A Review of FoxO1-Regulated Metabolic Diseases and Related Drug Discoveries


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
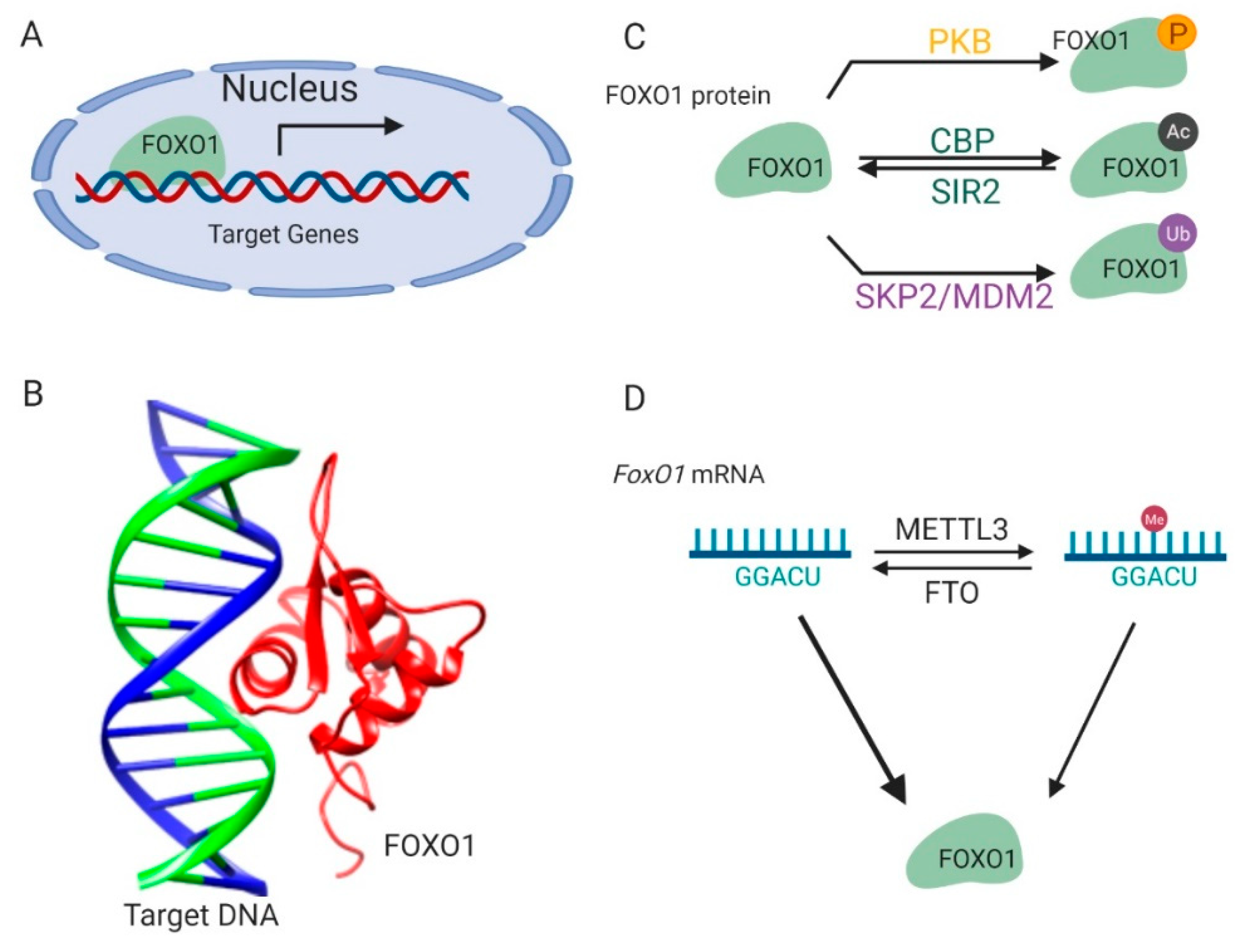
1. Introduction of FoxO1
2. Regulation of FoxO1
2.1. Protein Modifications
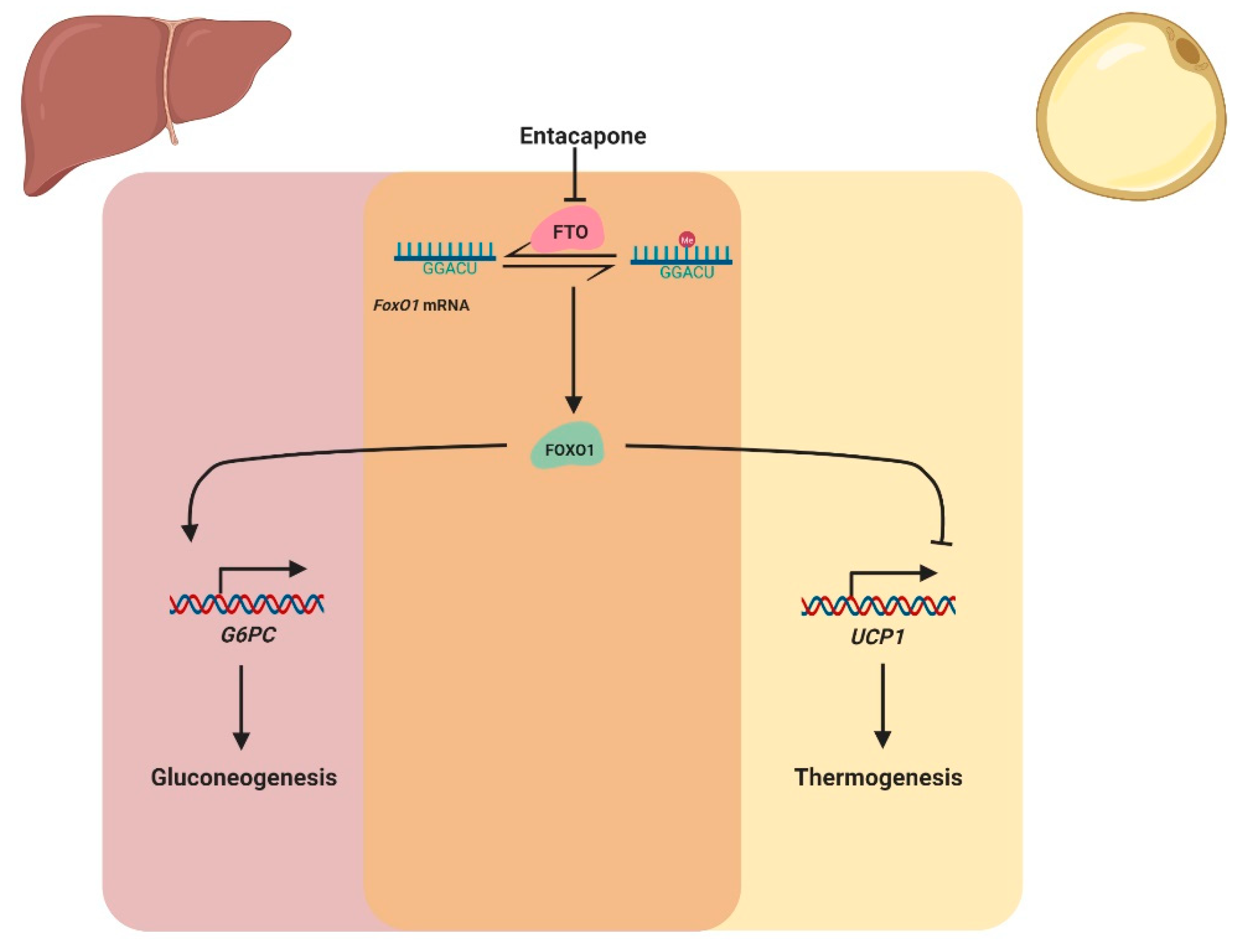
2.2. mRNA Methylation
3. FoxO1-Regulating Mechanism Behind Diseases
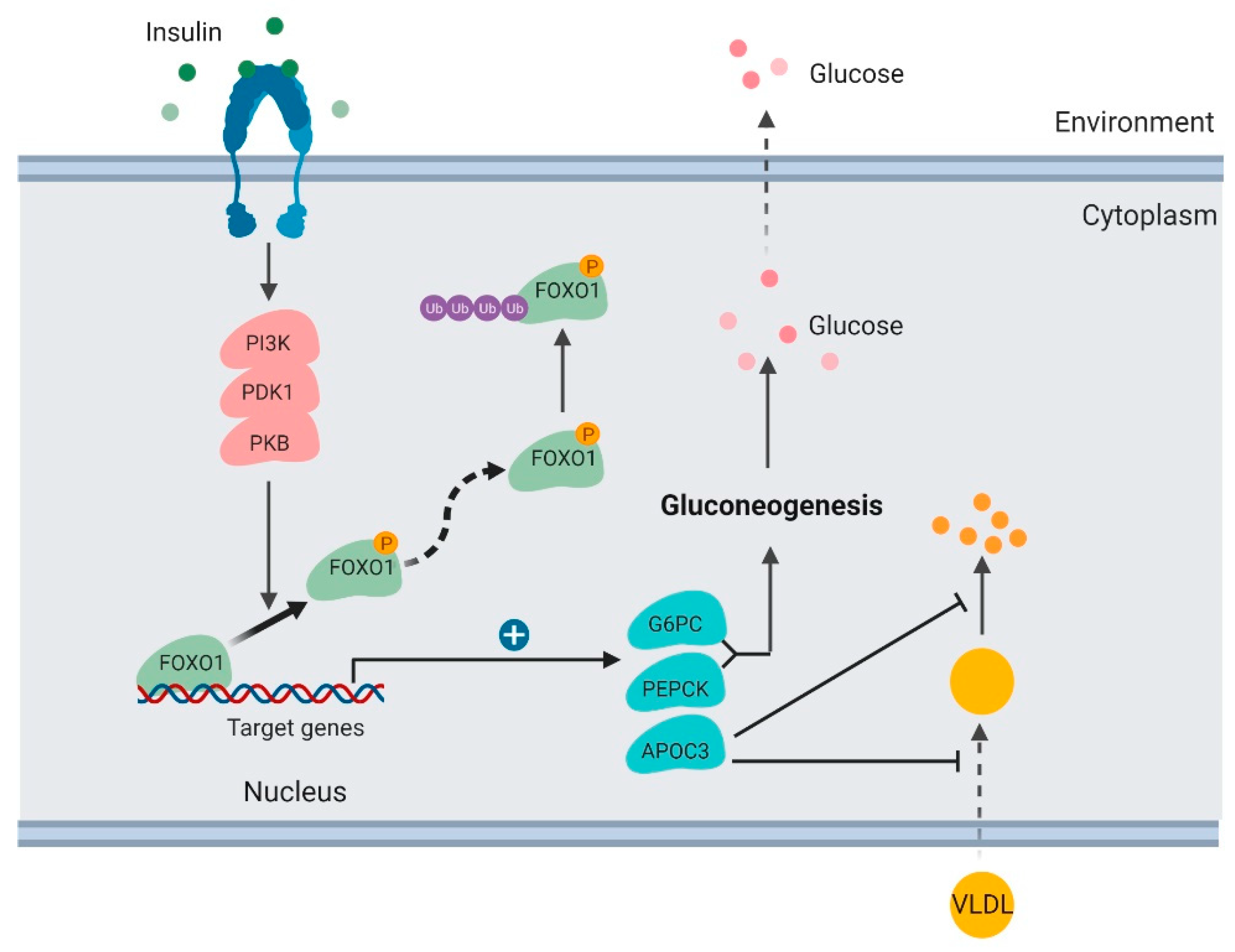
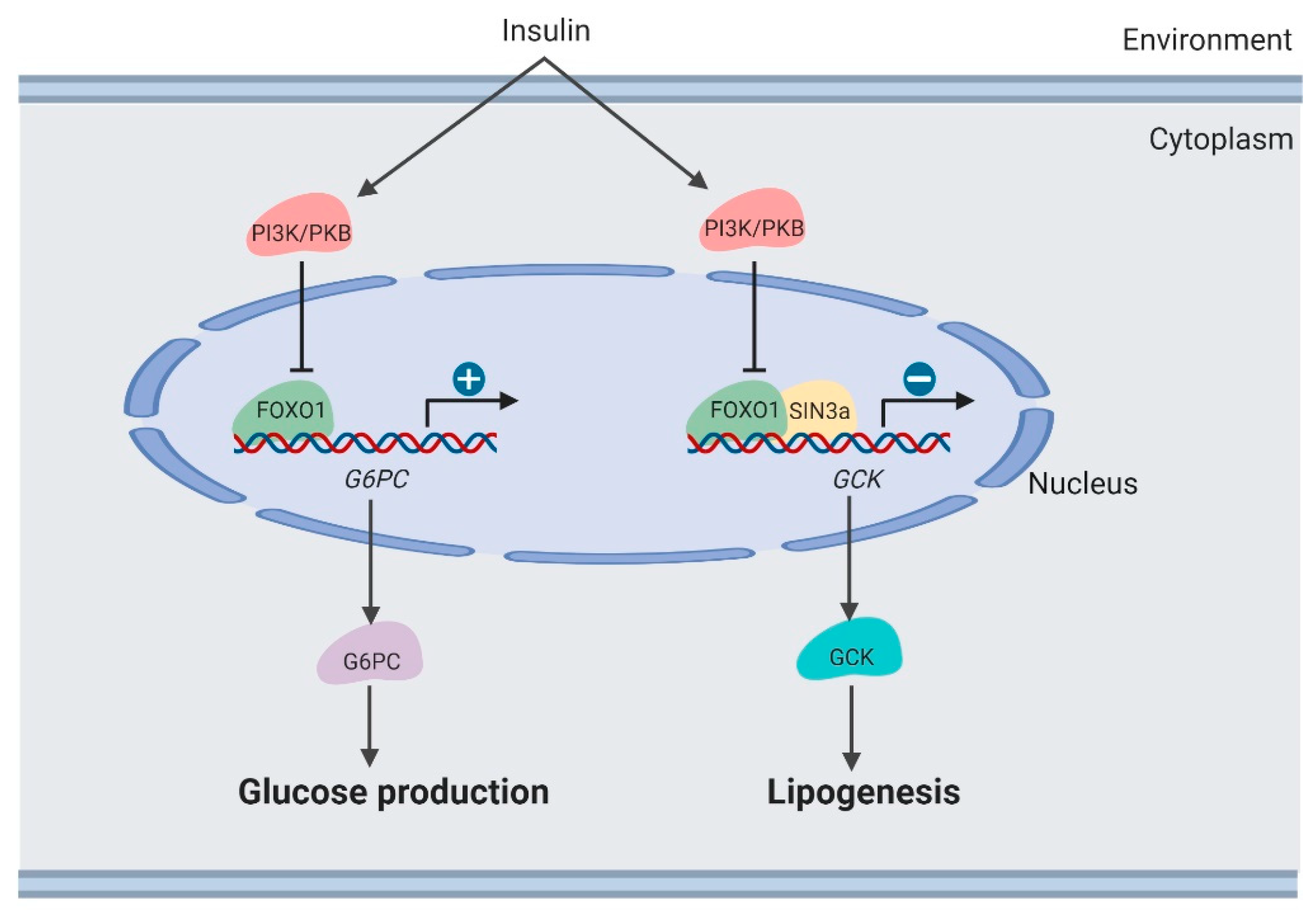
3.1. Glucose Production in the Liver
3.2. Lipoprotein Uptake in the Liver
3.3. Lipogenesis in the Liver
3.4. Lipid Metabolism in Adipocytes
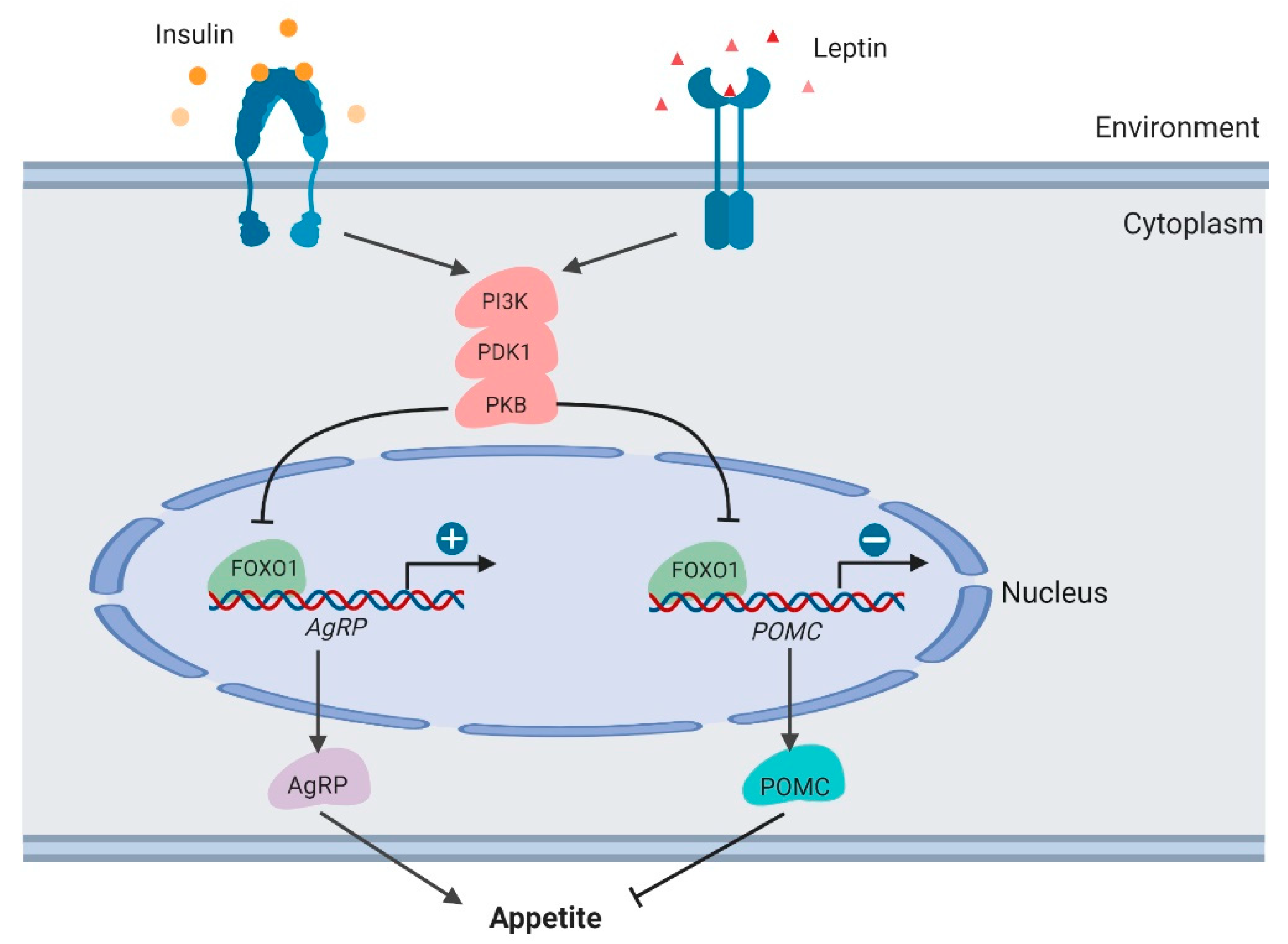
3.5. Feeding Behavior in the Hypothalamus
4. FoxO1-Related Drug Discovery
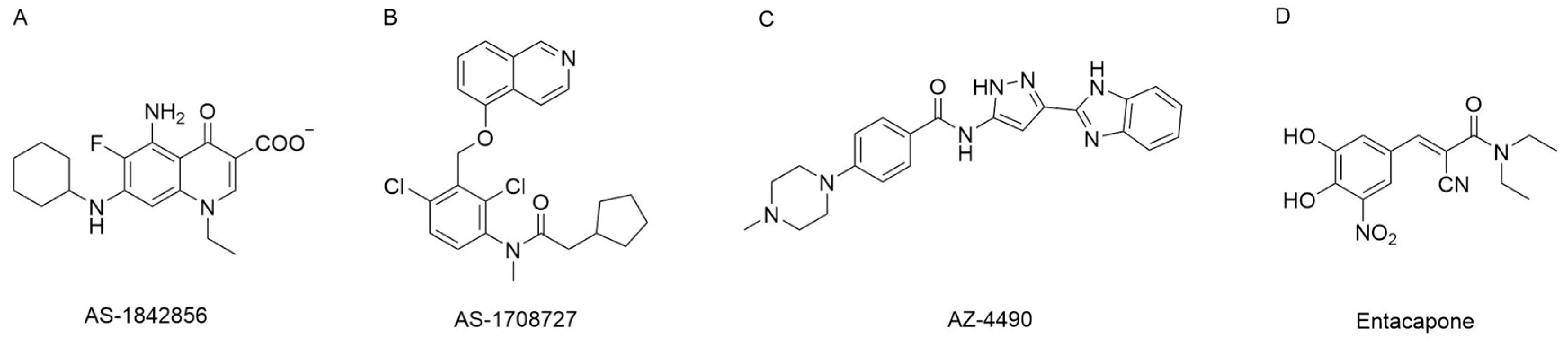
4.1. FOXO1 Protein Inhibitors
4.2. FoxO1 Pathway Modulators
Author Contributions
Funding
Conflicts of Interest
References
- Anderson, M.J.; Viars, C.S.; Czekay, S.; Cavenee, W.K.; Arden, K.C. Cloning and characterization of three human forkhead genes that comprise an fkhr-like gene subfamily. Genomics 1998, 47, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; Brunet, A. Foxo transcription factors. Curr. Biol. 2007, 17, R113–R114. [Google Scholar] [CrossRef]
- Galili, N.; Davis, R.J.; Fredericks, W.J.; Mukhopadhyay, S.; Rauscher, F.J., III; Emanuel, B.S.; Rovera, G.; Barr, F.G. Fusion of a fork head domain gene to pax3 in the solid tumour alveolar rhabdomyosarcoma. Nat. Genet. 1993, 5, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Boura, E.; Silhan, J.; Herman, P.; Vecer, J.; Sulc, M.; Teisinger, J.; Obsilova, V.; Obsil, T. Both the n-terminal loop and wing w2 of the forkhead domain of transcription factor foxo4 are important for DNA binding. J. Biol. Chem. 2007, 282, 8265–8275. [Google Scholar] [CrossRef]
- Brent, M.M.; Anand, R.; Marmorstein, R. Structural basis for DNA recognition by foxo1 and its regulation by posttranslational modification. Structure 2008, 16, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Psenakova, K.; Kohoutova, K.; Obsilova, V.; Ausserlechner, M.J.; Veverka, V.; Obsil, T. Forkhead domains of foxo transcription factors differ in both overall conformation and dynamics. Cells 2019, 8, 966. [Google Scholar] [CrossRef]
- Avila-Flores, A.; Arranz-Nicolas, J.; Merida, I. Transcriptional activity of foxo transcription factors measured by luciferase assays. Methods Mol. Biol. 2019, 1890, 91–102. [Google Scholar]
- Linardic, C.M. Pax3-foxo1 fusion gene in rhabdomyosarcoma. Cancer Lett. 2008, 270, 10–18. [Google Scholar] [CrossRef]
- Missiaglia, E.; Williamson, D.; Chisholm, J.; Wirapati, P.; Pierron, G.; Petel, F.; Concordet, J.P.; Thway, K.; Oberlin, O.; Pritchard-Jones, K.; et al. Pax3/foxo1 fusion gene status is the key prognostic molecular marker in rhabdomyosarcoma and significantly improves current risk stratification. J. Clin. Oncol. 2012, 30, 1670–1677. [Google Scholar] [CrossRef]
- Xing, Y.Q.; Li, A.; Yang, Y.; Li, X.X.; Zhang, L.N.; Guo, H.C. The regulation of foxo1 and its role in disease progression. Life Sci. 2018, 193, 124–131. [Google Scholar] [CrossRef]
- Dong, X.C. Foxo transcription factors in non-alcoholic fatty liver disease. Liver Res. 2017, 1, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Eijkelenboom, A.; Burgering, B.M. Foxos: Signalling integrators for homeostasis maintenance. Nat. Rev. Mol. Cell Biol. 2013, 14, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Link, W.; Fernandez-Marcos, P.J. Foxo transcription factors at the interface of metabolism and cancer. Int. J. Cancer 2017, 141, 2379–2391. [Google Scholar] [CrossRef]
- Maiese, K. Foxo transcription factors and regenerative pathways in diabetes mellitus. Curr. Neurovasc. Res. 2015, 12, 404–413. [Google Scholar] [CrossRef]
- Wagatsuma, A.; Shiozuka, M.; Takayama, Y.; Hoshino, T.; Mabuchi, K.; Matsuda, R. Effects of ageing on expression of the muscle-specific e3 ubiquitin ligases and akt-dependent regulation of foxo transcription factors in skeletal muscle. Mol. Cell. Biochem. 2016, 412, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K.; Daitoku, H.; Takahashi, Y.; Namiki, K.; Hisatake, K.; Kako, K.; Mukai, H.; Kasuya, Y.; Fukamizu, A. Arginine methylation of foxo transcription factors inhibits their phosphorylation by akt. Mol. Cell 2008, 32, 221–231. [Google Scholar] [CrossRef]
- Biggs, W.H., III; Meisenhelder, J.; Hunter, T.; Cavenee, W.K.; Arden, K.C. Protein kinase b/akt-mediated phosphorylation promotes nuclear exclusion of the winged helix transcription factor fkhr1. Proc. Natl. Acad. Sci. USA 1999, 96, 7421–7426. [Google Scholar] [CrossRef]
- Grabiec, A.M.; Angiolilli, C.; Hartkamp, L.M.; van Baarsen, L.G.; Tak, P.P.; Reedquist, K.A. Jnk-dependent downregulation of foxo1 is required to promote the survival of fibroblast-like synoviocytes in rheumatoid arthritis. Ann. Rheumatic Dis. 2015, 74, 1763–1771. [Google Scholar] [CrossRef]
- Yuan, Z.; Lehtinen, M.K.; Merlo, P.; Villen, J.; Gygi, S.; Bonni, A. Regulation of neuronal cell death by mst1-foxo1 signaling. J. Biol. Chem. 2009, 284, 11285–11292. [Google Scholar] [CrossRef]
- Brunet, A.; Sweeney, L.B.; Sturgill, J.F.; Chua, K.F.; Greer, P.L.; Lin, Y.; Tran, H.; Ross, S.E.; Mostoslavsky, R.; Cohen, H.Y.; et al. Stress-dependent regulation of foxo transcription factors by the sirt1 deacetylase. Science 2004, 303, 2011–2015. [Google Scholar] [CrossRef]
- Pramanik, K.C.; Fofaria, N.M.; Gupta, P.; Srivastava, S.K. Cbp-mediated foxo-1 acetylation inhibits pancreatic tumor growth by targeting sirt. Mol. Cancer Ther. 2014, 13, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Van der Heide, L.P.; Smidt, M.P. Regulation of foxo activity by cbp/p300-mediated acetylation. Trends Biochem. Sci. 2005, 30, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Ma, Q.; Chen, L.; Li, P.; Zhang, M.; Ramamoorthy, S.; Nawaz, Z.; Shimojima, T.; Wang, H.; Yang, Y.; et al. Mdm2 acts downstream of p53 as an e3 ligase to promote foxo ubiquitination and degradation. J. Biol. Chem. 2009, 284, 13987–14000. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Tindall, D.J. Regulation of foxo protein stability via ubiquitination and proteasome degradation. Biochim. Biophys. Acta 2011, 1813, 1961–1964. [Google Scholar] [CrossRef] [PubMed]
- Vogt, P.K.; Jiang, H.; Aoki, M. Triple layer control: Phosphorylation, acetylation and ubiquitination of foxo proteins. Cell Cycle 2005, 4, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6a rna methylomes revealed by m6a-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mrna methylation reveals enrichment in 3′ utrs and near stop codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef]
- Peng, S.; Xiao, W.; Ju, D.; Sun, B.; Hou, N.; Liu, Q.; Wang, Y.; Zhao, H.; Gao, C.; Zhang, S.; et al. Identification of entacapone as a chemical inhibitor of fto mediating metabolic regulation through foxo1. Sci. Transl. Med. 2019, 11, eaau7116. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. Ucsf chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Henriques, V.; Machado, S.; Link, W.; Ferreira, B.I. Monitoring the transcriptional activity of foxo transcription factors by analyzing their target genes. Methods Mol. Biol. 2019, 1890, 103–113. [Google Scholar]
- Nakae, J.; Cao, Y.; Oki, M.; Orba, Y.; Sawa, H.; Kiyonari, H.; Iskandar, K.; Suga, K.; Lombes, M.; Hayashi, Y. Forkhead transcription factor foxo1 in adipose tissue regulates energy storage and expenditure. Diabetes 2008, 57, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Altomonte, J.; Richter, A.; Harbaran, S.; Suriawinata, J.; Nakae, J.; Thung, S.N.; Meseck, M.; Accili, D.; Dong, H. Inhibition of foxo1 function is associated with improved fasting glycemia in diabetic mice. Am. J. Physiol. Endocrinol. Metabol. 2003, 285, E718–E728. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Pocai, A.; Rossetti, L.; Depinho, R.A.; Accili, D. Impaired regulation of hepatic glucose production in mice lacking the forkhead transcription factor foxo1 in liver. Cell Metabol. 2007, 6, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Pan, Q.; Yan, H.; Zhang, K.; Guo, X.; Xu, Z.; Yang, W.; Qi, Y.; Guo, C.A.; Hornsby, C.; et al. Novel mechanism of foxo1 phosphorylation in glucagon signaling in control of glucose homeostasis. Diabetes 2018, 67, 2167–2182. [Google Scholar] [CrossRef]
- Altomonte, J.; Cong, L.; Harbaran, S.; Richter, A.; Xu, J.; Meseck, M.; Dong, H.H. Foxo1 mediates insulin action on apoc-iii and triglyceride metabolism. J. Clin. Investig. 2004, 114, 1493–1503. [Google Scholar] [CrossRef]
- Mendivil, C.O.; Zheng, C.; Furtado, J.; Lel, J.; Sacks, F.M. Metabolism of very-low-density lipoprotein and low-density lipoprotein containing apolipoprotein c-iii and not other small apolipoproteins. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 239–245. [Google Scholar] [CrossRef]
- Bobik, A. Apolipoprotein ciii and atherosclerosis: Beyond effects on lipid metabolism. Circulation 2008, 118, 702–704. [Google Scholar] [CrossRef]
- Haeusler, R.A.; Hartil, K.; Vaitheesvaran, B.; Arrieta-Cruz, I.; Knight, C.M.; Cook, J.R.; Kammoun, H.L.; Febbraio, M.A.; Gutierrez-Juarez, R.; Kurland, I.J.; et al. Integrated control of hepatic lipogenesis versus glucose production requires foxo transcription factors. Nat. Commun. 2014, 5, 5190. [Google Scholar] [CrossRef]
- Langlet, F.; Haeusler, R.A.; Linden, D.; Ericson, E.; Norris, T.; Johansson, A.; Cook, J.R.; Aizawa, K.; Wang, L.; Buettner, C.; et al. Selective inhibition of foxo1 activator/repressor balance modulates hepatic glucose handling. Cell 2017, 171, 824–835. [Google Scholar] [CrossRef]
- Farmer, S.R. The forkhead transcription factor foxo1: A possible link between obesity and insulin resistance. Mol. Cell 2003, 11, 6–8. [Google Scholar] [CrossRef]
- Nakae, J.; Kitamura, T.; Kitamura, Y.; Biggs, W.H., III; Arden, K.C.; Accili, D. The forkhead transcription factor foxo1 regulates adipocyte differentiation. Dev. Cell 2003, 4, 119–129. [Google Scholar] [CrossRef]
- Kita, M.; Nakae, J.; Kawano, Y.; Asahara, H.; Takemori, H.; Okado, H.; Itoh, H. Zfp238 regulates the thermogenic program in cooperation with foxo1. iScience 2019, 12, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef]
- Inagaki, T.; Sakai, J.; Kajimura, S. Transcriptional and epigenetic control of brown and beige adipose cell fate and function. Nat. Rev. Mol. Cell Biol. 2016, 17, 480–495. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Cohen, P.; Spiegelman, B.M. Adaptive thermogenesis in adipocytes: Is beige the new brown? Genes Dev. 2013, 27, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Varela, L.; Horvath, T.L. Leptin and insulin pathways in pomc and agrp neurons that modulate energy balance and glucose homeostasis. EMBO Rep. 2012, 13, 1079–1086. [Google Scholar] [CrossRef]
- Belgardt, B.F.; Okamura, T.; Bruning, J.C. Hormone and glucose signalling in pomc and agrp neurons. J. Physiol. 2009, 587, 5305–5314. [Google Scholar] [CrossRef]
- Breen, T.L.; Conwell, I.M.; Wardlaw, S.L. Effects of fasting, leptin, and insulin on agrp and pomc peptide release in the hypothalamus. Brain Res. 2005, 1032, 141–148. [Google Scholar] [CrossRef]
- Belgardt, B.F.; Husch, A.; Rother, E.; Ernst, M.B.; Wunderlich, F.T.; Hampel, B.; Klockener, T.; Alessi, D.; Kloppenburg, P.; Bruning, J.C. Pdk1 deficiency in pomc-expressing cells reveals foxo1-dependent and -independent pathways in control of energy homeostasis and stress response. Cell Metab. 2008, 7, 291–301. [Google Scholar] [CrossRef]
- Cao, Y.; Nakata, M.; Okamoto, S.; Takano, E.; Yada, T.; Minokoshi, Y.; Hirata, Y.; Nakajima, K.; Iskandar, K.; Hayashi, Y.; et al. Pdk1-foxo1 in agouti-related peptide neurons regulates energy homeostasis by modulating food intake and energy expenditure. PLoS ONE 2011, 6, e18324. [Google Scholar] [CrossRef]
- Iskandar, K.; Cao, Y.; Hayashi, Y.; Nakata, M.; Takano, E.; Yada, T.; Zhang, C.; Ogawa, W.; Oki, M.; Chua, S., Jr.; et al. Pdk-1/foxo1 pathway in pomc neurons regulates pomc expression and food intake. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E787–E798. [Google Scholar] [CrossRef] [PubMed]
- Ropelle, E.R.; Pauli, J.R.; Prada, P.; Cintra, D.E.; Rocha, G.Z.; Moraes, J.C.; Frederico, M.J.; da Luz, G.; Pinho, R.A.; Carvalheira, J.B.; et al. Inhibition of hypothalamic foxo1 expression reduced food intake in diet-induced obesity rats. J. Physiol. 2009, 587, 2341–2351. [Google Scholar] [CrossRef] [PubMed]
- Auguste, G.; Gurha, P.; Lombardi, R.; Coarfa, C.; Willerson, J.T.; Marian, A.J. Suppression of activated foxo transcription factors in the heart prolongs survival in a mouse model of laminopathies. Circ. Res. 2018, 122, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Li, Y.; Zhang, X. The functions of foxo transcription factors in epithelial wound healing. Australasian J. Dermatol. 2019, 60, 105–109. [Google Scholar]
- O’Neill, B.T.; Bhardwaj, G.; Penniman, C.M.; Krumpoch, M.T.; Suarez Beltran, P.A.; Klaus, K.; Poro, K.; Li, M.; Pan, H.; Dreyfuss, J.M.; et al. Foxo transcription factors are critical regulators of diabetes-related muscle atrophy. Diabetes 2019, 68, 556–570. [Google Scholar] [CrossRef]
- Roupe, K.M.; Veerla, S.; Olson, J.; Stone, E.L.; Sorensen, O.E.; Hedrick, S.M.; Nizet, V. Transcription factor binding site analysis identifies foxo transcription factors as regulators of the cutaneous wound healing process. PLoS ONE 2014, 9, e89274. [Google Scholar] [CrossRef]
- Stitt, T.N.; Drujan, D.; Clarke, B.A.; Panaro, F.; Timofeyva, Y.; Kline, W.O.; Gonzalez, M.; Yancopoulos, G.D.; Glass, D.J. The igf-1/pi3k/akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting foxo transcription factors. Mol. Cell 2004, 14, 395–403. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, Y.; Graves, D.T. Foxo transcription factors: Their clinical significance and regulation. BioMed Res. Int. 2014, 2014, 925350. [Google Scholar] [CrossRef]
- Nagashima, T.; Shigematsu, N.; Maruki, R.; Urano, Y.; Tanaka, H.; Shimaya, A.; Shimokawa, T.; Shibasaki, M. Discovery of novel forkhead box o1 inhibitors for treating type 2 diabetes: Improvement of fasting glycemia in diabetic db/db mice. Mol. Pharmacol. 2010, 78, 961–970. [Google Scholar] [CrossRef]
- Zou, P.; Liu, L.; Zheng, L.; Liu, L.; Stoneman, R.E.; Cho, A.; Emery, A.; Gilbert, E.R.; Cheng, Z. Targeting foxo1 with as1842856 suppresses adipogenesis. Cell Cycle 2014, 13, 3759–3767. [Google Scholar] [CrossRef]
- Tanaka, H.; Nagashima, T.; Shimaya, A.; Urano, Y.; Shimokawa, T.; Shibasaki, M. Effects of the novel foxo1 inhibitor as1708727 on plasma glucose and triglyceride levels in diabetic db/db mice. Eur. J. Pharmacol. 2010, 645, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Johansson, A.; Petersson, M.; Fredenwall, M. Discovery of novel FOXO1 inhibitors for the treatment of diabetes. In Proceedings of the 258th ACS National Meeting & Exposition, San Diego, CA, USA, 25–29 August 2019. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, S.; Li, W.; Hou, N.; Huang, N. A Review of FoxO1-Regulated Metabolic Diseases and Related Drug Discoveries. Cells 2020, 9, 184. https://doi.org/10.3390/cells9010184
Peng S, Li W, Hou N, Huang N. A Review of FoxO1-Regulated Metabolic Diseases and Related Drug Discoveries. Cells. 2020; 9(1):184. https://doi.org/10.3390/cells9010184
Chicago/Turabian StylePeng, Shiming, Wei Li, Nannan Hou, and Niu Huang. 2020. "A Review of FoxO1-Regulated Metabolic Diseases and Related Drug Discoveries" Cells 9, no. 1: 184. https://doi.org/10.3390/cells9010184
APA StylePeng, S., Li, W., Hou, N., & Huang, N. (2020). A Review of FoxO1-Regulated Metabolic Diseases and Related Drug Discoveries. Cells, 9(1), 184. https://doi.org/10.3390/cells9010184