The Zinc Transporter Zip7 Is Downregulated in Skeletal Muscle of Insulin-Resistant Cells and in Mice Fed a High-Fat Diet

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Protein Extraction
2.3. RNA Extraction
2.4. cDNA Synthesis
2.5. Mice and Diets
2.6. Body Composition Analysis
2.7. Oral Glucose Tolerance Test
2.8. Mouse Tissue Collection and Protein Extraction
2.9. Fatty Acid Preparation and Cell Culture Treatment
2.10. Cell Viability Assay [1-(4,5-Dimethylthiazol-2-yl)-3,5-diphenylformazan] Assay
2.11. Insulin Receptor Inhibition
2.12. Western Blotting
2.13. Zip7 Overexpression
2.14. Quantitative Real-Time PCR (qPCR)
2.15. Insulin Signaling Pathway Array
2.16. Data Analysis
3. Results
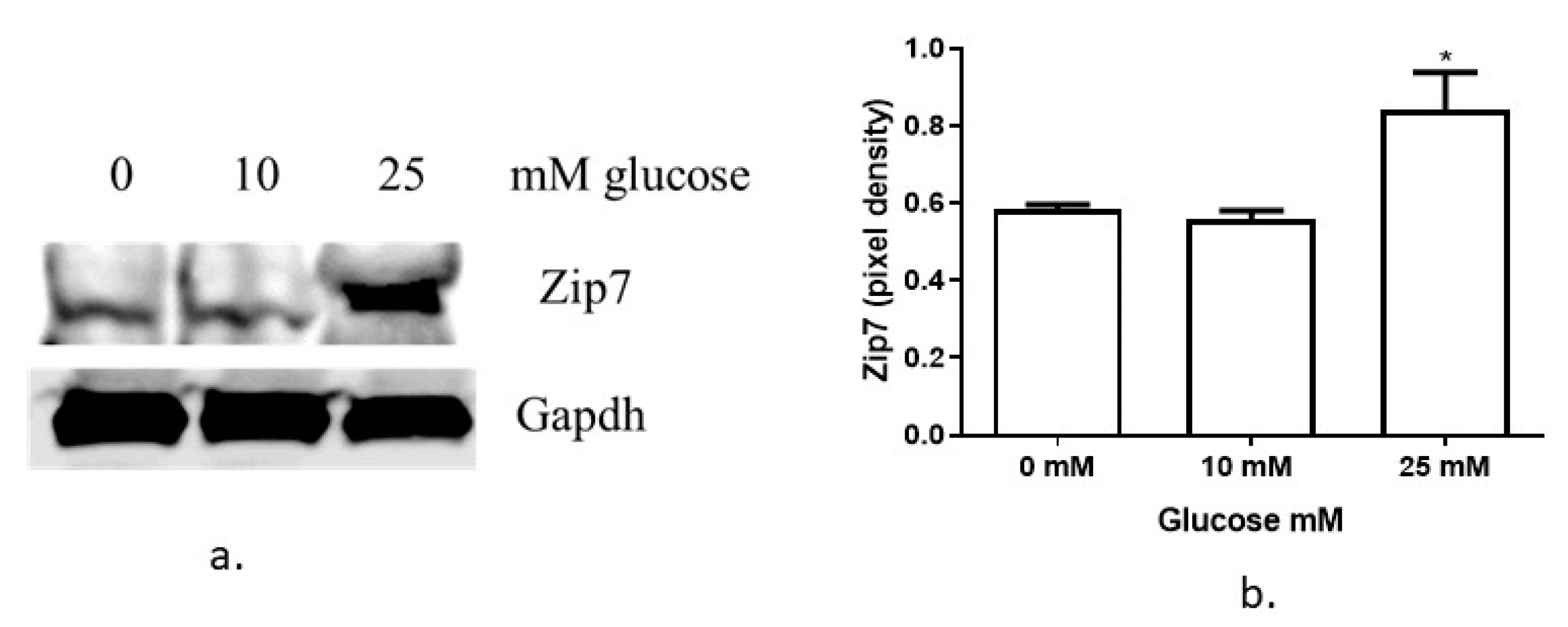
3.1. Glucose Regulates Zip7 Expression in C2C12 Skeletal Muscle Cells
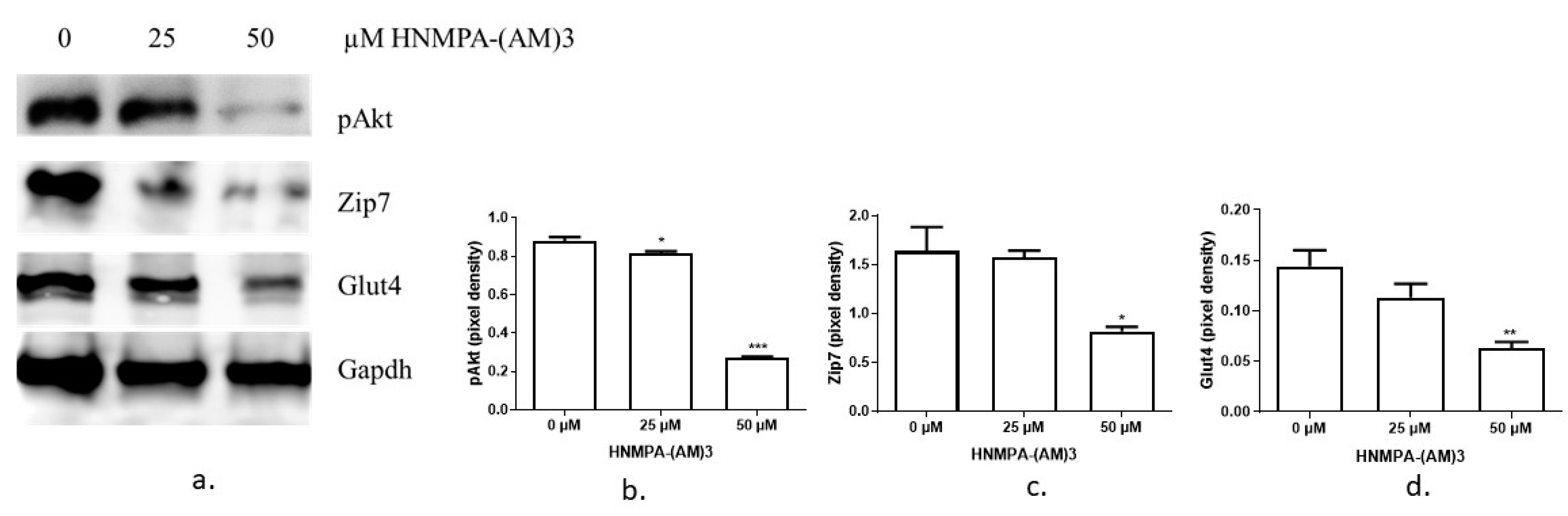
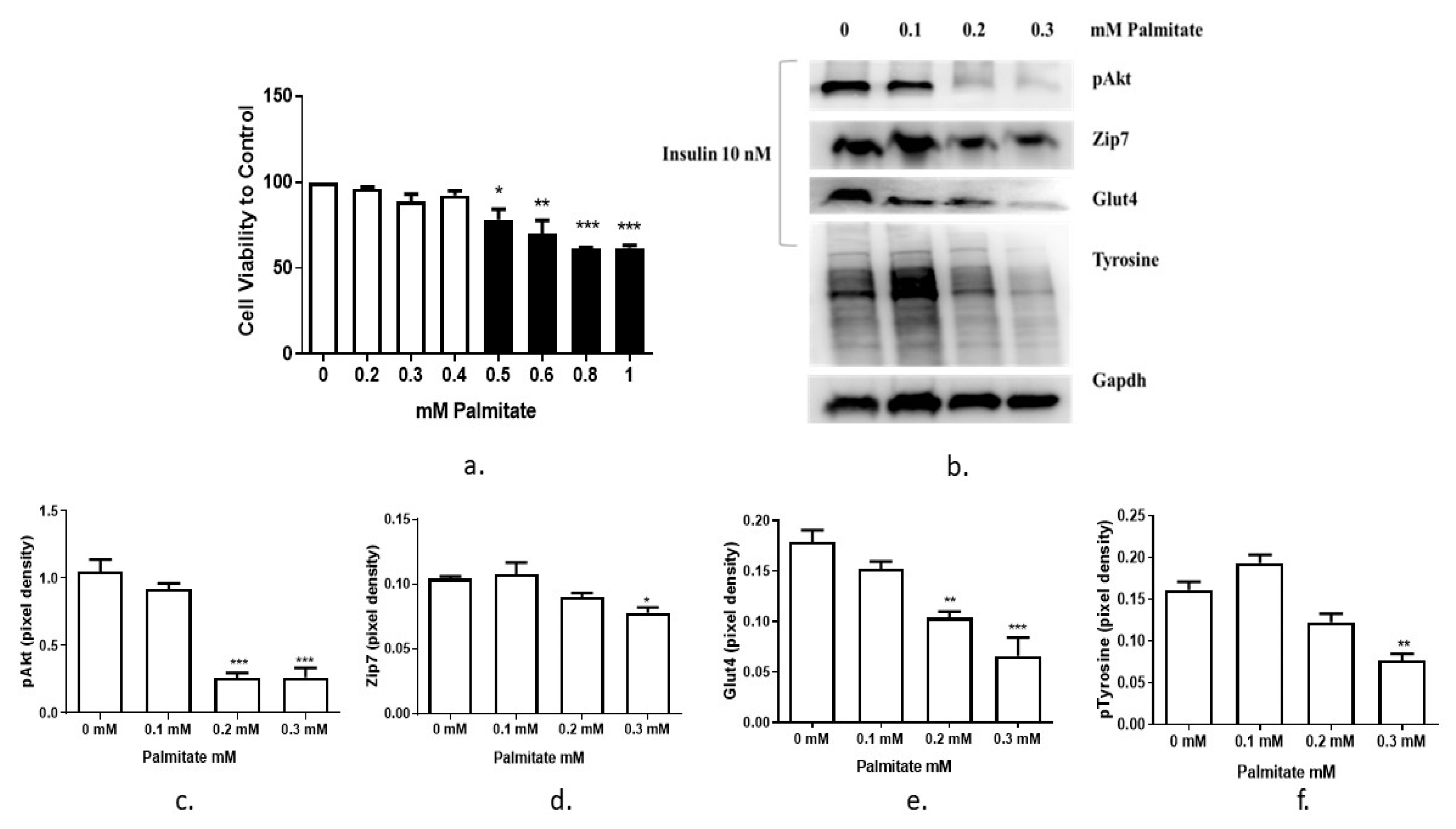
3.2. The Expression of Zip7 and Glut4 Is Suppressed in Insulin-Resistant C2C12 Cells Treated with Either an Insulin Receptor Inhibitor Hnmpa-(Am)3 or Palmitate
3.3. Testing the Function of ZIP7 in Controlling Genes Involved in Insulin Signaling
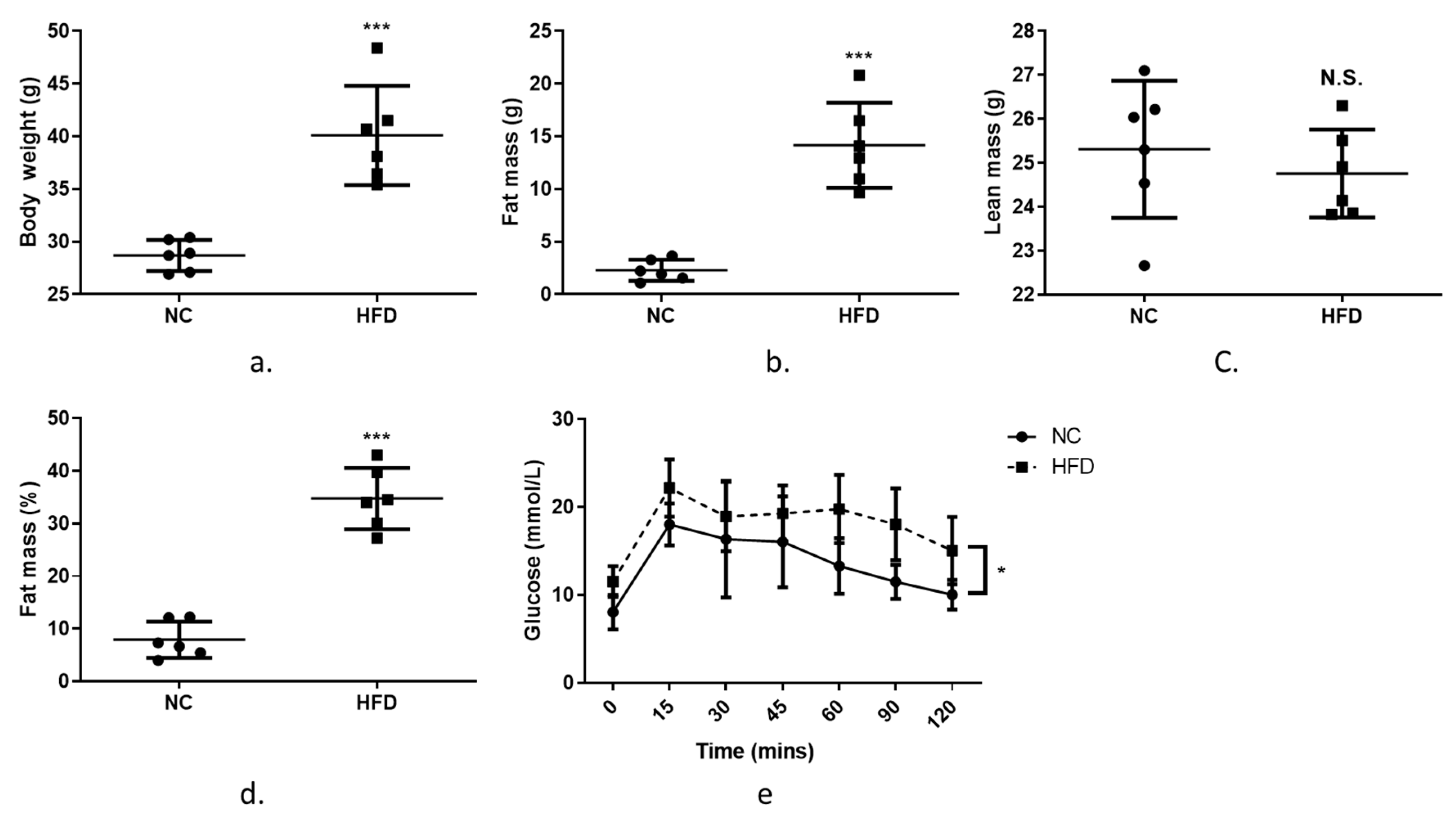
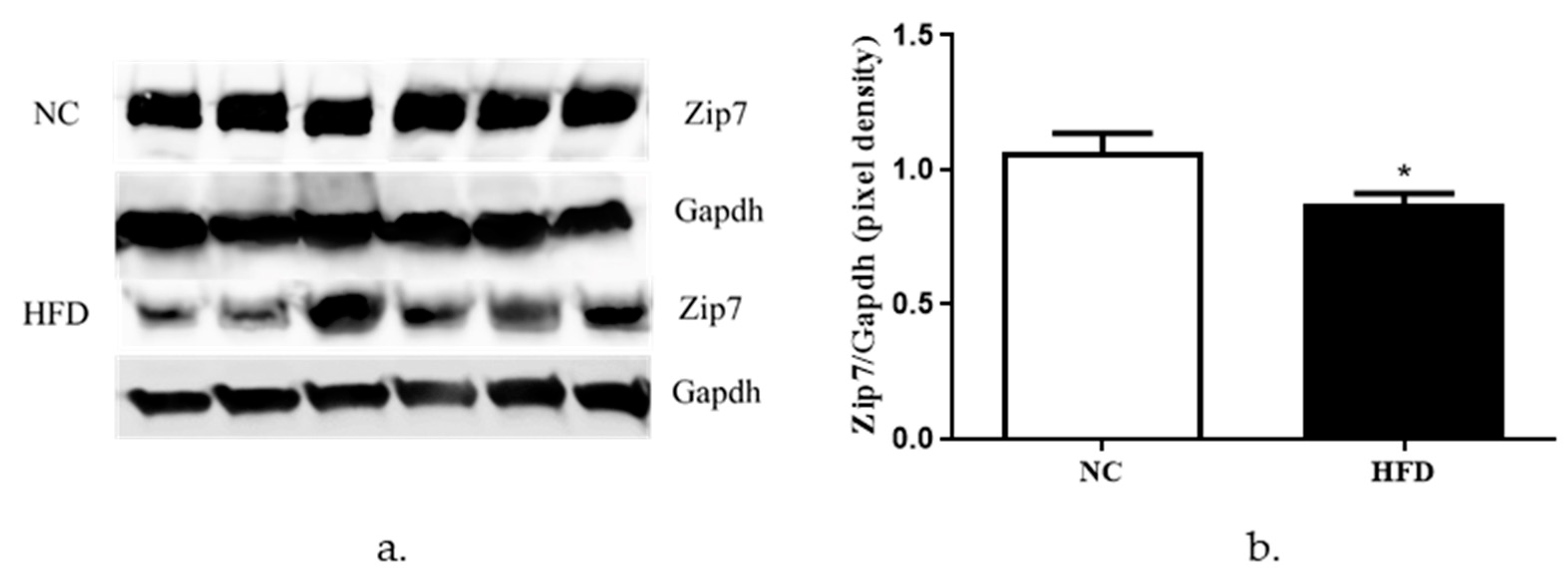
3.4. The Expression of Zip7 and Glut4 Is Reduced in the Skeletal Muscle of Mice Fed a High-Fat Diet
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Wild, S.; Roglic, G.; Green, A.; Sicree, R.; King, H. Global Prevalence of Diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care 2004, 27, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Hajer, G.R.; van Haeften, T.W.; Visseren, F.L.J. Adipose tissue dysfunction in obesity, diabetes, and vascular diseases. Eur. Heart J. 2008, 29, 2959–2971. [Google Scholar] [CrossRef] [PubMed]
- Marcadenti, A.; Fuchs, S.C.; Moreira, L.B.; Wiehe, M.; Gus, M.; Fuchs, F.D. Accuracy of Anthropometric Indexes of Obesity to Predict Diabetes Mellitus Type 2 among Men and Women with Hypertension. Am. J. Hypertens. 2011, 24, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Berumen, J.; Orozco, L.; Betancourt-Cravioto, M.; Gallardo, H.; Zulueta, M.; Mendizabal, L.; Simon, L.; Benuto, R.E.; Ramírez-Campos, E.; Marin, M. Influence of obesity, parental history of diabetes, and genes in type 2 diabetes: A case-control study. Sci. Rep. 2019, 9, 2748. [Google Scholar] [CrossRef] [PubMed]
- Janikiewicz, J.; Hanzelka, K.; Kozinski, K.; Kolczynska, K.; Dobrzyn, A. Islet β-cell failure in type 2 diabetes—Within the network of toxic lipids. Biochem. Biophys. Res. Commun. 2015, 460, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.A.; Nield, A.; Myers, M. Zinc Transporters, Mecahnisms of Action and Therapeutic Utility: Implications for Type 2 Diabetes Mellitus. J. Nutr. Metab. 2012. [Google Scholar] [CrossRef]
- Kambe, T.; Hashimoto, A.; Fujimoto, S. Current understanding of ZIP and ZnT zinc transporters in human health and diseases. Cell. Mol. Life Sci. 2014, 71, 3281–3295. [Google Scholar] [CrossRef]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The physiological, biochemical, and molecular roles of zinc transporters in zinc homeostasis and metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef]
- Cousins, R.J.; Liuzzi, J.P.; Lichten, L.A. Mammalian zinc transport, trafficking, and signals. JBC 2006, 281, 24085–24089. [Google Scholar] [CrossRef]
- Taylor, K.M.; Hiscox, S.; Nicholson, R.I.; Hogstrand, C.; Kille, P. Protein kinase CK2 triggers cytosolic zinc signaling pathways by phosphorylation of zinc channel ZIP7. Sci. Signal. 2012, 5, ra11. [Google Scholar] [CrossRef] [PubMed]
- Hogstrand, C.; Kille, P.; Nicholson, R.I.; Taylor, K.M. Zinc transporters and cancer: A potential role for ZIP7 as a hub for tyrosine kinase activation. Trends Mol. Med. 2009, 15, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, G.; Bouwkamp, C.G.; de Vrij, F.M.; Lovenberg, T.; Bonaventure, P.; Kushner, S.A.; Harrington, A.W. The Zinc Transporter SLC39A7 (ZIP7) Is Essential for Regulation of Cytosolic Zinc Levels. Mol. Pharm. 2018, 94, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Vichova, P.; Jordan, N.; Hiscox, S.; Hendley, R.; Nicholson, R.I. ZIP7-Mediated Intracellular Zinc Transport Contributes to Aberrant Growth Factor Signaling in Antihormone-Resistant Breast Cancer Cells. Endocrinology 2008, 149, 4912–4920. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.A.; Nield, A.; Chew, G.S.; Myers, M.A. The zinc transporter, Slc39a7 (Zip7) is implicated in glycaemic control in skeletal muscle cells. PLoS ONE 2013, 8, e79316. [Google Scholar] [CrossRef]
- Tuncay, E.; Bitirim, V.C.; Durak, A.; Carrat, G.R.J.; Taylor, K.M.; Rutter, G.A.; Turan, B. Hyperglycemia-Induced Changes in ZIP7 and ZnT7 Expression Cause Zn(2+) Release From the Sarco(endo)plasmic Reticulum and Mediate ER Stress in the Heart. Diabetes 2017, 66, 1346–1358. [Google Scholar] [CrossRef]
- Norouzi, S.; Adulcikas, J.; Sohal, S.S.; Myers, S. Zinc stimulates glucose oxidation and glycemic control by modulating the insulin signaling pathway in human and mouse skeletal muscle cell lines. PLoS ONE 2018, 13, e0191727. [Google Scholar] [CrossRef]
- Marshall, J.P.S.; Estevez, E.; Kammoun, H.L.; King, E.J.; Bruce, C.R.; Drew, B.G.; Qian, H.; Iliades, P.; Gregorevic, P.; Febbraio, M.A.; et al. Skeletal muscle-specific overexpression of heat shock protein 72 improves skeletal muscle insulin-stimulated glucose uptake but does not alter whole body metabolism. Diabetes Obes. Metab. 2018, 20, 1928–1936. [Google Scholar] [CrossRef]
- Yang, M.; Wei, D.; Mo, C.; Zhang, J.; Wang, X.; Han, X.; Wang, Z.; Xiao, H. Saturated fatty acid palmitate-induced insulin resistance is accompanied with myotube loss and the impaired expression of health benefit myokine genes in C2C12 myotubes. Lipids Health Dis. 2013, 12, 104. [Google Scholar] [CrossRef]
- Bellomo, E.A.; Meur, G.; Rutter, G.A. Glucose Regulates Free Cytosolic Zn2+ Concentration, Slc39 (ZiP), and Metallothionein Gene Expression in Primary Pancreatic Islet β-Cells. J. Biol. Chem. 2011, 286, 25778–25789. [Google Scholar] [CrossRef]
- Feng, X.T.; Wang, T.Z.; Leng, J.; Chen, Y.; Liu, J.B.; Liu, Y.; Wang, W.J. Palmitate contributes to insulin resistance through downregulation of the Src-mediated phosphorylation of Akt in C2C12 myotubes. Biosci. Biotechnol. Biochem. 2012, 76, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Takamura, T.; Matsuzawa-Nagata, N.; Takayama, H.; Misu, H.; Noda, H.; Nabemoto, S.; Kurita, S.; Ota, T.; Ando, H.; et al. Palmitate induces insulin resistance in H4IIEC3 hepatocytes through reactive oxygen species produced by mitochondria. J. Biol. Chem. 2009, 284, 14809–14818. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R. Insulin Resistance and Type 2 Diabetes. Diabetes 2012, 61, 778–779. [Google Scholar] [CrossRef] [PubMed]
- Bevan, P. Insulin signalling. J. Cell Sci. 2001, 114, 1429–1430. [Google Scholar] [PubMed]
- Tokarz, V.L.; MacDonald, P.E.; Klip, A. The cell biology of systemic insulin function. J. Cell Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lu, H.; Yang, H.; Li, C.; Sang, Q.; Liu, X.; Liu, Y.; Wang, Y.; Sun, Z. Zinc stimulates glucose consumption by modulating the insulin signaling pathway in L6 myotubes: Essential roles of Akt–GLUT4, GSK3β and mTOR–S6K1. J. Nutr. Biochem. 2016, 34, 126–135. [Google Scholar] [CrossRef]
- Hardy, A.B.; Wijesekara, N.; Genkin, I.; Prentice, K.J.; Bhattacharjee, A.; Kong, D.; Chimienti, F.; Wheeler, M.B. Effects of high-fat diet feeding on Znt8-null mice: Differences between beta-cell and global knockout of Znt8. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1084–E1096. [Google Scholar] [CrossRef]
- Fukunaka, A.; Fukada, T.; Bhin, J.; Suzuki, L.; Tsuzuki, T.; Takamine, Y.; Bin, B.-H.; Yoshihara, T.; Ichinoseki-Sekine, N.; Naito, H.; et al. Zinc transporter ZIP13 suppresses beige adipocyte biogenesis and energy expenditure by regulating C/EBP-β expression. PLoS Genet. 2017, 13, e1006950. [Google Scholar] [CrossRef]
- Liu, Y.; Batchuluun, B.; Ho, L.; Zhu, D.; Prentice, K.J.; Bhattacharjee, A.; Zhang, M.; Pourasgari, F.; Hardy, A.B.; Taylor, K.M. Characterization of Zinc Influx Transporters (ZIPs) in Pancreatic β Cells Roles in Regulating Cytosolic Zinc Homeostasis and Insulin Secretion. J. Biol. Chem. 2015, 290, 18757–18769. [Google Scholar] [CrossRef]
- Jewell, J.L.; Oh, E.; Ramalingam, L.; Kalwat, M.A.; Tagliabracci, V.S.; Tackett, L.; Elmendorf, J.S.; Thurmond, D.C. Munc18c phosphorylation by the insulin receptor links cell signaling directly to SNARE exocytosis. J. Cell Biol. 2011, 193, 185–199. [Google Scholar] [CrossRef]
- Song, E.K.; Lee, Y.R.; Kim, Y.R.; Yeom, J.H.; Yoo, C.H.; Kim, H.K.; Park, H.M.; Kang, H.S.; Kim, J.S.; Kim, U.H.; et al. NAADP mediates insulin-stimulated glucose uptake and insulin sensitization by PPARgamma in adipocytes. Cell Rep. 2012, 2, 1607–1619. [Google Scholar] [CrossRef] [PubMed]
- Zacherl, J.R.; Tourkova, I.; St Croix, C.M.; Robinson, L.J.; Peck Palmer, O.M.; Mihalik, S.J.; Blair, H.C. Elaidate, an 18-carbon trans-monoenoic fatty acid, but not physiological fatty acids increases intracellular Zn(2+) in human macrophages. J. Cell. Biochem. 2015, 116, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Zhang, L.; Biswas, S.; Schugar, R.C.; Brown, J.M.; Byzova, T.; Podrez, E. Akt3 inhibits adipogenesis and protects from diet-induced obesity via WNK1/SGK1 signaling. JCI Insight 2017, 2, e95687. [Google Scholar] [CrossRef] [PubMed]
- Brozinick, J.T.; Roberts, B.R.; Dohm, G.L. Defective Signaling Through Akt-2 and -3 but Not Akt-1 in Insulin-Resistant Human Skeletal Muscle. Potential Role Insul. Resist. 2003, 52, 935–941. [Google Scholar]
- Brown, D.M.; Williams, H.; Ryan, K.J.; Wilson, T.L.; Daniel, Z.C.; Mareko, M.H.; Emes, R.D.; Harris, D.W.; Jones, S.; Wattis, J.A.; et al. Mitochondrial phosphoenolpyruvate carboxykinase (PEPCK-M) and serine biosynthetic pathway genes are co-ordinately increased during anabolic agent-induced skeletal muscle growth. Sci. Rep. 2016, 6, 28693. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.K.; Singh, J.; Dey, C.S. PPAR-gamma expression modulates insulin sensitivity in C2C12 skeletal muscle cells. Br. J. Pharmacol. 2004, 143, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Miles, P.D.; Barak, Y.; He, W.; Evans, R.M.; Olefsky, J.M. Improved insulin-sensitivity in mice heterozygous for PPAR-gamma deficiency. J. Clin. Investig. 2000, 105, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Waki, H.; Murakami, K.; Motojima, K.; Komeda, K.; Ide, T.; Kubota, N.; Terauchi, Y.; Tobe, K.; et al. The mechanisms by which both heterozygous peroxisome proliferator-activated receptor gamma (PPARgamma) deficiency and PPARgamma agonist improve insulin resistance. J. Biol. Chem. 2001, 276, 41245–41254. [Google Scholar] [CrossRef]
- Sun, Q.; Li, Q.; Zhong, W.; Zhang, J.; Sun, X.; Tan, X.; Yin, X.; Sun, X.; Zhang, X.; Zhou, Z. Dysregulation of hepatic zinc transporters in a mouse model of alcoholic liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G313–G322. [Google Scholar] [CrossRef]
- Singh, J.; Verma, N.K.; Kansagra, S.M.; Kate, B.N.; Dey, C.S. Altered PPARgamma expression inhibits myogenic differentiation in C2C12 skeletal muscle cells. Mol. Cell. Biochem. 2007, 294, 163–171. [Google Scholar] [CrossRef]
- Mnatsakanyan, H.; i Serra, R.S.; Rico, P.; Salmerón-Sánchez, M. Zinc uptake promotes myoblast differentiation via Zip7 transporter and activation of Akt signalling transduction pathway. Sci. Rep. 2018, 8, 13642. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, H.; Inoue, A.; Toyama-Sorimachi, N.; Nagai, Y.; Yasuda, T.; Suzuki, H.; Horai, R.; Iwakura, Y.; Yamamoto, T.; Karasuyama, H.; et al. Dok-1 and Dok-2 are negative regulators of lipopolysaccharide-induced signaling. J. Exp. Med. 2005, 201, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Shirakata, M.; Iwama, A.; Ishii, A.; Ebihara, Y.; Osawa, M.; Honda, K.; Shinohara, H.; Sudo, K.; Tsuji, K.; et al. Role of Dok-1 and Dok-2 in myeloid homeostasis and suppression of leukemia. J. Exp. Med. 2004, 200, 1681–1687. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Hancock, M.K.; Pitman, J.L.; Orth, A.P.; Gekakis, N. Negative regulators of insulin signaling revealed in a genome-wide functional screen. PLoS ONE 2009, 4, e6871. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.C.; Huang, D.Y.; Shiah, S.G.; Wang, J.S.; Lin, W.W. Regulation of c-Fos gene expression by NF-kappaB: A p65 homodimer binding site in mouse embryonic fibroblasts but not human HEK293 cells. PLoS ONE 2013, 8, e84062. [Google Scholar] [CrossRef]
- Delghandi, M.P.; Johannessen, M.; Moens, U. The cAMP signalling pathway activates CREB through PKA, p38 and MSK1 in NIH 3T3 cells. Cell Signal. 2005, 17, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Hojyo, S.; Fukada, T.; Shimoda, S.; Ohashi, W.; Bin, B.H.; Koseki, H.; Hirano, T. The zinc transporter SLC39A14/ZIP14 controls G-protein coupled receptor-mediated signaling required for systemic growth. PLoS ONE 2011, 6, e18059. [Google Scholar] [CrossRef]
- Yan, G.; Zhang, Y.; Yu, J.; Yu, Y.; Zhang, F.; Zhang, Z.; Wu, A.; Yan, X.; Zhou, Y.; Wang, F. Slc39a7/zip7 plays a critical role in development and zinc homeostasis in zebrafish. PLoS ONE 2012, 7, e42939. [Google Scholar] [CrossRef]
- Fukada, T.; Yamasaki, S.; Nishida, K.; Murakami, M.; Hirano, T. Zinc homeostasis and signaling in health and diseases. J. Biol. Inorg. Chem. 2011, 16, 1123–1134. [Google Scholar] [CrossRef]
- Castellano, E.; Santos, E. Functional specificity of ras isoforms: So similar but so different. Genes Cancer 2011, 2, 216–231. [Google Scholar] [CrossRef]
- Klein, C.; Creach, K.; Irintcheva, V.; Hughes, K.J.; Blackwell, P.L.; Corbett, J.A.; Baldassare, J.J. Zinc induces ERK-dependent cell death through a specific Ras isoform. Apoptosis Int. J. Program. Cell Death 2006, 11, 1933–1944. [Google Scholar] [CrossRef] [PubMed]
- Hennigar, S.R.; Velasquez, V.; Kelleher, S.L. Obesity-Induced Inflammation Is Associated with Alterations in Subcellular Zinc Pools and Premature Mammary Gland Involution in Lactating Mice. J. Nutr. 2015, 145, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Tinkov, A.A.; Popova, E.V.; Gatiatulina, E.R.; Skalnaya, A.A.; Yakovenko, E.N.; Alchinova, I.B.; Karganov, M.Y.; Skalny, A.V.; Nikonorov, A.A. Decreased adipose tissue zinc content is associated with metabolic parameters in high fat fed Wistar rats. Acta Sci. Pol. Technol. Aliment. 2016, 15, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Cooper-Capetini, V.; de Vasconcelos, D.A.A.; Martins, A.R.; Hirabara, S.M.; Donato, J., Jr.; Carpinelli, A.R.; Abdulkader, F. Zinc Supplementation Improves Glucose Homeostasis in High Fat-Fed Mice by Enhancing Pancreatic beta-Cell Function. Nutrients 2017, 9, 1150. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.J.; Bao, S.; Bolin, E.R.; Burris, D.L.; Xu, X.; Sun, Q.; Killilea, D.W.; Shen, Q.; Ziouzenkova, O.; Belury, M.A.; et al. Zinc deficiency augments leptin production and exacerbates macrophage infiltration into adipose tissue in mice fed a high-fat diet. J. Nutr. 2013, 143, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, G.; Matera, M.G.; De Martino, B.; Vacca, C.; Di Martino, S.; Rossi, F. Relationship between zinc and obesity. J. Med. 1993, 24, 177–183. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Fold–Up or Downregulation | p–value |
|---|---|---|
| Insulin Receptor–Associated Proteins | ||
| Eukaryotic translation initiation factor 4E binding protein 1 (Eif4ebp1) | −5.42 | 0.109812 |
| Insulin−like growth factor 2 (Igf2) | −2.51 | 0.119985 |
| Growth factor receptor bound protein 2 (Grb2) | −1.81 | 0.044301 |
| Fibroblast growth factor receptor substrate 3 (Frs3) | −1.78 | 0.342511 |
| Sorbin and SH3 domain containing 1 (Sorbs1) | −1.57 | 0.261979 |
| Protein tyrosine phosphatase, receptor type, F (Ptprf) | −1.49 | 0.376424 |
| Growth factor receptor bound protein 2−associated protein 1 (Gab1) | −1.48 | 0.237770 |
| Non−catalytic region of tyrosine kinase adaptor protein 1 (Nck1) | −1.24 | 0.479138 |
| Fibroblast growth factor receptor substrate 2 (Frs2) | −1.21 | 0.514579 |
| Protein tyrosine phosphatase, non−receptor type 1 (Ptpn1) | −1.19 | 0.481621 |
| Heat shock protein 90 alpha (cytosolic), class B member 1 (Hsp90ab1) | 1.04 | 0.965684 |
| src homology 2 domain−containing transforming protein C1 (Shc1) | 1.13 | 0.659820 |
| Protein phosphatase 1 catalytic subunit alpha (Ppp1ca) | 1.15 | 0.595936 |
| Casitas B−lineage lymphoma (Cb1) | 1.15 | 0.759472 |
| Insulin receptor substrate 1 (Irs1) | 1.47 | 0.435010 |
| Insulin−like growth factor I receptor (Igfr) | 1.54 | 0.241275 |
| Insulin receptor substrate 2 (Irs2) | 1.67 | 0.240410 |
| Prolactin (Prl) | 1.82 | 0.518438 |
| Insulin 1 (Ins1) | 2.04 | 0.401028 |
| Insulin−like growth factor binding protein 1 (Igfbp1) | 2.14 | 0.391212 |
| Neuropeptide Y (Npy) | 2.60 | 0.057738 |
| Docking protein 1 (Dok1) | 2.91 | 0.103312 |
| Docking protein 3 (Dok3) | 3.30 | 0.315765 |
| Growth factor receptor bound protein 10 (Grb10) | 3.75 | 0.140065 |
| Insulin−like 3 (InsI3) | 4.33 | 0.287920 |
| Thyroglobulin (Tg) | 5.53 | 0.179122 |
| Docking protein 2 (Dok2) | 21.79 | 0.031048 |
| PI3 Kinase Signaling | ||
| Thymoma viral proto−oncogene 3 (Akt3) | −2.16 | 0.019032 |
| Phosphoinositide−3−kinase regulatory subunit 1 (Pik3r1) | −1.57 | 0.350159 |
| Thymoma viral proto−oncogene 2 (Akt2) | −1.46 | 0.384953 |
| Protein kinase C, gamma (Prkcg) | −1.42 | 0.281971 |
| Eukaryotic translation initiation factor 2B, subunit 1 (alpha) (Eif2b1) | −1.39 | 0.219592 |
| Phosphatidylinositol−4,5−bisphosphate 3−kinase catalytic subunit alpha (Pik3ca) | −1.38 | 0.333187 |
| Phosphatidylinositol−4,5−bisphosphate 3−kinase catalytic subunit beta (Pik3cb) | −1.30 | 0.444354 |
| 3−phosphoinositide dependent protein kinase 1 (Pdpk1) | −1.15 | 0.751995 |
| Protein kinase C, iota (Prkci) | −1.14 | 0.707935 |
| Mechanistic target of rapamycin kinase (Mtor) | 1.08 | 0.874784 |
| BCL2−like 1 (Bcl2l1) | 1.21 | 0.631682 |
| Phosphoinositide−3−kinase regulatory subunit 2 (Pik3r2) | 1.22 | 0.653162 |
| Adrenergic receptor, alpha 1d (Adra1d) | 1.25 | 0.391856 |
| Thymoma viral proto−oncogene 1 (Akt1) | 1.36 | 0.241863 |
| Dual specificity phosphatase 14 (Dusp14) | 1.42 | 0.465154 |
| Protein kinase C, zeta (Prkcz) | 2.00 | 0.327070 |
| Serine peptidase inhibitor, clade E, member 1 (Serpine 1) | 2.09 | 0.131939 |
| MAP Kinase Signaling | ||
| Related RAS viral (r−ras) oncogene 2 (Rras2) | −1.86 | 0.075688 |
| Braf transforming gene (Braf) | −1.78 | 0.127855 |
| Related RAS viral (r−ras) oncogene (Rras) | −1.63 | 0.070002 |
| Araf proto−oncogene, serine/threonine kinase (Araf) | −1.40 | 0.331800 |
| SOS Ras/Rac guanine nucleotide exchange factor 1 (Sos1) | −1.21 | 0.477393 |
| Excision repair cross−complementing rodent repair deficiency, complementation group 1 (Ercc1) | −1.02 | 0.884782 |
| v−raf−leukemia viral oncogene 1 (raf1) | 1.03 | 0.834133 |
| Uncoupling protein 1 (mitochondrial, proton carrier) (Ucp1) | 1.26 | 0.433977 |
| Mitogen−activated protein kinase kinase 1 (Map2k1) | 1.43 | 0.244012 |
| Mitogen−activated protein kinase 1 (Mapk1) | 1.82 | 0.134200 |
| Kruppel−like factor 10 (Klf10) | 2.12 | 0.143675 |
| Nitric oxide synthase 2, inducible (Nos2) | 2.21 | 0.030364 |
| Harvey rat sarcoma virus oncogene (Hras) | 2.30 | 0.027193 |
| Ribosomal protein S6 kinase polypeptide 1 (Rps6ka1) | 2.93 | 0.286011 |
| FBJ osteosarcoma oncogene (Fos) | 4.42 | 0.014490 |
| Carbohydrate Metabolism | ||
| Phosphoenolpyruvate carboxykinase 2 (mitochondrial) (Pck2) | −2.73 | 0.046319 |
| Glucuronidase, beta (Gusb) | −2.44 | 0.057494 |
| Pyruvate kinase liver (Pkl) | −2.00 | 0.361565 |
| AE binding protein 1 (Aebp1) | −1.36 | 0.346200 |
| acyl−Coenzyme A oxidase 1, palmitoyl (Acox1) | −1.23 | 0.517860 |
| Hexokinase 2 (Hk2) | 1.06 | 0.892955 |
| Glycogen synthase kinase 3 beta (Gsk3b) | 1.04 | 0.908617 |
| Glyceraldehyde−3−phosphate dehydrogenase (Gapdh) | 1.15 | 0.642638 |
| Glucose−6−phosphatase, catalytic (G6pc) | 1.22 | 0.432012 |
| Glycerol−3−phosphate dehydrogenase 1 (Gpd1) | 1.43 | 0.434684 |
| Solute carrier family 2 (facilitated glucose transporter), member 1 (Slc2a1) | 1.50 | 0.246541 |
| Glucokinase (Gck) | 1.96 | 0.316235 |
| Fructose bisphosphatase 1 (Fbp1) | 3.40 | 0.315244 |
| Glucose−6−phosphatase, catalytic, 2 (G6pc2) | 4.71 | 0.189288 |
| Lipid Metabolism | ||
| Complement factor D (adipsin) (Cfd) | −3.66 | 0.370569 |
| Peroxisome proliferator activated receptor gamma (Pparg) | −3.31 | 0.007128 |
| Sterol regulatory element binding transcription factor 1 (Srebf1) | −1.59 | 0.041612 |
| Resistin (Retn) | −1.71 | 0.786857 |
| Low density lipoprotein receptor (Ldlr) | −1.11 | 0.938288 |
| Leptin (Lep) | 1.31 | 0.490170 |
| Solute carrier family 27 (fatty acid transporter), member 4 (Slc27a4) | 1.33 | 0.350357 |
| Cell Growth and Differentiation | ||
| Kirsten rat sarcoma viral oncogene homolog (Kras) | 2.12 | 0.025606 |
| CCAAT/enhancer binding protein (C/EBP), alpha (Cebpa) | −1.41 | 0.700291 |
| Vascular endothelial growth factor A (Vegfa) | −1.14 | 0.660055 |
| CAP, adenylate cyclase−associated protein 1 (Cap1) | −1.13 | 0.673316 |
| Jun proto−oncogene (Jun) | 1.02 | 0.996474 |
| Beta−2 microglobulin (B2m) | 1.13 | 0.700177 |
| Actin, beta (Actb) | 1.28 | 0.577259 |
| CCAAT/enhancer binding protein (C/EBP), beta (Cebpb) | 2.33 | 0.145381 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Norouzi, S.; Adulcikas, J.; Henstridge, D.C.; Sonda, S.; Sohal, S.S.; Myers, S. The Zinc Transporter Zip7 Is Downregulated in Skeletal Muscle of Insulin-Resistant Cells and in Mice Fed a High-Fat Diet. Cells 2019, 8, 663. https://doi.org/10.3390/cells8070663
Norouzi S, Adulcikas J, Henstridge DC, Sonda S, Sohal SS, Myers S. The Zinc Transporter Zip7 Is Downregulated in Skeletal Muscle of Insulin-Resistant Cells and in Mice Fed a High-Fat Diet. Cells. 2019; 8(7):663. https://doi.org/10.3390/cells8070663
Chicago/Turabian StyleNorouzi, Shaghayegh, John Adulcikas, Darren C Henstridge, Sabrina Sonda, Sukhwinder Singh Sohal, and Stephen Myers. 2019. "The Zinc Transporter Zip7 Is Downregulated in Skeletal Muscle of Insulin-Resistant Cells and in Mice Fed a High-Fat Diet" Cells 8, no. 7: 663. https://doi.org/10.3390/cells8070663
APA StyleNorouzi, S., Adulcikas, J., Henstridge, D. C., Sonda, S., Sohal, S. S., & Myers, S. (2019). The Zinc Transporter Zip7 Is Downregulated in Skeletal Muscle of Insulin-Resistant Cells and in Mice Fed a High-Fat Diet. Cells, 8(7), 663. https://doi.org/10.3390/cells8070663