Drosophila HUWE1 Ubiquitin Ligase Regulates Endoreplication and Antagonizes JNK Signaling During Salivary Gland Development

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Strains and Genetics
2.2. Antibody Production
2.3. Other Antibodies Used
2.4. Salivary Gland and Fat Body Dissections, Tissue Preparation, Confocal Microscopy and IMARIS Analysis
2.5. EdU Labeling
2.6. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL) Assay
2.7. Plasmids and Constructs for Expression in S2 Cells
2.8. RNAi and Measurement of Protein Stability in S2 Cells
3. Results
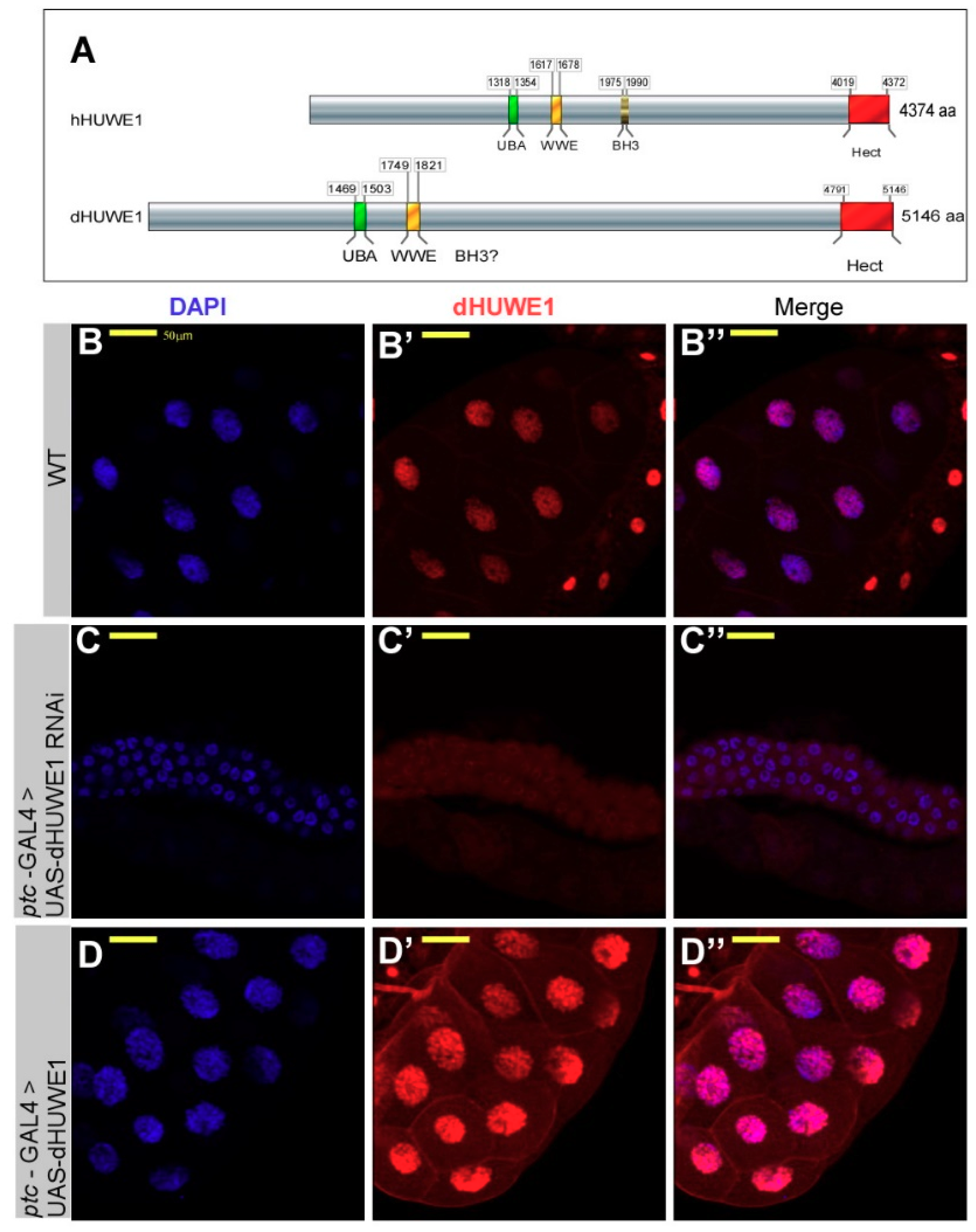
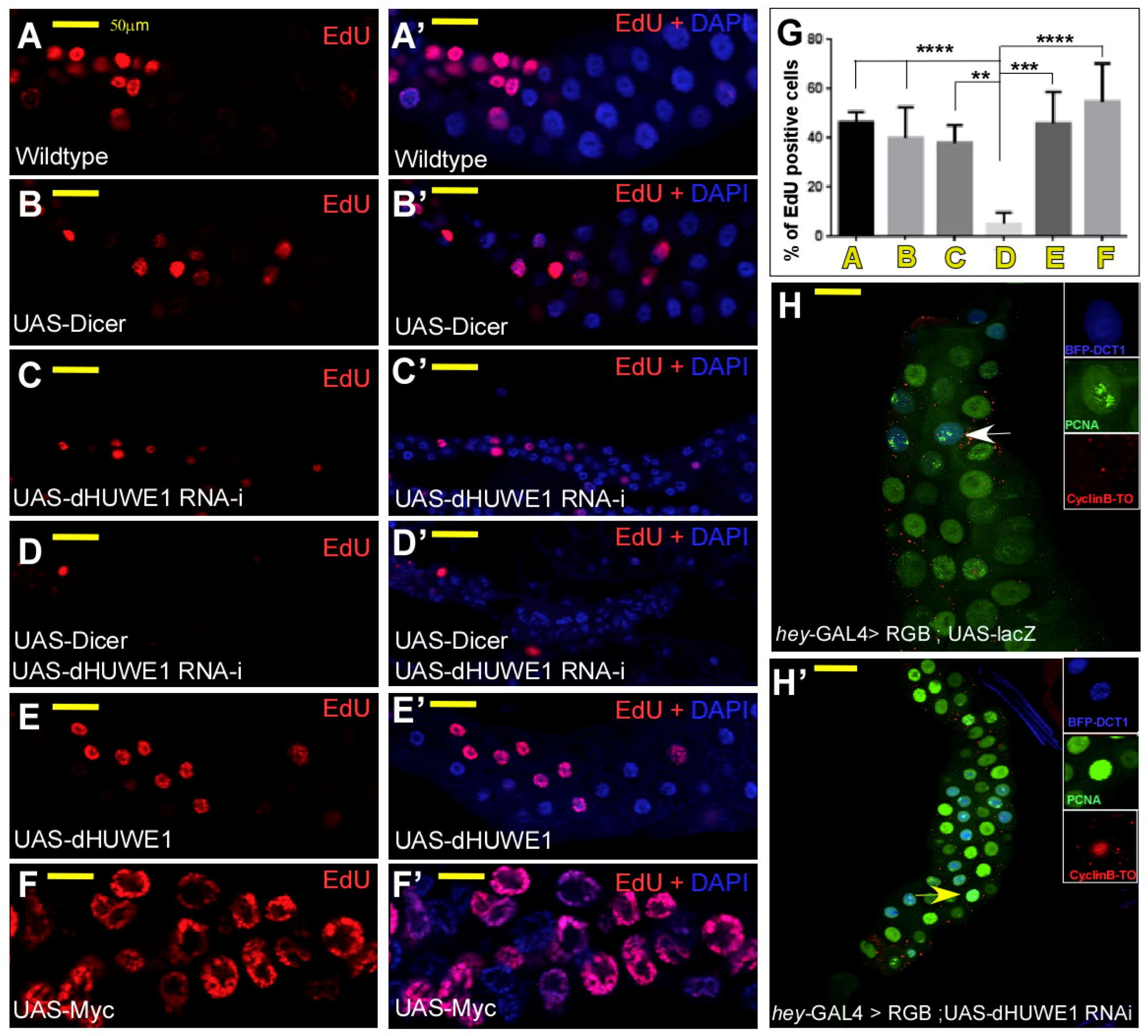
3.1. dHUWE1 Regulates Salivary Gland Development and Endoreplication
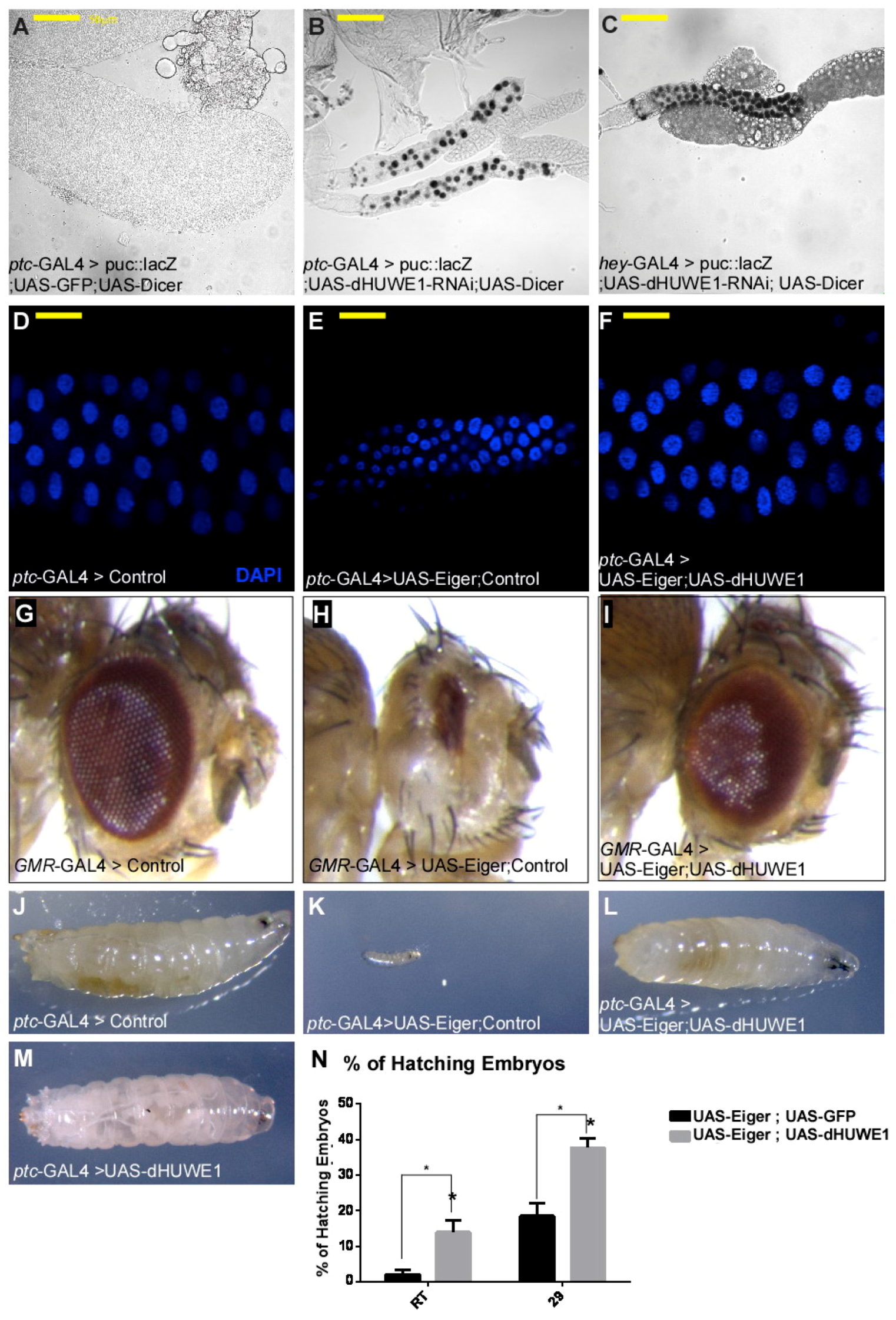
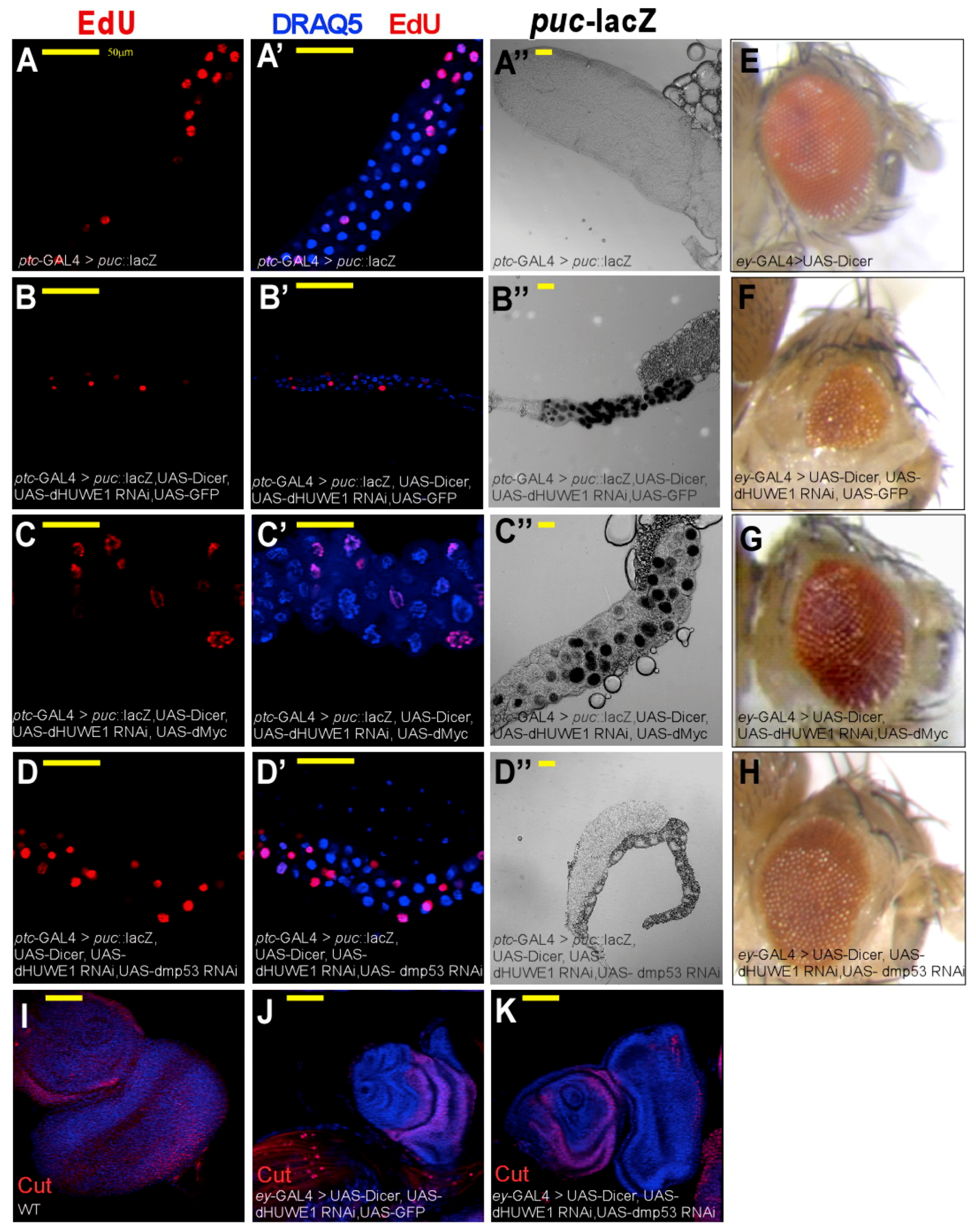
3.2. dHUWE1 Antagonizes JNK Signaling in vivo
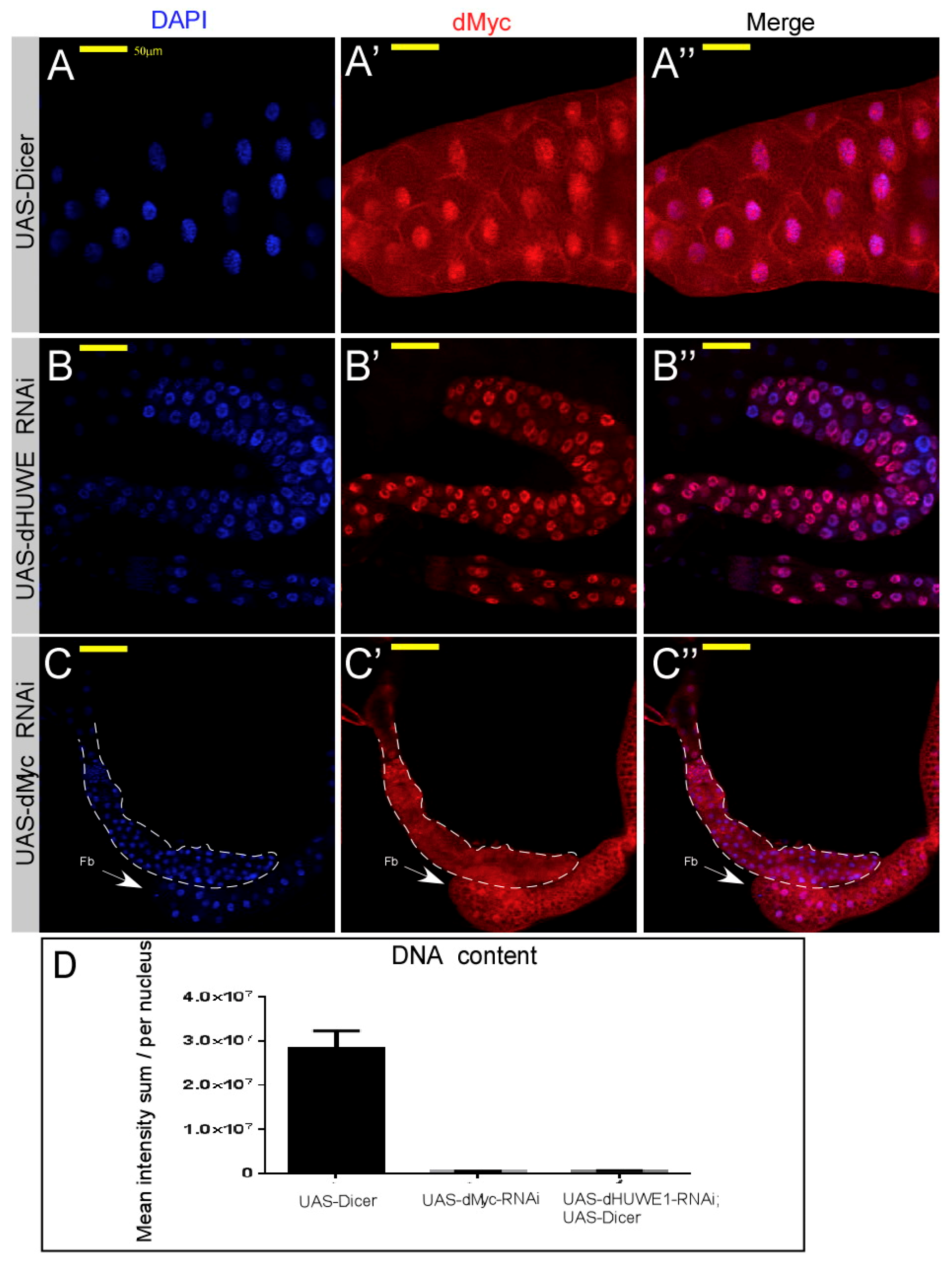
3.3. dHUWE1 Genetically Interacts with dMyc and dmP53
4. Discussion
4.1. dHUWE1 Regulates Endoreplication
4.2. dHUWE1 and the Regulation of the JNK Pathway
4.3. dHUWE1 and the Regulation of dMyc
4.4. Regulation of dHUWE1 by dmP53 in the Absence of Mcl1 and p14ARF
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huibregtse, J.M.; Scheffner, M.; Howley, P.M. A cellular protein mediates association of p53 with the E6 oncoprotein of human papillomavirus types 16 or 18. Embo J. 1991, 10, 4129–4135. [Google Scholar] [CrossRef] [PubMed]
- Bernassola, F.; Karin, M.; Ciechanover, A.; Melino, G. The HECT Family of E3 Ubiquitin Ligases: Multiple Players in Cancer Development. Cancer Cell 2008, 14, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Shabek, N. Ubiquitin Ligases: Structure, Function, and Regulation. Annu. Rev. Biochem. 2017, 86, 129–157. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, S. Structural mechanisms of HECT-type ubiquitin ligases. Biol. Chem. 2018, 399, 127–145. [Google Scholar] [CrossRef] [PubMed]
- Adhikary, S.; Marinoni, F.; Hock, A.; Hulleman, E.; Popov, N.; Beier, R.; Bernard, S.; Quarto, M.; Capra, M.; Goettig, S.; et al. The ubiquitin ligase HectH9 regulates transcriptional activation by Myc and is essential for tumor cell proliferation. Cell 2005, 123, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Kon, N.; Li, M.; Zhang, W.; Qin, J.; Gu, W. ARF-BP1/mule is a critical mediator of the ARF tumor suppressor. Cell 2005, 121, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Oughtred, R.; Wing, S.S. Characterization of E3Histone, a Novel Testis Ubiquitin Protein Ligase Which Ubiquitinates Histones. Mol. Cell. Biol. 2005, 25, 2819–2831. [Google Scholar] [CrossRef] [PubMed]
- Sander, B.; Xu, W.; Eilers, M.; Popov, N.; Lorenz, S. A conformational switch regulates the ubiquitin ligase HUWE1. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Michel, M.A.; Swatek, K.N.; Hospenthal, M.K.; Komander, D. Ubiquitin Linkage-Specific Affimers Reveal Insights into K6-Linked Ubiquitin Signaling. Mol. Cell 2017, 68, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Ohtake, F.; Saeki, Y.; Ishido, S.; Kanno, J.; Tanaka, K. The K48-K63 Branched Ubiquitin Chain Regulates NF-κB Signaling. Mol. Cell 2016, 64, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Shmueli, A.; Oren, M. Life, death, and ubiquitin: Taming the mule. Cell 2005, 121, 963–965. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, M.; Kim, J.; Geradts, J.; Matsuura, K.; Liu, L.; Ran, X.; Xia, W.; Ribar, T.J.; Henao, R.; Dewhirst, M.W.; et al. A network of substrates of the E3 ubiquitin ligases MDM2 and HUWE1 control apoptosis independently of p53. Sci. Signal. 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wilkinson, C.R.M.; Watt, S.; Penkett, C.J.; Toone, W.M.; Jones, N.; Bähler, J. High-Resolution Crystal Structure and In Vivo Function of a Kinesin-2 Homologue in Giardia intestinalis. Mol. Biol. Cell 2007, 19, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Leboucher, G.P.; Tsai, Y.C.; Yang, M.; Shaw, K.C.; Zhou, M.; Veenstra, T.D.; Glickman, M.H.; Weissman, A.M. Stress-Induced Phosphorylation and Proteasomal Degradation of Mitofusin 2 Facilitates Mitochondrial Fragmentation and Apoptosis. Mol. Cell 2012, 47, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Mandemaker, I.K.; Van Cuijk, L.; Janssens, R.C.; Lans, H.; Bezstarosti, K.; Hoeijmakers, J.H.; Demmers, J.A.; Vermeulen, W.; Marteijn, J.A. DNA damage-induced histone H1 ubiquitylation is mediated by HUWE1 and stimulates the RNF8-RNF168 pathway. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Herold, S.; Hock, A.; Herkert, B.; Berns, K.; Mullenders, J.; Beijersbergen, R.; Bernards, R.; Eilers, M. Miz1 and HectH9 regulate the stability of the checkpoint protein, TopBP1. EMBO J. 2008, 27, 2851–2861. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Gao, W.; Du, F.; Wang, X. Mule/ARF-BP1, a BH3-only E3 ubiquitin ligase, catalyzes the polyubiquitination of Mcl-1 and regulates apoptosis. Cell 2005, 121, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Schaub, F.X.; Cleveland, J.L. Tipping the MYC-MIZ1 balance: Targeting the HUWE1 ubiquitin ligase selectively blocks MYC-activated genes. EMBO Mol. Med. 2014, 6, 1509–1511. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Heng, J.I.T.; Guardavaccaro, D.; Jiang, R.; Pagano, M.; Guillemot, F.; Iavarone, A.; Lasorella, A. The HECT-domain ubiquitin ligase Huwe1 controls neural differentiation and proliferation by destabilizing the N-Myc oncoprotein. Nat. Cell Biol. 2008, 10, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Myant, K.B.; Cammareri, P.; Hodder, M.C.; Wills, J.; Von Kriegsheim, A.; Győrffy, B.; Rashid, M.; Polo, S.; Maspero, E.; Vaughan, L.; et al. HUWE1 is a critical colonic tumour suppressor gene that prevents MYC signalling, DNA damage accumulation and tumour initiation. EMBO Mol. Med. 2017, 9, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Brauer, C.; Khatun, R.; Elia, A.J.; Thu, K.L.; Ramachandran, P.; Baniasadi, S.P.; Hao, Z.; Jones, L.D.; Haight, J.; Sheng, Y.; et al. E3 ubiquitin ligase Mule targets β-catenin under conditions of hyperactive Wnt signaling. Proc. Natl. Acad. Sci. USA 2017, 114, E1148–E1157. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; D’Arca, D.; Lim, W.K.; Brahmachary, M.; Carro, M.S.; Ludwig, T.; Cardo, C.C.; Guillemot, F.; Aldape, K.; Califano, A.; et al. The N-Myc-DLL3 Cascade Is Suppressed by the Ubiquitin Ligase Huwe1 to Inhibit Proliferation and Promote Neurogenesis in the Developing Brain. Dev. Cell 2009, 17, 210–221. [Google Scholar] [CrossRef] [PubMed]
- D’Arca, D.; Zhao, X.; Xu, W.; Ramirez-Martinez, N.C.; Iavarone, A.; Lasorella, A. Huwe1 ubiquitin ligase is essential to synchronize neuronal and glial differentiation in the developing cerebellum. Proc. Natl. Acad. Sci. USA 2010, 107, 5875–5880. [Google Scholar] [CrossRef] [PubMed]
- Forget, A.; Bihannic, L.; Cigna, S.M.; Lefevre, C.; Remke, M.; Barnat, M.; Dodier, S.; Shirvani, H.; Mercier, A.; Mensah, A.; et al. Shh Signaling Protects Atoh1 from Degradation Mediated by the E3Ubiquitin Ligase Huwe1 in Neural Precursors. Dev. Cell 2014, 29, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Opperman, K.J.; Mulcahy, B.; Giles, A.C.; Risley, M.G.; Birnbaum, R.L.; Tulgren, E.D.; Dawson-Scully, K.; Zhen, M.; Grill, B. The HECT Family Ubiquitin Ligase EEL-1 Regulates Neuronal Function and Development. Cell Rep. 2017, 19, 822–835. [Google Scholar] [CrossRef] [PubMed]
- Noy, T.; Suad, O.; Taglicht, D.; Ciechanover, A. HUWE1 ubiquitinates MyoD and targets it for proteasomal degradation. Biochem. Biophys. Res. Commun. 2012, 418, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Martinez, S.S.; Hu, W.; Liu, Z.; Tjian, R. A specific E3 ligase/deubiquitinase pair modulates TBP protein levels during muscle differentiation. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Friez, M.J.; Brooks, S.S.; Stevenson, R.E.; Field, M.; Basehore, M.J.; Adès, L.C.; Sebold, C.; Mcgee, S.; Saxon, S.; Skinner, C.; et al. HUWE1 mutations in Juberg-Marsidi and Brooks syndromes: The results of an X-chromosome exome sequencing study. BMJ Open 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Orian, A.; Delrow, J.J.; Rosales Nieves, A.E.; Abed, M.; Metzger, D.; Paroush, Z.; Eisenman, R.N.; Parkhurst, S.M. A Myc Groucho complex integrates EGF and Notch signaling to regulate neural development. Proc. Natl. Acad. Sci. USA 2007, 104, 15771–15776. [Google Scholar] [CrossRef] [PubMed]
- Zielke, N.; Edgar, B.A.; DePamphilis, M.L. Endoreplication. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Handke, B.; Szabad, J.; Lidsky, P.V.; Hafen, E.; Lehner, C.F. Towards long term cultivation of Drosophila wing imaginal discs in vitro. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Prober, D.A.; Edgar, B.A. Interactions between Ras1, dMyc, and dPI3K signaling in the developing Drosophila wing. Genes Dev. 2002, 16, 2286–2299. [Google Scholar] [CrossRef] [PubMed]
- Aris, J.P.; Blobel, G. Identification and characterization of a yeast nucleolar protein that is similar to a rat liver nucleolar protein. J. Cell Biol. 1988, 107, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.J.; Abed, M.; Heuberger, J.; Novak, R.; Zohar, Y.; Beltran Lopez, A.P.; Trausch-Azar, J.S.; Ilagan, M.X.G.; Benhamou, D.; Dittmar, G.; et al. RNF4-Dependent Oncogene Activation by Protein Stabilization. Cell Rep. 2016, 16, 3388–3400. [Google Scholar] [CrossRef] [PubMed]
- Pierce, S.B. dMyc is required for larval growth and endoreplication in Drosophila. Development 2004, 131, 2317–2327. [Google Scholar] [CrossRef] [PubMed]
- Gallant, P.; Shiio, Y.; Cheng, P.F.; Parkhurst, S.M.; Eisenman, R.N. Myc and Max homologs in Drosophila. Science 1996, 274, 1523–1527. [Google Scholar] [CrossRef] [PubMed]
- Hammond, M.P.; Laird, C.D. Control of DNA replication and spatial distribution of defined DNA sequences in salivary gland cells of Drosophila melanogaster. Chromosoma 1985, 91, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Øvrebø, J.I.; Edgar, B.A. Polyploidy in tissue homeostasis and regeneration. Development 2018, 145, dev156034. [Google Scholar] [CrossRef] [PubMed]
- Shu, Z.; Row, S.; Deng, W.M. Endoreplication: The Good, the Bad, and the Ugly. Trends Cell Biol. 2018, 28, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zhou, Y.; Guo, C.; Gong, H.; Gong, Z.; Liu, L. Differential roles of the fan-shaped body and the ellipsoid body in Drosophila visual pattern memory. Learn. Mem. 2009, 16, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Zielke, N.; Kim, K.J.; Tran, V.; Shibutani, S.T.; Bravo, M.J.; Nagarajan, S.; Van Straaten, M.; Woods, B.; Von Dassow, G.; Rottig, C.; et al. Control of Drosophila endocycles by E2F and CRL4 CDT2. Nature 2011, 480, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Björklund, M.; Taipale, M.; Varjosalo, M.; Saharinen, J.; Lahdenperä, J.; Taipale, J. Identification of pathways regulating cell size and cell-cycle progression by RNAi. Nature 2006. [Google Scholar] [CrossRef] [PubMed]
- Edgar, B.A.; Zielke, N.; Gutierrez, C. Endocycles: A recurrent evolutionary innovation for post-mitotic cell growth. Nat. Rev. Mol. Cell Biol. 2014, 15, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Zielke, N.; Querings, S.; Rottig, C.; Lehner, C.; Sprenger, F. The anaphase-promoting complex/cyclosome (APC/C) is required for rereplication control in endoreplication cycles. Genes Dev. 2008, 22, 1690–1703. [Google Scholar] [CrossRef] [PubMed]
- Sakaue-Sawano, A.; Kurokawa, H.; Morimura, T.; Hanyu, A.; Hama, H.; Osawa, H.; Kashiwagi, S.; Fukami, K.; Miyata, T.; Miyoshi, H.; et al. Visualizing Spatiotemporal Dynamics of Multicellular Cell-Cycle Progression. Cell 2008, 132, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Zielke, N.; Edgar, B.A. FUCCI sensors: Powerful new tools for analysis of cell proliferation. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Arias, E.E.; Walter, J.C. PCNA functions as a molecular platform to trigger Cdt1 destruction and prevent re-replication. Nat. Cell Biol. 2006, 8, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, H.; Rahn, H.P.; Weinzierl, P.; Sporbert, A.; Cremer, T.; Zink, D.; Cardoso, M.C. Dynamics of DNA replication factories in living cells. J. Cell Biol. 2000, 149, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Zanet, J.; Pibre, S.; Jacquet, C.; Ramirez, A.; de Alborán, I.M.; Gandarillas, A. Endogenous Myc controls mammalian epidermal cell size, hyperproliferation, endoreplication and stem cell amplification. J. Cell Sci. 2005, 118, 1693–1704. [Google Scholar] [CrossRef] [PubMed]
- Orian, A.; Van Steensel, B.; Delrow, J.; Bussemaker, H.J.; Li, L.; Sawado, T.; Williams, E.; Loo, L.W.M.; Cowley, S.M.; Yost, C.; et al. Genomic binding by the Drosophila Myc, Max, Mad/Mnt transcription factor network. Genes Dev. 2003, 17, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Tavares, L.; Correia, A.; Santos, M.A.; Relvas, J.B.; Pereira, P.S. dMyc is required in retinal progenitors to prevent JNK-mediated retinal glial activation. PLoS Genet. 2017, 13. [Google Scholar] [CrossRef] [PubMed]
- Chittaranjan, S.; Xu, J.; Kuzyk, M.; Dullat, H.K.; Wilton, J.; DeVorkin, L.; Lebovitz, C.; Morin, G.B.; Marra, M.A.; Gorski, S.M. The Drosophila TIPE family member Sigmar interacts with the Ste20-like kinase Misshapen and modulates JNK signaling, cytoskeletal remodeling and autophagy. Biol. Open 2015, 4, 672–684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, C.; Wu, C.; Yang, Y.; Li, W.; Xue, L. The canonical Wg signaling modulates Bsk-mediated cell death in Drosophila. Cell Death Dis. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Huang, J.; Yang, L.; Yang, Y.; Li, W.; Xue, L. NOPO modulates Egr-induced JNK-independent cell death in Drosophila. Cell Res. 2012, 22, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Gowda, P.S.; Zhou, F.; Chadwell, L.V.; McEwen, D.G. p53 binding prevents phosphatase-mediated inactivation of diphosphorylated c-Jun N-terminal kinase. J. Biol. Chem. 2012, 287, 17554–17567. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.R.; Kow, E.; Nevis, K.R.; Lu, C.K.; Luce, K.S.; Zhong, Q.; Cook, J.G. Cdc6 Stability Is Regulated by the Huwe1 Ubiquitin Ligase after DNA Damage. Mol. Biol. Cell 2007. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.R.; Lee, H.O.; Bunker, B.D.; Dorn, E.S.; Rogers, G.C.; Duronio, R.J.; Cook, J.G. Cdt1 and Cdc6 are destabilized by rereplication-induced DNA damage. J. Biol. Chem. 2008, 283, 25356–25363. [Google Scholar] [CrossRef] [PubMed]
- Heidelberger, J.B.; Voigt, A.; Borisova, M.E.; Petrosino, G.; Ruf, S.; Wagner, S.A.; Beli, P. Proteomic profiling of VCP substrates links VCP to K6-linked ubiquitylation and c-Myc function. EMBO Rep. 2018, e44754. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, Y.; Eilers, M.; Lin, A. Miz1 is a signal- and pathway-specific modulator or regulator (SMOR) that suppresses TNF-alpha-induced JNK1 activation. Proc. Natl. Acad. Sci. USA 2009, 106, 18279–18284. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.K.; Yang, Y.; Chen, C.; Liu, J. Syk-mediated tyrosine phosphorylation of mule promotes TNF-induced JNK activation and cell death. Oncogene 2016, 35, 1988–1995. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.F.; Zhang, R.; Sun, J.; Li, Z.; Shin, D.M.; Wang, H.; Kovalchuk, A.L.; Sakai, T.; Xiong, H.; Kon, N.; et al. Homeostatic defects in B cells deficient in the E3 ubiquitin ligase ARF-BP1 are restored by enhanced expression of MYC. Leuk. Res. 2013, 37, 1680–1689. [Google Scholar] [CrossRef] [PubMed]
- Peter, S.; Bultinck, J.; Myant, K.; Jaenicke, L.A.; Walz, S.; Muller, J.; Gmachl, M.; Treu, M.; Boehmelt, G.; Ade, C.P.; et al. Tumor cell-specific inhibition of MYC function using small molecule inhibitors of the HUWE1 ubiquitin ligase. EMBO Mol. Med. 2014, 6, 1525–1541. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanku, Y.; Bitman-Lotan, E.; Zohar, Y.; Kurant, E.; Zilke, N.; Eilers, M.; Orian, A. Drosophila HUWE1 Ubiquitin Ligase Regulates Endoreplication and Antagonizes JNK Signaling During Salivary Gland Development. Cells 2018, 7, 151. https://doi.org/10.3390/cells7100151
Yanku Y, Bitman-Lotan E, Zohar Y, Kurant E, Zilke N, Eilers M, Orian A. Drosophila HUWE1 Ubiquitin Ligase Regulates Endoreplication and Antagonizes JNK Signaling During Salivary Gland Development. Cells. 2018; 7(10):151. https://doi.org/10.3390/cells7100151
Chicago/Turabian StyleYanku, Yifat, Eliya Bitman-Lotan, Yaniv Zohar, Estee Kurant, Norman Zilke, Martin Eilers, and Amir Orian. 2018. "Drosophila HUWE1 Ubiquitin Ligase Regulates Endoreplication and Antagonizes JNK Signaling During Salivary Gland Development" Cells 7, no. 10: 151. https://doi.org/10.3390/cells7100151
APA StyleYanku, Y., Bitman-Lotan, E., Zohar, Y., Kurant, E., Zilke, N., Eilers, M., & Orian, A. (2018). Drosophila HUWE1 Ubiquitin Ligase Regulates Endoreplication and Antagonizes JNK Signaling During Salivary Gland Development. Cells, 7(10), 151. https://doi.org/10.3390/cells7100151