 157, 230–246. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Adey, W.R. Interaction mechanisms of low-level electromagnetic fields in living systems; Norden, B., Ramel, K., Eds.; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Wang, R.; Fan, Y.; Wu, Y. Spontaneous electromagnetic induction promotes the formation of economical neuronal network structure via self-organization process. Sci. Rep. 2019, 9, 9698. [Google Scholar] [CrossRef] [PubMed]
- Caruso, L.; Wunderle, T.; Lewis, C.M.; Valadeiro, J.; Trauchessec, V.; Rosillo, J.T.; Amaral, J.P.; Ni, J.; Jendritza, P.; Fermon, C.; et al. In vivo magnetic recording of neuronal activity. Neuron 2017, 95, 1283–1291.e4. [Google Scholar] [CrossRef]
- Georgiev, D. Electric and magnetic fields inside neurons and their impact upon the cytoskeletal microtubules. Available online: http://cogprints.org/3190/ (accessed on 12 February 2020).
- Katz, B. Nerve, Muscle and Synapse; McGraw-Hilt: New York, NY, USA, 1966. [Google Scholar]
- Hales, C.G.; Pockett, S. The relationship between local field potentials [LFPs] and the electromagnetic fields that give rise to them. Front. Syst. Neurosci. 2014, 8, 233. [Google Scholar] [CrossRef]
- Vreugdenhil, M.; Bracci, E.; Jefferys, J.G. Layer-specific pyramidal cell oscillations evoked by tetanic stimulation in the rat hippocampal area CA1 in vitro and in vivo. J. Physiol. 2005, 562, 149–164. [Google Scholar] [CrossRef]
- Anastassiou, C.A.; Montgomery, S.M.; Barahona, M.; Buzsaki, G.; Koch, C. The effect of spatially inhomogeneous extracellular electric fields on neurons. J. Neurosci. 2010, 30, 1925–1936. [Google Scholar] [CrossRef]
- Furukawa, T.; Furshpan, E.J. Two inhibitory mechanisms in the Mauthner neurons of goldfish. J. Neurophysiol. 1963, 26, 140–176. [Google Scholar] [CrossRef] [PubMed]
- Faber, D.S.; Korn, H. Field effects trigger post-anodal rebound excitation in vertebrate CNS. Nature 1983, 305, 802–804. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Li, W.C.; Xiao, Q.; Wang, S.R. Electrical interaction between neurons in the pigeon isthmo-optic nucleus. Brain Res. Bull. 2000, 51, 159–163. [Google Scholar] [CrossRef]
- Weiss, S.A.; Preuss, T.; Faber, D.S. A role of electrical inhibition in sensorimotor integration. Proc. Natl. Acad. Sci. 2008, 105, 18047–18052. [Google Scholar] [CrossRef] [PubMed]
- Steriade, M.; Contreras, D.; Curro Dossi, R.; Nunez, A. The slow [<1 Hz] oscillation in reticular thalamic and thalamocortical neurons: Scenario of sleep rhythm generation in interacting thalamic and neocortical networks. J. Neurosci. 1993, 13, 3284–3299. [Google Scholar]
- Ye, H.; Steiger, A. Neuron matters: Electric activation of neuronal tissue is dependent on the interaction between the neuron and the electric field. J. Neuroeng. Rehabil. 2015, 12, 65. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Katz, B. The effect of sodium ions on the electrical activity of the giant axon of the squid. J. Physiol. 1949, 108, 37–77. [Google Scholar] [CrossRef]
- Malmivuo, J.; Plonsey, R. Bioelectromagnetism. Principles and Applications of Bioelectric and Biomagnetic Fields; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Soares, M.A.G.; Cruz, F.A.O.; Silva, D. Magnetic and electric fields across sodium and potassium channels. AIP Conf. Proc. 2015, 1702, 130003. [Google Scholar]
- Ingber, L. Computational algorithms derived from multiple scales of neocortical processing. Cogn. Comput. 2012, 4, 38–50. [Google Scholar] [CrossRef]
- Ingber, L.; Pappalepore, M.; Stesiak, R.R. Electroencephalographic field influence on calcium momentum waves. J. Ther. Biol. 2014, 343, 138–153. [Google Scholar] [CrossRef]
- Reyes-Puerta, V.; Yang, J.; Siwek, M.E.; Kilb, W.; Su, J.-J.; Luhmann, H.J. Propagation of spontaneous slow-wave activity across columns and layers of the adult rat barrel cortex in vivo. Brain Struct. Funct. 2016, 221, 4429–4449. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Du, Y.; Kiyoshi, C.M.; Wu, X.; Askwith, C.C.; McTigue, D.M.; Zhou, M. Syncytial isopotentiality: An electrical feature of spinal cord astrocyte networks. Neuroglia 2018, 1, 271–279. [Google Scholar] [CrossRef]
- Radman, T.; Su, Y.; An, J.H.; Parra, L.C.; Bikson, M. Spike timing amplifies the effect of electric fields on neurons: Implications for endogenous field effects. J. Neurosci. 2007, 27, 3030–3036. [Google Scholar] [CrossRef]
- Manninen, T.; Havela, R.; Linne, M.L. Computational models for calcium-mediated astrocyte functions. Front. Comput. Neurosci. 2018, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; Deneen, B. The emerging nature of astrocyte diversity. Annu Rev Neurosci 2019, 42, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C.; Nedergaard, M. Glial regulation of the cerebral microvasculature. Nat. Neurosci. 2007, 10, 1369–1376. [Google Scholar] [CrossRef]
- Gordon, G.R.; Choi, H.B.; Rungta, R.L.; Ellis-Davies, G.C.; MacVicar, B.A. Brain metabolism dictates the polarity of astrocyte control over arterioles. Nature 2008, 456, 745–749. [Google Scholar] [CrossRef]
- Giaume, C.; Koulakoff, A.; Roux, L.; Holcman, D.; Rouach, N. Astroglial networks: A step further in neuroglial and gliovascular interactions. Nat. Rev. Neurosci. 2010, 11, 87–99. [Google Scholar] [CrossRef]
- Girouard, H.; Bonev, A.D.; Hannah, R.M.; Meredith, A.; Aldrich, R.W.; Nelson, M.T. Astrocytic endfoot Ca2+ and BK channels determine both arteriolar dilation and constriction. Proc. Natl. Acad. Sci. USA 2010, 107, 3811–3816. [Google Scholar] [CrossRef]
- Fiacco, T.A.; Agulhon, C.; Taves, S.R.; Petravicz, J.; Casper, K.B.; Dong, X.; Chen, J.; McCarthy, K.D. Selective stimulation of astrocyte calcium in situ does not affect neuronal excitatory synaptic activity. Neuron 2007, 54, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Agulhon, C.; Fiacco, T.A.; McCarthy, K.D. Hippocampal short- and long-term plasticity are not modulated by astrocyte Ca2+ signaling. Science 2010, 327, 1250–1254. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, N.B.; Attwell, D. Do astrocytes really exocytose neurotransmitters? Nat. Rev. Neurosci. 2010, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Kuga, N.; Sasaki, T.; Takahara, Y.; Matsuki, N.; Ikegaya, Y. Large-scale calcium waves traveling through astrocytic networks in vivo. J. Neurosci. 2011, 31, 2607–2614. [Google Scholar] [CrossRef]
- Scemes, E.; Giaume, C. Astrocyte calcium waves: What they are and what they do. Glia 2006, 54, 716–725. [Google Scholar] [CrossRef]
- Peters, O.; Schipke, C.G.; Hashimoto, Y.; Kettenmann, H. Different mechanisms promote astrocyte Ca2+ waves and spreading depression in the mouse neocortex. J. Neurosci. 2003, 23, 9888–9896. [Google Scholar] [CrossRef]
- Saito, A.; Nakashima, Y.; Shimba, K.; Takayama, Y.; Kotanid, K.; Jimbod, Y. Modulation of neuronal network activity using magnetic nanoparticle-based astrocytic network integration. Biomater. Sci. 2015, 3, 1228–1235. [Google Scholar] [CrossRef]
- Newman, E.A. New roles for astrocytes: Regulation of synaptic transmission. Trends Neurosci. 2003, 26, 536–542. [Google Scholar] [CrossRef]
- De Pittà, M.; Brunel, N.; Volterra, A. Astrocytes: Orchestrating synaptic plasticity? Neuroscience 2016, 323, 43–61. [Google Scholar] [CrossRef]
- Bullock, T.H. Signals and signs in the nervous system: The dynamic anatomy of electrical activity is probably information-rich. Proc. Natl. Acad. Sci. USA 1997, 94, 1–6. [Google Scholar] [CrossRef]
- Deans, J.K.; Powell, A.D.; Jefferys, J.G. Sensitivity of coherent oscillations in rat hippocampus to AC electric fields. J. Physiol. 2007, 583, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Logothetis, N.K.; Kayser, C.; Oeltermann, A. In vivo measurement of cortical impedance spectrum in monkeys: Implications for signal propagation. Neuron 2007, 55, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Massimini, M.; Ferrarelli, F.; Esser, S.K.; Riedner, B.A.; Huber, R.; Murphy, M.; Peterson, M.J.; Tononi, G. Triggering sleep slow waves by transcranial magnetic stimulation. Proc. Natl. Acad. Sci. USA 2007, 104, 8496–8501. [Google Scholar] [CrossRef] [PubMed]
- Richards, P.; Persinger, M.A.; Koren, S.A. Modification of semantic memory in normal subjects by application across the temporal lobes of a weak [1 microT] magnetic field structure that promotes long-term potentiation in hippocampal slices. Eletro. Magn. 1996, 15, 141–148. [Google Scholar] [CrossRef]
- Marshall, L.; Helgadottir, H.; Molle, M.; Born, J. Boosting slow oscillations during sleep potentiates memory. Nature 2006, 444, 610–613. [Google Scholar] [CrossRef]
- Kanai, R.; Chaieb, L.; Antal, A.; Walsh, V.; Paulus, W. Frequency dependent electrical stimulation of the visual cortex. Curr. Biol. 2008, 18, 1839–1843. [Google Scholar] [CrossRef]
- Kirov, R.; Weiss, C.; Siebner, H.R.; Born, J.; Marshall, L. Slow oscillation electrical brain stimulation during waking promotes EEG theta activity and memory encoding. Proc. Natl. Acad. Sci. USA 2009, 106, 15460–15465. [Google Scholar] [CrossRef]
- Ardolino, G.; Bossi, B.; Barbieri, S.; Priori, A. Non-synaptic mechanisms underlie the after-effects of cathodal transcutaneous direct current stimulation of the human brain. J. Physiol. 2005, 568, 653–663. [Google Scholar] [CrossRef]
- Chien, J.H.; Anderson, W. Understanding transcranial magnetic stimulation: A new study of high-temporal-resolution cortical single-neuron responses with extensive artifact reduction. Neurosurgery 2014, 75, N12–N13. [Google Scholar] [CrossRef][Green Version]
- Aspart, F.; Remme, M.W.H.; Obermayer, K. Differential polarization of cortical pyramidal neuron dendrites through weak extracellular fields. PLoS Comput. Biol. 2018, 14, e1006124. [Google Scholar] [CrossRef]
- Cavopol, A.V.; Wamil, A.W.; Holcomb, R.R.; McLean, M.J. Measurement and analysis of static magnetic fields that block action potentials in cultured neurons. Bioelectromagnetics 1995, 16, 197–206. [Google Scholar] [CrossRef]
- Vargas, J.P.; Siegel, J.J.; Bingman, V.P. The effects of a changing ambient magnetic field on single-unit activity in the homing pigeon hippocampus. Brain Res. Bull. 2006, 70, 158–164. [Google Scholar] [CrossRef]
- Cullen, C.L.; Young, K.M. How does transcranial magnetic stimulation influence glial cells in the central nervous system? Front. Neural Circuits 2016, 10, 26. [Google Scholar] [CrossRef]
- Chvatal, A.; Anderova, M.; Ziak, D.; Sykova, E. Glial depolarization evokes a larger potassium accumulation around oligodendrocytes than around astrocytes in gray matter of rat spinal cord slices. J. Neurosci. Res. 1999, 56, 493–505. [Google Scholar] [CrossRef]
- Bikson, M.; Lian, J.; Hahn, P.J.; Stacey, W.C.; Sciortino, C.; Durand, D.M. Suppression of epileptiform activity by high frequency sinusoidal fields in rat hippocampal slices. J. Physiol. 2001, 531, 181–191. [Google Scholar] [CrossRef]
- Ozen, S.; Sirota, A.; Belluscio, M.A.; Anastassiou, C.A.; Stark, E.; Koch, C.; Buzsáki, G. Transcranial electric stimulation entrains cortical neuronal populations in rats. J. Neurosci. 2010, 30, 11476–11485. [Google Scholar] [CrossRef]
- Hirase, H.; Qian, L.; Bartho, P.; Buzsaki, G. Calcium dynamics of cortical astrocytic networks in vivo. PLoS Biol. 2004, 2, e96. [Google Scholar] [CrossRef]
- Poskanzer, K.E.; Yuste, R. Astrocytic regulation of cortical UP states. Proc. Natl. Acad. Sci. USA 2011, 108, 18453–18458. [Google Scholar] [CrossRef]
- Poskanzer, K.E.; Yuste, R. Astrocytes regulate cortical state switching in vivo. Proc. Natl. Acad. Sci. USA 2016, 113, E2675–E2684. [Google Scholar] [CrossRef]
- Pannasch, U.; Rouach, N. Emerging role for astroglial networks in information processing: From synapse to behavior. Trends Neurosci. 2013, 36, 405–417. [Google Scholar] [CrossRef]
- Ruohonen, J.; Karhu, J. tDCS possibly stimulates glial cells. Clin. Neurophysiol. 2012, 123, 2006–2009. [Google Scholar] [CrossRef] [PubMed]
- Monai, H.; Ohkura, M.; Tanaka, M.; Oe, Y.; Konno, A.; Hirai, H.; Mikoshiba, K.; Itohara, S.; Nakai, J.; Iwai, Y.; et al. Calcium imaging reveals glial involvement in transcranial direct current stimulation-induced plasticity in mouse brain. Nat. Commun. 2016, 7, 11100. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, A.; Mordillo-Mateos, L.; Arias, P.; Panyavin, I.; Foffani, G.; Aguilar, J. Transcranial static magnetic field stimulation of the human motor cortex. J. Physiol. 2011, 589, 4949–4958. [Google Scholar] [CrossRef] [PubMed]
- Fontenele, A.J.; de Vasconcelos, N.A.P.; Feliciano, T.; Aguiar, L.A.A.; Soares-Cunha, C.; Coimbra, B.; Dalla Porta, L.; Ribeiro, S.; Rodrigues, A.J.; Sousa, N.; et al. Criticality between cortical states. Phys. Rev. Lett. 2019, 122, 208101. [Google Scholar] [CrossRef] [PubMed]
- Hahn, G.; Ponce-Alvarez, A.; Monier, C.; Benvenuti, G.; Kumar, A.; Chavane, F.; Deco, G.; Frégnac, Y. Spontaneous cortical activity is transiently poised close to criticality. PLoS Comput. Biol. 2017, 13, e1005543. [Google Scholar] [CrossRef] [PubMed]
- Wilting, J.; Priesemann, V. Between perfectly critical and fully irregular: A reverberating model captures and predicts cortical spike propagation. Cereb. Cortex 2019, 29, 2759–2770. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, S.; Kang, C.; Hwang, S.; Lee, Y.; Yu, K. Toward a brain functional connectivity mapping modality by simultaneous imaging of coherent brainwaves. Neuroimage 2014, 91, 63–69. [Google Scholar] [CrossRef][Green Version]
- Barbieri, F.; Trauchessec, V.; Caruso, L.; Trejo-Rosillo, J.; Telenczuk, B.; Paul, E.; Bal, T.; Destexhe, A.; Fermon, C.; Pannetier-Lecoeur, M.; et al. Local recording of biological magnetic fields using Giant Magneto Resistance-based micro-probes. Sci. Rep. 2016, 6, 39330. [Google Scholar] [CrossRef]




© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Banaclocha, M. Astroglial Isopotentiality and Calcium-Associated Biomagnetic Field Effects on Cortical Neuronal Coupling. Cells 2020, 9, 439. https://doi.org/10.3390/cells9020439
Martinez-Banaclocha M. Astroglial Isopotentiality and Calcium-Associated Biomagnetic Field Effects on Cortical Neuronal Coupling. Cells. 2020; 9(2):439. https://doi.org/10.3390/cells9020439
Chicago/Turabian StyleMartinez-Banaclocha, Marcos. 2020. "Astroglial Isopotentiality and Calcium-Associated Biomagnetic Field Effects on Cortical Neuronal Coupling" Cells 9, no. 2: 439. https://doi.org/10.3390/cells9020439
APA StyleMartinez-Banaclocha, M. (2020). Astroglial Isopotentiality and Calcium-Associated Biomagnetic Field Effects on Cortical Neuronal Coupling. Cells, 9(2), 439. https://doi.org/10.3390/cells9020439