Association of the Rheumatoid Arthritis Severity Variant rs26232 with the Invasive Activity of Synovial Fibroblasts

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of RASF
2.2. rs26232 Genotyping
2.3. RASF Invasion
2.4. RASF Migration
2.5. RASF Proliferation
2.6. Flow Cytometry
2.7. Cytokine and Protease Production
2.8. Real-Time PCR
2.9. Data Analysis
3. Results
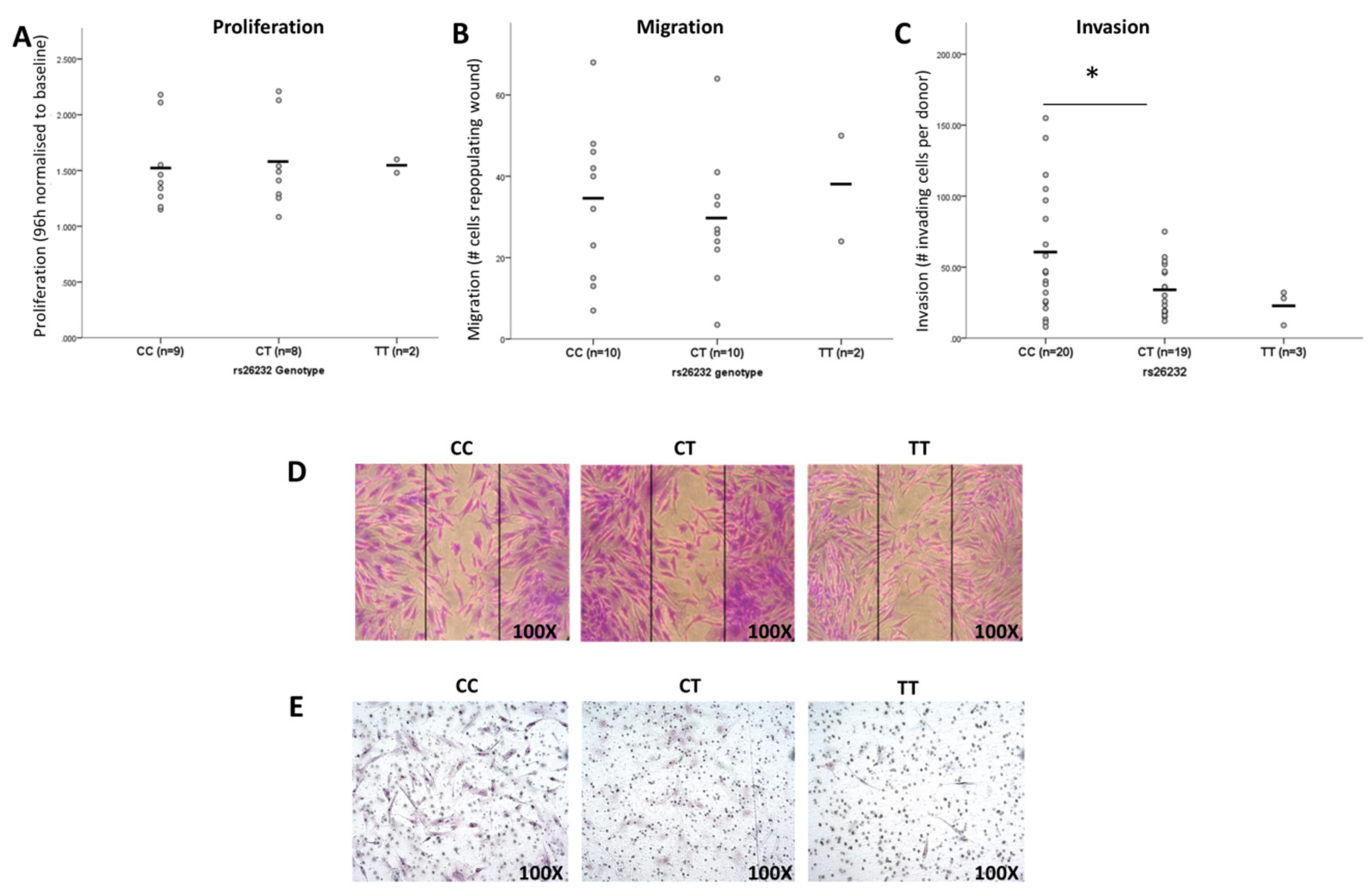
3.1. Association of the rs26232 CC Genotype with Greater Invasiveness of RASFs In Vitro
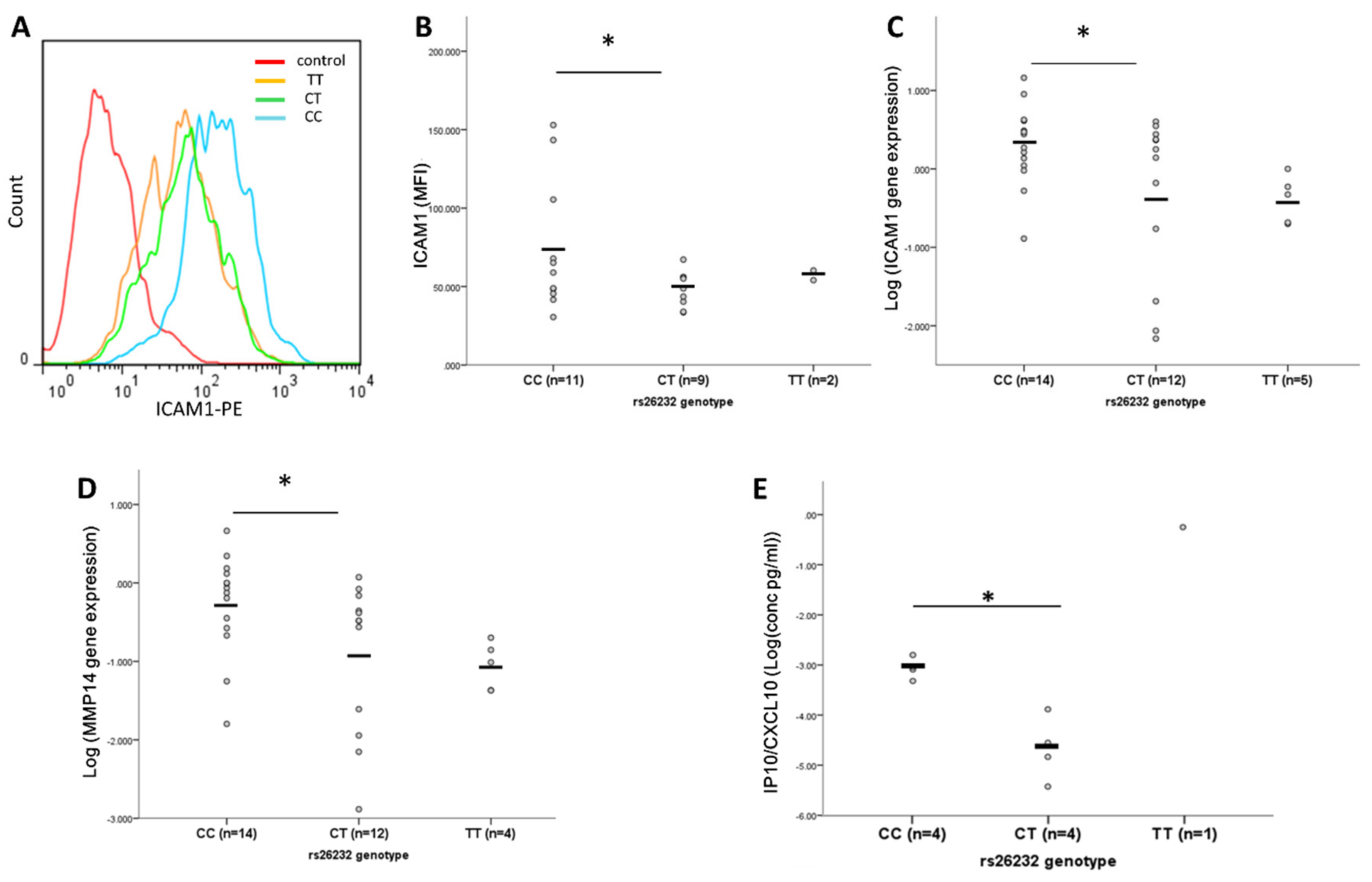
3.2. Higher ICAM-1, MMP14, and IP-10 Production by RASFs of CC Compared to CT Genotype
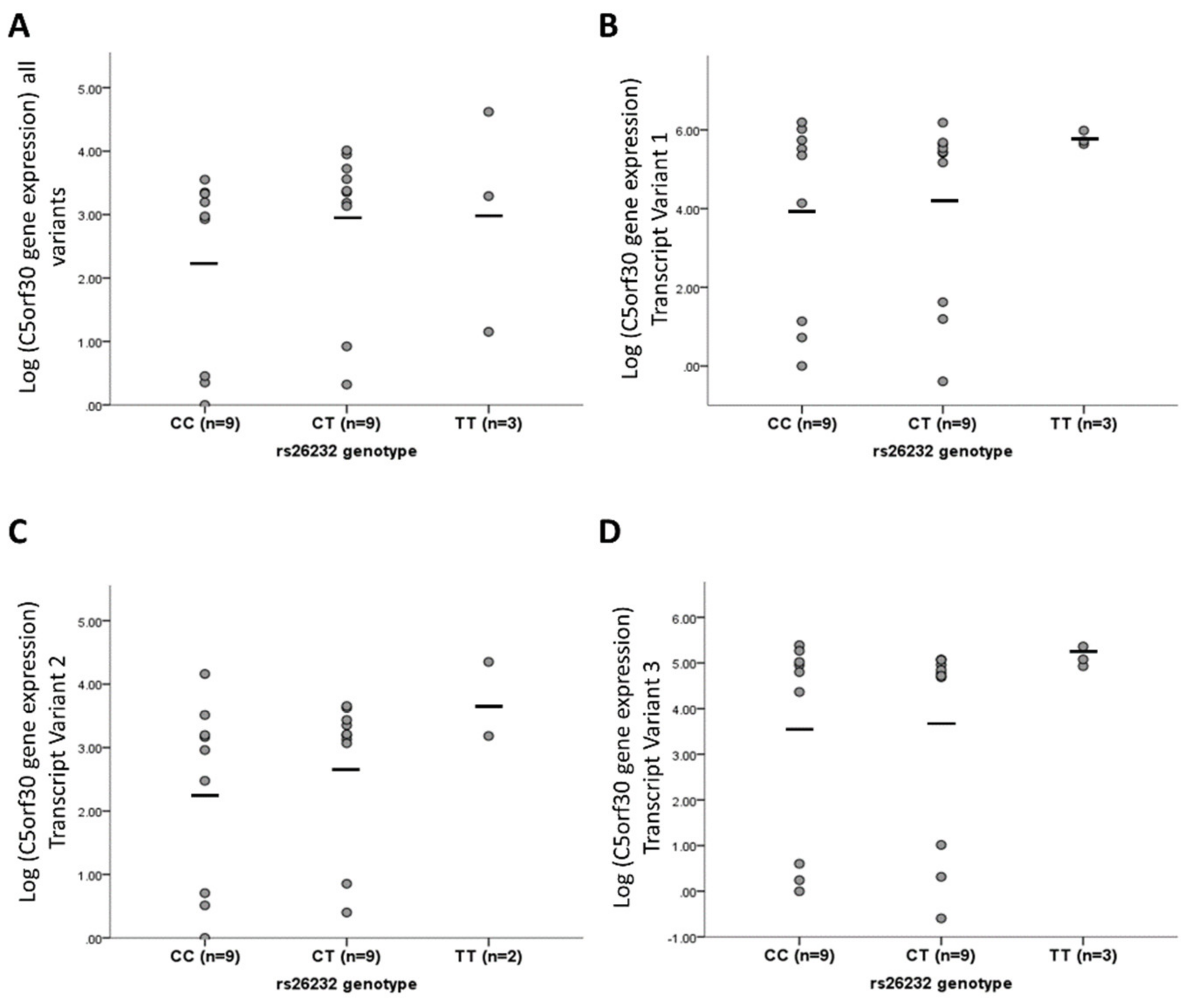
3.3. rs26232 Genotype Is Not Associated with C5orf30 mRNA Expression in RASFs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Stahl, E.A.; Raychaudhuri, S.; Remmers, E.F.; Xie, G.; Eyre, S.; Thomson, B.P.; Li, Y.; Kurreeman, F.A.; Zhernakova, A.; Hinks, A.; et al. Genome-wide association study meta-analysis identifies seven new rheumatoid arthritis risk loci. Nat. Genet. 2010, 42, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Cordell, H.J.; Han, Y.; Mells, G.F.; Li, Y.; Hirschfield, G.M.; Greene, C.S.; Xie, G.; Juran, B.D.; Zhu, D.; Qian, D.C.; et al. International genome-wide meta-analysis identifies new primary biliary cirrhosis risk loci and targetable pathogenic pathways. Nat. Commun. 2015, 6, 8019. [Google Scholar] [CrossRef] [PubMed]
- Teare, M.D.; Knevel, R.; Morgan, M.D.; Kleszcz, A.; Emery, P.; Moore, D.J.; Conaghan, P.; Huizinga, T.W.; Morgan, A.W.; van der Helm-van Mil, A.H.; et al. Allele-dose association of the C5orf30 rs26232 variant with joint damage in rheumatoid arthritis. Arthritis Rheum. 2013, 65, 2555–2561. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. New Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef]
- Muthana, M.; Hawtree, S.; Wilshaw, A.; Linehan, E.; Roberts, H.; Khetan, S.; Adeleke, G.; Wright, F.; Akil, M.; Fearon, U.; et al. C5orf30 is a negative regulator of tissue damage in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2015, 112, 11618–11623. [Google Scholar] [CrossRef]
- Dorris, E.R.; Tazzyman, S.J.; Moylett, J.; Ramamoorthi, N.; Hackney, J.; Townsend, M.; Muthana, M.; Lewis, M.J.; Pitzalis, C.; Wilson, A.G. The Autoimmune Susceptibility Gene C5orf30 Regulates Macrophage-Mediated Resolution of Inflammation. J. Immunol. 2019, 202, 1069–1078. [Google Scholar] [CrossRef]
- Abdul-Sater, A.A.; Edilova, M.I.; Clouthier, D.L.; Mbanwi, A.; Kremmer, E.; Watts, T.H. The signaling adaptor TRAF1 negatively regulates Toll-like receptor signaling and this underlies its role in rheumatic disease. Nat. Immunol. 2017, 18, 26–35. [Google Scholar] [CrossRef]
- Noss, E.H.; Nguyen, H.N.; Chang, S.K.; Watts, G.F.; Brenner, M.B. Genetic polymorphism directs IL-6 expression in fibroblasts but not selected other cell types. Proc. Natl. Acad. Sci. USA 2015, 112, 14948–14953. [Google Scholar] [CrossRef]
- Dorris, E.R.; Blackshields, G.; Sommerville, G.; Alhashemi, M.; Dias, A.; McEneaney, V.; Smyth, P.; O’Leary, J.J.; Sheils, O. Pluripotency markers are differentially induced by MEK inhibition in thyroid and melanoma BRAFV600E cell lines. Cancer Biol. Ther. 2016, 17, 526–542. [Google Scholar] [CrossRef]
- Dorris, E.; O’Neill, A.; Hanrahan, K.; Treacy, A.; Watson, R.W. MARCKS promotes invasion and is associated with biochemical recurrence in prostate cancer. Oncotarget 2017, 8, 72021–72030. [Google Scholar] [CrossRef]
- Huber, L.C.; Distler, O.; Tarner, I.; Gay, R.E.; Gay, S.; Pap, T. Synovial fibroblasts: Key players in rheumatoid arthritis. Rheumatology 2006, 45, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Niederer, F.; Trenkmann, M.; Ospelt, C.; Karouzakis, E.; Neidhart, M.; Stanczyk, J.; Kolling, C.; Gay, R.E.; Detmar, M.; Gay, S.; et al. Down-regulation of microRNA-34a* in rheumatoid arthritis synovial fibroblasts promotes apoptosis resistance. Arthritis Rheum. 2012, 64, 1771–1779. [Google Scholar] [CrossRef]
- Muller-Ladner, U.; Kriegsmann, J.; Franklin, B.N.; Matsumoto, S.; Geiler, T.; Gay, R.E.; Gay, S. Synovial fibroblasts of patients with rheumatoid arthritis attach to and invade normal human cartilage when engrafted into SCID mice. Am. J. Pathol. 1996, 149, 1607–1615. [Google Scholar] [PubMed]
- Tolboom, T.C.; van der Helm-Van Mil, A.H.; Nelissen, R.G.; Breedveld, F.C.; Toes, R.E.; Huizinga, T.W. Invasiveness of fibroblast-like synoviocytes is an individual patient characteristic associated with the rate of joint destruction in patients with rheumatoid arthritis. Arthritis Rheum. 2005, 52, 1999–2002. [Google Scholar] [CrossRef] [PubMed]
- Fearon, U.; Mullan, R.; Markham, T.; Connolly, M.; Sullivan, S.; Poole, A.R.; FitzGerald, O.; Bresnihan, B.; Veale, D.J. Oncostatin M induces angiogenesis and cartilage degradation in rheumatoid arthritis synovial tissue and human cartilage cocultures. Arthritis Rheum. 2006, 54, 3152–3162. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuka, K.; Tanaka, Y.; Hubscher, S.; Abe, M.; Wake, A.; Saito, K.; Morimoto, I.; Eto, S. Rheumatoid synovial fibroblasts are stimulated by the cellular adhesion to T cells through lymphocyte function associated antigen-1/intercellular adhesion molecule-1. J. Rheumatol. 1997, 24, 458–464. [Google Scholar]
- Hanyuda, M.; Kasama, T.; Isozaki, T.; Matsunawa, M.M.; Yajima, N.; Miyaoka, H.; Uchida, H.; Kameoka, Y.; Ide, H.; Adachi, M. Activated leucocytes express and secrete macrophage inflammatory protein-1 α upon interaction with synovial fibroblasts of rheumatoid arthritis via a β 2-integrin/ICAM-1 mechanism. Rheumatology 2003, 42, 1390–1397. [Google Scholar] [CrossRef]
- Hanaoka, R.; Kasama, T.; Muramatsu, M.; Yajima, N.; Shiozawa, F.; Miwa, Y.; Negishi, M.; Ide, H.; Miyaoka, H.; Uchida, H.; et al. A novel mechanism for the regulation of IFN-gamma inducible protein-10 expression in rheumatoid arthritis. Arthritis Res. Ther. 2003, 5, R74–R81. [Google Scholar] [CrossRef]
- Pap, T.; Shigeyama, Y.; Kuchen, S.; Fernihough, J.K.; Simmen, B.; Gay, R.E.; Billingham, M.; Gay, S. Differential expression pattern of membrane-type matrix metalloproteinases in rheumatoid arthritis. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 2000, 43, 1226–1232. [Google Scholar] [CrossRef]
- Konttinen, Y.; Ceponis, A.; Takagi, M.; Ainola, M.; Sorsa, T.; Sutinen, M.; Salo, T.; Ma, J.; Santavirta, S.; Seiki, M. New collagenolytic enzymes/cascade identified at the pannus-hard tissue junction in rheumatoid arthritis: Destruction from above. Matrix Biol. 1998, 17, 585–601. [Google Scholar] [CrossRef]
- Mizoguchi, F.; Slowikowski, K.; Wei, K.; Marshall, J.L.; Rao, D.A.; Chang, S.K.; Nguyen, H.N.; Noss, E.H.; Turner, J.D.; Earp, B.E.; et al. Functionally distinct disease-associated fibroblast subsets in rheumatoid arthritis. Nat. Commun. 2018, 9, 789. [Google Scholar] [CrossRef] [PubMed]
- Croft, A.P.; Campos, J.; Jansen, K.; Turner, J.D.; Marshall, J.; Attar, M.; Savary, L.; Wehmeyer, C.; Naylor, A.J.; Kemble, S.; et al. Distinct fibroblast subsets drive inflammation and damage in arthritis. Nature 2019, 570, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Joehanes, R.; Chen, B.H.; Huan, T.; Ying, S.; Munson, P.J.; Johnson, A.D.; Levy, D.; O'Donnell, C.J. Identification of common genetic variants controlling transcript isoform variation in human whole blood. Nat. Genet. 2015, 47, 345–352. [Google Scholar] [CrossRef] [PubMed]
- eGTEx Project. Enhancing GTEx by bridging the gaps between genotype, gene expression, and disease. Nat. Genet. 2017, 49, 1664–1670. [Google Scholar] [CrossRef]
- Consortium, E.P. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57. [Google Scholar] [CrossRef]
- Robert, F.; Pelletier, J. Exploring the Impact of Single-Nucleotide Polymorphisms on Translation. Front Genet 2018, 9, 507. [Google Scholar] [CrossRef]
- Nishizaki, S.S.; Boyle, A.P. Mining the Unknown: Assigning Function to Noncoding Single Nucleotide Polymorphisms. Trends Genet 2017, 33, 34–45. [Google Scholar] [CrossRef]
- McDaniell, R.; Lee, B.-K.; Song, L.; Liu, Z.; Boyle, A.P.; Erdos, M.R.; Scott, L.J.; Morken, M.A.; Kucera, K.S.; Battenhouse, A. Heritable individual-specific and allele-specific chromatin signatures in humans. Science 2010, 328, 235–239. [Google Scholar] [CrossRef]
- Maurano, M.T.; Humbert, R.; Rynes, E.; Thurman, R.E.; Haugen, E.; Wang, H.; Reynolds, A.P.; Sandstrom, R.; Qu, H.; Brody, J. Systematic localization of common disease-associated variation in regulatory DNA. Science 2012, 337, 1190–1195. [Google Scholar] [CrossRef]
- Schaub, M.A.; Boyle, A.P.; Kundaje, A.; Batzoglou, S.; Snyder, M. Linking disease associations with regulatory information in the human genome. Genome Res. 2012, 22, 1748–1759. [Google Scholar] [CrossRef]
- Kasowski, M.; Grubert, F.; Heffelfinger, C.; Hariharan, M.; Asabere, A.; Waszak, S.M.; Habegger, L.; Rozowsky, J.; Shi, M.; Urban, A.E. Variation in transcription factor binding among humans. Science 2010, 328, 232–235. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Li, W.; Liyanarachchi, S.; Srinivas, M.; Wang, Y.; Akagi, K.; Wang, Y.; Wu, D.; Wang, Q.; Jin, V.; et al. Multiple functional variants in long-range enhancer elements contribute to the risk of SNP rs965513 in thyroid cancer. Proc. Natl. Acad. Sci. 2015, 112, 6128–6133. [Google Scholar] [CrossRef] [PubMed]
- Gusev, A.; Lee, S.H.; Trynka, G.; Finucane, H.; Vilhjálmsson, B.J.; Xu, H.; Zang, C.; Ripke, S.; Bulik-Sullivan, B.; Stahl, E. Partitioning heritability of regulatory and cell-type-specific variants across 11 common diseases. Am. J. Hum. Genet. 2014, 95, 535–552. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The Human Genome Browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef]
- Bisogno, L.S.; Keene, J.D. RNA regulons in cancer and inflammation. Curr. Opin. Genet. Dev. 2018, 48, 97–103. [Google Scholar] [CrossRef]
- Wang, T.; Chen, Y.-H.; Hong, H.; Zeng, Y.; Zhang, J.; Lu, J.-P.; Jeansonne, B.; Lu, Q. Increased nucleotide polymorphic changes in the 5′-untranslated region of δ-catenin (CTNND2) gene in prostate cancer. Oncogene 2009, 28, 555. [Google Scholar] [CrossRef]
- Corradin, O.; Saiakhova, A.; Akhtar-Zaidi, B.; Myeroff, L.; Willis, J.; Cowper-Sal·lari, R.; Lupien, M.; Markowitz, S.; Scacheri, P.C. Combinatorial effects of multiple enhancer variants in linkage disequilibrium dictate levels of gene expression to confer susceptibility to common traits. Genome Res. 2014, 24, 1–13. [Google Scholar] [CrossRef]
- Neumann, E.; Riepl, B.; Knedla, A.; Lefèvre, S.; Tarner, I.H.; Grifka, J.; Steinmeyer, J.; Schölmerich, J.; Gay, S.; Müller-Ladner, U. Cell culture and passaging alters gene expression pattern and proliferation rate in rheumatoid arthritis synovial fibroblasts. Arthritis Res. Ther. 2010, 12, R83. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorris, E.R.; Linehan, E.; Trenkmann, M.; Veale, D.J.; Fearon, U.; Wilson, A.G. Association of the Rheumatoid Arthritis Severity Variant rs26232 with the Invasive Activity of Synovial Fibroblasts. Cells 2019, 8, 1300. https://doi.org/10.3390/cells8101300
Dorris ER, Linehan E, Trenkmann M, Veale DJ, Fearon U, Wilson AG. Association of the Rheumatoid Arthritis Severity Variant rs26232 with the Invasive Activity of Synovial Fibroblasts. Cells. 2019; 8(10):1300. https://doi.org/10.3390/cells8101300
Chicago/Turabian StyleDorris, Emma R, Eimear Linehan, Michelle Trenkmann, Douglas J Veale, Ursula Fearon, and Anthony G. Wilson. 2019. "Association of the Rheumatoid Arthritis Severity Variant rs26232 with the Invasive Activity of Synovial Fibroblasts" Cells 8, no. 10: 1300. https://doi.org/10.3390/cells8101300
APA StyleDorris, E. R., Linehan, E., Trenkmann, M., Veale, D. J., Fearon, U., & Wilson, A. G. (2019). Association of the Rheumatoid Arthritis Severity Variant rs26232 with the Invasive Activity of Synovial Fibroblasts. Cells, 8(10), 1300. https://doi.org/10.3390/cells8101300