The IL4I1 Enzyme: A New Player in the Immunosuppressive Tumor Microenvironment

Abstract
1. Introduction
2. L-Amino-Acid Oxidase Interleukin-Four Induced Gene 1 (IL4I1)
3. Expression of IL4I1
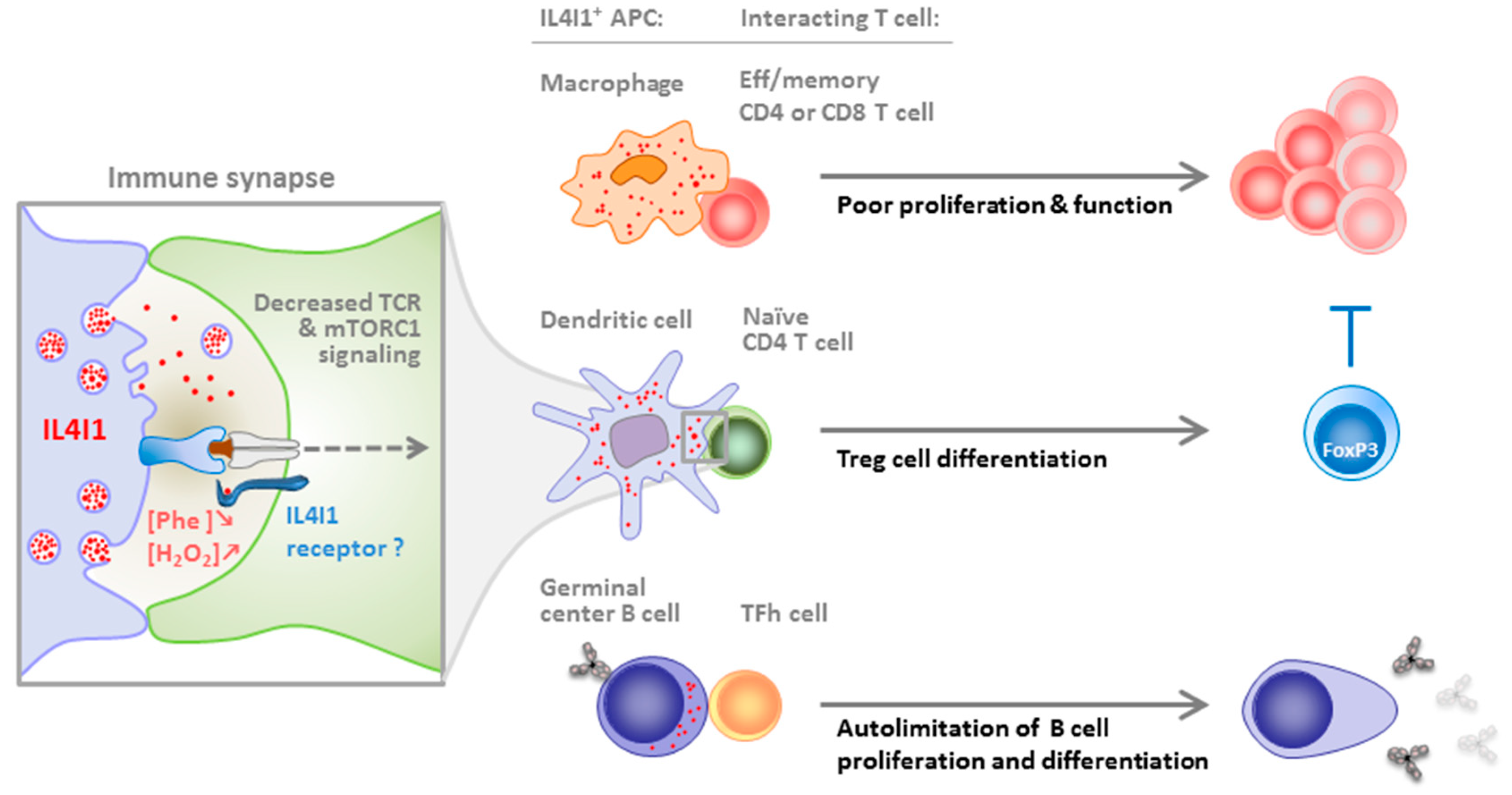
4. IL4I1 Inhibition of T and B Cells
5. The Role of IL4I1 in Infection and the Control of Immunopathology
6. The Role of IL4I1 in the Immune Escape of Tumors
7. Conclusions
Funding
Acknowledgments
References
- Matzinger, P. Tolerance, danger, and the extended family. Annu Rev. Immunol 1994, 12, 991–1045. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.C.; Paul, W.E. Fig1, an interleukin 4-induced mouse B cell gene isolated by cDNA representational difference analysis. Proc. Natl. Acad. Sci. USA 1997, 94, 2507–2512. [Google Scholar] [CrossRef] [PubMed]
- Chavan, S.S.; Tian, W.; Hsueh, K.; Jawaheer, D.; Gregersen, P.K.; Chu, C.C. Characterization of the human homolog of the IL-4 induced gene-1 (Fig1). Biochim. Biophys. Acta 2002, 1576, 70–80. [Google Scholar] [CrossRef]
- Houston, B.; Curry, B.; Aitken, R.J. Human spermatozoa possess an IL4I1 l-amino acid oxidase with a potential role in sperm function. Reproduction 2015, 149, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, S.; Kolb-Kokocinski, A.; Poustka, A. Alternative pre-mRNA processing regulates cell-type specific expression of the IL4l1 and NUP62 genes. BMC Biol. 2005, 3, 16. [Google Scholar] [CrossRef]
- Boulland, M.L.; Marquet, J.; Molinier-Frenkel, V.; Moller, P.; Guiter, C.; Lasoudris, F.; Copie-Bergman, C.; Baia, M.; Gaulard, P.; Leroy, K.; et al. Human IL4I1 is a secreted L-phenylalanine oxidase expressed by mature dendritic cells that inhibits T-lymphocyte proliferation. Blood 2007, 110, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Molinier-Frenkel, V.; Mestivier, D.; Castellano, F. Alterations of the immunosuppressive IL4I1 enzyme activity induced by naturally occurring SNP/mutations. Genes Immun. 2016, 17, 148–152. [Google Scholar] [CrossRef]
- Yue, Y.; Huang, W.; Liang, J.; Guo, J.; Ji, J.; Yao, Y.; Zheng, M.; Cai, Z.; Lu, L.; Wang, J. IL4I1 Is a Novel Regulator of M2 Macrophage Polarization That Can Inhibit T Cell Activation via L-Tryptophan and Arginine Depletion and IL-10 Production. PLoS ONE 2015, 10, e0142979. [Google Scholar] [CrossRef]
- De Kok, A.; Veeger, C. Studies on L-amino-acid oxidase. I. Effects of pH and competitive inhibitors. Biochim. Biophys. Acta 1968, 167, 35–47. [Google Scholar] [CrossRef]
- Marquet, J.; Lasoudris, F.; Cousin, C.; Puiffe, M.; Martin-Garcia, N.; Baud, V.; Chéreau, F.; Farcet, J.; Molinier-Frenkel, V.; Castellano, F. Dichotomy between factors inducing the immunosuppressive enzyme IL4I1 in B lymphocytes and mononuclear phagocytes. Eur. J. Immunol. 2010, 40, 2557–2568. [Google Scholar] [CrossRef]
- Park, S.H.; Kang, K.; Giannopoulou, E.; Qiao, Y.; Kang, K.; Kim, G.; Park-Min, K.H.; Ivashkiv, L.B. Type I interferons and the cytokine TNF cooperatively reprogram the macrophage epigenome to promote inflammatory activation. Nat. Immunol. 2017, 18, 1104–1116. [Google Scholar] [CrossRef]
- Carbonnelle-Puscian, A.; Copie-Bergman, C.; Baia, M.; Martin-Garcia, N.; Allory, Y.; Haioun, C.; Cremades, A.; Abd-Alsamad, I.; Farcet, J.P.; Gaulard, P.; et al. The novel immunosuppressive enzyme IL4I1 is expressed by neoplastic cells of several B-cell lymphomas and by tumor-associated macrophages. Leukemia 2009, 23, 952–960. [Google Scholar] [CrossRef]
- Caron, G.; Le Gallou, S.; Lamy, T.; Tarte, K.; Fest, T. CXCR4 expression functionally discriminates centroblasts versus centrocytes within human germinal center B cells. J. Immunol. 2009, 182, 7595–7602. [Google Scholar] [CrossRef]
- Victora, G.D.; Schwickert, T.A.; Fooksman, D.R.; Kamphorst, A.O.; Meyer-Hermann, M.; Dustin, M.L.; Nussenzweig, M.C. Germinal center dynamics revealed by multiphoton microscopy with a photoactivatable fluorescent reporter. Cell 2010, 143, 592–605. [Google Scholar] [CrossRef]
- Santarlasci, V.; Maggi, L.; Mazzoni, A.; Capone, M.; Querci, V.; Rossi, M.C.; Beltrame, L.; Cavalieri, D.; De Palma, R.; Liotta, F.; et al. IL-4-induced gene 1 maintains high Tob1 expression that contributes to TCR unresponsiveness in human T helper 17 cells. Eur. J. Immunol. 2014, 44, 654–661. [Google Scholar] [CrossRef]
- Santarlasci, V.; Maggi, L.; Capone, M.; Querci, V.; Beltrame, L.; Cavalieri, D.; D’Aiuto, E.; Cimaz, R.; Nebbioso, A.; Liotta, F.; et al. Rarity of human T helper 17 cells is due to retinoic acid orphan receptor-dependent mechanisms that limit their expansion. Immunity 2012, 36, 201–214. [Google Scholar] [CrossRef]
- Scarlata, C.M.; Celse, C.; Pignon, P.; Ayyoub, M.; Valmori, D. Differential expression of the immunosuppressive enzyme IL4I1 induce Aiolos, but not natural Helios, FOXP3 Treg cells. Eur. J. Immunol. 2014. [Google Scholar] [CrossRef]
- Bulitta, B.; Zuschratter, W.; Bernal, I.; Bruder, D.; Klawonn, F.; von Bergen, M.; Garritsen, H.S.P.; Jansch, L. Proteomic definition of human mucosal-associated invariant T cells determines their unique molecular effector phenotype. Eur. J. Immunol. 2018, 48, 1336–1349. [Google Scholar] [CrossRef]
- Cousin, C.; Aubatin, A.; Le Gouvello, S.; Apetoh, L.; Castellano, F.; Molinier-Frenkel, V. The immunosuppressive enzyme IL4I1 promotes FoxP3(+) regulatory T lymphocyte differentiation. Eur. J. Immunol. 2015, 45, 1772–1782. [Google Scholar] [CrossRef]
- Aubatin, A.; Sako, N.; Decrouy, X.; Donnadieu, E.; Molinier-Frenkel, V.; Castellano, F. IL4-induced gene 1 is secreted at the immune synapse and modulates TCR activation independently of its enzymatic activity. Eur. J. Immunol. 2017. [Google Scholar] [CrossRef]
- Aalto, K.; Autio, A.; Kiss, E.A.; Elima, K.; Nymalm, Y.; Veres, T.Z.; Marttila-Ichihara, F.; Elovaara, H.; Saanijoki, T.; Crocker, P.R.; et al. Siglec-9 is a novel leukocyte ligand for vascular adhesion protein-1 and can be used in PET imaging of inflammation and cancer. Blood 2011, 118, 3725–3733. [Google Scholar] [CrossRef]
- Pallotta, M.T.; Orabona, C.; Volpi, C.; Vacca, C.; Belladonna, M.L.; Bianchi, R.; Servillo, G.; Brunacci, C.; Calvitti, M.; Bicciato, S.; et al. Indoleamine 2,3-dioxygenase is a signaling protein in long-term tolerance by dendritic cells. Nat. Immunol. 2011, 12, 870–878. [Google Scholar] [CrossRef]
- Bod, L.; Douguet, L.; Auffray, C.; Lengagne, R.; Bekkat, F.; Rondeau, E.; Molinier-Frenkel, V.; Castellano, F.; Richard, Y.; Prevost-Blondel, A. IL-4-Induced Gene 1: A Negative Immune Checkpoint Controlling B Cell Differentiation and Activation. J. Immunol. 2018, 200, 1027–1038. [Google Scholar] [CrossRef]
- Romagnani, S. IL4I1: Key immunoregulator at a crossroads of divergent T-cell functions. Eur. J. Immunol. 2016, 46, 2302–2305. [Google Scholar] [CrossRef]
- Puiffe, M.L.; Lachaise, I.; Molinier-Frenkel, V.; Castellano, F. Antibacterial properties of the mammalian L-amino acid oxidase IL4I1. PLoS ONE 2013, 8, e54589. [Google Scholar] [CrossRef]
- Seddigh, P.; Bracht, T.; Molinier-Frenkel, V.; Castellano, F.; Kniemeyer, O.; Schuster, M.; Weski, J.; Hasenberg, A.; Kraus, A.; Poschet, G.; et al. Quantitative Analysis of Proteome Modulations in Alveolar Epithelial Type II Cells in Response to Pulmonary Aspergillus fumigatus Infection. Mol. Cell. Proteom. 2017, 16, 2184–2198. [Google Scholar] [CrossRef]
- Reales-Calderon, J.A.; Vaz, C.; Monteoliva, L.; Molero, G.; Gil, C. Candida albicans Modifies the Protein Composition and Size Distribution of THP-1 Macrophage-Derived Extracellular Vesicles. J. Proteom. Res. 2017, 16, 87–105. [Google Scholar] [CrossRef]
- Baas, T.; Baskin, C.R.; Diamond, D.L.; Garcia-Sastre, A.; Bielefeldt-Ohmann, H.; Tumpey, T.M.; Thomas, M.J.; Carter, V.S.; Teal, T.H.; Van Hoeven, N.; et al. Integrated molecular signature of disease: analysis of influenza virus-infected macaques through functional genomics and proteomics. J. Virol. 2006, 80, 10813–10828. [Google Scholar] [CrossRef]
- Anyanwu, S.I.; Doherty, A.; Powell, M.D.; Obialo, C.; Huang, M.B.; Quarshie, A.; Mitchell, C.; Bashir, K.; Newman, G.W. Detection of HIV-1 and Human Proteins in Urinary Extracellular Vesicles from HIV+ Patients. Adv. Virol. 2018, 2018, 7863412. [Google Scholar] [CrossRef]
- Feng, M.; Xie, T.; Li, Y.; Zhang, N.; Lu, Q.; Zhou, Y.; Shi, M.; Sun, J.; Zhang, X. A balanced game: chicken macrophage response to ALV-J infection. Vet. Res. 2019, 50, 20. [Google Scholar] [CrossRef]
- Liu, J.; Karypis, G.; Hippen, K.L.; Vegoe, A.L.; Ruiz, P.; Gilkeson, G.S.; Behrens, T.W. Genomic view of systemic autoimmunity in MRLlpr mice. Genes Immun. 2006, 7, 156–168. [Google Scholar] [CrossRef]
- Psachoulia, K.; Chamberlain, K.A.; Heo, D.; Davis, S.E.; Paskus, J.D.; Nanescu, S.E.; Dupree, J.L.; Wynn, T.A.; Huang, J.K. IL4I1 augments CNS remyelination and axonal protection by modulating T cell driven inflammation. Brain 2016. [Google Scholar] [CrossRef]
- Copie-Bergman, C.; Boulland, M.L.; Dehoulle, C.; Moller, P.; Farcet, J.P.; Dyer, M.J.; Haioun, C.; Romeo, P.H.; Gaulard, P.; Leroy, K. Interleukin 4-induced gene 1 is activated in primary mediastinal large B-cell lymphoma. Blood 2003, 101, 2756–2761. [Google Scholar] [CrossRef]
- Lasoudris, F.; Cousin, C.; Prevost-Blondel, A.; Martin-Garcia, N.; Abd-Alsamad, I.; Ortonne, N.; Farcet, J.P.; Castellano, F.; Molinier-Frenkel, V. IL4I1: an inhibitor of the CD8(+) antitumor T-cell response in vivo. Eur. J. Immunol. 2011, 41, 1629–1638. [Google Scholar] [CrossRef]
- Bod, L.; Lengagne, R.; Wrobel, L.; Ramspott, J.P.; Kato, M.; Avril, M.F.; Castellano, F.; Molinier-Frenkel, V.; Prevost-Blondel, A. IL4-induced gene 1 promotes tumor growth by shaping the immune microenvironment in melanoma. Oncoimmunology 2017, 6, e1278331. [Google Scholar] [CrossRef]
- Prevost-Blondel, A.; Richard, Y. Interleukin 4–Induced Gene 1 as an Emerging Regulator of B-Cell Biology and its Role in Cutaneous Melanoma. Crit. Rev. Immunol. 2019, 39, 39–57. [Google Scholar] [CrossRef]
- Ramspott, J.P.; Bekkat, F.; Bod, L.; Favier, M.; Terris, B.; Salomon, A.; Djerroudi, L.; Zaenker, K.S.; Richard, Y.; Molinier-Frenkel, V.; et al. Emerging role of IL4-induced gene 1 as a prognostic biomarker affecting the local T cell response in human cutaneous melanoma. J. Invest. Dermatol. 2018. [Google Scholar] [CrossRef]
- Finak, G.; Bertos, N.; Pepin, F.; Sadekova, S.; Souleimanova, M.; Zhao, H.; Chen, H.; Omeroglu, G.; Meterissian, S.; Omeroglu, A.; et al. Stromal gene expression predicts clinical outcome in breast cancer. Nat. Med. 2008, 14, 518–527. [Google Scholar] [CrossRef]
- Regev, A.; Teichmann, S.A.; Lander, E.S.; Amit, I.; Benoist, C.; Birney, E.; Bodenmiller, B.; Campbell, P.; Carninci, P.; Clatworthy, M.; et al. The Human Cell Atlas. eLife 2017, 6. [Google Scholar] [CrossRef]
- Poplawski, P.; Tohge, T.; Boguslawska, J.; Rybicka, B.; Tanski, Z.; Trevino, V.; Fernie, A.R.; Piekielko-Witkowska, A. Integrated transcriptomic and metabolomic analysis shows that disturbances in metabolism of tumor cells contribute to poor survival of RCC patients. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 744–752. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Enzyme Acronym | Inducible Nitric Oxide Synthase (iNOS) | Arginase (Arg1 and Arg2) | Indoleamine 2,3-Dioxygenase (IDO1 and IDO2) | Tryptophan 2,3 Dioxygenase (TDO) | Interleukin 4-Induced Gene 1 (IL4I1) |
|---|---|---|---|---|---|
| Substrate and reaction | Arginine→ Citrulline + NO | Arginine→ Ornithine + Urea | Tryptophan +O2→ Kynurenines | Tryptophan +O2→ Kynurenines | Phe→ H2O2 + NH3 + Phenylpyruvate |
| Expression in the immune system | Myeloid cells | Myeloid cells | Myeloid cells | Myeloid cells | Myeloid cells, T and B lymphocytes |
| Control by soluble factors | IFNγ | IL-4 and IL-10 in mouse macrophages | IFNγ | Corticosteroids and glucagon | Type I and II IFN in myeloid cells (humans) and IL-4 in B cells (humans and mice) |
| Localization | Intracellular | Intracellular | Intracellular | Intracellular | Secreted |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molinier-Frenkel, V.; Prévost-Blondel, A.; Castellano, F. The IL4I1 Enzyme: A New Player in the Immunosuppressive Tumor Microenvironment. Cells 2019, 8, 757. https://doi.org/10.3390/cells8070757
Molinier-Frenkel V, Prévost-Blondel A, Castellano F. The IL4I1 Enzyme: A New Player in the Immunosuppressive Tumor Microenvironment. Cells. 2019; 8(7):757. https://doi.org/10.3390/cells8070757
Chicago/Turabian StyleMolinier-Frenkel, Valérie, Armelle Prévost-Blondel, and Flavia Castellano. 2019. "The IL4I1 Enzyme: A New Player in the Immunosuppressive Tumor Microenvironment" Cells 8, no. 7: 757. https://doi.org/10.3390/cells8070757
APA StyleMolinier-Frenkel, V., Prévost-Blondel, A., & Castellano, F. (2019). The IL4I1 Enzyme: A New Player in the Immunosuppressive Tumor Microenvironment. Cells, 8(7), 757. https://doi.org/10.3390/cells8070757