Cannabis Sativa Revisited—Crosstalk between microRNA Expression, Inflammation, Oxidative Stress, and Endocannabinoid Response System in Critically Ill Patients with Sepsis


 ,
,
Abstract
1. Introduction
2. Pathophysiological and Clinical Aspects in Sepsis
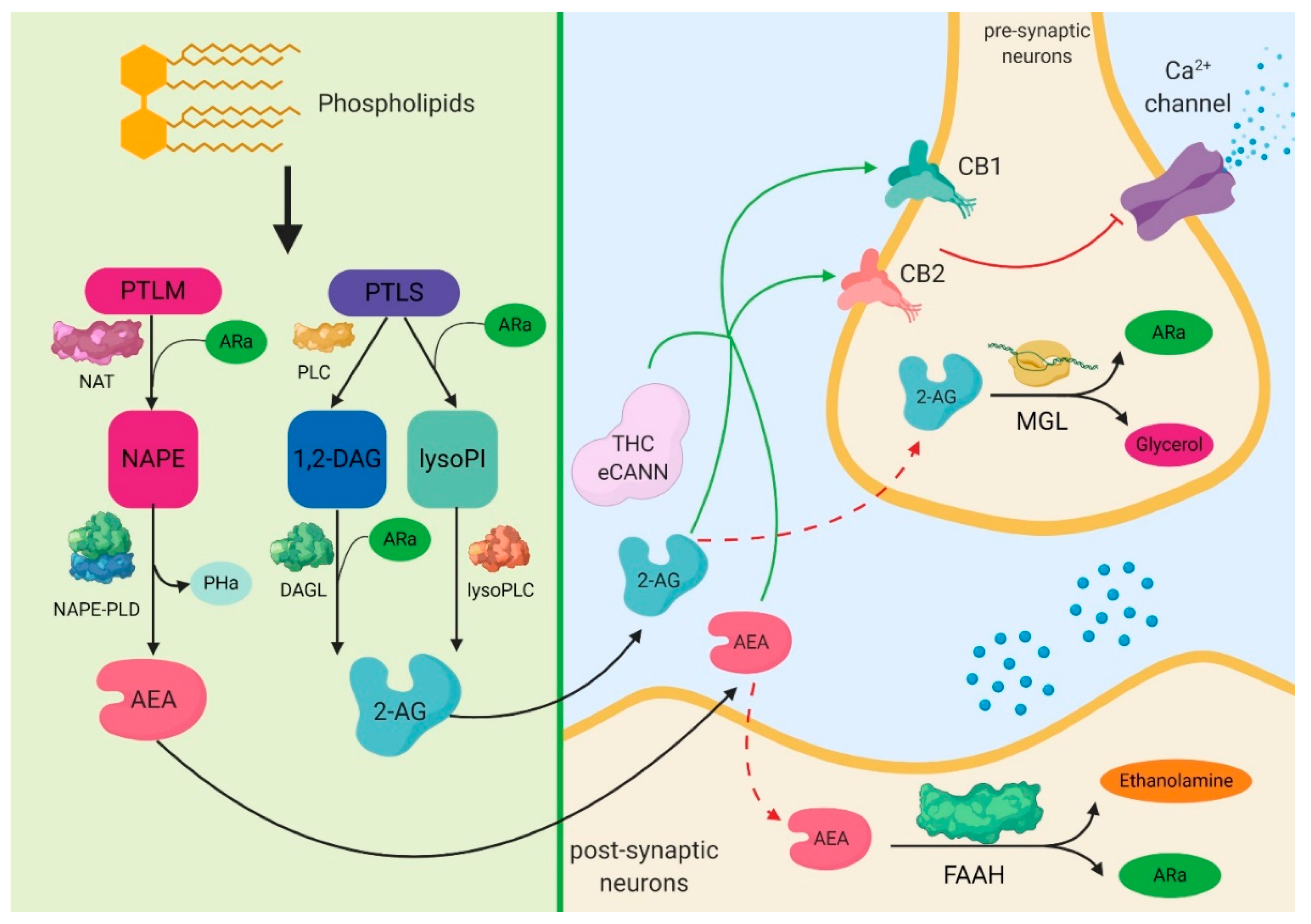
3. Biochemical Aspects of Cannabinoids
4. The Expression of Cannabinoid Signaling System in Sepsis
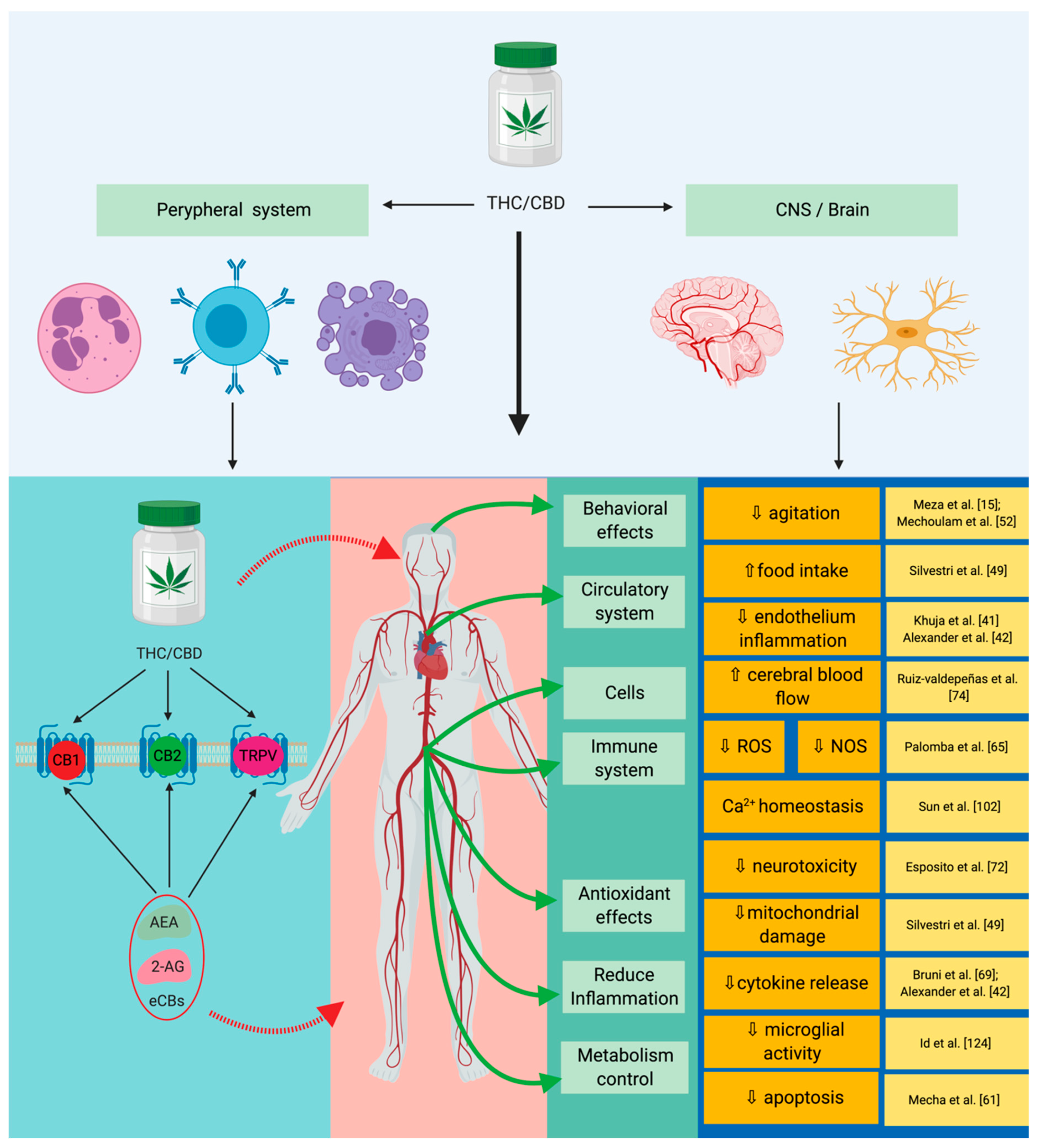
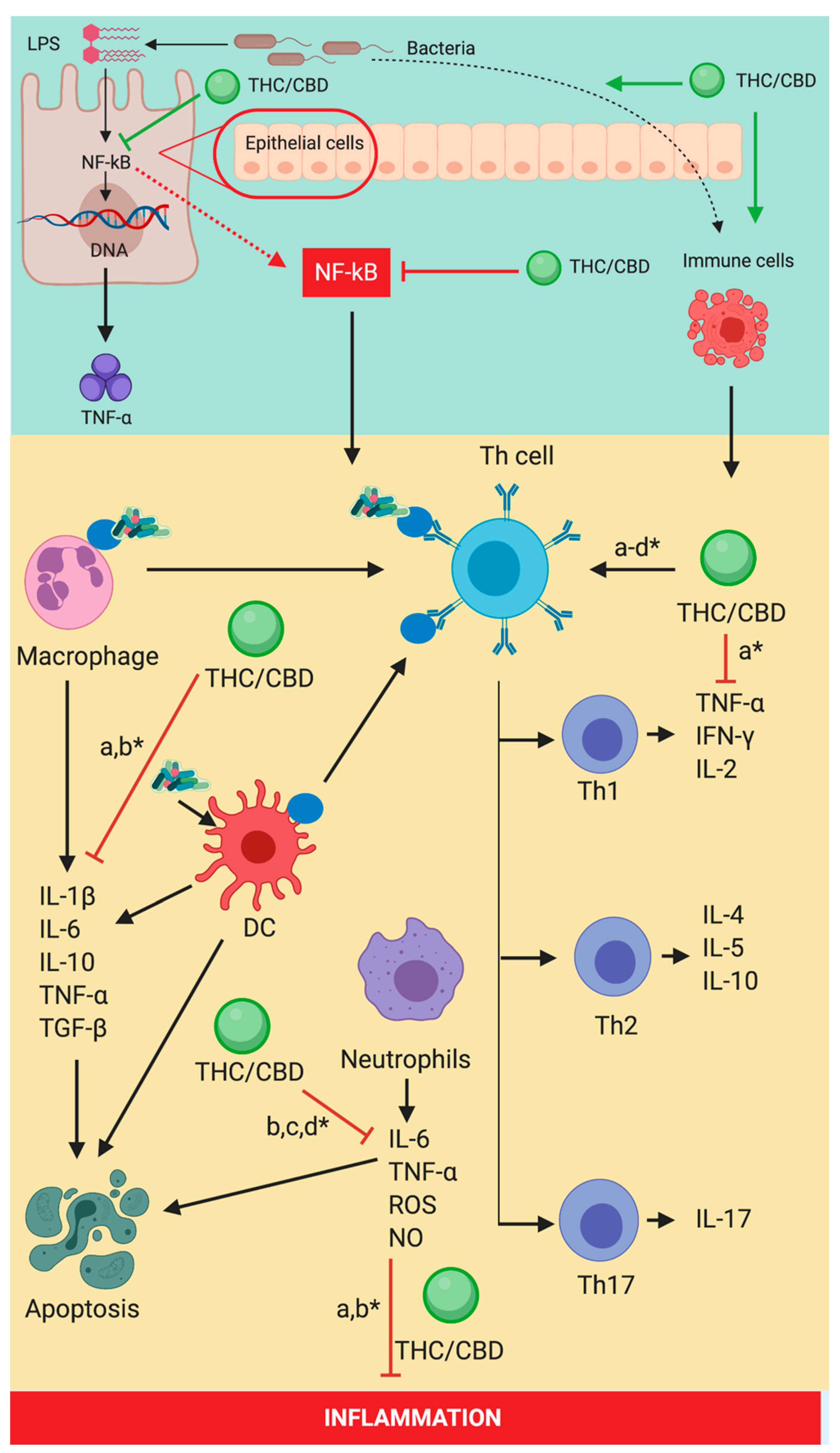
5. The Cannabinoid Signaling System and Inflammation-Linked with Sepsis
6. Cannabinoid Signaling System and Redox Activity-Linked with Sepsis
7. MicroRNAs Expression in Sepsis-Induced by Cannabinoid Signaling System
8. Immune System Expression-Induced by Cannabinoid Signaling System
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Angus, D.C.; van der Poll, T.; Wacker, D.A.; Winters, M.E.; Finfer, S.R.; Vincent, J.-L.; De Backer, D.; Martin, G.S.; Mutschler, M.; Paffrath, T.; et al. Severe sepsis and septic shock. Emerg. Med. Clin. N. Am. 2013, 45 (Suppl. 3), 747–758. [Google Scholar] [CrossRef] [PubMed]
- Coelho, F.R.; Martins, J.O. Diagnostic methods in sepsis: The need of speed. Rev. Assoc. Med. Bras. 2012, 58, 498–504. [Google Scholar] [CrossRef]
- David, V.L.; Ercisli, F.; Florin, A.; Boia, E.S.; Nitu, R. Early Prediction of Sepsis Incidence in Critically Ill Patients Using Specific Genetic Polymorphisms. Biochem. Genet. 2017, 55, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E. NF-κB activation. Crit. Care Med. 2000, 28, N100–N104. [Google Scholar] [CrossRef]
- Cimolai, M.C.; Alvarez, S.; Bode, C.; Bugger, H. Mitochondrial Mechanisms in Septic Cardiomyopathy. Int. J. Mol. Sci. 2015, 16, 17763–17778. [Google Scholar] [CrossRef]
- Horhat, F.G.; Gundogdu, F.; David, L.V.; Boia, E.S.; Pirtea, L.; Horhat, R.; Cucui-Cozma, A.; Ciuca, I.; Diaconu, M.; Nitu, R.; et al. Early Evaluation and Monitoring of Critical Patients with Acute Respiratory Distress Syndrome (ARDS) Using Specific Genetic Polymorphisms. Biochem. Genet. 2017, 55, 204–211. [Google Scholar] [CrossRef]
- Koekkoek, W.A.C.; van Zanten, A.R.H. Antioxidant Vitamins and Trace Elements in Critical Illness. Nutr. Clin. Pract. 2016, 31, 457–474. [Google Scholar] [CrossRef]
- Akinosoglou, K.; Alexopoulos, D. Use of antiplatelet agents in sepsis: A glimpse into the future. Thromb. Res. 2014, 133, 131–138. [Google Scholar] [CrossRef]
- Pop-Began, V.; Păunescu, V.; Grigorean, V.; Pop-Began, D.; Popescu, C. Molecular mechanisms in the pathogenesis of sepsis. J. Med. Life 2014, 7, 38–41. [Google Scholar]
- Xie, L.X. New biomarkers for sepsis. Med. J. Chin. People’s Lib. Army 2013, 38, 6–9. [Google Scholar]
- Bartz, R.R.; Fu, P.; Suliman, H.B.; Crowley, S.D.; MacGarvey, N.C.; Welty-Wolf, K.; Piantadosi, C.A. Staphylococcus aureus sepsis induces early renal mitochondrial DNA repair and mitochondrial biogenesis in mice. PLoS ONE 2014, 9, e100912. [Google Scholar] [CrossRef] [PubMed]
- Id, A.S.; Theilla, M.; Hellerman, M.; Singer, P.; Maggiore, U.; Barbagallo, M.; Regolisti, G.; Fiaccadori, E. Energy and Protein in Critically Ill Patients with AKI: A Prospective, Multicenter Observational Study Using Indirect Calorimetry and Protein Catabolic Rate. Nutrients 2017, 9, 802. [Google Scholar]
- Daniel Lafreniere, J.; Lehmann, C. Parameters of the endocannabinoid system as novel biomarkers in sepsis and septic shock. Metabolites 2017, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Rogobete, A.F.; Sandesc, D.; Bedreag, O.H.; Papurica, M.; Popovici, S.E.; Bratu, T.; Popoiu, C.M.; Nitu, R.; Dragomir, T.; AAbed, H.I.M.; et al. MicroRNA Expression is Associated with Sepsis Disorders in Critically Ill Polytrauma Patients. Cells 2018, 7, 271. [Google Scholar] [CrossRef] [PubMed]
- Meza, A.; Lehmann, C. Betacaryophyllene—A phytocannabinoid as potential therapeutic modality for human sepsis? Med. Hypotheses 2018, 110, 68–70. [Google Scholar] [CrossRef]
- Chiarlone, A.; Börner, C.; Martín-Gómez, L.; Jiménez-González, A.; García-Concejo, A.; García-Bermejo, M.L.; Lorente, M.; Blázquez, C.; García-Taboada, E.; de Haro, A.; et al. MicroRNA let-7d is a target of cannabinoid CB1 receptor and controls cannabinoid signaling. Neuropharmacology 2016, 108, 345–352. [Google Scholar] [CrossRef]
- Fox, E.D.; Heffernan, D.S.; Cioffi, W.G.; Reichner, J.S. Neutrophils from critically ill septic patients mediate profound loss of endothelial barrier integrity. Crit. Care 2013, 17, R226. [Google Scholar] [CrossRef]
- Gu, W.; Jiang, J. Genetic polymorphisms and posttraumatic complications. Comp. Funct. Genom. 2010, 2010. [Google Scholar] [CrossRef]
- Wenceslau, C.F.; McCarthy, C.G.; Goulopoulou, S.; Szasz, T.; NeSmith, E.G.; Webb, R.C. Mitochondrial-derived N-formyl peptides: Novel links between trauma, vascular collapse and sepsis. Med. Hypotheses 2013, 81, 532–535. [Google Scholar] [CrossRef]
- Huber-Lang, M.; Kovtun, A.; Ignatius, A. The role of complement in trauma and fracture healing. Semin. Immunol. 2013, 25, 73–78. [Google Scholar] [CrossRef]
- Sun, S.; Sursal, T.; Adibnia, Y.; Zhao, C.; Zheng, Y.; Li, H.; Otterbein, L.E.; Hauser, C.J.; Itagaki, K. Mitochondrial DAMPs Increase Endothelial Permeability through Neutrophil Dependent and Independent Pathways. PLoS ONE 2013, 8, e59989. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Chaiworapongsa, T.; Alpay Savasan, Z.; Xu, Y.; Hussein, Y.; Dong, Z.; Kusanovic, J.P.; Kim, C.J.; Hassan, S.S. Damage-associated molecular patterns (DAMPs) in preterm labor with intact membranes and preterm PROM: A study of the alarmin HMGB1. J. Matern.-Fetal Neonatal Med. 2011, 24, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Bronkhorst, M.W.G.A.; Boyé, N.D.A.; Lomax, M.A.Z.; Vossen, R.H.A.M.; Bakker, J.; Patka, P.; Van Lieshout, E.M.M. Single-nucleotide polymorphisms in the Toll-like receptor pathway increase susceptibility to infections in severely injured trauma patients. J. Trauma Acute Care Surg. 2013, 74, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, A.M.; Walley, K.R.; Russell, J.A. Polymorphisms in CD14, mannose-binding lectin, and Toll-like receptor-2 are associated with increased prevalence of infection in critically ill adults*. Crit. Care Med. 2005, 33, 638–644. [Google Scholar] [CrossRef]
- Szilágyi, B.; Fejes, Z.; Pócsi, M.; Kappelmayer, J.; Nagy, B., Jr. Role of sepsis modulated circulating microRNAs. EJIFCC 2019, 30, 128–145. [Google Scholar]
- Davis, S.M.; Clark, E.A.S.; Nelson, L.T.; Silver, R.M. The association of innate immune response gene polymorphisms and puerperal group a streptococcal sepsis. Am. J. Obstet. Gynecol. 2010, 202, 308 e1–308 e8. [Google Scholar] [CrossRef]
- Stanilova, S.A.; Miteva, L.D.; Karakolev, Z.T.; Stefanov, C.S. Interleukin-10-1082 promoter polymorphism in association with cytokine production and sepsis susceptibility. Intensive Care Med. 2006, 32, 260–266. [Google Scholar] [CrossRef]
- Unit, I.C.; Paz, H. La Interleukin-1 receptor antagonist gene polymorphism and mortality in patients with severe sepsis. Clin. Exp. Immunol. 2002, 127, 331–336. [Google Scholar]
- Das, U.N. Serum adipocyte fatty acid-binding protein in the critically ill. Crit. Care 2013, 17, 121. [Google Scholar] [CrossRef][Green Version]
- Kołakowska, B. Nutrition challenges in polytrauma patients. New trends in energy expenditure measurements. Central European J. of Clin. Res. 2019, 2, 51–57. [Google Scholar] [CrossRef]
- Pravda, J. Metabolic theory of septic shock. World J. Crit. Care Med. 2014, 3, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, M.C.; Davies, D.C.; Moss, R.F.; Tighe, D.; Bennett, E.D. Pathophysiology of septic encephalopathy: A review. Crit. Care Med. 2000, 28, 3019–3024. [Google Scholar] [CrossRef] [PubMed]
- Malbrain, M.L.N.G.; Marik, P.E.; Witters, I.; Cordemans, C.; Kirkpatrick, A.W.; Roberts, D.J.; Van Regenmortel, N. Fluid overload, de-resuscitation, and outcomes in critically ill or injured patients: A systematic review with suggestions for clinical practice. Anaesthesiol. Intensive Ter. 2014, 46, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Oudemans-van Straaten, H.M.; Man, A.M.S.; de Waard, M.C. Vitamin C revisited. Crit. Care 2014, 18, 460. [Google Scholar] [CrossRef]
- Gill, S.E.; Rohan, M.; Mehta, S. Role of pulmonary microvascular endothelial cell apoptosis in murine sepsis-induced lung injury in vivo. Respir. Res. 2015, 16, 109. [Google Scholar] [CrossRef] [PubMed]
- Biron, B.M.; Ayala, A.; Lomas-neira, J.L. Biomarkers for Sepsis: What Is and What Might Be? Biomark Insights. 2015, 10, 7–17. [Google Scholar] [CrossRef]
- Binkowska, A.M.; Michalak, G.; Słotwiński, R. Current views on the mechanisms of immune responses to trauma and infection. Cent.-Eur. J. Immunol. 2015, 40, 206–216. [Google Scholar] [CrossRef]
- Leite, H.P.; de Lima, L.F.P. Metabolic resuscitation in sepsis: A necessary step beyond the hemodynamic? J. Thorac. Dis. 2016, 8, E552–E557. [Google Scholar] [CrossRef]
- Arlati, S.; Storti, E.; Pradella, V.; Bucci, L.; Vitolo, A.; Pulici, M. Decreased fluid volume to reduce organ damage: A new approach to burn shock resuscitation? A preliminary study. Resuscitation 2007, 72, 371–378. [Google Scholar] [CrossRef]
- Iseppi, R.; Brighenti, V.; Licata, M.; Lambertini, A.; Sabia, C.; Messi, P.; Pellati, F.; Benvenuti, S. Chemical characterization and evaluation of the antibacterial activity of essential oils from fibre-type cannabis sativa L. (Hemp). Molecules 2019, 24, 7–12. [Google Scholar] [CrossRef]
- Khuja, I.; Yekhtin, Z.; Or, R.; Almogi-Hazan, O. Cannabinoids reduce inflammation but inhibit lymphocyte recovery in murine models of bone marrow transplantation. Int. J. Mol. Sci. 2019, 20, 668. [Google Scholar] [CrossRef] [PubMed]
- Alexander, A.; Smith, P.F.; Rosengren, R.J. Cannabinoids in the treatment of cancer. Cancer Lett. 2009, 285, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiu, V.; Leuti, A.; Cencioni, M.T.; Albanese, M.; De Bardi, M.; Bisogno, T.; Centonze, D.; Battistini, L.; Maccarrone, M. Modulation of monocytes by bioactive lipid anandamide in multiple sclerosis involves distinct Toll-like receptors. Pharmacol. Res. 2016, 113, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Leishman, E.; Murphy, M.; Mackie, K.; Bradshaw, H.B. BBA—Molecular and Cell Biology of Lipids Δ9-Tetrahydrocannabinol changes the brain lipidome and transcriptome di ff erentially in the adolescent and the adult. BBA-Mol. Cell Biol. Lipids 2018, 1863, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiu, V.; Cencioni, M.T.; Bisicchia, E.; De Bardi, M.; Gasperini, C.; Borsellino, G.; Centonze, D.; Battistini, L.; Maccarrone, M. Distinct modulation of human myeloid and plasmacytoid dendritic cells by anandamide in multiple sclerosis. Ann. Neurol. 2013, 73, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Matthews, A.T.; Ross, M.K. Oxyradical Stress, Endocannabinoids, and Atherosclerosis. Toxics 2015, 3, 481–498. [Google Scholar] [CrossRef] [PubMed]
- Chanda, D.; Kim, D.-K.; Li, T.; Kim, Y.-H.; Koo, S.-H.; Lee, C.-H.; Chiang, J.Y.L.; Choi, H.-S. Cannabinoid receptor type 1 (CB1R) signaling regulates hepatic gluconeogenesis via induction of endoplasmic reticulum-bound transcription factor cAMP-responsive element-binding protein H (CREBH) in primary hepatocytes. J. Biol. Chem. 2011, 286, 27971–27979. [Google Scholar] [CrossRef]
- Bisogno, T.; Ligresti, A.; Di Marzo, V. The endocannabinoid signalling system: Biochemical aspects. Pharmacol. Biochem. Behav. 2005, 81, 224–238. [Google Scholar] [CrossRef]
- Silvestri, C.; Di Marzo, V. The Endocannabinoid System in Energy Homeostasis and the Etiopathology of Metabolic Disorders. Cell Metab. 2013, 17, 475–490. [Google Scholar] [CrossRef]
- Maione, S.; Costa, B.; Di Marzo, V. Endocannabinoids: A unique opportunity to develop multitarget analgesics. PAIN 2013, 154, S87–S93. [Google Scholar] [CrossRef]
- Di Marzo, V. Targeting the endocannabinoid system: To enhance or reduce? Nat. Rev. Drug Discov. 2008, 7, 438–455. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Parker, L.A. The Endocannabinoid System and the Brain. Annu. Rev. Psychol. 2013, 64, 21–47. [Google Scholar] [CrossRef] [PubMed]
- McHugh, D.; Tanner, C.; Mechoulam, R.; Pertwee, R.G.; Ross, R.A. Inhibition of Human Neutrophil Chemotaxis by Endogenous Cannabinoids and Phytocannabinoids: Evidence for a Site Distinct from CB1 and CB2. Mol. Pharmacol. 2008, 73, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Tabarkiewicz, J. Endocannabinoid system as a regulator of tumor cell malignancy—Biological pathways and clinical significance. Onco Targets Ther. 2016, 9, 4323–4336. [Google Scholar]
- Gardner, E.L. Endocannabinoid signaling system and brain reward: Emphasis on dopamine. Pharmacol. Biochem. Behav. 2005, 81, 263–284. [Google Scholar] [CrossRef]
- Watkins, B.A.; Hutchins, H.; Li, Y.; Seifert, M.F. The endocannabinoid signaling system: A marriage of PUFA and musculoskeletal health. J. Nutr. Biochem. 2010, 21, 1141–1152. [Google Scholar] [CrossRef]
- Dogjani, A.; Zatriqi, S.; Uranues, S.; Latifi, R. Biology-based nutritional support of critically ill and injured patients. Eur. Surg. 2011, 43, 7–12. [Google Scholar] [CrossRef]
- Vallejo, K.P.; Martínez, C.M.; Adames, A.A.M.; Fuchs-tarlovsky, V.; Carlos, G.; Nogales, C.; Enrique, R.; Paz, R.; Perman, M.I.; Isabel, M.; et al. Current clinical nutrition practices in critically ill patients in Latin America: A multinational observational study. Crit. Care 2017, 21, 227. [Google Scholar] [CrossRef]
- Maday, K.R. Energy Estimation in the Critically Ill: A Literature Review. Int. J. Clin. Med. 2013, 1, 39–43. [Google Scholar]
- Rogobete, A.F.; Sandesc, D.; Papurica, M.; Stoicescu, E.R.; Popovici, S.E.; Bratu, L.M.; Vernic, C.; Sas, A.M.; Stan, A.T.; Bedreag, O.H. The influence of metabolic imbalances and oxidative stress on the outcome of critically ill polytrauma patients: A review. Burn. Trauma 2017, 5, 8. [Google Scholar] [CrossRef]
- Mecha, M.; Torrao, A.S.; Mestre, L.; Carrillo-salinas, F.J.; Mechoulam, R.; Guaza, C. Cannabidiol protects oligodendrocyte progenitor cells from inflammation-induced apoptosis by attenuating endoplasmic reticulum stress. Cell Death Discov. 2012, 3, e331. [Google Scholar] [CrossRef]
- Simon, L.; Song, K.; Stouwe, C.V.; Hollenbach, A.; Amedee, A.; Mohan, M.; Winsauer, P.; Molina, P. Δ9-Tetrahydrocannabinol (Δ9-THC) Promotes Neuroimmune-Modulatory MicroRNA Profile in Striatum of Simian Immunodeficiency Virus (SIV)-Infected Macaques. J. Neuroimmune Pharmacol. 2016, 11, 192–213. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Peters, M.; Murillo-Rodriguez, E.; Hanus, L.O. Cannabidiol--recent advances. Chem. Biodivers. 2007, 4, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- Brown, I.; Cascio, M.G.; Rotondo, D.; Pertwee, R.G.; Heys, S.D.; Wahle, K.W.J. Cannabinoids and omega-3/6 endocannabinoids as cell death and anticancer modulators. Prog. Lipid Res. 2013, 52, 80–109. [Google Scholar] [CrossRef]
- Palomba, L.; Silvestri, C.; Imperatore, R.; Morello, G.; Piscitelli, F.; Martella, A.; Cristino, L.; Marzo, V. Di Negative Regulation of Leptin-induced Reactive Oxygen Species (ROS) Formation by Cannabinoid CB 1 Receptor Activation in Hypothalamic Neurons. J. Biol. Chem. 2015, 290, 13669–13677. [Google Scholar] [CrossRef]
- Lau, F.C.; Bagchi, M.; Sen, C.; Roy, S.; Bagchi, D. Nutrigenomic Analysis of Diet-Gene Interactions on Functional Supplements for Weight Management. Curr. Genom. 2008, 9, 239–251. [Google Scholar] [CrossRef][Green Version]
- Endocannabinoids, S.C.; Mukhopadhyay, P.; Horiguchi, N.; Jeong, W.; Osei-hyiaman, D.; Park, O.; Liu, J.; Harvey-white, J.; Marsicano, G.; Lutz, B.; et al. Paracrine Activation of Hepatic CB1 Receptors Mediates Alcoholic Fatty Liver. Cell. Metab. 2008, 7, 227–235. [Google Scholar]
- Prester, L.; Mikoli, A.; Juri, A.; Fuchs, N.; Neuberg, M.; Luci, A.; Br, I. Chemico-Biological Interactions Effects of Δ9-tetrahydrocannabinol on irinotecan-induced clinical effects in rats. Chem.-Biol. Interact. 2018, 294, 128–134. [Google Scholar] [CrossRef]
- Bruni, N.; Della Pepa, C.; Oliaro-Bosso, S.; Pessione, E.; Gastaldi, D.; Dosio, F. Cannabinoid delivery systems for pain and inflammation treatment. Molecules 2018, 23, 2478. [Google Scholar] [CrossRef]
- Ladak, N.; Beishon, L.; Thompson, J.P.; Lambert, D.G. Trends in Anaesthesia and Critical Care Cannabinoids and sepsis. Trends Anaesth. Crit. Care 2011, 1, 191–198. [Google Scholar] [CrossRef]
- Malfait, A.M.; Gallily, R.; Sumariwalla, P.F.; Malik, A.S.; Andreakos, E.; Mechoulam, R.; Feldmann, M. The nonpsychoactive cannabis constituent cannabidiol is an oral anti-arthritic therapeutic in murine collagen-induced arthritis. Proc. Natl. Acad. Sci. USA 2000, 97, 9561–9566. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Scuderi, C.; Valenza, M.; Togna, G.I.; Latina, V.; Iuvone, T.; Steardo, L.; De Filippis, D.; Cipriano, M.; Carratu, M.R. Cannabidiol Reduces Aβ-Induced Neuroinflammation and Promotes Hippocampal Neurogenesis through PPARγ Involvement. PLoS ONE 2011, 6, e28668. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.M.; Homsi, C.; Ferreira, J.; Mateus-vasconcelos, E.C.L.; Moroni, R.M.; Maria, L.; Brito, O.; Gustavo, L.; Brito, O. Case Report Physical Therapy in the Management of Pelvic Floor Muscles Hypertonia in a Woman with Hereditary Spastic Paraplegia. Case Rep. Obstet. Gynecol. 2014, 2014, 306028. [Google Scholar]
- Ruiz-valdepeñas, L.; Martínez-orgado, J.A.; Benito, C.; Millán, Á.; Tolón, R.M. Cannabidiol reduces lipopolysaccharide-induced vascular changes and inflammation in the mouse brain: An intravital microscopy study Cannabidiol reduces lipopolysaccharide-induced vascular changes and inflammation in the mouse brain: An intravital micros. J. Neuroinflamm. 2011, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Kaplan, B.L.F.; Pike, S.T.; Topper, L.A.; Lichorobiec, N.R.; Simmons, S.O.; Ramabhadran, R.; Kaminski, N.E. Magnitude of stimulation dictates the cannabinoid-mediated differential T cell response to HIVgp120. J. Leukoc. Biol. 2012, 92, 1093–1102. [Google Scholar] [CrossRef]
- Cassol, O.J., Jr.; Comim, C.M.; Silva, B.R.; Hermani, F.V.; Constantino, L.S.; Felisberto, F.; Petronilho, F.; Hallak, J.E.C.; De Martinis, B.S.; Zuardi, A.W.; et al. Treatment with cannabidiol reverses oxidative stress parameters, cognitive impairment and mortality in rats submitted to sepsis by cecal ligation and puncture. Brain Res. 2010, 1348, 128–138. [Google Scholar] [CrossRef]
- Vuolo, F.; Petronilho, F.; Sonai, B.; Ritter, C.; Hallak, J.E.C.; Zuardi, A.W.; Crippa, J.A.; Dal-pizzol, F. Evaluation of Serum Cytokines Levels and the Role of Cannabidiol Treatment in Animal Model of Asthma. Mediat. Inflamm. 2015, 2015, 538670. [Google Scholar] [CrossRef]
- Nissen, L.; Zatta, A.; Stefanini, I.; Grandi, S.; Sgorbati, B.; Biavati, B.; Monti, A. Characterization and antimicrobial activity of essential oils of industrial hemp varieties (Cannabis sativa L.). Fitoterapia 2010, 81, 413–419. [Google Scholar] [CrossRef]
- Hernández-cervantes, R.; Méndez-díaz, M.; Prospéro-garcía, Ó. Immunoregulatory Role of Cannabinoids during Infectious Disease. Neuroimmunomodulation 2017, 24, 183–199. [Google Scholar] [CrossRef]
- Wasim, K.; Haq, I.; Ashraf, M. Antimicrobial studies of the leaf of cannabis sativa L. Pak. J. Pharm. Sci. 1995, 8, 29–38. [Google Scholar]
- Elphick, M.R. BfCBR: A cannabinoid receptor ortholog in the cephalochordate Branchiostoma floridae (Amphioxus). Gene 2007, 399, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Appendino, G.; Gibbons, S.; Giana, A.; Pagani, A.; Grassi, G.; Stavri, M.; Smith, E.; Rahman, M.M. Antibacterial cannabinoids from Cannabis sativa: A structure-activity study. J. Nat. Prod. 2008, 71, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Bass, R.; Engelhard, D.; Trembovler, V.; Shohami, E. A novel nonpsychotropic cannabinoid, HU-211, in the treatment of experimental pneumococcal meningitis. J. Infect. Dis. 1996, 173, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Bisoi, S.; Chattopadhyay, D.; Mishra, R. A Study on Demographic and Clinical Profile of Burn Patients in an Apex Institute of West Bengal. Indian J. Public Health. 2010, 54, 92–94. [Google Scholar]
- Bellocchio, L.; Lafenetre, P.; Cannich, A.; Cota, D.; Puente, N.; Grandes, P.; Chaouloff, F.; Piazza, P.V.; Marsicano, G. Bimodal control of stimulated food intake by the endocannabinoid system. Nat. Neurosci. 2010, 13, 281–283. [Google Scholar] [CrossRef]
- Sardinha, J.; Kelly, M.E.M.; Zhou, J.; Lehmann, C. Experimental Cannabinoid 2 Receptor-Mediated Immune Modulation in Sepsis. Mediat. Inflamm. 2014, 2014, 978678. [Google Scholar] [CrossRef]
- Wang, L.-L.; Zhao, R.; Li, J.-Y.; Li, S.-S.; Liu, M.; Wang, M.; Zhang, M.-Z.; Dong, W.-W.; Jiang, S.-K.; Zhang, M.; et al. Pharmacological activation of cannabinoid 2 receptor attenuates inflammation, fibrogenesis, and promotes re-epithelialization during skin wound healing. Eur. J. Pharmacol. 2016, 786, 128–136. [Google Scholar] [CrossRef]
- Eisenstein, T.K.; Meissler, J.J. Effects of Cannabinoids on T-cell Function and Resistance to Infection. J. Neuroimmune Pharmacol. 2015, 10, 204–216. [Google Scholar] [CrossRef]
- Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.L.; Vitoretti, L.B.; Mariano-Souza, D.P.; Quinteiro-Filho, W.M.; Akamine, A.T.; Almeida, V.I.; Quevedo, J.; Dal-Pizzol, F.; et al. Cannabidiol, a non-psychotropic plant-derived cannabinoid, decreases inflammation in a murine model of acute lung injury: Role for the adenosine A2A receptor. Eur. J. Pharmacol. 2012, 678, 78–85. [Google Scholar] [CrossRef]
- Obeid, R.; Herrmann, W. Mechanisms of homocysteine neurotoxicity in neurodegenerative diseases with special reference to dementia. FEBS Lett. 2006, 580, 2994–3005. [Google Scholar] [CrossRef]
- Naviaux, R.K. Mitochondrion Metabolic features of the cell danger response. MITOCH 2014, 16, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Horhat, F.G.; Rogobete, A.F.; Papurica, M.; Sandesc, D.; Tanasescu, S.; Dumitrascu, V.; Licker, M.; Nitu, R.; Cradigati, C.A.; Sarandan, M.; et al. The Use of Lipid Peroxidation Expression as a Biomarker for the Molecular Damage in the Critically Ill Polytrauma Patient. Clin. Lab. 2016, 62, 1601–1607. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.C.; Valença, S.S.; Gitirana, L.B.; Santos, J.C.; Ribeiro, L.M.; Machado, M.N.; Magalhães, C.B.; Zin, W.A.; Porto, L.C. Redox markers and inflammation are differentially affected by atorvastatin, pravastatin or simvastatin administered before endotoxin-induced acute lung injury. Int. Immunopharmacol. 2013, 17, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Moise, A. Vitamin D in Critically Ill Patients - From Molecular Damage Interactions to Clinical Outcomes Benefits. When, Why, How? Central. European. J. Clin. Res. 2018, 1, 59–66. [Google Scholar] [CrossRef][Green Version]
- Sandesc, M.; Rogobete, A.F.; Bedreag, O.H.; Dinu, A.; Papurica, M.; Cradigati, C.A.; Sarandan, M.; Popovici, S.E.; Bratu, L.M.; Bratu, T.; et al. Analysis of oxidative stress-related markers in critically ill polytrauma patients: An observational prospective single-center study. Bosn. J. Basic Med. Sci. 2018, 18, 191–197. [Google Scholar] [CrossRef]
- Bedreag, O.H.; Rogobete, A.F.; Sandesc, D.; Cradigati, C.A.; Sarandan, M.; Popovici, S.E.; Dumache, R.; Horhat, F.G.; Vernic, C.; Sima, L.V.; et al. Modulation of the Redox Expression and Inflammation Response in the Critically Ill Polytrauma Patient with Thoracic Injury. Statistical Correlations between Antioxidant Therapy and Clinical Aspects. A Retrospective Single Center Study. Clin. Lab. 2016, 62, 1747–1759. [Google Scholar] [CrossRef]
- Hu, Y.; Deng, H.; Xu, S.; Zhang, J. MicroRNAs Regulate Mitochondrial Function in Cerebral Ischemia-Reperfusion Injury. Int. J. Mol. Sci. 2015, 16, 24895–24917. [Google Scholar] [CrossRef]
- Fredriksson, K.; Tjäder, I.; Keller, P.; Petrovic, N.; Ahlman, B.; Schéele, C.; Wernerman, J.; Timmons, J.A.; Rooyackers, O. Dysregulation of mitochondrial dynamics and the muscle transcriptome in ICU patients suffering from sepsis induced multiple organ failure. PLoS ONE 2008, 3, e3686. [Google Scholar] [CrossRef]
- Ross, J.A.; Tolar, J.; Spector, L.G.; DeFor, T.; Lund, T.C.; Weisdorf, D.J.; Langer, E.; Hooten, A.J.; Thyagarajan, B.; Gleason, M.K.; et al. An exploratory analysis of mitochondrial haplotypes and allogeneic hematopoietic cell transplantation outcomes. Biol. Blood Marrow Transplant. 2015, 21, 81–88. [Google Scholar] [CrossRef][Green Version]
- Yao, X.; Carlson, D.; Sun, Y.; Ma, L.; Wolf, S.E.; Minei, J.P.; Zang, Q.S. Mitochondrial ROS induces cardiac inflammation via a pathway through mtDNA damage in a pneumonia-related sepsis model. PLoS ONE 2015, 10, e0139416. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Z. The pathophysiological role of mitochondrial oxidative stress in lung diseases. J. Transl. Med. 2017, 15, 207. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Hu, F.; Wu, J.; Zhang, S. Redox Biology Cannabidiol attenuates OGD/R-induced damage by enhancing mitochondrial bioenergetics and modulating glucose metabolism via pentose-phosphate pathway in hippocampal neurons. Redox Biol. 2017, 11, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Burstein, S. Bioorganic & Medicinal Chemistry Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorg. Med. Chem. 2015, 23, 1377–1385. [Google Scholar] [PubMed]
- Carrier, E.J.; Auchampach, J.A.; Hillard, C.J. Inhibition of an equilibrative nucleoside transporter by cannabidiol: A mechanism of cannabinoid immunosuppression. Proc. Natl. Acad. Sci. USA 2006, 103, 7895–7900. [Google Scholar] [CrossRef] [PubMed]
- Castillo, R.L.; Loza, R.C. Pathophysiological Approaches of Acute Respiratory Distress syndrome: Novel Bases for Study of Lung Injury. Open Respir. Med. J. 2015, 9, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Hebert-Chatelain, E.; Desprez, T.; Serrat, R.; Bellocchio, L.; Soria-Gomez, E.; Busquets-Garcia, A.; Pagano Zottola, A.C.; Delamarre, A.; Cannich, A.; Vincent, P.; et al. A cannabinoid link between mitochondria and memory. Nature 2016, 539, 555. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.; Drysdale, A.J.; Lafourcade, C.; Pertwee, R.G.; Platt, B. Cannabidiol Targets Mitochondria to Regulate Intracellular Ca+2 Levels. J. Neurosci. 2009, 29, 2053–2063. [Google Scholar] [CrossRef]
- Resstel, L.B.M.; Tavares, R.F.; Lisboa, S.F.S.; Joca, S.R.L.; Corrêa, F.M.A.; Guimarães, F.S. 5-HT1A receptors are involved in the cannabidiol-induced attenuation of behavioural and cardiovascular responses to acute restraint stress in rats. Br. J. Pharmacol. 2009, 156, 181–188. [Google Scholar] [CrossRef]
- Nalivaiko, E.; Ootsuka, Y.; Blessing, W.W. Activation of 5-HT1A receptors in the medullary raphe reduces cardiovascular changes elicited by acute psychological and inflammatory stresses in rabbits. Am. J. Physiol. Integr. Comp. Physiol. 2005, 289, R596–R604. [Google Scholar] [CrossRef]
- Mccoy, K.L. Interaction between Cannabinoid System and Toll-Like Receptors Controls Inflammation. Mediat. Inflamm. 2016. [Google Scholar] [CrossRef]
- Kennett, G.A.; Dourish, C.T.; Curzon, G. Antidepressant-like action of 5-HT1A agonists and conventional antidepressants in an animal model of depression. Eur. J. Pharmacol. 1987, 134, 265–274. [Google Scholar] [CrossRef]
- Popova, N.K.; Amstislavskaya, T.G. Involvement of the 5-HT1A and 5-HT1B serotonergic receptor subtypes in sexual arousal in male mice. Psychoneuroendocrinology 2002, 27, 609–618. [Google Scholar] [CrossRef]
- Kathmann, M.; Flau, K.; Redmer, A.; Tränkle, C.; Schlicker, E. Cannabidiol is an allosteric modulator at mu- and delta-opioid receptors. Naunyn. Schmiedebergs. Arch. Pharmacol. 2006, 372, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Iuvone, T.; Esposito, G.; De Filippis, D.; Scuderi, C.; Steardo, L. Cannabidiol: A Promising Drug for Neurodegenerative Disorders? CNS Neurosci. Ther. 2009, 15, 65–75. [Google Scholar] [CrossRef]
- Jhaveri, M.D.; Sagar, D.R.; Elmes, S.J.R.; Kendall, D.A.; Chapman, V. Cannabinoid CB2 Receptor-Mediated Anti-nociception in Models of Acute and Chronic Pain. Mol. Neurobiol. 2007, 36, 26–35. [Google Scholar] [CrossRef]
- Rog, D.J.; Nurmikko, T.J.; Friede, T.; Young, C.A. Randomized, controlled trial of cannabis-based medicine in central pain in multiple sclerosis. Neurology 2005, 65, 812. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Lee, S.T.; Lin, W.-W. Effects of cannabinoids on LPS-stimulated inflammatory mediator release from macrophages: Involvement of eicosanoids. J. Cell. Biochem. 2001, 81, 715–723. [Google Scholar] [CrossRef]
- Baldwin, G.C.; Tashkin, D.P.; Buckley, D.M.; Park, A.N.; Dubinett, S.M.; Roth, M.D. Marijuana and cocaine impair alveolar macrophage function and cytokane production. Am. J. Respir. Crit. Care Med. 1997, 156, 1606–1613. [Google Scholar] [CrossRef]
- Sido, J.M.; Nagarkatti, P.S.; Nagarkatti, M. Δ9-Tetrahydrocannabinol attenuates allogeneic host-versus-graft response and delays skin graft rejection through activation of cannabinoid receptor 1 and induction of myeloid-derived suppressor cells. J. Leukoc. Biol. 2015, 98, 435–447. [Google Scholar] [CrossRef]
- Lee, W.; Erdelyi, K.; Matyas, C.; Mukhopadhyay, P.; Varga, Z.V. Cannabidiol Limits T Cell – Mediated Chronic Autoimmune Myocarditis: Implications to Autoimmune Disorders and Organ Transplantation. Mol. Med. 2016, 22, 136–146. [Google Scholar] [CrossRef]
- Pandey, R.; Hegde, V.L.; Nagarkatti, M.; Nagarkatti, P.S.; Carolina, S. Targeting Cannabinoid Receptors as a Novel Approach in the Treatment of Graft-versus-Host Disease: Evidence from an Experimental Murine Model. J. Pharmacol. Exp. Ther. 2011, 338, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Kozela, E.; Juknat, A.; Gao, F.; Kaushansky, N.; Coppola, G.; Vogel, Z. Pathways and gene networks mediating the regulatory effects of cannabidiol, a nonpsychoactive cannabinoid, in autoimmune T cells. J. Neuroinflammation 2016, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Rock, E.M.; Sullivan, M.T.; Pravato, S.; Pratt, M.; Limebeer, C.L.; Parker, L.A. Effect of combined doses of Δ(9)-tetrahydrocannabinol and cannabidiol or tetrahydrocannabinolic acid and cannabidiolic acid on acute nausea in male Sprague-Dawley rats. Psychopharmacology (Berl). 2020. [Google Scholar] [CrossRef] [PubMed]
- Id, A.J.; Gao, F.; Coppola, G.; Vogel, Z.; Kozela, E. miRNA expression profiles and molecular networks in resting and LPS-activated BV-2 microglia—Effect of cannabinoids. PLoS ONE 2019, 14, e0212039. [Google Scholar]
- Dumache, R.; Ciocan, V.; Muresan, C.; Rogobete, A.F.; Enache, A. Circulating microRNAs as promising biomarkers in forensic body fluids identification. Clin. Lab. 2015, 61, 1129–1135. [Google Scholar] [CrossRef]
- Dumache, R.; Rogobete, A.F.; Bedreag, O.H.; Sarandan, M.; Cradigati, A.C.; Papurica, M.; Dumbuleu, C.M.; Nartita, R.; Sandesc, D. Use of miRNAs as Biomarkers in Sepsis. Anal. Cell. Pathol. 2015, 2015, 186716. [Google Scholar] [CrossRef]
- Papurica, M.; Rogobete, A.F.; Sandesc, D.; Dumache, R.; Cradigati, C.A.; Sarandan, M.; Nartita, R.; Popovici, S.E.; Bedreag, O.H. Advances in Biomarkers in Critical Ill Polytrauma Patients. Clin. Lab. 2016, 62, 977–986. [Google Scholar] [CrossRef]
- Negoita, S.I.; Sandesc, D.; Rogobete, A.F.; Dutu, M.; Bedreag, O.H.; Papurica, M.; Ercisli, M.F.; Popovici, S.E.; Dumache, R.; Sandesc, M.; et al. MiRNAs expressions and interaction with biological systems in patients with Alzheimer’s disease. Using miRNAs as a diagnosis and prognosis biomarker. Clin. Lab. 2017, 63, 1315–1321. [Google Scholar] [CrossRef]
- Ivan, M.V.; Rogobete, A.F.; Bedreag, O.H.; Papurica, M.; Popovici, S.E.; Dinu, A.; Sandesc, M.; Beceanu, A.; Bratu, L.M.; Popoiu, C.M.; et al. New Molecular and Epigenetic Expressions as Novel Biomarkers in Critically Ill Polytrauma Patients with Acute Kidney Injury (AKI). Clin. Lab. 2018, 64, 663–668. [Google Scholar] [CrossRef]
- Koukos, G.; Polytarchou, C.; Kaplan, J.L.; Morley-Fletcher, A.; Gras-Miralles, B.; Kokkotou, E.; Baril-Dore, M.; Pothoulakis, C.; Winter, H.S.; Iliopoulos, D. MicroRNA-124 regulates STAT3 expression and is down-regulated in colon tissues of pediatric patients with ulcerative colitis. Gastroenterology 2013, 145, 842–852. [Google Scholar] [CrossRef]
- Ma, C.; Li, Y.; Li, M.; Deng, G.; Wu, X.; Zeng, J.; Hao, X.; Wang, X.; Liu, J.; Cho, W.C.S.; et al. microRNA-124 negatively regulates TLR signaling in alveolar macrophages in response to mycobacterial infection. Mol. Immunol. 2014, 62, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, N.; Peng, Y.; Tan, Z.; Ciraolo, G.; Wang, D.; Li, R. miRNAs in mtDNA-less cell mitochondria. Cell Death Discov. 2015, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, J.; Chi, H.; Zhang, F.; Zhu, X. MicroRNA-181c targets Bcl-2 and regulates mitochondrial morphology in myocardial cells. 2015, 19, 2084–2097. J. Cell. Mol. Med. 2015, 19, 20. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Icli, B.; Wara, A.K.; Belkin, N.; He, S.; Kobzik, L.; Hunninghake, G.M.; Vera, M.P.; Registry, M.; Blackwell, T.S.; et al. MicroRNA-181b regulates NF- κ B – mediated vascular inflammation. J. Clin. Investig. 2012, 122, 1973–1990. [Google Scholar] [CrossRef]
- Mu, N.; Gu, J.; Huang, T.; Zhang, C.; Shu, Z.; Li, M.; Hao, Q.; Li, W.; Zhang, W.; Zhao, J.; et al. A novel NF-κB/YY1/microRNA-10a regulatory circuit in fibroblast-like synoviocytes regulates inflammation in rheumatoid arthritis. Sci. Rep. 2016, 6, 20059. [Google Scholar] [CrossRef]
- Jeker, L.T.; Zhou, X.; Gershberg, K.; De Kouchkovsky, D.; Morar, M.M.; Stadthagen, G.; Lund, A.H.; Bluestone, J.A.
- Sheedy, F.J.; Palsson-McDermott, E.; Hennessy, E.J.; Martin, C.; O’Leary, J.J.; Ruan, Q.; Johnson, D.S.; Chen, Y.; O’Neill, L.A.J. Negative regulation of TLR4 via targeting of the proinflammatory tumor suppressor PDCD4 by the microRNA miR-21. Nat. Immunol. 2010, 11, 141–147. [Google Scholar] [CrossRef]
- Liu, S.S.; Wang, Y.U.S.H.I.; Sun, Y.A.N.F.; Miao, L.I.X.I.A. Plasma microRNA - 320, microRNA - let - 7e and microRNA - 21 as novel potential biomarkers for the detection of retinoblastoma. Biomedical Reports 2014, 424–428. [Google Scholar] [CrossRef]
- Lin, Q.; Geng, Y.; Zhao, M.; Lin, S.; Zhu, Q.; Tian, Z. MiR-21 Regulates TNF-alpha-Induced CD40 Expression via the SIRT1-NF-kappaB Pathway in Renal Inner Medullary Collecting Duct Cells. Cell. Physiol. Biochem. 2017, 41, 124–136. [Google Scholar] [CrossRef]
- Canfran-Duque, A.; Rotllan, N.; Zhang, X.; Fernandez-Fuertes, M.; Ramirez-Hidalgo, C.; Araldi, E.; Daimiel, L.; Busto, R.; Fernandez-Hernando, C.; Suarez, Y. Macrophage deficiency of miR-21 promotes apoptosis, plaque necrosis, and vascular inflammation during atherogenesis. EMBO Mol. Med. 2017, 9, 1244–1262. [Google Scholar] [CrossRef]
- Caescu, C.I.; Guo, X.; Tesfa, L.; Bhagat, T.D.; Verma, A.; Zheng, D.; Stanley, E.R. Colony stimulating factor-1 receptor signaling networks inhibit mouse macrophage inflammatory responses by induction of microRNA-21. Blood 2015, 125, e1–e13. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Ganesh, K.; Khanna, S.; Sen, C.K.; Roy, S. Engulfment of apoptotic cells by macrophages: A role of microRNA-21 in the resolution of wound inflammation. J. Immunol. 2014, 192, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Cd, H.S.-; Cells, S.-C.D.; Wu, Q.; Zhan, J.; Li, Y.; Wang, X.; Xu, L.; Yu, J.; Pu, S.; Zhou, Z. Differentiation-Associated MicroRNA Alterations in Mouse. Stem cells International 2016, 2016. [Google Scholar]
- Suarez, Y.; Wang, C.; Manes, T.D.; Pober, J.S. Cutting edge: TNF-induced microRNAs regulate TNF-induced expression of E-selectin and intercellular adhesion molecule-1 on human endothelial cells: Feedback control of inflammation. J. Immunol. 2010, 184, 21–25. [Google Scholar] [CrossRef]
- Ganan-Gomez, I.; Wei, Y.; Yang, H.; Pierce, S.; Bueso-Ramos, C.; Calin, G.; Boyano-Adanez, M.D.C.; Garcia-Manero, G. Overexpression of miR-125a in myelodysplastic syndrome CD34+ cells modulates NF-kappaB activation and enhances erythroid differentiation arrest. PLoS ONE 2014, 9, e93404. [Google Scholar] [CrossRef]
- Ho, J.; Chan, H.; Wong, S.H.; Wang, M.H.T.; Yu, J.; Xiao, Z.; Liu, X.; Choi, G.; Leung, C.C.H.; Wong, W.T.; et al. The involvement of regulatory non-coding RNAs in sepsis: A systematic review. Crit. Care 2016, 20, 1–12. [Google Scholar] [CrossRef]
- Gaston, T.E.; Friedman, D. Pharmacology of cannabinoids in the treatment of epilepsy. Epilepsy. Behav. 2017, 70, 313–318. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Bowie, A.G. The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat. Rev. Immunol. 2007, 7, 353–364. [Google Scholar] [CrossRef]
- Chen, X.-M.; Splinter, P.L.; O’Hara, S.P.; LaRusso, N.F. A cellular micro-RNA, let-7i, regulates Toll-like receptor 4 expression and contributes to cholangiocyte immune responses against Cryptosporidium parvum infection. J. Biol. Chem. 2007, 282, 28929–28938. [Google Scholar] [CrossRef]
- Ma, C.; Li, Y.; Zeng, J.; Wu, X.; Liu, X.; Wang, Y. Mycobacterium bovis BCG Triggered MyD88 Induces miR-124 Feedback Negatively Regulates Immune Response in Alveolar Epithelial Cells. PLoS ONE 2014, 9, e92419. [Google Scholar] [CrossRef]
- Liang, H.; Yan, X.; Pan, Y.; Wang, Y.; Wang, N.; Li, L.; Liu, Y.; Chen, X. MicroRNA-223 delivered by platelet-derived microvesicles promotes lung cancer cell invasion via targeting tumor suppressor EPB41L3. Mol. Cancer 2015, 14, 58. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Qiao, Y.; Wang, P.; Li, S.; Zhao, W.; Gao, C. microRNA-210 negatively regulates LPS-induced production of proinflammatory cytokines by targeting NF-kappaB1 in murine macrophages. FEBS Lett. 2012, 586, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.-T.; Chen, R.; Chen, C.; Su, K.; Li, W.; Tang, L.-H.; Liu, H.-M.; Xue, R.; Sun, Q.; Leng, Y.; et al. Transcription factors Nrf2 and NF-κB contribute to inflammation and apoptosis induced by intestinal ischemia-reperfusion in mice. Int. J. Mol. Med. 2017, 40, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Liu, Z.; Li, B.-S.; Tang, B.; Li, W.; Guo, G.; Shi, Y.; Wang, F.; Wu, Y.; Tong, W.-D.; et al. Induction of microRNA-155 during Helicobacter pylori infection and its negative regulatory role in the inflammatory response. J. Infect. Dis. 2009, 200, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Gu, J.T.; Yi, B.I.N.; Tang, Z.Z.H.I.; Tao, G.U. microRNA-23b regulates the expression of inflammatory factors in vascular endothelial cells during sepsis. Exp. Ther. Med. 2015, 9, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Dalli, J.; Chiang, N.; Baron, R.M.; Quintana, C.; Serhan, C.N. Article Plasticity of Leukocytic Exudates in Resolving Acute Inflammation Is Regulated by MicroRNA and Proresolving Mediators. Immunity 2013, 39, 885–898. [Google Scholar] [CrossRef]
- Ma, X.; Buscaglia, L.E.B.; Barker, J.R.; Li, Y. MicroRNAs in NF-kB signaling. J. Mol. Cell Biol. 2011, 3, 159–166. [Google Scholar] [CrossRef]
- Yang, X.; Bam, M.; Nagarkatti, P.S.; Nagarkatti, M. RNA-seq Analysis of δ9-Tetrahydrocannabinol-treated T Cells Reveals Altered Gene Expression Profiles That Regulate Immune Response and Cell Proliferation. J. Biol. Chem. 2016, 291, 15460–15472. [Google Scholar] [CrossRef]
- Rao, R.; Nagarkatti, P.S.; Nagarkatti, M. Δ9 Tetrahydrocannabinol attenuates Staphylococcal enterotoxin B-induced inflammatory lung injury and prevents mortality in mice by modulation of miR-17-92 cluster and induction of T-regulatory cells Tables of Links. Br. J. Pharmacol. 2015, 172, 1792–1806. [Google Scholar] [CrossRef]
- Ingram, G.; Pearson, O.R. Cannabis and multiple sclerosis. Pract Neurol. 2019, 19, 310–315. [Google Scholar] [CrossRef]
- Sripada, L.; Tomar, D.; Singh, R. Mitochondrion Mitochondria: One of the destinations of miRNAs. MITOCH 2012, 12, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Kren, B.T.; Wong, P.Y.; Sarver, A.; Zhang, X.; Zeng, Y.; Steer, C.J. MicroRNAs identified in highly purified liver-derived mitochondria may play a role in apoptosis. RNA Biol. 2009, 6, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Sripada, L.; Tomar, D.; Prajapati, P.; Singh, R.; Singh, A.K.; Singh, R. Systematic Analysis of Small RNAs Associated with Human Mitochondria by Deep Sequencing: Detailed Analysis of Mitochondrial Associated miRNA. PLoS ONE 2012, 7, e44873. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Peng, X.; Huang, F.; Yao, D.; Yu, Q.; Liang, H. The Spectroscopy Study of the Binding of an Active Ingredient of Dioscorea Species with Bovine Serum Albumin with or without Co 2+. Evid. Based Complementary Altern. Med. 2014. [Google Scholar] [CrossRef]
- Zhou, W.; Su, L.; Duan, X.; Chen, X.; Hays, A.; Upadhyayula, S.; Shivde, J.; Wang, H.; Li, Y.; Huang, D.; et al. MicroRNA-21 down-regulates inflammation and inhibits periodontitis. Mol. Immunol. 2018, 101, 608–614. [Google Scholar] [CrossRef]
- Sido, J.M.; Jackson, A.R.; Nagarkatti, P.S.; Nagarkatti, M. Marijuana-derived Δ-9-tetrahydrocannabinol suppresses Th1/Th17 cell-mediated delayed-type hypersensitivity through microRNA regulation. J. Mol. Med. 2016, 94, 1039–1051. [Google Scholar] [CrossRef]
- Al-Ghezi, Z.Z.; Miranda, K.; Nagarkatti, M.; Nagarkatti, P.S. Combination of Cannabinoids, Δ9- Tetrahydrocannabinol and Cannabidiol, Ameliorates Experimental Multiple Sclerosis by Suppressing Neuroinflammation Through Regulation of miRNA-Mediated Signaling Pathways. Front. Immunol. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Lehmann, C.; Kianian, M.; Zhou, J.; Küster, I.; Kuschnereit, R.; Whynot, S.; Hung, O.; Shukla, R.; Johnston, B.; Cerny, V.; et al. Cannabinoid receptor 2 activation reduces intestinal leukocyte recruitment and systemic inflammatory mediator release in acute experimental sepsis. Crit. Care 2012, 16. [Google Scholar] [CrossRef]
- Çakır, M.; Tekin, S.; Okan, A.; Çakan, P.; Doğanyiğit, Z. The ameliorating effect of cannabinoid type 2 receptor activation on brain, lung, liver and heart damage in cecal ligation and puncture-induced sepsis model in rats. Int. Immunopharmacol. 2019, 78. [Google Scholar] [CrossRef]
- Newton, C.A.; Klein, T.W.; Friedman, H. Secondary immunity to Legionella pneumophila and Th1 activity are suppressed by delta-9-tetrahydrocannabinol injection. Infect. Immun. 1994, 62, 4015–4020. [Google Scholar] [CrossRef]
- Gardner, B.; Zu, L.X.; Sharma, S.; Liu, Q.; Makriyannis, A.; Tashkin, D.P.; Dubinett, S.M. Autocrine and Paracrine Regulation of Lymphocyte CB2 Receptor Expression by TGF-β. Biochem. Biophys. Res. Commun. 2002, 290, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Croxford, J.L.; Yamamura, T. Cannabinoids and the immune system: Potential for the treatment of inflammatory diseases? J. Neuroimmunol. 2005, 166, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Mageed, R.A.; Adams, G.; Woodrow, D.; Podhajcer, O.L.; Chernajovsky, Y. Prevention of collagen-induced arthritis by gene delivery of soluble p75 tumour necrosis factor receptor. Gene Ther. 1998, 5, 1584–1592. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Triantaphyllopoulos, K.A.; Williams, R.O.; Tailor, H.; Chernajovsky, Y. Amelioration of collagen-induced arthritis and suppression of interferon-γ, interleukin-12, and tumor necrosis factor α production by interferon-β gene therapy. Arthritis Rheum. 1999, 42, 90–99. [Google Scholar] [CrossRef]
- David, V.L.; Izvernariu, D.A.; Popoiu, C.M.; Puiu, M.; Boia, E.S. Morphologic, morphometrical and histochemical proprieties of the costal cartilage in children with pectus excavatum. Rom. J. Morphol. Embryol. 2011, 52, 625–629. [Google Scholar]
- Ross, R.A.; Brockie, H.C.; Pertwee, R.G. Inhibition of nitric oxide production in RAW264.7 macrophages by cannabinoids and palmitoylethanolamide. Eur. J. Pharmacol. 2000, 401, 121–130. [Google Scholar] [CrossRef]
- Watzl, B.; Scuderi, P.; Watson, R.R. marijuana components stimulate human peripheral bloodmonon uclear cell secretion of interferon - gammaandsuppress interleukin-1 alpha in vitro. Int. J. Immunopharmacol. 1991, 13, 1091–1097. [Google Scholar] [CrossRef]
- Srivastava, M.D.; Srivastava, B.I.S.; Brouhard, B. Δ9 Tetrahydrocannabinol and cannabidiol alter cytokine production by human immune cells. Immunopharmacology 1998, 40, 179–185. [Google Scholar] [CrossRef]
- Yuan, M.; Kiertscher, S.M.; Cheng, Q.; Zoumalan, R.; Tashkin, D.P.; Roth, M.D. Δ9-Tetrahydrocannabinol regulates Th1/Th2 cytokine balance in activated human T cells. J. Neuroimmunol. 2002, 133, 124–131. [Google Scholar] [CrossRef]
- Aziz, M.; Jacob, A.; Yang, W.-L.; Matsuda, A.; Wang, P. Current trends in inflammatory and immunomodulatory mediators in sepsis. J. Leukoc. Biol. 2013, 93, 329–342. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Disorder | Observations | Reference |
|---|---|---|
| Pain | CB1 receptor agonists have a nociceptive action on the interneurons in the spinal cord, CB2 acts directly on reducing inflammation, and the CB2 receptor was shown to have an increased immunomodulatory response. | [63] |
| Cancer | The following effects have been reported: Anti-inflammatory, anti-proliferative, pro-apoptotic, anti-invasive, and anti-metastatic. | [50,51,64] |
| Hepatic metabolism | Directly acts on the modulation of the hepatic metabolism through gluconeogenesis and lipogenesis, and the CB2 receptor has a protective action on the phenomenon induced by ischemia reperfusion injury. | [47,49,65,66,67] |
| Gastrointestinal system | CB1 and CB2 receptors inhibit the pro-inflammatory and pro-oxidative activities specifically for the colon. | [49,65,68] |
| Cardiovascular system | CB2 receptor reduces inflammation specifically related to atheromatous plaques and reduces thrombosis risk; CB1 activates AMP-activated protein kinase (AMPK), reduces insulin resistance, and mimics all of the effects that encompass ischemia-reperfusion injury (IR). | [40,68,69,70] |
| Immune system/inflammation response | Reduces iNOS activity, reduces IL-6 expression; reduces TNF-α and IL-1β expression; reduces specific inflammation of ARDS/ALI; modulates and reduces the activity of TNF-α and COX-2 in the context of LPS-induced inflammation; inhibits neutrophil chemotaxis; and modulates the expression of IFN-γ, leading to the decrease of IL-2 expression. | [53,71,72,73,74,75] |
| Cannabinoid | Observations | References |
|---|---|---|
| CBD in vitro and in vivo studies |
| [108,109,110,111,112,113,114,115,116,117,118,119,120,121,122] |
| THC in vivo and in vitro studies |
| [105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dinu, A.R.; Rogobete, A.F.; Bratu, T.; Popovici, S.E.; Bedreag, O.H.; Papurica, M.; Bratu, L.M.; Sandesc, D. Cannabis Sativa Revisited—Crosstalk between microRNA Expression, Inflammation, Oxidative Stress, and Endocannabinoid Response System in Critically Ill Patients with Sepsis. Cells 2020, 9, 307. https://doi.org/10.3390/cells9020307
Dinu AR, Rogobete AF, Bratu T, Popovici SE, Bedreag OH, Papurica M, Bratu LM, Sandesc D. Cannabis Sativa Revisited—Crosstalk between microRNA Expression, Inflammation, Oxidative Stress, and Endocannabinoid Response System in Critically Ill Patients with Sepsis. Cells. 2020; 9(2):307. https://doi.org/10.3390/cells9020307
Chicago/Turabian StyleDinu, Anca Raluca, Alexandru Florin Rogobete, Tiberiu Bratu, Sonia Elena Popovici, Ovidiu Horea Bedreag, Marius Papurica, Lavinia Melania Bratu, and Dorel Sandesc. 2020. "Cannabis Sativa Revisited—Crosstalk between microRNA Expression, Inflammation, Oxidative Stress, and Endocannabinoid Response System in Critically Ill Patients with Sepsis" Cells 9, no. 2: 307. https://doi.org/10.3390/cells9020307
APA StyleDinu, A. R., Rogobete, A. F., Bratu, T., Popovici, S. E., Bedreag, O. H., Papurica, M., Bratu, L. M., & Sandesc, D. (2020). Cannabis Sativa Revisited—Crosstalk between microRNA Expression, Inflammation, Oxidative Stress, and Endocannabinoid Response System in Critically Ill Patients with Sepsis. Cells, 9(2), 307. https://doi.org/10.3390/cells9020307