Identification of CNS Injury-Related microRNAs as Novel Toll-Like Receptor 7/8 Signaling Activators by Small RNA Sequencing

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice and Ethics Statement
2.2. Synthetic microRNA Mimics
2.3. Primary Cortical Neurons
2.4. Primary Microglia
2.5. Co-Cultures of Neurons and Microglia
2.6. Immunofluorescence Microscopy
2.7. Induction of Apoptosis in Neurons
2.8. Small RNA Enrichment
2.9. Library Generation and Small RNA Sequencing
2.10. Data Processing
2.11. HEK-Blue TLR Reporter Cell Lines
2.12. TNF Enzyme Linked Immunosorbent Assay
2.13. Multiplex Immune Assay
2.14. Gene Ontology and KEGG Analyses
2.15. Statistical Analyses
3. Results
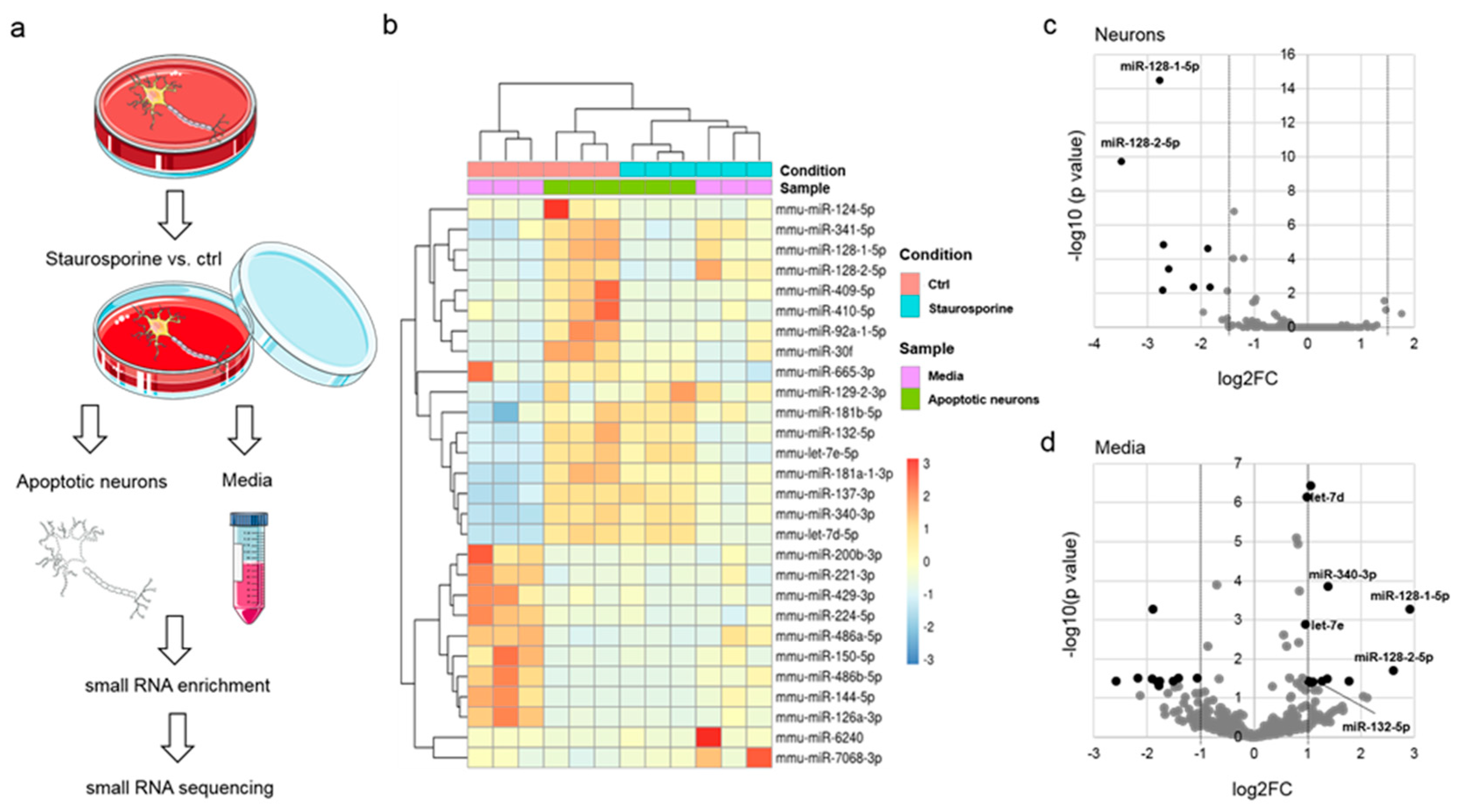
3.1. Identification of miRNAs Released from Apoptotic Neurons
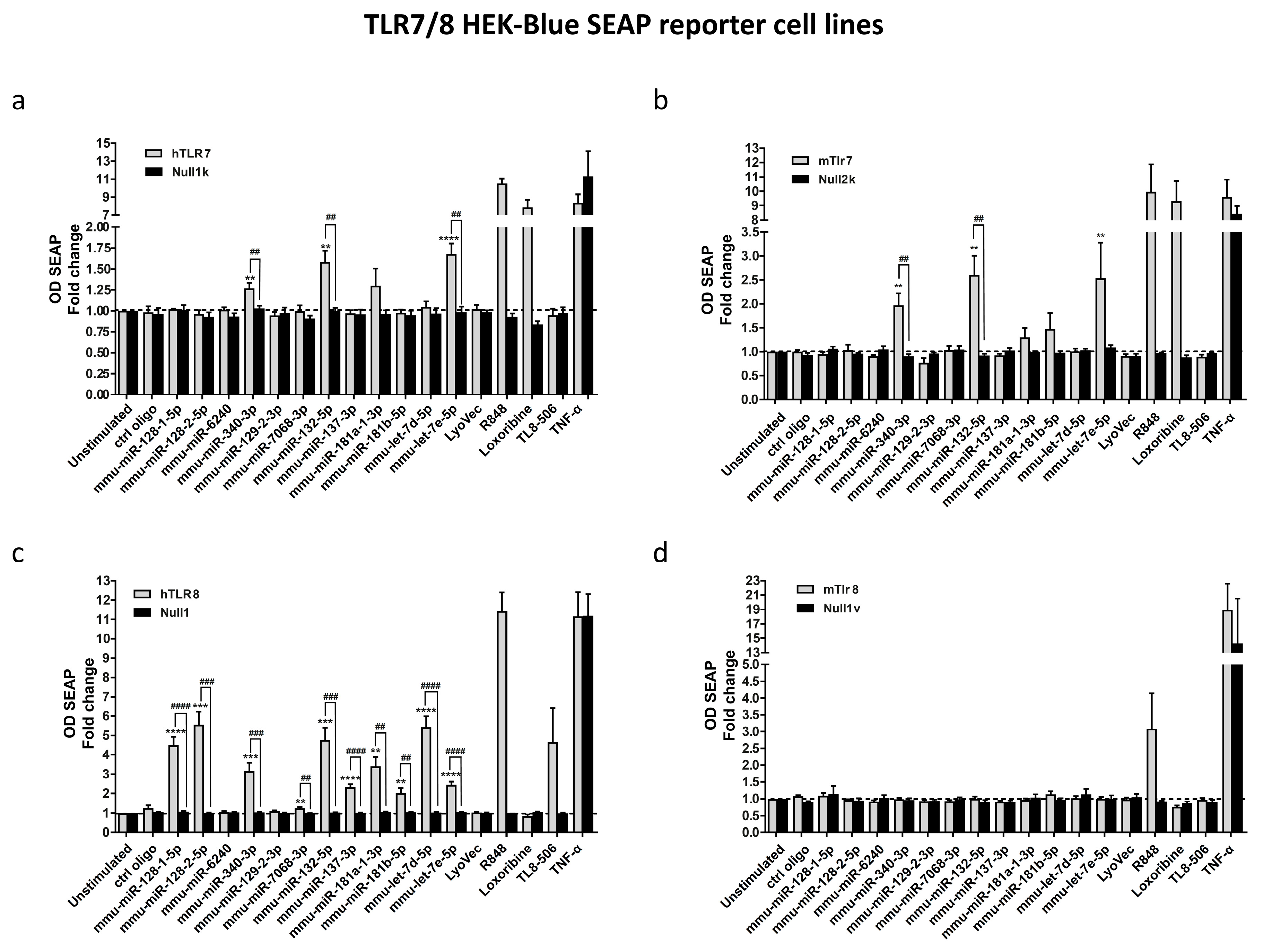
3.2. Select miRNAs Released from Injured Neurons Activate TLR7/8 Expressed in HEK293-Derived TLR Reporter Cells
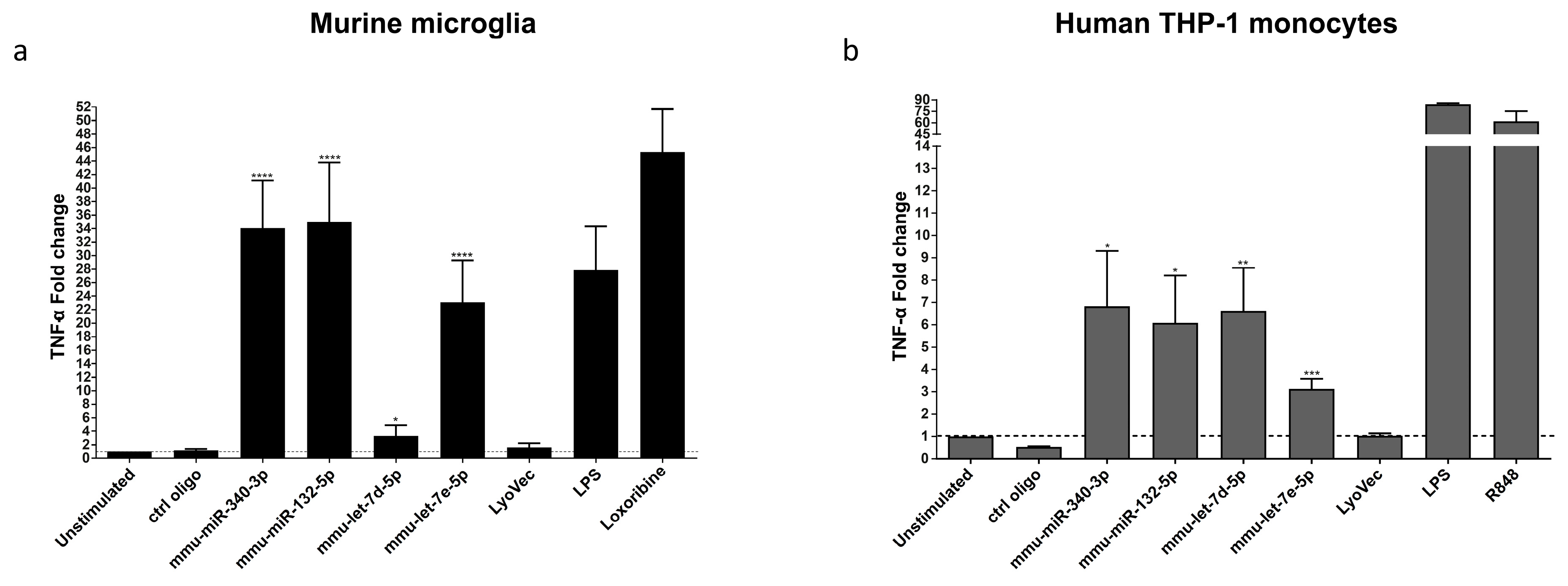
3.3. miR-340-3p and miR-132-5p Induce Cytokine and Chemokine Release from Murine Microglia and Human-Derived Monocytes
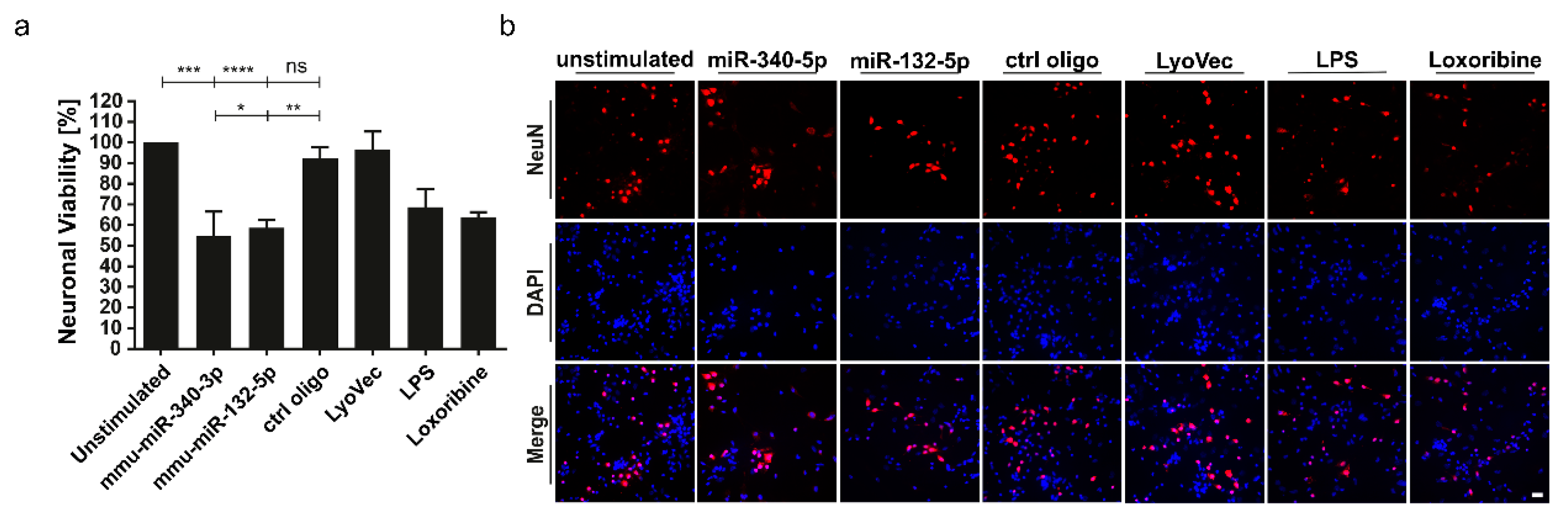
3.4. miR-340-3p and miR-132-5p Trigger Neuronal Injury
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heneka, M.T.; Kummer, M.P.; Latz, E. Innate immune activation in neurodegenerative disease. Nat. Rev. Immunol. 2014, 14, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Kielian, T. Overview of Toll-Like Receptors in the CNS, in Toll-like Receptors: Roles in Infection and Neuropathology; Kielian, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–14. [Google Scholar]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Forsbach, A.; Nemorin, J.G.; Montino, C.; Müller, C.; Samulowitz, U.; Vicari, A.P.; Jurk, M.; Mutwiri, G.K.; Krieg, A.M.; Lipford, G.B.; et al. Identification of RNA sequence motifs stimulating sequence-specific TLR8-dependent immune responses. J. Immunol. 2008, 180, 3729–3738. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [PubMed]
- Lund, J.M.; Alexopoulou, L.; Sato, A.; Karow, M.; Adams, N.C.; Gale, N.W.; Iwasaki, A.; Flavell, R.A. Recognition of single-stranded RNA viruses by Toll-like receptor 7. Proc. Natl. Acad. Sci. USA 2004, 101, 5598–5603. [Google Scholar] [CrossRef] [PubMed]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; E Sousa, C.R. Innate Antiviral Responses by Means of TLR7-Mediated Recognition of Single-Stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.M.; Krüger, C.; Park, B.; Derkow, K.; Rosenberger, K.; Baumgart, J.; Trimbuch, T.; Eom, G.; Hinz, M.; Kaul, D.; et al. An unconventional role for miRNA: Let-7 activates Toll-like receptor 7 and causes neurodegeneration. Nat. Neurosci. 2012, 15, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, A.D.; Fonken, L.K.; Watkins, L.R.; Nelson, R.J.; Popovich, P.G. MicroRNAs: Roles in Regulating Neuroinflammation. Neurosci. 2017, 24, 221–245. [Google Scholar] [CrossRef]
- Derkow, K.; Rössling, R.; Schipke, C.; Krüger, C.; Bauer, J.; Fähling, M.; Stroux, A.; Schott, E.; Ruprecht, K.; Peters, O.; et al. Distinct expression of the neurotoxic microRNA family let-7 in the cerebrospinal fluid of patients with Alzheimer’s disease. PLoS ONE 2018, 13, e0200602. [Google Scholar] [CrossRef]
- Cogswell, J.P.; Ward, J.; Taylor, I.A.; Waters, M.; Shi, Y.; Cannon, B.; Kelnar, K.; Kemppainen, J.; Brown, D.; Chen, C.; et al. Identification of miRNA changes in Alzheimer’s disease brain and CSF yields putative biomarkers and insights into disease pathways. J. Alzheimers. Dis. 2008, 14, 27–41. [Google Scholar] [CrossRef]
- Glynn, C.L.; Khan, S.; Kerin, M.J.; Dwyer, R.M. Isolation of Secreted microRNAs (miRNAs) from Cell-conditioned Media. MicroRNA 2013, 2, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef] [PubMed]
- Koh, J.-Y.; Wie, M.B.; Gwag, B.J.; Sensi, S.L.; Canzoniero, L.M.; Demaro, J.; Csernansky, C.; Choi, D.W. Staurosporine-Induced Neuronal Apoptosis. Exp. Neurol. 1995, 135, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Krohn, A.J.; Preis, E.; Prehn, J.H.M. Staurosporine-Induced Apoptosis of Cultured Rat Hippocampal Neurons Involves Caspase-1-Like Proteases as Upstream Initiators and Increased Production of Superoxide as a Main Downstream Effector. J. Neurosci. 1998, 18, 8186–8197. [Google Scholar] [CrossRef] [PubMed]
- López, E.; Ferrer, I. Staurosporine- and H-7-induced cell death in SH-SY5Y neuroblastoma cells is associated with caspase-2 and caspase-3 activation, but not with activation of the FAS/FAS-L-caspase-8 signaling pathway. Mol. Brain Res. 2000, 85, 61–67. [Google Scholar] [CrossRef]
- Deshmukh, M.; Johnson, E.M. Staurosporine-induced neuronal death: Multiple mechanisms and methodological implications. Cell Death Differ. 2000, 7, 250–261. [Google Scholar] [CrossRef]
- Hutchison, E.R.; Kawamoto, E.M.; Taub, D.D.; Lal, A.; Abdelmohsen, K.; Zhang, Y.; Wood, W.H.; Lehrmann, E.; Camandola, S.; Becker, K.G.; et al. Evidence for miR-181 involvement in neuroinflammatory responses of astrocytes. Glia 2013, 61, 1018–1028. [Google Scholar] [CrossRef]
- Cao, W.; Manicassamy, S.; Tang, H.; Kasturi, S.P.; Pirani, A.; Murthy, N.; Pulendran, B. Toll-like receptor–mediated induction of type I interferon in plasmacytoid dendritic cells requires the rapamycin-sensitive PI (3) K-mTOR-p70S6K pathway. Nat. Immunol. 2008, 9, 1157–1164. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Jurk, M.; Heil, F.; Vollmer, J.; Schetter, C.; Krieg, A.M.; Wagner, H.; Lipford, G.; Bauer, S. Human TLR7 or TLR8 independently confer responsiveness to the antiviral compound R-848. Nat. Immunol. 2002, 3, 499. [Google Scholar] [CrossRef]
- Heil, F.; Ahmad-Nejad, P.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Gellert, T.; Dietrich, H.; Lipford, G.; Takeda, K.; Akira, S.; et al. The Toll-like receptor 7 (TLR7)-specific stimulus loxoribine uncovers a strong relationship within the TLR7, 8 and 9 subfamily. Eur. J. Immunol. 2003, 33, 2987–2997. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Dietsch, G.N.; Matthews, M.A.; Yang, Y.; Ghanekar, S.; Inokuma, M.; Suni, M.; Maino, V.C.; Henderson, K.E.; Howbert, J.J.; et al. VTX-2337 is a novel TLR8 agonist that activates NK cells and augments ADCC. Clin. Cancer Res. 2012, 18, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, C.; Hsu, L.C.; Luo, Y.; Xiang, R.; Chuang, T.H. A five-amino-acid motif in the undefined region of the TLR8 ectodomain is required for species-specific ligand recognition. Mol. Immunol. 2010, 47, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.K.; Miller, S.D. Microglia Initiate Central Nervous System Innate and Adaptive Immune Responses through Multiple TLRs. J. Immunol. 2004, 173, 3916–3924. [Google Scholar] [CrossRef]
- Zarember, K.A.; Godowski, P.J. Tissue Expression of Human Toll-Like Receptors and Differential Regulation of Toll-Like Receptor mRNAs in Leukocytes in Response to Microbes, Their Products, and Cytokines. J. Immunol. 2002, 168, 554–561. [Google Scholar] [CrossRef]
- Wang, J.; Shao, Y.; Bennett, T.A.; Shankar, R.A.; Wightman, P.D.; Reddy, L.G. The functional effects of physical interactions among Toll-like receptors 7, 8, and 9. J. Biol. Chem. 2006, 281, 37427–37434. [Google Scholar] [CrossRef]
- Lehmann, S.M.; Rosenberger, K.; Krüger, C.; Habbel, P.; Derkow, K.; Kaul, D.; Rybak, A.; Brandt, C.; Schott, E.; Wulczyn, F.G.; et al. Extracellularly Delivered Single-Stranded Viral RNA Causes Neurodegeneration Dependent on TLR7. J. Immunol. 2012, 189, 1448–1458. [Google Scholar] [CrossRef]
- Lehnardt, S.; Massillon, L.; Follett, P.; Jensen, F.E.; Ratan, R.; Rosenberg, P.A.; Volpe, J.J.; Vartanian, T. Activation of innate immunity in the CNS triggers neurodegeneration through a Toll-like receptor 4-dependent pathway. Proc. Natl. Acad. Sci. USA 2003, 100, 8514–8519. [Google Scholar] [CrossRef]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef]
- Feng, Y.; Zou, L.; Yan, D.; Chen, H.; Xu, G.; Jian, W.; Cui, P.; Chao, W. Extracellular MicroRNAs Induce Potent Innate Immune Responses via TLR7/MyD88-Dependent Mechanisms. J. Immunol. 2017, 199, 2106–2117. [Google Scholar] [CrossRef]
- Adlakha, Y.K.; Saini, N. Brain microRNAs and insights into biological functions and therapeutic potential of brain enriched miRNA-128. Mol. Cancer 2014, 13, 33. [Google Scholar] [CrossRef] [PubMed]
- Pigati, L.; Yaddanapudi, S.C.S.; Iyengar, R.; Kim, D.-J.; Hearn, S.A.; Danforth, D.; Hastings, M.L.; Duelli , D.M. Selective Release of MicroRNA Species from Normal and Malignant Mammary Epithelial Cells. PLoS ONE 2010, 5, e13515. [Google Scholar] [CrossRef] [PubMed]
- Lukiw, W.J. Micro-RNA speciation in fetal, adult and Alzheimer’s disease hippocampus. NeuroReport 2007, 18, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-l.; Min, L.; Guo, Q.D.; Zhang, J.X.; Jiang, H.L.; Shao, S.; Xing, J.G.; Yin, L.L.; Liu, J.H.; Liu, R.; et al. Profiling microRNA from Brain by Microarray in a Transgenic Mouse Model of Alzheimer’s Disease. BioMed Res. Int. 2017, 2017, 8030369. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Zhang, W.; Tan, E.K.; Zeng, L. Deciphering the Function and Regulation of microRNAs in Alzheimer’s Disease and Parkinson’s Disease. ACS Chem. Neurosci. 2014, 5, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Harischandra, D.S.; Ghaisas, S.; Rokad, D.; Zamanian, M.; Jin, H.; Anantharam, V.; Kimber, M.; Kanthasamy, A.; Kanthasamy, A.G. Environmental neurotoxicant manganese regulates exosome-mediated extracellular miRNAs in cell culture model of Parkinson’s disease: Relevance to α-synuclein misfolding in metal neurotoxicity. NeuroToxicology 2018, 64, 267–277. [Google Scholar] [CrossRef]
- Qian, Y.; Song, J.; Ouyang, Y.; Han, Q.; Chen, W.; Zhao, X.; Xie, Y.; Chen, Y.; Yuan, W.; Fan, C. Advances in Roles of miR-132 in the Nervous System. Front. Pharmacol. 2017, 8, 770. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Li, R.; Rezk, A.; Misirliyan, H.; Moore, C.; Farooqi, N.; Solis, M.; Goiry, L.G.; Junior, O.D.F.; Dang, V.D.; et al. A Novel MicroRNA-132-Surtuin-1 Axis Underlies Aberrant B-cell Cytokine Regulation in Patients with Relapsing-Remitting Multiple Sclerosis. PLoS ONE 2014, 9, e105421. [Google Scholar] [CrossRef]
- Huang, D.; Qiu, S.; Ge, R.; He, L.; Li, M.; Li, Y.; Peng, Y. miR-340 suppresses glioblastoma multiforme. Oncotarget 2015, 6, 9257–9270. [Google Scholar] [CrossRef]
- Fernandez, S.; Risolino, M.; Mandia, N.; Talotta, F.; Soini, Y.; Incoronato, M.; Condorelli, G.; Banfi, S.; Verde, P. miR-340 inhibits tumor cell proliferation and induces apoptosis by targeting multiple negative regulators of p27 in non-small cell lung cancer. Oncogene 2015, 34, 3240–3250. [Google Scholar] [CrossRef]
- Li, X.; Gong, X.; Chen, J.; Zhang, J.; Sun, J.; Guo, M. miR-340 inhibits glioblastoma cell proliferation by suppressing CDK6, cyclin-D1 and cyclin-D2. Biochem. Biophys. Res. Commun. 2015, 460, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Cheng, Z.; Zhang, J.; Li, R.; Zhao, P.; Fu, Z.; You, Y. hsa-mir-181a and hsa-mir-181b function as tumor suppressors in human glioma cells. Brain Res. 2008, 1236, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.-M.; Xu, L.; Giffard, R.G. Inhibition of microRNA-181 Reduces Forebrain Ischemia-Induced Neuronal Loss. J. Cereb. Blood Flow Metab. 2013, 33, 1976–1982. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Log2FC | p Value | Mature Sequence mirBASE 22.1 | hTLR8 | hTLR7/8 | GU % | AU % |
|---|---|---|---|---|---|---|---|
| Media | |||||||
| mmu-miR-128-1-5p | 2.9 | 0.0005 | CGGGGCCGUAGCACUGUCUGA | 57.1 | 33.3 | ||
| mmu-miR-128-2-5p | 2.6 | 0.0198 | GGGGGCCGAUGCACUGUAAGAGA | 56.5 | 39.1 | ||
| mmu-miR-6240 | 1.8 | 0.0363 | CCAAAGCAUCGCGAAGGCCCACGGG | 34.6 | 30.8 | ||
| mmu-miR-340-3p | 1.4 | 0.0001 | UCCGUCUCAGUUACUUUAUAGC | UUAUA | UCUC | 54.5 | 59.1 |
| mmu-miR-129-2-3p | 1.4 | 0.0316 | AAGCCCUUACCCCAAAAAGCAU | 22.7 | 54.5 | ||
| mmu-miR-7068-3p | 1.3 | 0.0363 | UCACCCUGGACUGACUCUCAG | 42.9 | 42.9 | ||
| mmu-miR-132-5p | 1.1 | 0.0391 | AACCGUGGCUUUCGAUUGUUAC | UUUC UUGU | 59.1 | 54.5 | |
| mmu-miR-137-3p | 1.1 | 0.0410 | UUAUUGCUUAAGAAUACGCGUAG | UUAUU | 56.5 | 65.2 | |
| mmu-miR-181a-1-3p | 1.1 | 0.0000 | ACCAUCGACCGUUGAUUGUACC | GUAC | UUGU | 45.5 | 50.0 |
| mmu-miR-181b-5p | 1.0 | 0.0373 | AACAUUCAUUGCUGUCGGUGGGUU | 66.7 | 54.2 | ||
| mmu-let-7d-5p | 1.0 | 0.0000 | AGAGGUAGUAGGUUGCAUAGUU | 68.2 | 59.1 | ||
| mmu-let-7e-5p | 1.0 | 0.0013 | UGAGGUAGGAGGUUGUAUAGUU | UAUA | UUGU | 77.3 | 59.1 |
| Neurons | |||||||
| mmu-miR-128-2-5p | −3.5 | 0.0000 | GGGGGCCGAUGCACUGUAAGAGA | 56.5 | 39.1 | ||
| mmu-miR-128-1-5p | −2.8 | 0.0000 | CGGGGCCGUAGCACUGUCUGA | 57.1 | 33.3 | ||
| mmu-miR-410-5p | −2.7 | 0.0072 | AGGUUGUCUGUGAUGAGUUCG | UUGU | 76.2 | 52.4 | |
| mmu-miR-409-5p | −2.7 | 0.0000 | AGGUUACCCGAGCAACUUUGCAU | 47.8 | 52.2 | ||
| mmu-miR-124-5p | −2.6 | 0.0004 | CGUGUUCACAGCGGACCUUGAU | UGUUC | 54.5 | 45.5 | |
| mmu-miR-341-5p | −2.1 | 0.0051 | CGGUCGGCCGAUCGCUCGGUC | 57.1 | 23.8 | ||
| mmu-miR-92a-1-5p | −1.9 | 0.0000 | AGGUUGGGAUUUGUCGCAAUGCU | UUGU | 69.6 | 52.2 | |
| mmu-miR-30f | −1.8 | 0.0051 | GUAAACAUCCGACUGAAAGCUC | 36.4 | 54.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wallach, T.; Wetzel, M.; Dembny, P.; Staszewski, O.; Krüger, C.; Buonfiglioli, A.; Prinz, M.; Lehnardt, S. Identification of CNS Injury-Related microRNAs as Novel Toll-Like Receptor 7/8 Signaling Activators by Small RNA Sequencing. Cells 2020, 9, 186. https://doi.org/10.3390/cells9010186
Wallach T, Wetzel M, Dembny P, Staszewski O, Krüger C, Buonfiglioli A, Prinz M, Lehnardt S. Identification of CNS Injury-Related microRNAs as Novel Toll-Like Receptor 7/8 Signaling Activators by Small RNA Sequencing. Cells. 2020; 9(1):186. https://doi.org/10.3390/cells9010186
Chicago/Turabian StyleWallach, Thomas, Max Wetzel, Paul Dembny, Ori Staszewski, Christina Krüger, Alice Buonfiglioli, Marco Prinz, and Seija Lehnardt. 2020. "Identification of CNS Injury-Related microRNAs as Novel Toll-Like Receptor 7/8 Signaling Activators by Small RNA Sequencing" Cells 9, no. 1: 186. https://doi.org/10.3390/cells9010186
APA StyleWallach, T., Wetzel, M., Dembny, P., Staszewski, O., Krüger, C., Buonfiglioli, A., Prinz, M., & Lehnardt, S. (2020). Identification of CNS Injury-Related microRNAs as Novel Toll-Like Receptor 7/8 Signaling Activators by Small RNA Sequencing. Cells, 9(1), 186. https://doi.org/10.3390/cells9010186