Heat Stress-Induced Multiple Multipolar Divisions of Human Cancer Cells


 and
and 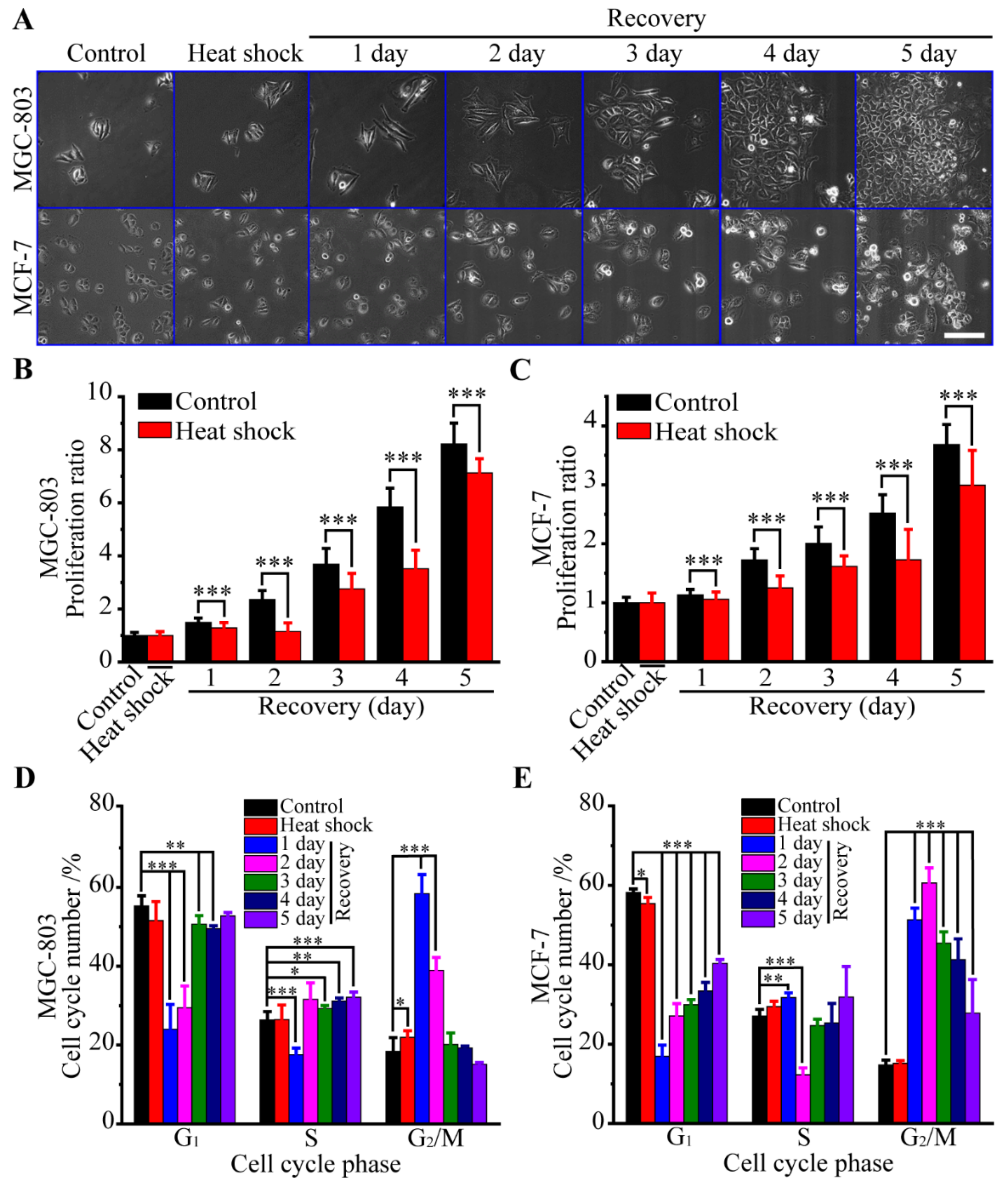{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Heat Stress Treatment
2.3. MTT Assay
2.4. Flow Cytometry
2.5. Long-Term Live Imaging
2.6. Immunofluorescence Assay
2.7. Statistical Analysis
3. Results
3.1. Biological Responses of Human Cancer Cells to Heat Stress
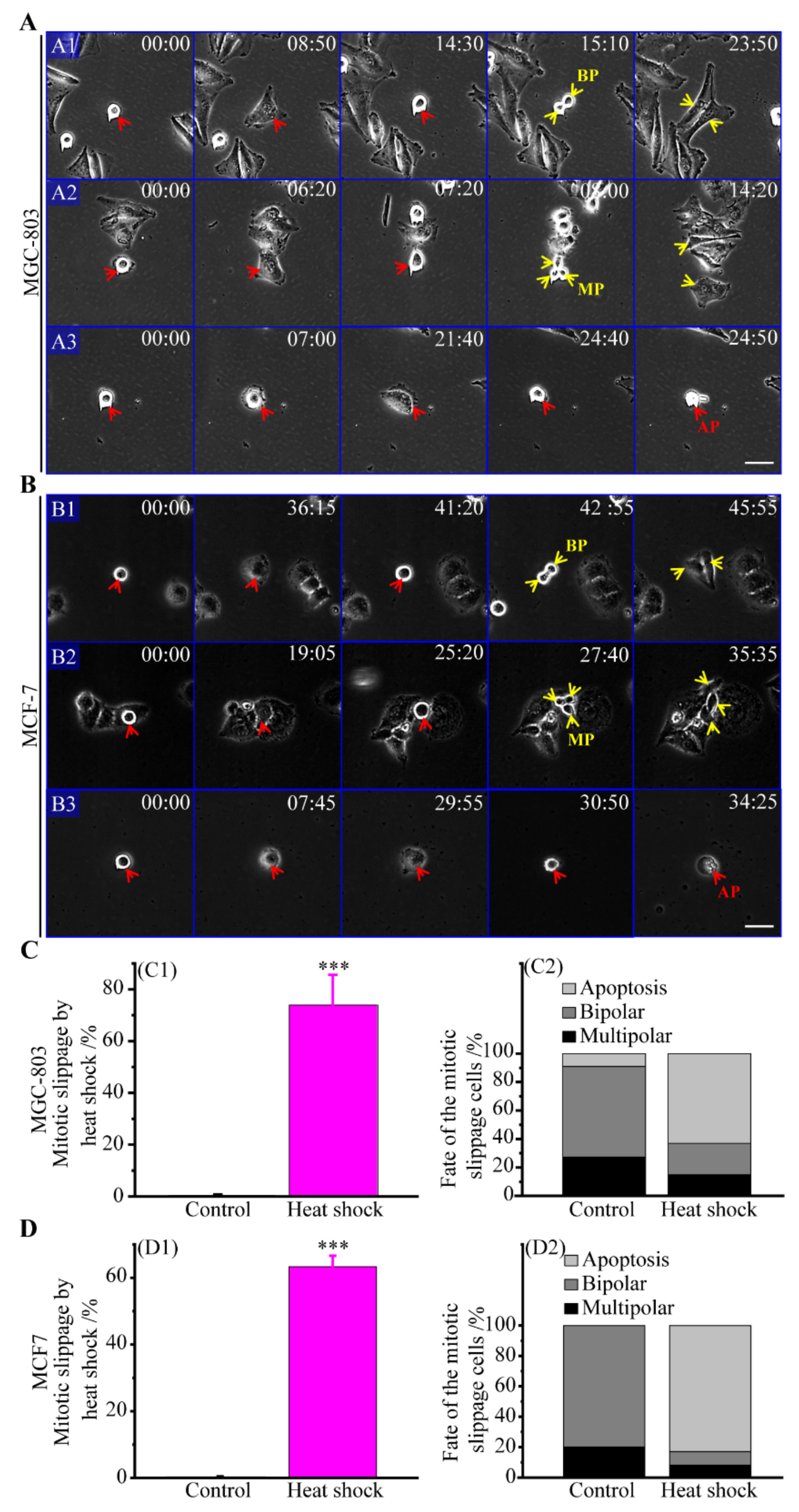
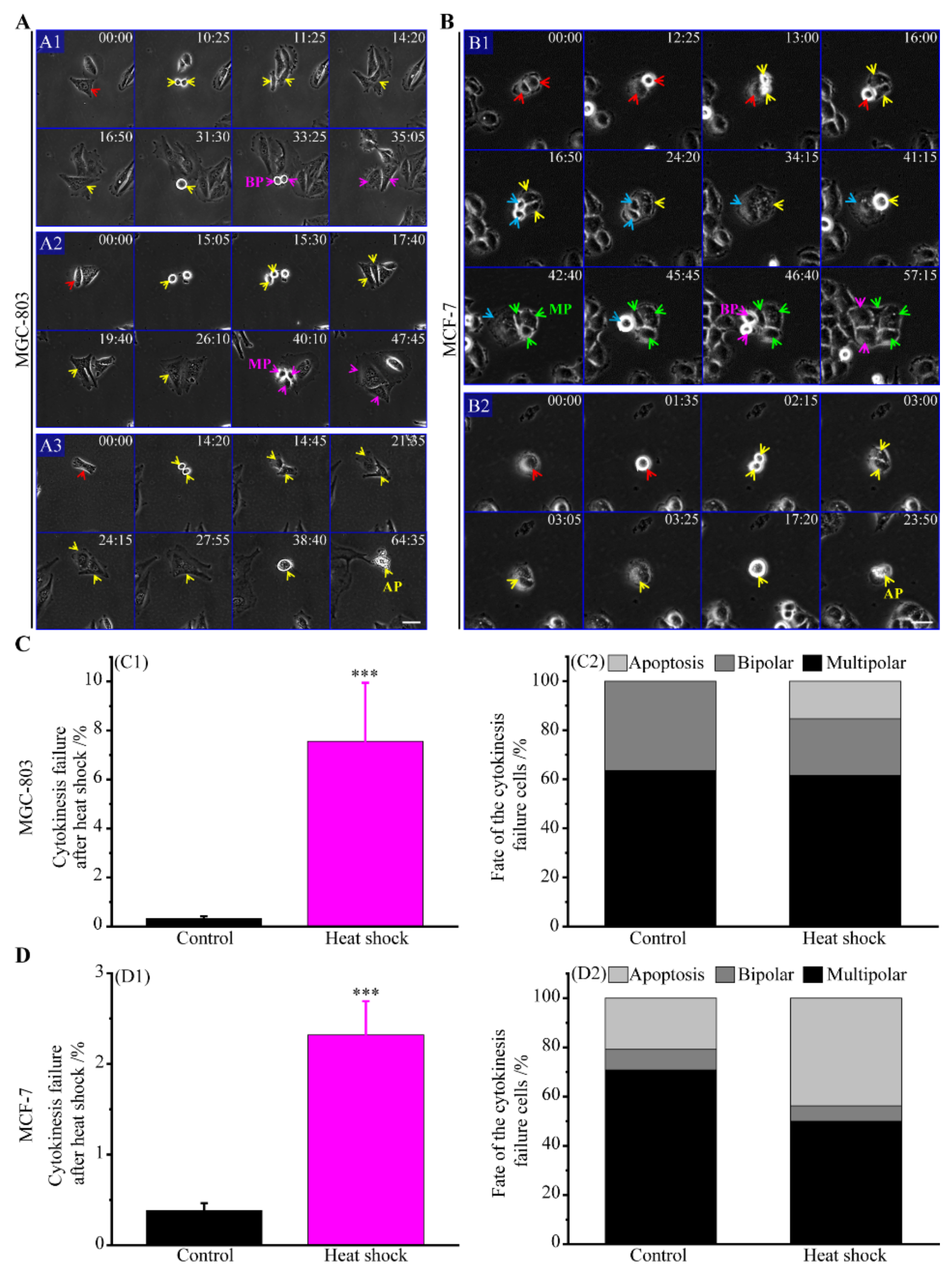
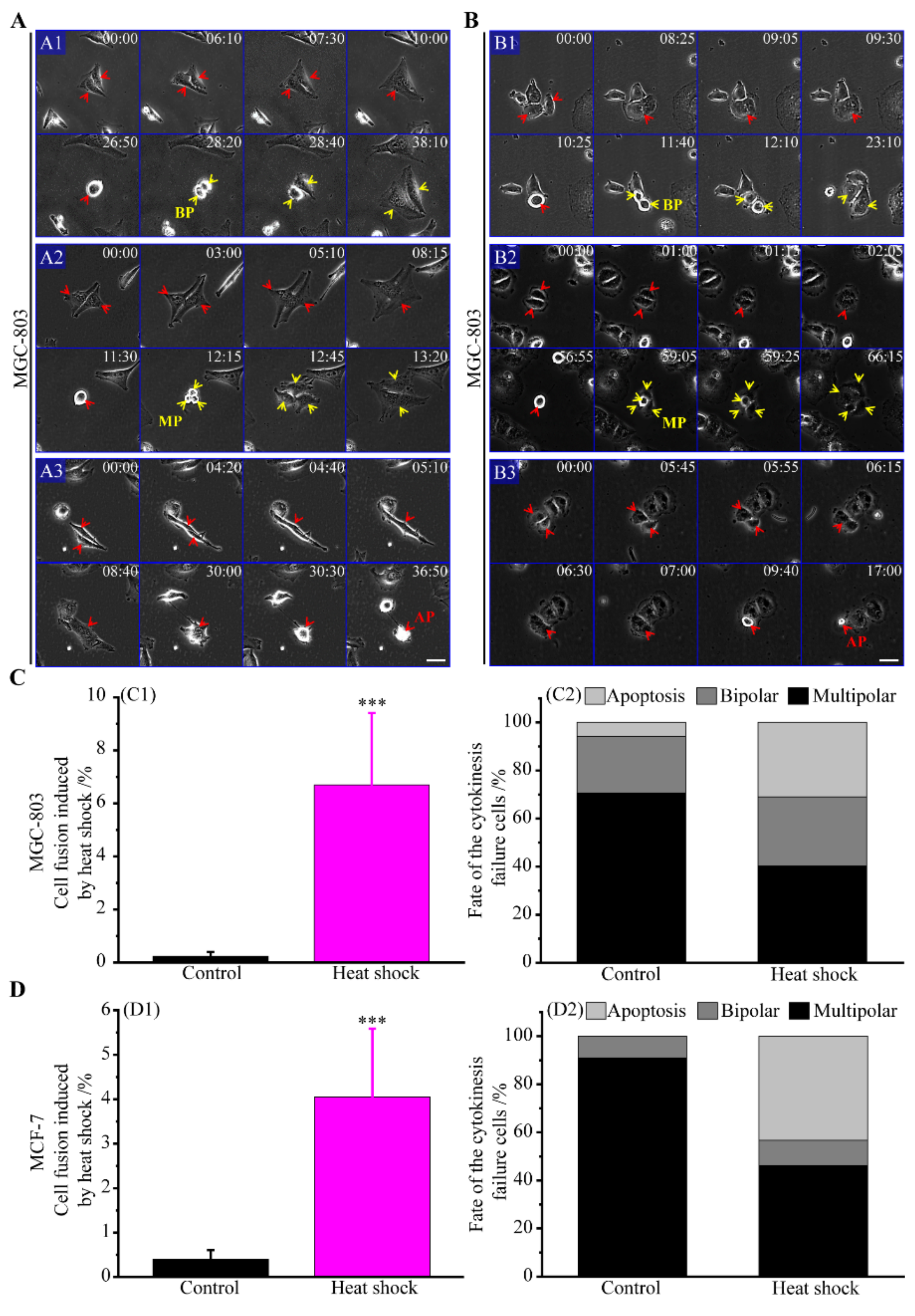
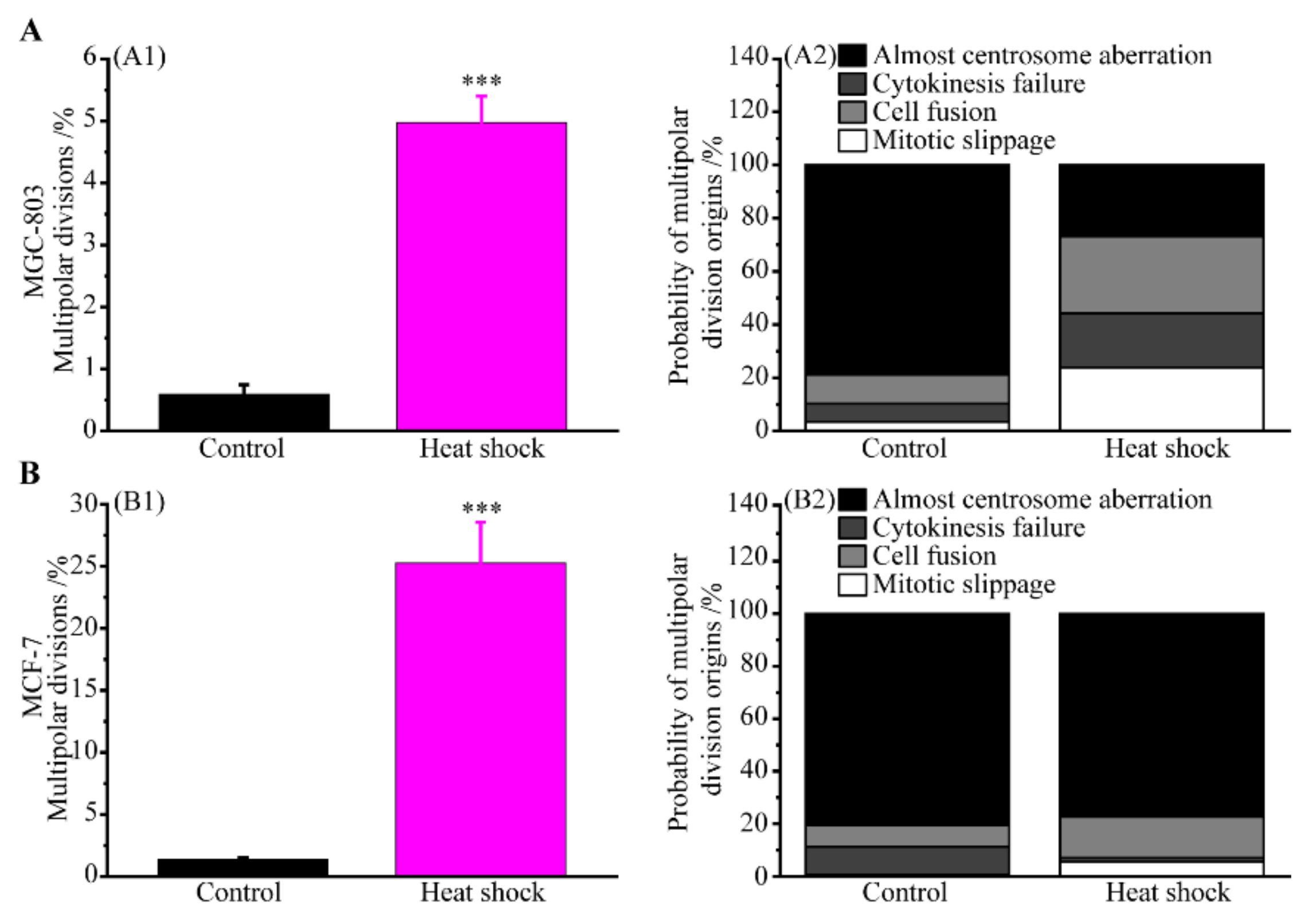
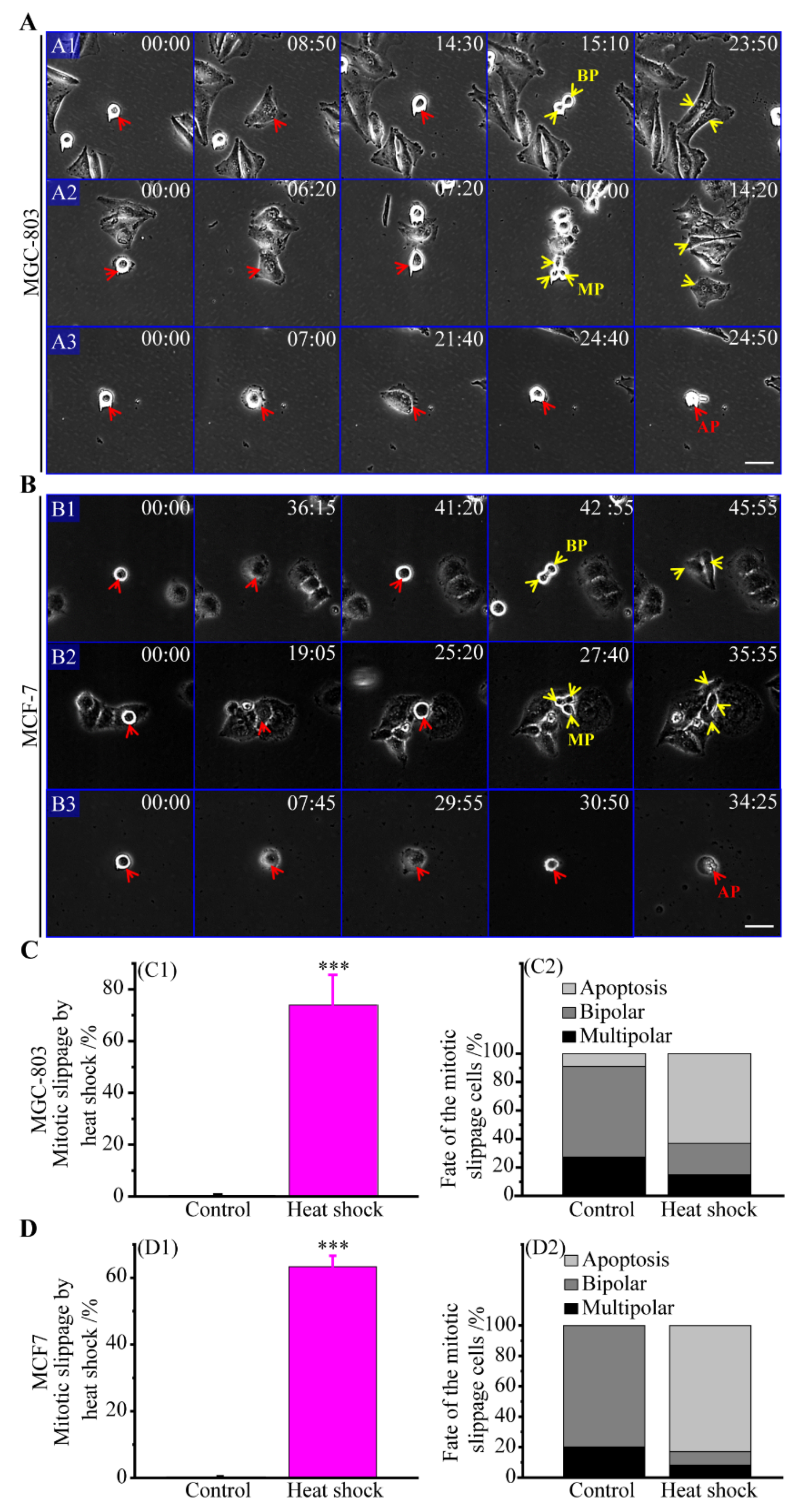
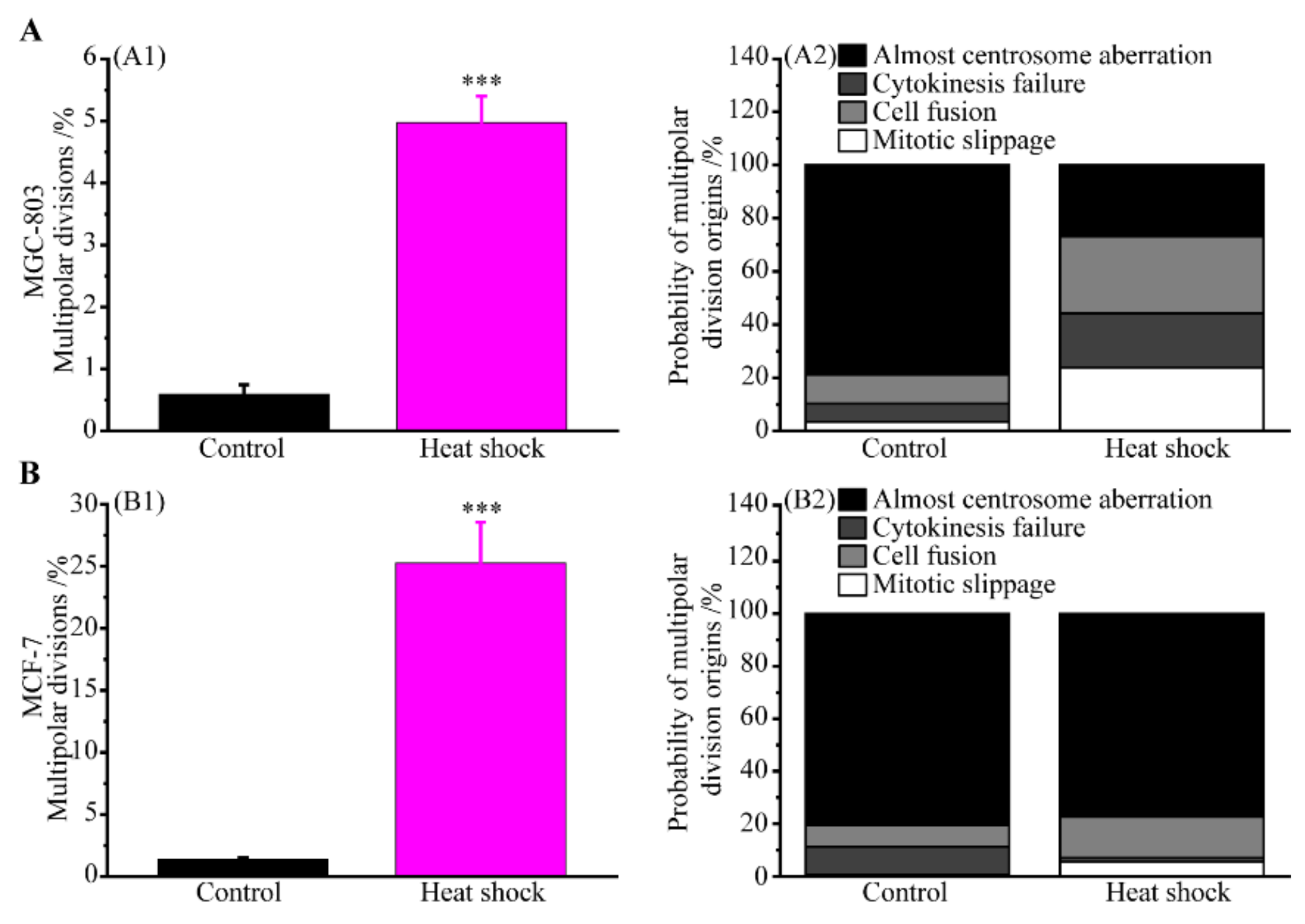
3.2. Multipolar Divisions Induced by Heat Stress
3.2.1. The Route of Mitotic Slippage
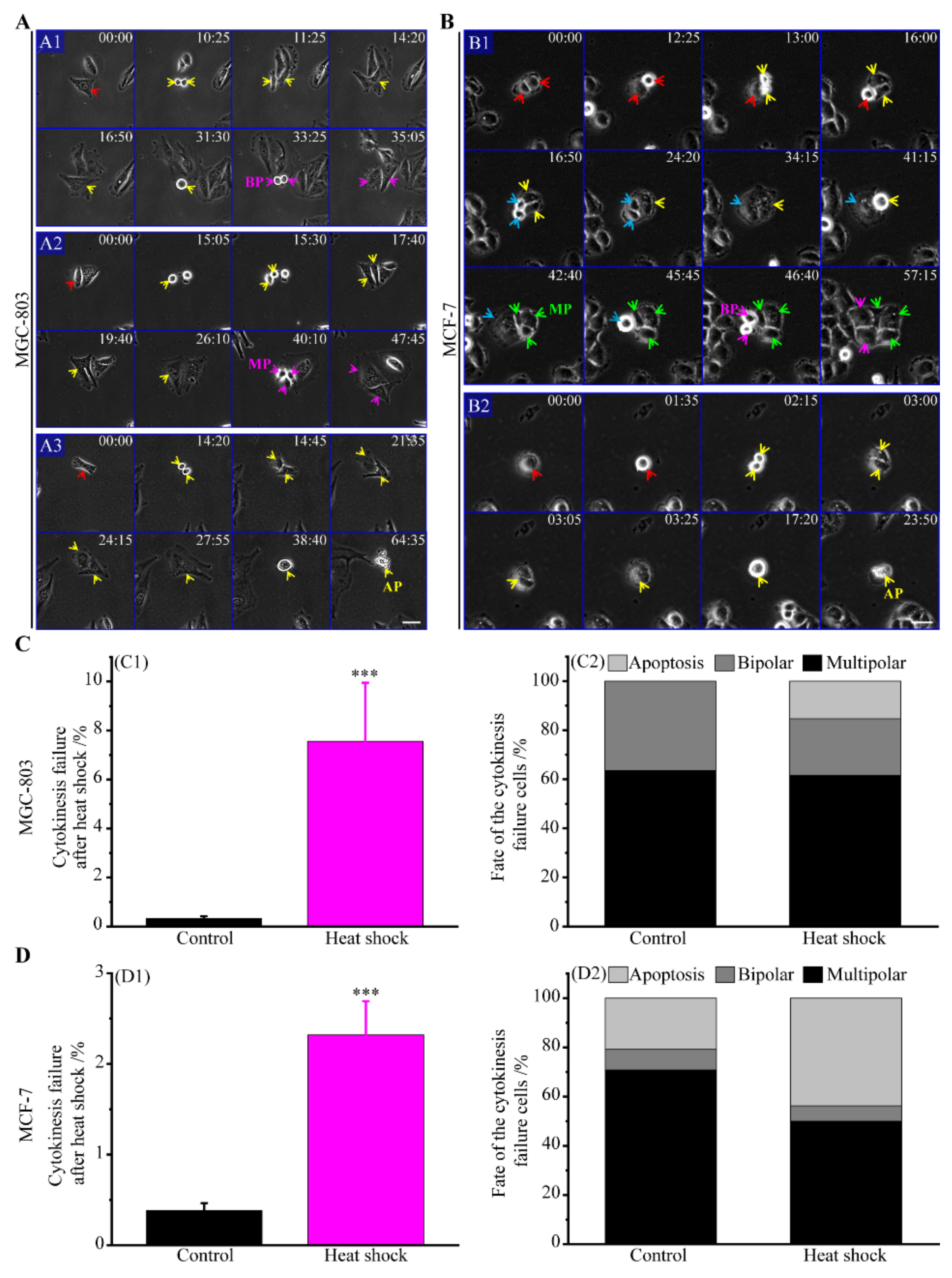
3.2.2. The Route of Cytokinesis Failure
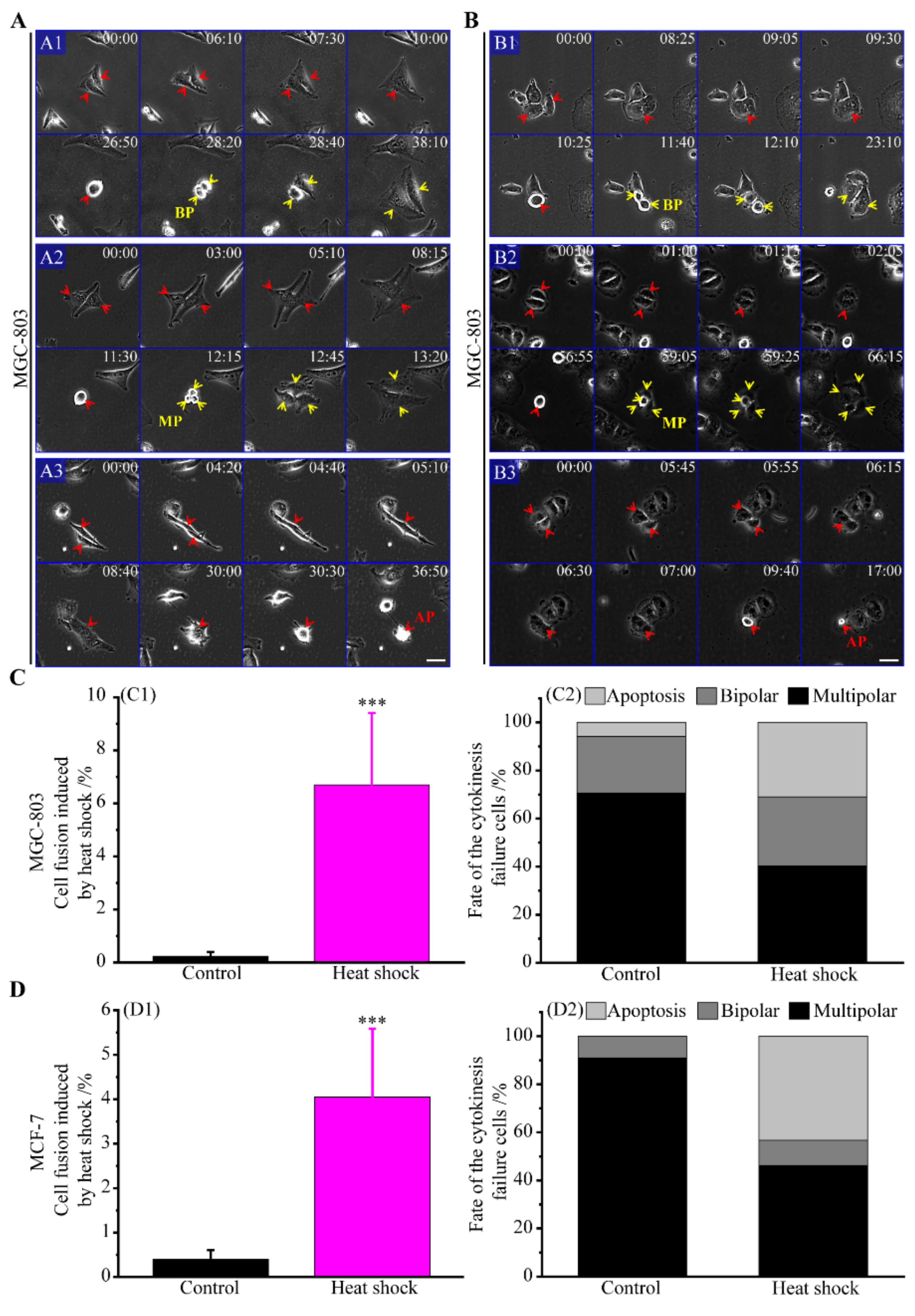
3.2.3. The Route of Cell Fusion
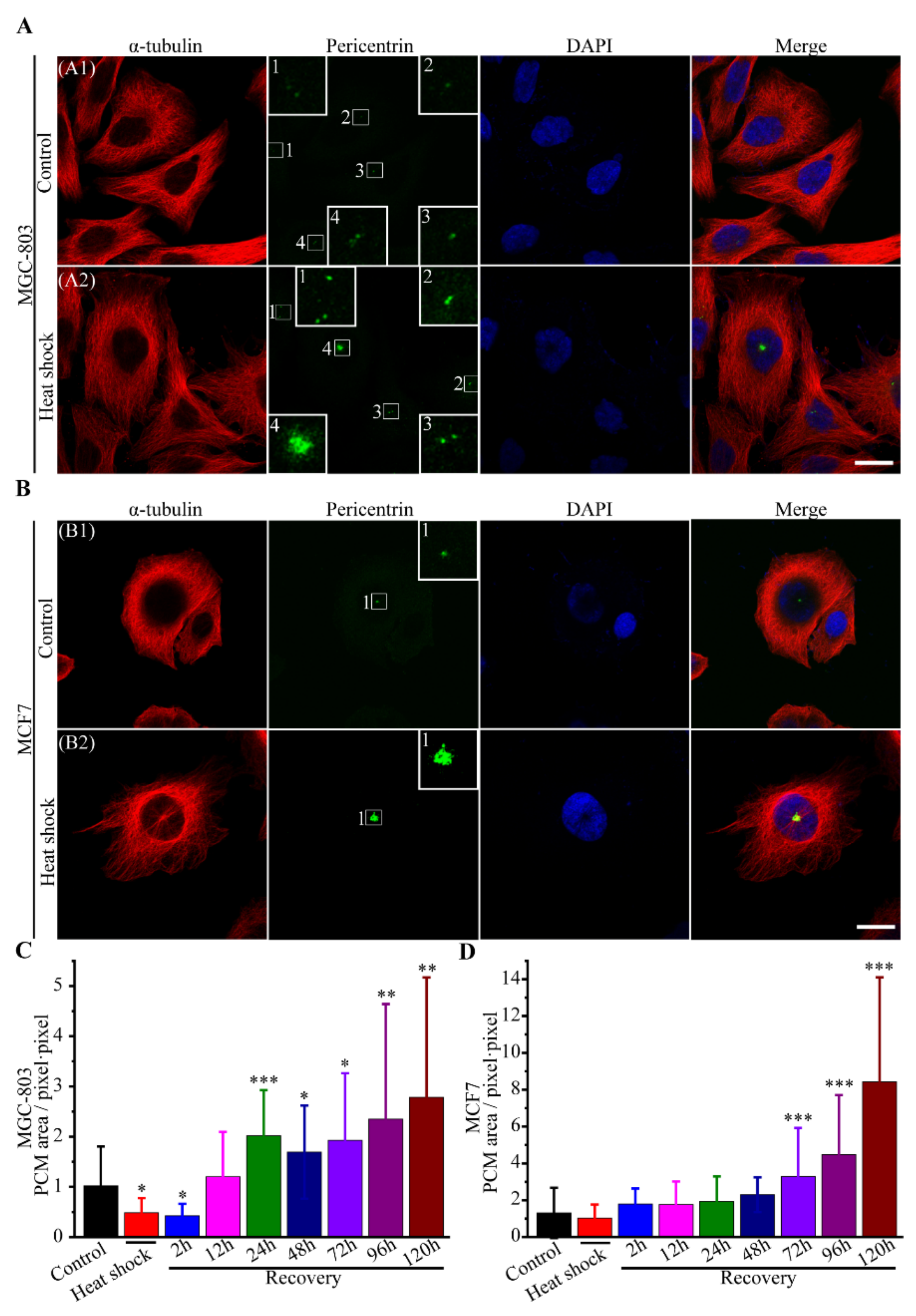
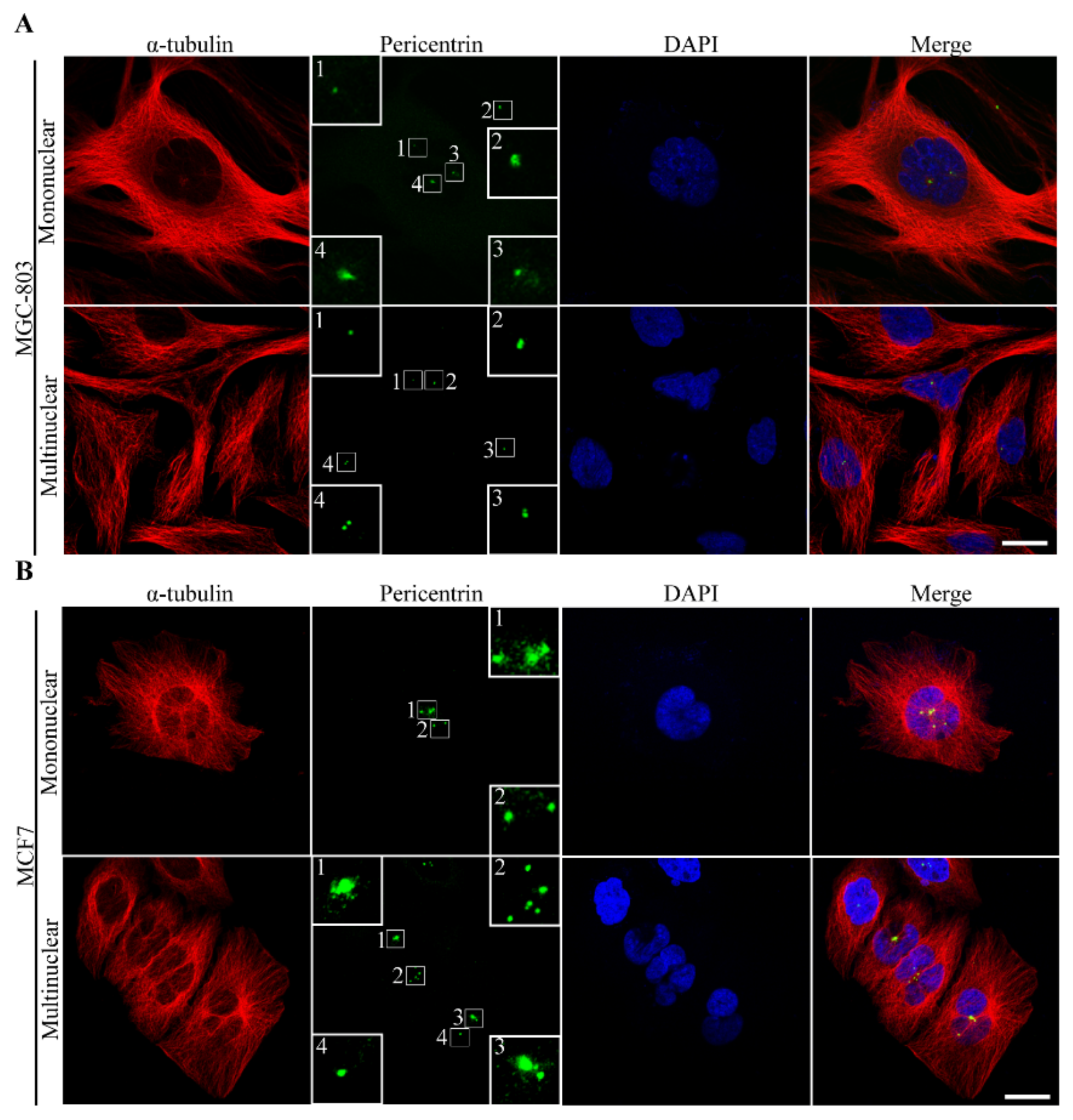
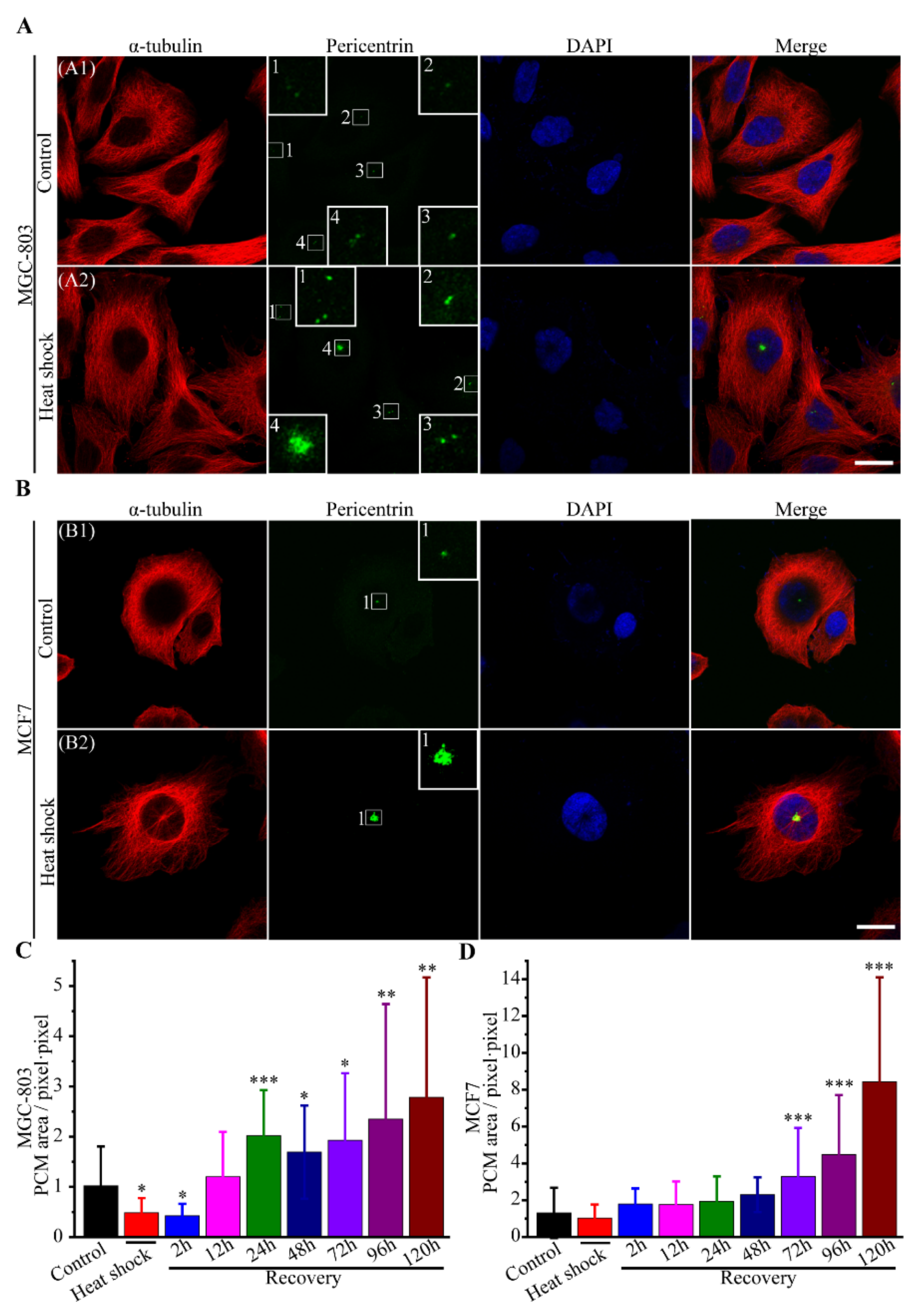
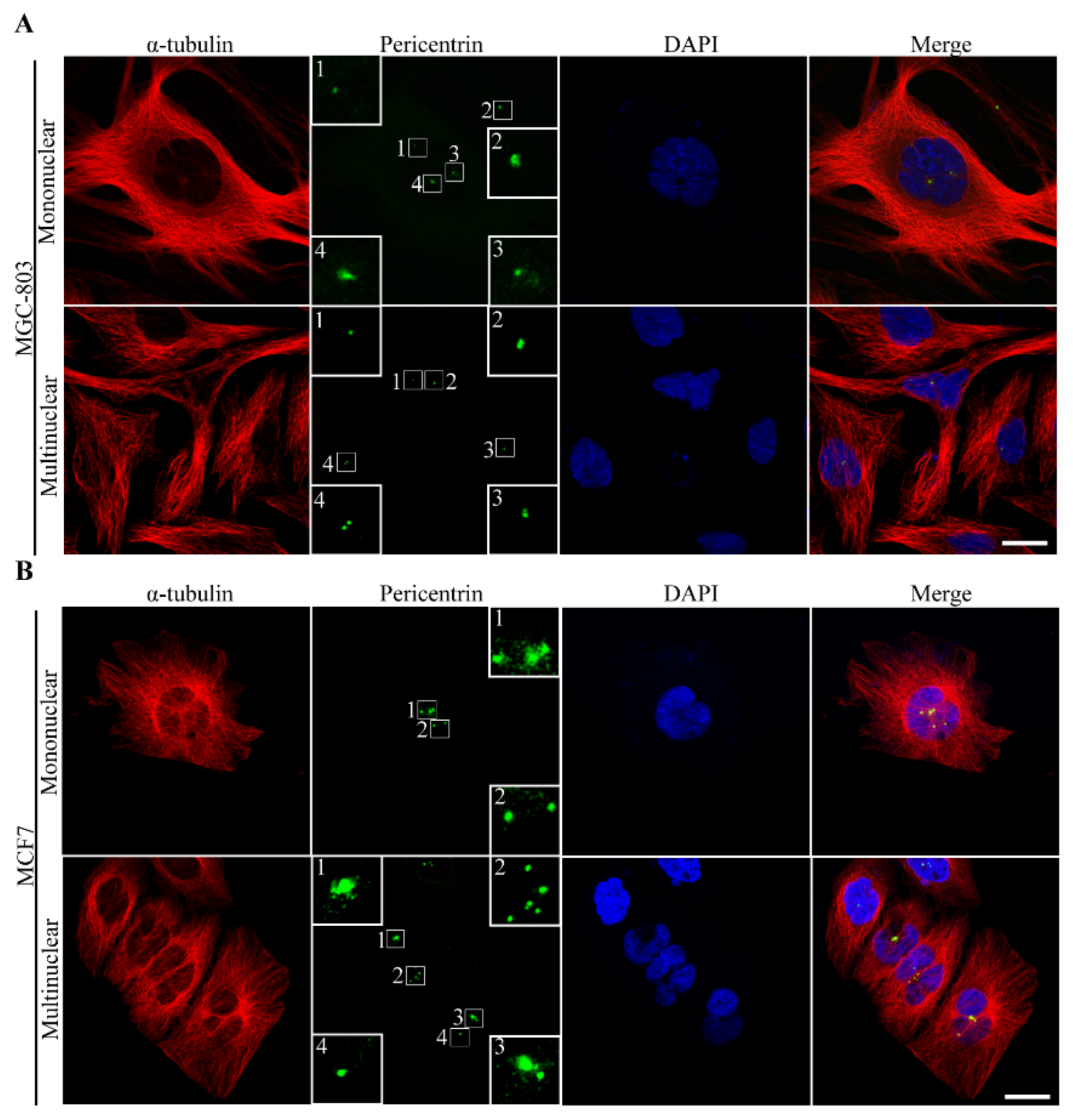
3.2.4. The Route of Centrosome Aberration
3.2.5. Incomes of Multiple Multipolar Divisions Induced by Heat Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nigg, E.A. Centrosome aberrations: cause or consequence of cancer progression? Nat. Rev. Cancer 2002, 2, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Nigg, E.A. Origins and consequences of centrosome aberrations in human cancers. Int. J. Cancer 2006, 119, 2717–2723. [Google Scholar] [CrossRef] [PubMed]
- Rajagopalan, H.; Lengauer, C. Aneuploidy and cancer. Nature 2004, 432, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Fukasawa, K. Centrosome amplification, chromosome instability and cancer development. Cancer Lett. 2005, 230, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Boveri, T. Concerning the origin of malignant tumours by Theodor Boveri. Translated and annotated by Henry Harris. J. Cell Sci. 2008, 121, 1–84. [Google Scholar] [CrossRef] [PubMed]
- Pihan, G.A.; Purohit, A.; Wallace, J.; Knecht, H.; Woda, B.; Quesenberry, P.; Doxsey, S.J. Centrosome defects and genetic instability in malignant tumors. Cancer Res. 1998, 58, 3974–3985. [Google Scholar] [PubMed]
- Yang, Z.; Lon arek, J.; Khodjakov, A.; Rieder, C.L. Extra centrosomes and/or chromosomes prolong mitosis in human cells. Nat. Cell Biol. 2008, 10, 748–751. [Google Scholar] [CrossRef] [Green Version]
- Bian, M.; Fu, J.; Yan, Y.; Chen, Q.; Yang, C.; Shi, Q.; Jiang, Q.; Zhang, C. Short exposure to paclitaxel induces multipolar spindle formation and aneuploidy through promotion of acentrosomal pole assembly. Sci. China Life Sci. 2010, 53, 1322–1329. [Google Scholar] [CrossRef]
- Timonen, S.; Therman, E. The changes in the mitotic mechanism of human cancer cells. Cancer Res. 1950, 10, 431–439. [Google Scholar]
- Rohnalter, V.; Roth, K.; Finkernagel, F.; Adhikary, T.; Obert, J.; Dorzweiler, K.; Bensberg, M.; Müller-Brüsselbach, S.; Müller, R. A multi-stage process including transient polyploidization and EMT precedes the emergence of chemoresistent ovarian carcinoma cells with a dedifferentiated and pro-inflammatory secretory phenotype. Oncotarget 2015, 6, 40005–40025. [Google Scholar] [CrossRef]
- Kittur, H.; Weaver, W.; Di Carlo, D. Well-plate mechanical confinement platform for studies of mechanical mutagenesis. Biomed. Microdevices 2014, 16, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Tse, H.T.K.; Weaver, W.M.; Di Carlo, D. Increased asymmetric and multi-daughter cell division in mechanically confined microenvironments. PLoS ONE 2012, 7, 38986–38993. [Google Scholar] [CrossRef] [PubMed]
- Parry, E.M.; Sharp, D.; Parry, J.M. The observation of mitotic division aberrations in mammalian cells exposed to chemical and radiation treatments. Mutat. Res. 1985, 150, 369–381. [Google Scholar] [CrossRef]
- Zielke-Temme, B.; Hopwood, L. Time-Lapse Cinemicrographic Observations of Heated G1-Phase Chinese Hamster Ovary Cells: II. Cell Death, Fusion, and Multipolar Divisions. Radiat. Res. 1982, 92, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Vidair, C.; Doxsey, S.; Dewey, W. Heat shock alters centrosome organization leading to mitotic dysfunction and cell death. J. Cell. Physiol. 1993, 154, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Srinivas, U.K. Heat shock induces chromosomal instability in near-tetraploid embryonal carcinoma cells. Cancer Biol. Ther. 2008, 7, 1471–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahata, K.; Miyakoda, M.; Suzuki, K.; Kodama, S.; Watanabe, M. Heat shock induces centrosomal dysfunction, and causes non-apoptotic mitotic catastrophe in human tumour cells. Int. J. Hyperthermia 2002, 18, 332–343. [Google Scholar] [CrossRef]
- Petrova, N.V.; Velichko, A.K.; Razin, S.V.; Kantidze, O.L. Early S-phase cell hypersensitivity to heat stress. Cell Cycle 2016, 15, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Asami, T.; Onozato, H. Polypolar spindle formation during first cell cycle in rainbow trout Oncorhynchus mykiss embryos after heat-shock treatment. Fish. Sci. 2007, 73, 1325–1331. [Google Scholar]
- Baek, I.K.; Jang, Y.K.; Lee, T.H.; Lee, J. Kinetic analysis of de novo centriole assembly in heat-shocked mammalian cells. Cytoskeleton 2017, 74, 18–28. [Google Scholar] [CrossRef]
- Stebbins, G.L., Jr. Types of polyploids: their classification and significance. Adv. Genet. 1947, 1, 403–429. [Google Scholar] [PubMed]
- Otto, S.P.; Whitton, J. Polyploid incidence and evolution. Annu. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [PubMed]
- Storchova, Z.; Pellman, D. From polyploidy to aneuploidy, genome instability and cancer. Nat. Rev. Mol. Cell Biol. 2004, 5, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Ganem, N.J.; Storchova, Z.; Pellman, D. Tetraploidy, aneuploidy and cancer. Curr. Opin. Genet. Dev. 2007, 17, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Storchova, Z.; Kuffer, C. The consequences of tetraploidy and aneuploidy. J. Cell Sci. 2008, 121, 3859–3866. [Google Scholar] [CrossRef] [Green Version]
- Pincus, G.; Waddington, C.H. The effects of mitosis-inhibiting treaments on normally fertilized pre-cleavage rabbit eggs: The comparative behavior of mamunalian eggs in vivo and in vitro-V. J. Hered. 1939, 30, 515–518. [Google Scholar] [CrossRef]
- Beatty, R.; Fischberg, M. Heteroploidy in mammals. III. Induction of tetraploidy in pre-implantation mouse eggs. J. Genet 1952, 50, 471–479. [Google Scholar] [CrossRef]
- Ostertag, W. The change in ploidy of human cells in culture after temperature shock. Exp. Cell Res. 1964, 34, 194–198. [Google Scholar] [CrossRef]
- Westra, A.; Dewey, W. Variation in sensitivity to heat shock during the cell-cycle of Chinese hamster cells in vitro. Int. J. Radiat. Biol. 1971, 19, 467–477. [Google Scholar] [CrossRef]
- Coss, R.A.; Dewey, W.C.; Bamburg, J.R. Effects of hyperthermia (41.5°) on Chinese hamster ovary cells analyzed in mitosis. Cancer Res. 1979, 39, 1911–1918. [Google Scholar]
- Coss, R.A.; Dewey, W.C.; Bamburg, J.R. Effects of hyperthermia on dividing Chinese hamster ovary cells and on microtubules in vitro. Cancer Res. 1982, 42, 1059–1071. [Google Scholar] [PubMed]
- Tan, Z.; Chan, Y.J.A.; Chua, Y.J.K.; Rutledge, S.D.; Pavelka, N.; Cimini, D.; Rancati, G. Environmental stresses induce karyotypic instability in colorectal cancer cells. Mol. Biol. Cell 2019, 30, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Bandi, M.; Nitta, M.; Ivanova, E.V.; Bronson, R.T.; Pellman, D. Cytokinesis failure generating tetraploids promotes tumorigenesis in p53-null cells. Nature 2005, 437, 1043–1047. [Google Scholar] [CrossRef] [PubMed]
- Andreassen, P.R.; Lohez, O.D.; Lacroix, F.B.; Margolis, R.L. Tetraploid state induces p53-dependent arrest of nontransformed mammalian cells in G1. Mol. Biol. Cell 2001, 12, 1315–1328. [Google Scholar] [CrossRef] [PubMed]
- Dittmar, T.; Nagler, C.; Schwitalla, S.; Reith, G.; Niggemann, B.; Znker, K.S. Recurrence cancer stem cells-made by cell fusion? Med.l Hypotheses 2009, 73, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Berndt, B.; Zanker, K.S.; Dittmar, T. Cell fusion is a potent inducer of aneuploidy and drug resistance in tumor cell/normal cell hybrids. Crit. Rev. Oncog. 2013, 18, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Ciocca, D.R.; Fuqua, S.A.; Lock-Lim, S.; Toft, D.O.; Welch, W.J.; McGuire, W.L. Response of human breast cancer cells to heat shock and chemotherapeutic drugs. Cancer Res. 1992, 52, 3648–3654. [Google Scholar] [PubMed]
- Lee, A.J.; Swanton, C. Tumour heterogeneity and drug resistance: personalising cancer medicine through functional genomics. Biochem. Pharmacol. 2012, 83, 1013–1020. [Google Scholar] [CrossRef]
- Vargas, H.I.; Dooley, W.C.; Gardner, R.A.; Gonzalez, K.D.; Venegas, R.; Heywang-Kobrunner, S.H.; Fenn, A.J. Focused microwave phased array thermotherapy for ablation of early-stage breast cancer: results of thermal dose escalation. Ann. Surg. Oncol. 2004, 11, 139–146. [Google Scholar] [CrossRef]
- Yang, X.-J.; Huang, C.-Q.; Suo, T.; Mei, L.-J.; Yang, G.-L.; Cheng, F.-L.; Zhou, Y.-F.; Xiong, B.; Yonemura, Y.; Li, Y. Cytoreductive surgery and hyperthermic intraperitoneal chemotherapy improves survival of patients with peritoneal carcinomatosis from gastric cancer: Final results of a phase III randomized clinical trial. Ann. Surg. Oncol. 2011, 18, 1575–1581. [Google Scholar] [CrossRef]
- Yoshida, S.; Kornek, M.; Ikenaga, N.; Schmelzle, M.; Masuzaki, R.; Csizmadia, E.; Wu, Y.; Robson, S.C.; Schuppan, D. Sublethal heat treatment promotes epithelial-mesenchymal transition and enhances the malignant potential of hepatocellular carcinoma. Hepatology 2013, 58, 1667–1680. [Google Scholar] [CrossRef] [PubMed]
- Obara, K.; Matsumoto, N.; Okamoto, M.; Kobayashi, M.; Ikeda, H.; Takahashi, H.; Katakura, Y.; Matsunaga, K.; Ishii, T.; Okuse, C. Insufficient radiofrequency ablation therapy may induce further malignant transformation of hepatocellular carcinoma. Hepatol. Int. 2008, 2, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozarowski, P.; Darzynkiewicz, Z. Analysis of cell cycle by flow cytometry. In Checkpoint Controls and Cancer; Springer: Clifton, NJ, USA, 2004; pp. 301–311. [Google Scholar]
- Chen, B.; Zhou, M.; Xu, L.X. Study of vascular endothelial cell morphology during hyperthermia. J. Therm.Biol. 2005, 30, 111–117. [Google Scholar] [CrossRef]
- Vitale, I.; Galluzzi, L.; Senovilla, L.; Criollo, A.; Jemaa, M.; Castedo, M.; Kroemer, G. Illicit survival of cancer cells during polyploidization and depolyploidization. Cell Death Differ. 2011, 18, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Ganem, N.J.; Godinho, S.A.; Pellman, D. A mechanism linking extra centrosomes to chromosomal instability. Nature 2009, 460, 278–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewénius, Y.; Gorunova, L.; Jonson, T.; Larsson, N.; Hoglund, M.; Mandahl, N.; Mertens, F.; Mitelman, F.; Gisselsson, D. Structural and numerical chromosome changes in colon cancer develop through telomere-mediated anaphase bridges, not through mitotic multipolarity. Proc. Natl. Acad. Sci. USA 2005, 102, 5541–5546. [Google Scholar] [CrossRef] [Green Version]
- Vavylonis, D.; Wu, J.Q.; Hao, S.; O’shaughnessy, B.; Pollard, T.D. Assembly mechanism of the contractile ring for cytokinesis by fission yeast. Science 2008, 319, 97–100. [Google Scholar] [CrossRef]
- Cao, L.; Wang, Y.-L. Mechanism of the formation of contractile ring in dividing cultured animal cells. II. Cortical movement of microinjected actin filaments. J. Cell Biol. 1990, 111, 1905–1911. [Google Scholar] [CrossRef]
- Murthy, K.; Wadsworth, P. Myosin-II-dependent localization and dynamics of F-actin during cytokinesis. Curr. Biol. 2005, 15, 724–731. [Google Scholar] [CrossRef]
- Glass, J.R.; DeWitt, R.G.; Cress, A.E. Rapid loss of stress fibers in Chinese hamster ovary cells after hyperthermia. Cancer Res. 1985, 45, 258–262. [Google Scholar]
- Coss, R.; Linnemans, W. The effects of hyperthermia on the cytoskeleton: a review. Int. J. Hyperthermia 1996, 12, 173–196. [Google Scholar] [CrossRef] [PubMed]
- Hurwitz, C.; Tolmach, L. Time-Lapse Cinemicrographic Studies of X-Irradiated HeLa S3 Cells: I. Cell Progression and Cell Disintegration. Biophys. J. 1969, 9, 607–633. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, H.; Hayashi, H. A nuclear thread bridging the sister cells prior to radiation-induced cell fusion. Radiat. Res. 1979, 77, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; King, R.W. Chromosome nondisjunction yields tetraploid rather than aneuploid cells in human cell lines. Nature 2005, 437, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Miller, F.R.; McInerney, D.; Rogers, C.; Miller, B.E. Spontaneous fusion between metastatic mammary tumor subpopulations. J. Cell. Biochem. 1988, 36, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, K.; Lichtenberg, J.; Thomsen, P.; Larsson, L.I. Spontaneous fusion between cancer cells and endothelial cells. Cell. Mol. Life Sci. 2004, 61, 2125–2131. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Kang, Y. Cell fusion as a hidden force in tumor progression. Cancer Res. 2009, 69, 8536–8539. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Kang, Y. Efficient acquisition of dual metastasis organotropism to bone and lung through stable spontaneous fusion between MDA-MB-231 variants. Proc. Natl. Acad. Sci. USA 2009, 106, 9385–9390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duelli, D.; Lazebnik, Y. Cell fusion: a hidden enemy? Cancer Cell 2003, 3, 445–448. [Google Scholar] [CrossRef]
- Dittmar, T.; Schwitalla, S.; Seidel, J.; Haverkampf, S.; Reith, G.; Meyer-Staeckling, S.N.; Brandt, B.H.; Niggemann, B.; Zanker, K.S. Characterization of hybrid cells derived from spontaneous fusion events between breast epithelial cells exhibiting stem-like characteristics and breast cancer cells. Clin. Exp. Metastasis 2011, 28, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Maier, B.; Bartek, J. Centrosome clustering and chromosomal (in) stability: a matter of life and death. Mol. Oncol. 2011, 5, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Godinho, S.A.; Chandhok, N.S.; Ganem, N.J.; Azioune, A.; Thery, M.; Pellman, D. Mechanisms to suppress multipolar divisions in cancer cells with extra centrosomes. Genes Dev. 2008, 22, 2189–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintyne, N.J.; Reing, J.E.; Hoffelder, D.R.; Gollin, S.M.; Saunders, W.S. Spindle multipolarity is prevented by centrosomal clustering. Science 2005, 307, 127–129. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Liu, M.; Huang, H.; Li, B.; Zhao, H.; Feng, X.-Q.; Zhao, H.-P. Heat Stress-Induced Multiple Multipolar Divisions of Human Cancer Cells. Cells 2019, 8, 888. https://doi.org/10.3390/cells8080888
Chen S, Liu M, Huang H, Li B, Zhao H, Feng X-Q, Zhao H-P. Heat Stress-Induced Multiple Multipolar Divisions of Human Cancer Cells. Cells. 2019; 8(8):888. https://doi.org/10.3390/cells8080888
Chicago/Turabian StyleChen, Shaoyong, Mingyue Liu, Huiming Huang, Bo Li, Hucheng Zhao, Xi-Qiao Feng, and Hong-Ping Zhao. 2019. "Heat Stress-Induced Multiple Multipolar Divisions of Human Cancer Cells" Cells 8, no. 8: 888. https://doi.org/10.3390/cells8080888
APA StyleChen, S., Liu, M., Huang, H., Li, B., Zhao, H., Feng, X.-Q., & Zhao, H.-P. (2019). Heat Stress-Induced Multiple Multipolar Divisions of Human Cancer Cells. Cells, 8(8), 888. https://doi.org/10.3390/cells8080888