
Neuro-Inflammatory and Behavioral Changes Are Selectively Reversed by Sceletium tortuosum (Zembrin®) and Mesembrine in Male Rats Subjected to Unpredictable Chronic Mild Stress


Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Materials and Chemical Profiling
2.2. Animals
2.3. Drug Treatment
2.4. Study Layout
2.5. The UCMS Protocol
2.6. Behavioral Tests
2.6.1. Sucrose Preference Test (SPT)
2.6.2. Elevated Plus Maze (EPM)
2.6.3. Barnes Maze (BM)
2.6.4. Open Field Test (OFT)
2.6.5. Forced Swim Test (FST)
2.7. Neurochemical Measures
2.7.1. Sample Collection and Preparation
2.7.2. Phosphodiesterase 4B, IL-10, and TNF-α
2.7.3. Monoamines
2.7.4. Tyrosine, 3-Chlorotyrosine, GSH, and GSSG
2.8. Statistical Analyses
3. Results
3.1. Validation of the Model
3.1.1. Effects of UCMS on Behavior (Figure 2; Table A1 in Appendix A.1)
3.1.2. Effects of UCMS on Neurochemical Markers (Figure 3; Table A2 in Appendix A.1)

3.2. Treatment Response and Influence of Sex
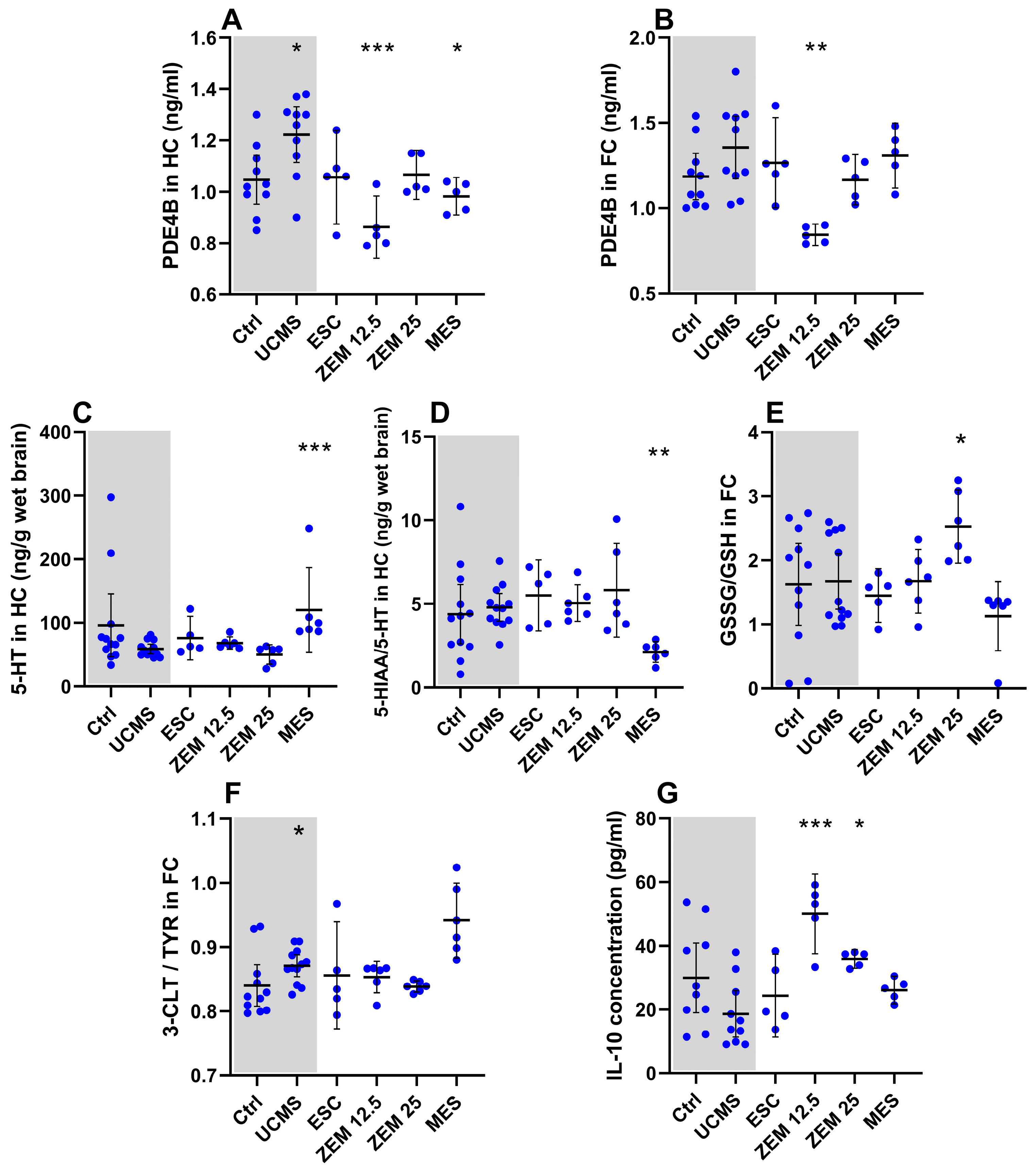
3.2.1. Treatment Effects on Behavior (Figure 4; Table A3 in Appendix A)

3.2.2. Treatment Effects on Neurochemical Markers (Figure 5; Table A4 in Appendix A)
4. Discussion
5. Conclusions and Unifying Hypothesis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3-CLT | 3-chlorotyrosine |
| 5-HIAA | 5-hydroxyindoleacetic acid |
| 5-HT | 5-hydroxytryptamine |
| BDNF | Brain-derived neurotrophic factor |
| cAMP | Cyclic adenosine monophosphate |
| CoV | Coefficient of variance |
| Ctrl | Control |
| DA | Dopamine |
| DOPAC | 3,4-Dihydroxyphenylacetic acid |
| ELISA | Enzyme-linked immunosorbent assay |
| ESC | Escitalopram |
| FC | Frontal cortex |
| GABA | Gamma-aminobutyric acid |
| HC | Hippocampus |
| HPA axis | Hypothalamic–pituitary–adrenal axis |
| IFN-γ | Interferon-gamma |
| HPLC-ECD | High-Performance Liquid Chromatography with Electrochemical Detection |
| MES | Mesembrine |
| NWU | North-West University |
| OFT | Open field test |
| PDE4 | Phosphodiesterase 4 |
| PND | Postnatal day |
| SPT | Sucrose preference test |
| ST | Sceletium tortuosum |
| TYR | Tyrosine |
| TNF-α | Tumor necrosis factor-alpha |
| UCMS | Unpredictable chronic mild stress |
| ZEM | Zembrin® |
Appendix A
Appendix A.1. Summary of the Effects of UCMS on Males, Females, and Both Sexes Combined

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combined (Males + Females) | Males | Females | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | Ctrl vs. UCMS mean ± SD (n) | p | d [95% CI] | Ctrl vs. UCMS mean ± SD (n) | p | d [95% CI] | Ctrl vs. UCMS mean ± SD (n) | p | d [95% CI] |
| Sucrose Preference Test (SPT) | |||||||||
| Sucrose preference PND50 | 0.75 ± 0.14 (24O) 0.69 ± 0.23 (24) | 0.8983 | 0.34 [−0.91; 0.23] | 0.79 ± 0.09 (12O) 0.66 ± 0.25 (12) | 0.4681 | 0.7 [−1.54; 0.11] | 0.71 ± 0.16 (12O) 0.72 ± 0.22 (12) | 0.2829 | 0.02 [−0.78; 0.82] |
| Sucrose preference PND64 | 0.79 ± 0.14 (24) 0.74 ± 0.19 (24) | 0.7938 | 0.26 [−0.84; 0.30] | 0.82 ± 0.14 (12O) 0.65 ± 0.22 (12) | 0.0265 | 0.89 [−1.76; −0.07] | 0.76 ± 0.14 (12) 0.84 ± 0.10 (12O) | 0.0490 | 0.62 [−0.19; 1.46] |
| Sucrose preference PND78 | 0.82 ± 0.13 (24O) 0.77 ± 0.15 (24O) | 0.0618 | 0.35 [−0.93; 0.21] | 0.87 ± 0.04 (12) 0.71 ± 0.18 (12O) | 0.0013 | 1.16 [−2.07; −0.32] | 0.78 ± 0.17 (12) 0.84 ± 0.09 (12O) | 0.8301 | 0.39 [−0.41; 1.21] |
| Sucrose preference PND92 | 0.77 ± 0.19 (24) 0.73 ± 0.23 (24) | 0.4005 | 0.18 [−0.75; 0.39] | 0.82 ± 0.12 (12O) 0.62 ± 0.30 (12) | 0.0996 | 0.87 [−1.75; −0.03] | 0.72 ± 0.23 (12) 0.83 ± 0.05 (12) | 0.6596 | 0.66 [−0.15; 1.5] |
| Sucrose preference PND101 | 0.84 ± 0.12 (24O) 0.72 ± 0.27 (24) | 0.2387 | 0.56 [−1.14; 0.01] | 0.83 ± 0.13 (12O) 0.59 ± 0.32 (12) | 0.1292 | 0.96 [−1.84; −0.13] | 0.86 ± 0.11 (12O) 0.86 ± 0.07 (12) | 0.7870 | 0.06 [−0.74; 0.86] |
| Forced Swim Test (FST) | |||||||||
| Swimming (s) | 48.44 ± 29.65 (24) 91.47 ± 46.33 (24) | 0.0003 | 1.09 [0.49; 1.71] | 38.90 ± 20.76 (12O) 94.68 ± 55.64 (12) | 0.0011 | 1.28 [0.43; 2.21] | 57.98 ± 34.76 (12) 88.26 ± 37.02 (12) | 0.0684 | 0.81 [0.01; 1.67] |
| Immobility (s) | 242.5 ± 29.27 (24) 196.3 ± 46.28 (24) | 0.0001 | 1.17 [−1.8; −0.57] | 248.8 ± 22.83 (12O) 198.0 ± 53.11 (12) | 0.0056 | 1.2 [−2.11; −0.35] | 236.1 ± 34.38 (12) 194.6 ± 40.64 (12) | 0.0068 | 1.06 [−1.95; −0.23] |
| Struggling (s) | 9.47 ± 7.22 (24) 12.30 ± 12.71 (24O) | 0.9268 | 0.27 [−0.3; 0.84] | 12.65 ± 7.60 (12) 7.25 ± 8.71 (12) | 0.0332 | 0.64 [−1.48; 0.17] | 6.28 ± 5.40 (12) 17.34 ± 14.37 (12O) | 0.0688 | 0.98 [0.13; 1.88] |
| Open Field Test (OFT) | |||||||||
| Distance (cm) | 7652 ± 2105 (24) 9549 ± 1235 (24) | 0.0014 | 1.08 [0.49; 1.70] | 7376 ± 1990 (12) 9965 ± 1105 (12) | 0.0007 | 1.55 [0.67; 2.52] | 7928 ± 2267 (12) 9132 ± 1261 (12) | 0.2657 | 0.63 [−0.17; 1.47] |
| Center duration (s) | 69.89 ± 25.54 (24) 62.58 ± 14.71 (24) | 0.4188 | 0.35 [−0.92; 0.222] | 69.31 ± 28.76 (12) 64.92 ± 12.21 (12) | 0.6707 | 0.19 [−1.0; 0.61] | 70.46 ± 23.17 (12O) 60.23 ± 17.07 (12) | 0.4095 | 0.49 [−1.31; 0.32] |
| Corner duration (s) | 70.76 ± 22.43 (24) 73.27 ± 16.60 (24) | >0.9999 | 0.13 [−0.44; 0.69] | 65.02 ± 22.70 (12O) 61.60 ± 8.29 (12) | 0.5512 | 0.19 [−1.0; 0.6] | 76.51 ± 21.57 (12) 84.93 ± 14.51 (12) | 0.4428 | 0.44 [−0.36; 1.27] |
| Elevated Plus Maze (EPM) | |||||||||
| Distance (cm) | 1691 ± 311.7 (24) 1797 ± 235.1 (24) | 0.2550 | 1.08 [0.49; 1.7] | 1684 ± 347.6 (12) 1763 ± 202.1 (12) | 0.7125 | 0.27 [−0.53; 1.08] | 1697 ± 286.8 (12) 1832 ± 268.6 (12) | 0.2415 | 0.47 [−0.34; 1.29] |
| Open/closed arm duration | 1.83 ± 1.37 (24) 1.59 ± 0.56 (24) | 0.8461 | 0.23 [−0.8; 0.34] | 1.54 ± 1.17 (12) 1.62 ± 0.65 (12) | 0.3777 | 0.08 [−0.72; 0.88] | 2.11 ± 1.53 (12O) 1.56 ± 0.47 (12) | 0.6707 | 0.48 [−1.3; 0.33] |
| Barnes Maze (BM) | |||||||||
| Distance (c) | 1294 ± 250.6 (24) 1321 ± 295.7 (24O) | 0.9919 | 0.1 [−0.47; 0.66] | 1239 ± 186.1 (12) 1299 ± 164.0 (12) | 0.7125 | 0.33 [−0.47; 1.14] | 1349 ± 300.2 (12) 1343 ± 393.6 (12O) | 0.8428 | 0.02 [−0.82; 0.78] |
| Primary latency (s) | 40.33 ± 29.59 (24) 16.7% not found in time 31.67 ± 31.09 (24) 12.5% not found in time | 0.1508 | 0.32 [−0.89; 0.25] | 38.58 ± 29.46 (12) 8.3% not found in time 29.17 ± 30.99 (12) 16.7% not found in time | 0.3541 | 0.3 [−1.11; 0.5] | 26.11 ± 12.94 (12) 25% not found in time 29.09 ± 28.48 (12) 8.3% not found in time | 0.5637 | 0.13 [−0.67; 0.93] |
| Primary error rate | 9.55 ± 6.74 (20) 16.7% no errors 5.84 ± 5.73 (19O) 20.8% no errors | 0.0677 | 0.58 [−1.23; 0.05] | 11.33 ± 7.69 (12) 0% no errors 3.89 ± 3.41 (12) 25% no errors | 0.0072 | 1.14 [−2.12; −0.23] | 6.88 ± 4.09 (12) 16.7% no errors 7.60 ± 6.93 (12) 16.7% no errors | 0.9127 | 0.12 [−0.81; 1.05] |
| Edge exploration (head dips) | 13.04 ± 4.14 (24) 11.38 ± 3.70 (24) | 0.1815 | 0.42 [−1; 0.15] | 11.25 ± 3.25 (12) 12.58 ± 3.99 (12) | 0.4331 | 0.35 [−0.45; 1.17] | 14.83 ± 4.28 (12) 10.17 ± 3.1 (12) | 0.0054 | 1.01 [−2.12; −0.36] |
| Probe zone duration (s) | 29.57 ± 9.43 (24) 30.33 ± 13.51 (24) | 0.9593 | 0.06 [−0.5; 0.63] | 29.80 ± 11.32 (12) 34.32 ± 12.82 (12) | 0.3777 | 0.36 [−0.44; 1.18] | 29.35 ± 7.6 (12) 26.35 ± 13.50 (12) | 0.4428 | 0.26 [−1.08; 0.534] |
| Probe/Not probe duration | 0.53 ± 0.27 (24) 0.59 ± 0.43 (24O) | 0.9878 | 0.19 [−0.38; 0.76] | 0.55 ± 0.33 (12) 0.71 ± 0.47 (12O) | 0.4013 | 0.38 [−0.42; 1.2] | 0.50 ± 0.19 (12) 0.48 ± 0.37 (12) | 0.4010 | 0.08 [−0.89; 0.72] |
| Outer/inner duration | 59.74 ± 57.23 (24) 49.93 ± 113.3 (24O) | 0.0093 | 0.11 [−0.68; 0.46] | 49.50 ± 63.40 (12) 10.58 ± 17.17 (12O) | 0.0081 | 0.81 [−1.67; 0.01] | 69.99 ± 50.98 89.29 ± 152.2 | 0.1978 | 0.16 [−0.63; 0.97] |
| Combined (Males + Females) | Male Rats | Female Rats | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | Ctrl vs. UCMS mean ± SD (n) | p | d [95% CI] | Ctrl vs. UCMS mean ± SD (n) | p | d [95% CI] | Ctrl vs. UCMS mean ± SD (n) | p | d [95% CI] |
| Frontal Cortex Analyses | |||||||||
| PDE4B (ng/mL) | 1.22 ± 0.21 (20) 1.29 ± 0.22 (20) | 0.4169 | 0.29 [−0.33; 0.92] | 1.19 ± 0.19 (10) 1.36 ± 0.26 (10) | 0.1091 | 0.72 [−0.17; 1.66] | 1.26 ± 0.24 (10) 1.22 ± 0.17 (10) | 0.4926 | 0.19 [−1.07; 0.69] |
| 5-HT (ng/g) | 151.60 ± 69.46 (24O) 119.60 ± 37.24 (23R) | 0.1402 | 0.27 [−0.295; 0.84] | 168.70 ± 80.50 (11R) 110.60 ± 32.61 (11R) | 0.0557 | 0.94 [−1.86; −0.08] | 134.60 ± 54.56 (12) 127.80 ± 40.66 (12) | 0.9774 | 0.14 [−0.94; 0.66] |
| 5-HIAA (ng/g) | 243.80 ± 53.90 (24) 215.70 ± 11.00 (24) | 0.0930 | 0.51 [−1.087; 0.06] | 253.40 ± 63.11 (12) 224.80 ± 58.09 (12) | 0.4095 | 0.46 [−1.28; 0.35] | 234.30 ± 43.50 (12) 206.60 ± 53.59 (12O) | 0.1600 | 0.55 [−1.38; 0.26] |
| 5HIAA/5-HT | 1.89 ± 0.80 (24) 1.54 ± 0.82 (24) | 0.2818 | 0.42 [−1.00; 0.15] | 1.75 ± 0.71 (12) 1.92 ± 0.77 (12O) | 0.5137 | 0.22 [−0.58; 1.02] | 2.02 ± 0.91 (12) 1.16 ± 0.70 (12) | 0.0387 | 0.02 [−0.82; 0.78] |
| DA (ng/g) | 183.80 ± 102.40 (24O) 216.0 ± 162.40 (24) | 0.8461 | 0.23 [−0.332; 0.80] | 144.60 ± 55.22 (12) 252.60 ± 187.80 (12) | 0.1432 | 0.75 [−0.06; 1.61] | 223.00 ± 124.6 (12O) 179.30 ± 130.20 (12) | 0.3474 | 0.33 [−1.15; 0.47] |
| DOPAC (ng/g) | 121.00 ± 38.86 (24O) 123.60 ± 64.45 (24O) | 0.8143 | 0.05 [−0.519; 0.61] | 105.70 ± 30.20 (12) 132.80 ± 30.97 (11R) | 0.0595 | 0.9 [0.02; 1.74] | 136.40 ± 41.60 (12) 94.60 ± 35.93 (12) | 0.0036 | 1.04 [−1.92; −0.21] |
| DOPAC/DA | 0.75 ± 0.28 (24) 0.73 ± 0.41 (23R) | 0.5336 | 0.27 [−0.296; 0.84] | 0.78 ± 0.21 (12) 0.74 ± 0.47 (11R) | 0.4865 | 0.38 [−0.42; 1.2] | 0.73 ± 0.34 (12) 0.72 ± 0.37 (12) | 0.9323 | 0.02 [−0.82; 0.78] |
| NA (ng/g) | 3576 ± 1759 (24) 4637 ± 2215 (23R) | 0.1714 | 0.41 [−0.158; 0.99] | 3544 ± 2105 (12) 4707 ± 2262 (11R) | 0.2351 | 0.51 [−0.31; 1.36] | 3607 ± 1428 (12) 4574 ± 2270 (12) | 0.4428 | 0.49 [−0.31; 1.32] |
| TYR (ng/g) | 103.7 ± 33.12 (24O) 106.1 ± 38.51 (24O) | 0.9268 | 0.07 [−0.5; 0.63] | 103.8 ± 42.07 (12O) 103.2 ± 28.10 (12O) | 0.7125 | 0.02 [−0.82; 0.78] | 103.7 ± 22.89 (12) 109.1 ± 47.88 (12) | 0.9323 | 0.14 [−0.66; 0.94] |
| 3-CLT (ng/g) | 85.20 ± 21.99 (24) 93.17 ± 32.13 (24O) | 0.5464 | 0.29 [−0.28; 0.86] | 83.22 ± 26.66 (12) 89.97 ± 24.91 (12) | 0.3186 | 0.25 [−0.55; 1.06] | 87.19 ± 17.09 (12) 96.36 ± 38.93 (12) | 0.9774 | 0.3 [−0.5; 1.11] |
| 3-CLT/TYR | 0.84 ± 0.05 (23R) 0.88 ± 0.06 (24) | 0.0028 | 0.77 [0.19; 1.38] | 0.84 ± 0.05 (11R) 0.87 ± 0.03 (12) | 0.0317 | 0.77 [−0.06; 1.64] | 0.85 ± 0.05 (12) 0.90 ± 0.08 (12) | 0.0597 | 0.79 [−0.03; 1.65] |
| GSH (ng/g) | 73.90 ± 19.16 (24) 80.96 ± 25.05 (24) | 0.2465 | 0.31 [−0.26; 0.88] | 74.34 ± 19.23 (12O) 85.32 ± 30.01 (12) | 0.6707 | 0.42 [−0.38; 1.24] | 73.46 ± 19.94 (12) 76.60 ± 19.25 (12O) | 0.4776 | 0.16 [−0.64; 0.96] |
| GSSG (ng/g) | 143.1 ± 67.47 (23R) 142.7 ± 71.47 (24O) | 0.5761 | 0.01 [−0.58; 0.57] | 116.9 ± 61.18 (11R) 124.7 ± 15.63 (12O) | 0.9279 | 0.17 [−0.64; 1.0] | 167.0 ± 66.25 (12) 160.7 ± 98.64 (12) | 0.4428 | 0.07 [−0.88; 0.73] |
| GSSG/GSH | 2.078 ± 1.171 (23R) 1.900 ± 0.9881 (24O) | 0.5618 | 0.16 [−0.74; 0.41] | 1.63 ± 0.95 (11R) 1.67 ± 0.68 (12) | 0.8801 | 0.05 [−0.76; 0.87] | 2.49 ± 1.24 (12) 2.13 ± 1.21 (12) | 0.5512 | 0.29 [−1.1; 0.51] |
| Hippocampus Analyses | |||||||||
| PDE4B (ng/mL) | 1.11 ± 0.14 (20) 1.21 ± 0.17 (20) | 0.0566 | 0.60 [−0.02; 1.25] | 1.05 ± 0.13 (10) 1.22 ± 0.15 (10) | 0.0108 | 1.18 [0.26; 2.18] | 1.18 ± 0.12 (10) 1.19 ± 0.19 (10) | 0.9560 | 0.08 [−0.8; 0.96] |
| 5-HT (ng/g) | 93.41 ± 58.27 (24O) 90.25 ± 81.45 (24O) | 0.3007 | 0.04 [−0.61; 0.52] | 96.19 ± 77.65 (12O) 58.98 ± 12.02 (12) | 0.1600 | 0.65 [−1.49; 0.16] | 90.63 ± 32.45 (12) 121.50 ± 107.7 (12O) | 0.8874 | 0.38 [−0.424; 1.19] |
| 5-HIAA (ng/g) | 302.80 ± 78.43 (23R) 279.50 ± 60.61 (24) | 0.3256 | 0.33 [−0.91; 0.24] | 294.60 ± 96.93 (12) 272.90 ± 51.75 (12) | 0.7125 | 0.27 [−1.08; 0.53] | 311.80 ± 55.02 (11R) 286.10 ± 70.06 (12) | 0.3470 | 0.39 [−1.228; 0.43] |
| 5HIAA/5-HT | 4.83 ± 4.28 (24O) 3.97 ± 1.49 (24) | >0.9999 | 0.27 [−0.84; 0.3] | 4.38 ± 2.79 (12) 4.80 ± 1.30 (12) | 0.4095 | 0.18 [−0.62; 1.0] | 5.28 ± 5.49 (11R) 3.14 ± 1.20 (12) | 0.5658 | 0.43 [−1.265; 0.39] |
| DA (ng/g) | 74.27 ± 31.17 (23R) 67.02 ± 26.81 (24) | 0.5062 | 0.25 [−0.82; 0.33] | 73.75 ± 35.24 (11R) 66.56 ± 17.69 (12) | 0.9279 | 0.25 [−1.08; 0.56] | 74.75 ± 28.53 (12) 67.48 ± 34.49 (12O) | 0.2913 | 0.22 [−1.03; 0.58] |
| DOPAC (ng/g) | 92.12 ± 56.42 (23R) 85.95 ± 31.55 (24O) | 0.9580 | 0.31 [−0.89; 0.25] | 76.97 ± 20.23 (11R) 88.42 ± 28.44 (12O) | 0.3164 | 0.44 [−0.38; 1.29] | 106.00 ± 74.52 (12O) 83.48 ± 35.49 (12O) | 0.4776 | 0.37 [−1.19; 0.43] |
| DOPAC/DA | 1.13 ± 0.26 (23R) 1.33 ± 0.38 (23R) | 0.1483 | 0.54 [−0.04; 1.14] | 1.09 ± 0.29 (12) 1.39 ± 0.51 (11R) | 0.21 | 0.56 [−0.26; 1.41] | 1.17 ± 0.23 (11R) 1.28 ± 0.20 (12) | 0.4134 | 0.47 [−0.35; 1.313] |
| NA (ng/g) | 8096 ± 12,356 (24O) 3634 ± 1895 (24O) | 0.3007 | 0.41 [−0.992; 0.164] | 3473 ± 1375 (12) 3581 ± 1188 (12) | 0.7553 | 0.08 [−0.72; 0.88] | 12,719 ± 16,453 (12) 3687 ± 2469 (12) | 0.1277 | 0.741 [−1.591; 0.072] |
| TYR (ng/g) | 139.7 ± 39.07 (24) 131.7 ± 47.17 (24O) | 0.4188 | 0.18 [−0.75; 0.38] | 129.6 ± 30.08 (12) 125.8 ± 23.92 (12) | 0.9774 | 0.14 [−0.94; 0.66] | 149.8 ± 45.43 (12) 137.6 ± 63.28 (12O) | 0.3186 | 0.21 [−0.02; 0.58] |
| 3-CLT (ng/g) | 120.8 ± 38.29 (168 BD) 110.2 ± 55.77 (1113 BD) | 0.1945 | 0.22 [−0.79; 0.346] | 108.3 ± 29.64 (915BD) 83.40 ± 10.43 (222BD) | 0.3273 | 1.08 [−1.98; −0.24] | 137.0 ± 44.21 (75BD) 116.1 ± 60.46 (93BD) | 0.2105 | 0.38 [−1.2; 0.42] |
| 3-CLT/TYR | 0.81 ± 0.02 (168 BD) 0.84 ± 0.04 (1113 BD) | 0.2122 | 0.7 [0.125; 1.293] | 0.81 ± 0.02 (93BD) 0.82 ± 0.001 (210BD) | 0.7273 | 0.57 [−0.23; 1.41] | 0.81 ± 0.02 (75BD) 0.84 ± 0.04 (93BD) | 0.2523 | 0.71 [−0.1; 1.56] |
| GSH (ng/g) | 80.72 ± 15.39 (24) 81.91 ± 20.76 (24) | 0.6165 | 0.06 [−0.5; 0.63] | 79.83 ± 16.38 (12O) 72.41 ± 21.14 (12) | 0.4428 | 0.38 [−1.2; 0.42] | 81.60 ± 15.00 (12) 91.40 ± 16.05 (12) | 0.1600 | 0.61 [−0.2; 1.45] |
| GSSG (ng/g) | 192.2 ± 63.97 (24) 187.5 ± 71.39 (23R) | 0.5336 | 0.07 [−0.64; 0.5] | 179.7 ± 63.77 (12) 161.3 ± 41.10 (12) | 0.4095 | 0.33 [−1.15; 0.47] | 204.7 ± 64.43 (12) 216.0 ± 87.39 (11R) | 0.8801 | 0.14 [−0.67; 0.97] |
| GSSG/GSH | 2.421 ± 0.7473 (24) 2.578 ± 1.130 (24) | 0.5740 | 0.16 [−0.4; 0.73] | 2.34 ± 0.87 (12) 2.48 ± 1.11 (12) | 0.7125 | 0.13 [−0.67; 0.94] | 2.5 ± 0.62 (12) 2.68 ± 1.19 (12) | 0.6297 | 0.18 [−0.62; 0.99] |
| Plasma Analyses | |||||||||
| IL-10 (pg/g) | 23.26 ± 14.37 (20) 17.81 ± 8.683 (20) | 0.2504 | 0.45 [−1.08; 0.17] | 29.97 ± 15.26 (10) 18.66 ± 10.22 (10) | 0.0612 | 0.83 [−1.78; 0.06] | 16.54 ± 10.15 (10) 16.95 ± 7.29 (10) | 0.8098 | 0.04 [−0.83; 0.92] |
| TNF-α (pg/g) | 8.220 ± 4.125 (20) 7.802 ± 3.301 (20) | 0.9411 | 0.11 [−0.73; 0.51] | 7.86 ± 3.28 (10) 8.14 ± 3.30 (10) | 0.9292 | 0.08 [−0.79; 0.96] | 8.58 ± 4.99 (10) 7.46 ± 3.44 (10) | 0.8688 | −0.25 [−1.14; 0.62] |
Appendix A.2. The Effects of Treatment on UCMS-Exposed Male Rats
| Parameter | Kruskal–Wallis Test | Rx Group | n | Mean ± SD | p-Value | d-Value [95% CI] |
|---|---|---|---|---|---|---|
| Sucrose Preference Test (SPT) | ||||||
| Sucrose preference PND50 | X2(4) = 6.265, p = 0.18 | UCMS | 12 | 0.66 ± 0.25 | ||
| ESC | 5 | 0.58 ± 0.22 | >0.9999 | 0.30 [−1.36; 0.74] | ||
| ZEM12.5 | 6 | 0.68 ± 0.16 | >0.9999 | 0.07 [−0.91; 1.05] | ||
| ZEM25 | 6O | 0.80 ± 0.19 | 0.6165 | 0.57 [−0.41; 1.59] | ||
| MES | 6O | 0.76 ± 0.21 | >0.9999 | 0.41 [−0.57; 1.42] | ||
| Sucrose preference PND64 | X2(4) = 13.29, p = 0.0099 | UCMS | 12 | 0.65 ± 0.22 | ||
| ESC | 5O | 0.81 ± 0.21 | 0.0629 | 0.69 [−0.36; 1.79] | ||
| ZEM12.5 | 6 | 0.80 ± 0.21 | 0.8382 | 0.71 [−0.28; 1.75] | ||
| ZEM25 | 6 | 0.77 ± 0.12 | >0.9999 | 0.59 [−0.4; 1.51] | ||
| MES | 6 | 0.89 ± 0.03 | 0.0056 | 1.25 [0.22; 2.37] | ||
| Sucrose preference PND78 | X2(4) = 17.86, p = 0.0013 | UCMS | 12O | 0.71 ± 0.18 | ||
| ESC | 5 | 0.72 ± 0.14 | >0.9999 | 0.05 [−1.00; 1.09] | ||
| ZEM12.5 | 6 | 0.85 ± 0.05 | 0.1991 | 0.88 [−0.12; 1.94] | ||
| ZEM25 | 6 | 0.83 ± 0.05 | 0.5118 | 0.75 [−0.24; 1.79] | ||
| MES | 6 | 0.92 ± 0.03 | 0.0003 | 1.32 [0.28; 2.46] | ||
| Sucrose preference PND92 | X2(4) = 3.834, p = 0.429 | UCMS | 11 | 0.62 ± 0.30 | ||
| ESC | 5O | 0.75 ± 0.29 | 0.6615 | 0.43 [−0.62; 1.52] | ||
| ZEM12.5 | 6 | 0.82 ± 0.11 | 0.5338 | 0.77 [−0.24; 1.83] | ||
| ZEM25 | 6 | 0.78 ± 0.12 | >0.9999 | 0.59 [−0.409; 1.629] | ||
| MES | 6 | 0.80 ± 0.16 | 0.6057 | 0.67 [−0.331; 1.720] | ||
| Sucrose preference PND101 | X2(4) = 8.898, p = 0.0637 | UCMS | 12 | 0.59 ± 0.32 | ||
| ESC | 5O | 0.83 ± 0.14 | 0.5689 | 0.82 [−0.234; 1.941] | ||
| ZEM12.5 | 6 | 0.90 ± 0.04 | 0.0230 | 1.13 [0.107; 2.230] | ||
| ZEM25 | 6 | 0.82 ± 0.05 | >0.9999 | 0.84 [−0.156; 1.895] | ||
| MES | 6 | 0.72 ± 0.25 | >0.9999 | 0.43 [−0.547; 1.437] | ||
| Forced Swim Test (FST) | ||||||
| Swimming (s) | X2(4) = 11.55, p = 0.0211 | UCMS | 12 | 94.68 ± 55.64 | ||
| ESC | 5 | 52.23 ± 22.62 | 0.3284 | 0.82 [−1.94; 0.24] | ||
| ZEM12.5 | 6O | 41.13 ± 32.36 | 0.0615 | 1.03 [−2.11; −0.02] | ||
| ZEM25 | 6 | 121.6 ± 54.92 | >0.9999 | 0.46 [−0.52; 1.47] | ||
| MES | 6 | 75.99 ± 26.69 | >0.9999 | 0.37 [−1.37; 0.61] | ||
| Immobility (s) | X2(4) = 8.44. p = 0.0766 | UCMS | 12 | 198.0 ± 53.11 | ||
| ESC | 5 | 232.9 ± 33.14 | 0.7974 | 0.68 [−0.37; 1.78] | ||
| ZEM12.5 | 6 | 249.4 ± 34.34 | 0.1748 | 1.02 [0.01; 2.1] | ||
| ZEM25 | 6 | 171.4 ± 55.71 | >0.9999 | 0.469 [−1.48; 0.51] | ||
| MES | 6 | 200.3 ± 41.79 | >0.9999 | 0.04 [−0.94; 1.03] | ||
| Struggling (s) | X2(4) = 3.82, p = 0.4316 | UCMS | 12 | 7.25 ± 8.71 | ||
| ESC | 5 | 14.14 ± 13.55 | >0.9999 | 0.64 [−0.41; 1.73] | ||
| ZEM12.5 | 6 | 9.72 ± 13.39 | >0.9999 | 0.23 [−0.75; 1.22] | ||
| ZEM25 | 6 | 7.45 ± 4.46 | >0.9999 | 0.03 [−0.95; 1.01] | ||
| MES | 6 | 23.80 ± 22.41 | 0.2640 | 1.09 [0.07; 2.19] | ||
| Open Field Test (OFT) | ||||||
| Distance (cm) | X2(4) = 6.025, p = 0.1973 | UCMS | 12 | 9965 ± 1105 | ||
| ESC | 5 | 8885 ± 1461 | 0.2857 | 0.85 [−1.97; 0.21] | ||
| ZEM12.5 | 6 | 8834 ± 837 | 0.2740 | 1.05 [−2.14; −0.03] | ||
| ZEM25 | 6O | 9249 ± 1397 | 0.3884 | 0.57 [−1.59; 0.42] | ||
| MES | 6 | 8643 ± 1537 | 0.4737 | 1.00 [−2.08; 0.00] | ||
| Center duration (s) | X2(4) = 8.07, p = 0.0889 | UCMS | 12 | 64.92 ± 12.21 | ||
| ESC | 5 | 46.54 ± 7.36 | 0.1686 | 1.57 [−2.83; −0.44] | ||
| ZEM12.5 | 6 | 54.88 ± 22.65 | 0.9404 | 0.59 [−1.61; 0.39] | ||
| ZEM25 | 6 | 77.52 ± 25.16 | >0.9999 | 0.69 [−0.3; 1.73] | ||
| MES | 6 | 60.67 ± 16.46 | >0.9999 | 0.3 [−1.29; 0.68] | ||
| Corner duration (s) | X2(4) = 7.42, p = 0.1152 | UCMS | 12 | 61.60 ± 8.29 | ||
| ESC | 5 | 86.81 ± 20.98 | 0.0444 | 1.85 [0.67; 3.18] | ||
| ZEM12.5 | 6O | 74.73 ± 24.48 | 0.8900 | 0.82 [−0.18; 1.87] | ||
| ZEM25 | 6 | 69.73 ± 22.79 | >0.9999 | 0.54 [−0.45; 1.55] | ||
| MES | 6 | 73.93 ± 21.82 | 0.3755 | 0.84 [−0.16; 1.89] | ||
| Elevated Plus Maze (EPM) | ||||||
| Distance (cm) | X2(4) = 3.17, p = 0.5299 | UCMS | 12 | 1763 ± 202.1 | ||
| ESC | 5 | 1899 ± 275.7 | 0.6652 | 0.58 [−0.47; 1.66] | ||
| ZEM12.5 | 6 | 1780 ± 374.4 | >0.9999 | 0.06 [−0.92; 1.04] | ||
| ZEM25 | 6 | 1908 ± 398.4 | 0.7081 | 0.5 [−0.48; 1.51] | ||
| MES | 6 | 1817 ± 111.4 | >0.9999 | 0.29 [−0.69; 1.28] | ||
| Open/closed arm duration | X2(4) = 5.56, p = 0.2349 | UCMS | 12 | 1.62 ± 0.65 | ||
| ESC | 5 | 1.27 ± 0.64 | >0.9999 | 0.51 [−1.59; 0.53] | ||
| ZEM12.5 | 6 | 2.35 ± 1.02 | 0.3388 | 0.89 [−0.11; 1.95] | ||
| ZEM25 | 6 | 2.24 ± 1.49 | >0.9999 | 0.59 [−0.39; 1.62] | ||
| MES | 6 | 1.7 ± 0.45 | >0.9999 | 0.12 [−0.85; 1.11] | ||
| Barnes Maze (BM) | ||||||
| Distance (s) | X2(4) = 6.43, p = 0.1696 | UCMS | 12 | 1299 ± 164.0 | ||
| ESC | 5 | 1112 ± 237.6 | 0.5969 | 0.95 [−2.09; 0.12] | ||
| ZEM12.5 | 6 | 990.1 ± 159.6 | 0.0228 | 1.81 [−3.06; −0.7] | ||
| ZEM25 | 6 | 1395 ± 118.5 | 0.9663 | 0.61 [−0.38; 1.63] | ||
| MES | 6 | 1267 ± 235.4 | >0.9999 | 0.16 [−1.15; 0.82] | ||
| Primary latency (s) | X2(4) = 2.746, p = 0.6011 | UCMS | 12 | 29.17 ± 30.99 | ||
| ESC | 5 | 43.00 ± 29.10 | 0.7237 | 0.43 [−0.61; 1.5] | ||
| ZEM12.5 | 6 | 42.67 ± 32.15 | >0.9999 | 0.41 [−0.57; 1.42] | ||
| ZEM25 | 6O | 22.50 ± 17.76 | >0.9999 | 0.23 [−1.22; 0.75] | ||
| MES | 6 | 40.17 ± 39.18 | >0.9999 | 0.31 [−0.67; 1.31] | ||
| Primary error rate | X2(4) = 6.9, p = 0.1414 | UCMS | 12 | 3.89 ± 3.41 | ||
| ESC | 5 | 11.25 ± 4.99 | 0.1064 | 1.75 [0.45; 3.27] | ||
| ZEM12.5 | 6 | 11.20 ± 9.26 | 0.2312 | 1.14 [−0.002; 2.39] | ||
| ZEM25 | 6 | 5.17 ± 3.31 | >0.9999 | 0.36 [−0.67; 1.42] | ||
| MES | 6 | 6.20 ± 5.45 | >0.9999 | 0.52 [−0.58; 1.65] | ||
| Edge exploration (head dips) | X2(4) = 6.83, p = 0.1453 | UCMS | 12 | 12.58 ± 3.99 | ||
| ESC | 5 | 7.80 ± 1.924 | 0.0604 | 1.28 [−2.47; −0.18] | ||
| ZEM12.5 | 6 | 10.33 ± 3.14 | >0.9999 | 0.57 [−1.59; 0.41] | ||
| ZEM25 | 6 | 10.33 ± 2.66 | 0.8381 | 0.59 [−1.61; 0.39] | ||
| MES | 6 | 11.67 ± 3.39 | >0.9999 | 0.23 [−1.22; 0.75] | ||
| Probe zone duration (s) | X2(4) = 2.61, p = 0.6253 | UCMS | 12 | 34.32 ± 12.82 | ||
| ESC | 5 | 28.64 ± 16.57 | >0.9999 | 0.39 [−1.45; 0.65] | ||
| ZEM12.5 | 6 | 37.08 ± 12.23 | >0.9999 | 0.21 [−0.77; 1.2] | ||
| ZEM25 | 6 | 29.41 ± 11.07 | >0.9999 | 0.38 [−1.38; 0.6] | ||
| MES | 6 | 28.48 ± 10.52 | >0.9999 | 0.46 [−1.47; 0.52] | ||
| Probe/not probe duration | X2(4) = 2.722, p = 0.6054 | UCMS | 12O | 0.71 ± 0.47 | ||
| ESC | 5 | 0.55 ± 0.42 | >0.9999 | 0.32 [−1.38; 0.72] | ||
| ZEM12.5 | 6 | 0.78 ± 0.44 | >0.9999 | 0.15 [−0.83; 1.13] | ||
| ZEM25 | 6 | 0.53 ± 0.32 | >0.9999 | 0.39 [−1.4; 0.58] | ||
| MES | 6 | 0.50 ± 0.27 | >0.9999 | 0.47 [−1.48; 0.51] | ||
| Outside duration (s) | X2(4) = 1.48, p = 0.8306 | UCMS | 12O | 6.76 ± 10.89 | ||
| ESC | 5O | 7.51 ± 9.86 | >0.9999 | 0.07 [−0.97; 1.11] | ||
| ZEM12.5 | 6 | 8.65 ± 9.01 | >0.9999 | 0.17 [−0.8; 1.16] | ||
| ZEM25 | 6 | 12.44 ± 13.64 | >0.9999 | 0.46 [−0.52; 1.47] | ||
| MES | 6 | 11.00 ± 14.66 | >0.9999 | 0.33 [−0.65; 1.33] | ||
| Inside duration (s) | X2(4) = 8.87, p = 0.0644 | UCMS | 12 | 1.39 ± 1.03 | ||
| ESC | 5 | 1.12 ± 0.79 | >0.9999 | 0.26 [−1.32; 0.78] | ||
| ZEM12.5 | 6 | 0.57 ± 0.32 | 0.1962 | 0.89 [−1.95; 0.11] | ||
| ZEM25 | 6 | 1.46 ± 0.62 | >0.9999 | 0.07 [−0.91; 1.05] | ||
| MES | 6 | 0.63 ± 0.39 | 0.2456 | 0.82 [−1.87; 0.18] | ||
| Outer/inner duration (s) | X2(4) = 1.751, p = 0.7814 | UCMS | 12O | 10.58 ± 17.17 | ||
| ESC | 5 | 10.52 ± 11.19 | >0.9999 | 0.003 [−1.05; 1.04] | ||
| ZEM12.5 | 6 | 25.92 ± 29.01 | >0.9999 | 0.67 [−0.31; 1.71] | ||
| ZEM25 | 6 | 10.94 ± 12.10 | >0.9999 | 0.02 [−0.96; 1.00] | ||
| MES | 6 | 32.99 ± 39.72 | >0.9999 | 0.81 [−0.19; 1.86] | ||
| Marker | Kruskal–Wallis | Rx Group | n | Mean ± SD | p-Value | d [95% CI] |
|---|---|---|---|---|---|---|
| Frontal Cortex Analyses | ||||||
| PDE4B (ng/mL) | X2(4) = 13.6, p = 0.0087 | UCMS | 10 | 1.36 ± 0.26 | ||
| ESC | 5 | 1.27 ± 0.21 | >0.9999 | 0.35 [−1.45; 0.72] | ||
| ZEM12.5 | 5 | 0.84 ± 0.05 | 0.0024 | 2.24 [−3.76; −0.95] | ||
| ZEM25 | 5 | 1.17 ± 0.12 | >0.9999 | 0.80 [−1.96; 0.28] | ||
| MES | 5 | 1.31 ± 0.12 | >0.9999 | 0.2 [−1.28; 0.87] | ||
| 5-HT (ng/g) | X2(4) = 4.679, p = 0.3219 | UCMS | 11R | 110.60 ± 32.61 | ||
| ESC | 5 | 140.40 ± 31.39 | 0.2097 | 0.87 [−0.20; 2.02] | ||
| ZEM12.5 | 6 | 116.40 ± 31.86 | >0.9999 | 0.17 [−0.82; 1.17] | ||
| ZEM25 | 6O | 121.50 ± 49.12 | >0.9999 | 0.27 [−0.72; 1.28] | ||
| MES | 5R | 93.75 ± 13.33 | >0.9999 | 0.56 [−1.66; 0.5] | ||
| 5-HIAA (ng/g) | X2(4) = 17.32, p = 0.0017 | UCMS | 12 | 224.80 ± 58.09 | ||
| ESC | 5O | 331.80 ± 59.00 | 0.0515 | 1.74 [0.58; 3.04] | ||
| ZEM12.5 | 6 | 275.00 ± 32.94 | 0.3884 | 0.93 [−0.07; 2.0] | ||
| ZEM25 | 6 | 197.90 ± 27.68 | >0.9999 | 0.51 [−1.52; 0.47] | ||
| MES | 6 | 194.00 ± 33.50 | 0.6475 | 0.57 [−1.59; 0.42] | ||
| 5-HIAA/5-HT | X2(4) = 7.88, p = 0.0963 | UCMS | 12O | 1.92 ± 0.77 | ||
| ESC | 5 | 2.40 ± 0.33 | 0.4550 | 0.67 [−0.38; 1.76] | ||
| ZEM12.5 | 6 | 2.47 ± 0.47 | 0.2933 | 0.76 [−0.24; 1.80] | ||
| ZEM25 | 6 | 1.77 ± 0.50 | >0.9999 | 0.2 [−1.19; 0.78] | ||
| MES | 5R | 2.03 ± 0.17 | >0.9999 | 0.17 [−0.87; 1.22] | ||
| DA (ng/g) | X2(4) = 8.40, p = 0.0780 | UCMS | 12 | 252.60 ± 187.80 | ||
| ESC | 5O | 183.90 ± 125.20 | >0.9999 | 0.38 [−1.44; 0.66] | ||
| ZEM12.5 | 6O | 223.10 ± 132.80 | >0.9999 | 0.16 [−1.15; 0.81] | ||
| ZEM25 | 6 | 235.20 ± 84.82 | >0.9999 | 0.10 [−1.09; 0.88] | ||
| MES | 5R | 123.90 ± 49.94 | 0.0936 | 0.75 [−1.86; 0.30] | ||
| DOPAC (ng/g) | X2(4) = 2.36, p = 0.6702 | UCMS | 12O | 152.60 ± 74.58 | ||
| ESC | 5O | 142.30 ± 57.40 | >0.9999 | 0.14 [−1.19; 0.90] | ||
| ZEM12.5 | 6O | 171.40 ± 118.6 | >0.9999 | 0.2 [−0.78; 1.19] | ||
| ZEM25 | 6O | 162.20 ± 121.7 | >0.9999 | 0.10 [−0.88; 1.08] | ||
| MES | 6 | 113.60 ± 25.30 | 0.5909 | 0.58 [−1.61; 0.4] | ||
| DOPAC/DA | X2(4) = 2.13, p = 0.7128 | UCMS | 12 | 0.74 ± 0.47 | ||
| ESC | 5 | 0.86 ± 0.21 | >0.9999 | 0.28 [−0.77; 1.36] | ||
| ZEM12.5 | 6 | 0.77 ± 0.21 | >0.9999 | 0.08 [−0.92; 1.08] | ||
| ZEM25 | 6 | 0.65 ± 0.24 | >0.9999 | 0.21 [−1.22; 0.78] | ||
| MES | 6 | 0.83 ± 0.44 | >0.9999 | 0.18 [−0.81; 1.19] | ||
| NA (ng/g) | X2(4) = 4.67, p = 0.3225 | UCMS | 11R | 4707 ± 2262 | ||
| ESC | 5O | 3485 ± 1225 | 0.9606 | 0.57 [−1.68; 0.49] | ||
| ZEM12.5 | 6 | 3685 ± 1876 | >0.9999 | 0.45 [−1.48; 0.54] | ||
| ZEM25 | 6 | 2806 ± 406.6 | 0.1456 | 0.97 [−2.07; 0.05] | ||
| MES | 6 | 3599 ± 1053 | >0.9999 | 0.54 [−1.58; 0.45] | ||
| TYR (ng/g) | X2(4) = 5.25, p = 0.2622 | UCMS | 12O | 103.2 ± 28.10 | ||
| ESC | 5O | 99.70 ± 19.47 | >0.9999 | 0.13 [−1.18; 0.91] | ||
| ZEM12.5 | 6 | 106.5 ± 29.07 | >0.9999 | 0.11 [−0.87; 1.1] | ||
| ZEM25 | 6 O | 81.76 ± 12.88 | 0.1748 | 0.84 [−1.89; 0.16] | ||
| MES | 6 | 97.51 ± 17.88 | >0.9999 | 0.21 [−1.20; 0.76] | ||
| 3-CLT (ng/g) | X2(4) = 7.51, p = 0.1111 | UCMS | 12 | 89.97 ± 24.91 | ||
| ESC | 5 | 84.90 ± 14.63 | >0.9999 | 0.21 [−1.27; 0.83] | ||
| ZEM12.5 | 6 | 90.92 ± 25.37 | >0.9999 | 0.04 [−0.94; 1.02] | ||
| ZEM25 | 6O | 68.58 ± 10.90 | 0.0873 | 0.95 [−2.02; 0.06] | ||
| MES | 6 | 91.20 ± 12.99 | >0.9999 | 0.05 [−0.93; 1.04] | ||
| 3-CLT/TYR | X2(4) = 16.30, p = 0.0026 | UCMS | 11R | 0.87 ± 0.03 | ||
| ESC | 5 | 0.86 ± 0.07 | 0.5469 | 0.35 [−1.41; 0.69] | ||
| ZEM12.5 | 6O | 0.85 ± 0.02 | >0.9999 | 0.65 [−1.68; 0.33] | ||
| ZEM25 | 6 | 0.84 ± 0.01 | 0.1682 | 1.35 [−2.49; −0.31] | ||
| MES | 6 | 0.94 ± 0.06 | 0.1325 | 1.76 [0.66; 2.99] | ||
| GSH (ng/g) | X2(4) = 9.98, p = 0.0408 | UCMS | 12 | 85.32 ± 30.01 | ||
| ESC | 5O | 81.12 ± 20.20 | >0.9999 | 0.14 [−1.19; 0.9] | ||
| ZEM12.5 | 6O | 85.74 ± 19.20 | >0.9999 | 0.02 [−0.97; 1.0] | ||
| ZEM25 | 6 | 52.52 ± 7.04 | 0.0291 | 1.24 [−2.35; −0.21] | ||
| MES | 6 | 81.03 ± 10.18 | >0.9999 | 0.16 [−1.15; 0.82] | ||
| GSSG (ng/g) | X2(4) = 8.5, p = 0.075 | UCMS | 12O | 124.7 ± 15.63 | ||
| ESC | 5 | 117.1 ± 34.19 | >0.9999 | 0.33 [−1.39; 0.71] | ||
| ZEM12.5 | 6 | 137.7 ± 25.23 | >0.9999 | 0.64 [−0.34; 1.67] | ||
| ZEM25 | 6 | 130.6 ± 21.91 | >0.9999 | 0.31 [−0.66; 1.31] | ||
| MES | 6O | 90.14 ± 42.64 | 0.0734 | 1.21 [−2.33; −0.19] | ||
| GSSG/GSH | X2(4) = 10.98, p = 0.0268 | UCMS | 12 | 1.67 ± 0.68 | ||
| ESC | 5 | 1.45 ± 0.34 | >0.9999 | 0.35 [−1.41; 0.69] | ||
| ZEM12.5 | 6 | 1.68 ± 0.47 | >0.9999 | 0.004 [−0.98; 0.98] | ||
| ZEM25 | 6O | 2.53 ± 0.55 | 0.0513 | 1.27 [0.23; 2.40] | ||
| MES | 6 | 1.13 ± 0.51 | >0.9999 | 0.82 [−1.87; 0.18] | ||
| Hippocampus Analyses | ||||||
| PDE4B (ng/mL) | X2(4) = 16.18, p = 0.0028 | UCMS | 10 | 1.22 ± 0.1511 | ||
| ESC | 5 | 1.06 ± 0.1467 | 0.5847 | 1.04 [−2.24; 0.06] | ||
| ZEM12.5 | 5O | 0.86 ± 0.09783 | 0.0007 | 2.47 [−4.07; −1.14] | ||
| ZEM25 | 5 | 1.07 ± 0.07701 | 0.3870 | 1.11 [−2.32; 0.01] | ||
| MES | 5 | 0.98 ± 0.05891 | 0.0378 | 1.74 [−3.11; −0.54] | ||
| 5-HT (ng/g) | X2(4) = 18.27, p = 0.0011 | UCMS | 12 | 58.98 ± 12.02 | ||
| ESC | 5 | 75.98 ± 27.43 | 0.6614 | 0.92 [−0.14; 2.06] | ||
| ZEM12.5 | 6O | 68.11 ± 9.16 | 0.4737 | 0.78 [−0.22; 1.82] | ||
| ZEM25 | 6 | 50.33 ± 14.35 | >0.9999 | 0.64 [−1.67; 0.34] | ||
| MES | 6O | 120.10 ± 63.58 | 0.0009 | 1.58 [0.50; 2.77] | ||
| 5-HIAA (ng/g) | X2(4) = 14.08, p = 0.005 | UCMS | 12 | 272.90 ± 51.75 | ||
| ESC | 5 | 394.80 ± 98.69 | 0.1192 | 1.71 [0.56; 3.01] | ||
| ZEM12.5 | 6 | 342.10 ± 70.33 | 0.4017 | 1.13 [0.11; 2.24] | ||
| ZEM25 | 6 | 261.20 ± 37.20 | >0.9999 | 0.23 [−1.22; 0.74] | ||
| MES | 6 | 228.30 ± 36.34 | 0.3507 | 0.89 [−1.96; 0.11] | ||
| 5-HIAA/5-HT | X2(4) = 14.56, p = 0.0057 | UCMS | 12 | 4.79 ± 1.297 | ||
| ESC | 5 | 5.51 ± 1.71 | >0.9999 | 0.48 [−0.56; 1.55] | ||
| ZEM12.5 | 6 | 5.05 ± 1.04 | >0.9999 | 0.2 [−0.78; 1.19] | ||
| ZEM25 | 6 | 5.81 ± 2.67 | >0.9999 | 0.53 [−0.45; 1.55] | ||
| MES | 6 | 2.12 ± 0.59 | 0.0076 | 2.26 [−3.63; −1.08] | ||
| DA (ng/g) | X2(4) = 3.35, p = 0.5007 | UCMS | 12 | 66.56 ± 17.69 | ||
| ESC | 4R | 55.91 ± 14.87 | >0.9999 | 0.59 [−1.77; 0.54] | ||
| ZEM12.5 | 6 | 71.33 ± 9.29 | >0.9999 | 0.29 [−0.68; 1.29] | ||
| ZEM25 | 6 | 59.68 ± 16.57 | >0.9999 | 0.38 [−1.38; 0.60] | ||
| MES | 6 | 85.15 ± 57.20 | >0.9999 | 0.50 [−0.48; 1.52] | ||
| DOPAC (ng/g) | X2(4) = 6.48, p = 0.1662 | UCMS | 12 | 88.42 ± 28.44 | ||
| ESC | 4R | 71.14 ± 17.14 | >0.9999 | 0.62 [−1.8; 0.51] | ||
| ZEM12.5 | 6 | 100.4 ± 35.41 | >0.9999 | 0.37 [−0.61; 1.37] | ||
| ZEM25 | 6 | 94.97 ± 30.32 | >0.9999 | 0.22 [−0.76; 1.21] | ||
| MES | 6 | 68.72 ± 26.50 | 0.2625 | 0.67 [−1.71; 0.31] | ||
| DOPAC/DA | X2(4) = 9.37, p = 0.0446 | UCMS | 12 | 1.39 ± 0.51 | ||
| ESC | 5 | 1.23 ± 0.26 | >0.9999 | 0.10 [−1.17; 0.95] | ||
| ZEM12.5 | 6 | 1.44 ± 0.61 | >0.9999 | 0.40 [−0.6; 1.42] | ||
| ZEM25 | 6 | 1.59 ± 0.32 | 0.4168 | 1.04 [0.01; 2.15] | ||
| MES | 6 | 0.92 ± 0.27 | 0.2455 | 1.15 [−2.28; −0.11] | ||
| NA (ng/g) | X2(4) = 10.5, p = 0.0328 | UCMS | 12 | 3581 ± 1188 | ||
| ESC | 5 | 2266 ± 1158 | 0.1007 | 1.06 [−2.21; 0.02] | ||
| ZEM12.5 | 6 | 3825 ± 9340 | >0.9999 | 0.21 [−0.77; 1.20] | ||
| ZEM25 | 6 | 3609 ± 1271 | >0.9999 | 0.02 [−0.96; 1.003] | ||
| MES | 6 | 2313 ± 617.6 | 0.1221 | 1.13 [−2.3; −0.05] | ||
| TYR (ng/g) | X2(4) = 9.04, p = 0.0602 | UCMS | 12 | 125.8 ± 23.92 | ||
| ESC | 5 | 109.5 ± 15.31 | 0.6956 | 0.70 [−1.81; 0.35] | ||
| ZEM12.5 | 6 | 142.7 ± 23.70 | >0.9999 | 0.68 [−0.31; 1.71] | ||
| ZEM25 | 6 | 123.6 ± 37.82 | >0.9999 | 0.07 [−1.05; 0.91] | ||
| MES | 6 | 101.2 ± 15.53 | 0.1682 | 1.08 [−2.17; −0.06] | ||
| 3-CLT (ng/g) | Inconclusive: Below limit of detection | |||||
| 3-CLT/TYR | Inconclusive: Below limit of detection | |||||
| GSH (ng/g) | X2(4) = 10.75, p = 0.0295 | UCMS | 12 | 72.41 ± 21.14 | ||
| ESC | 5 | 72.47 ± 13.79 | >0.9999 | 0.003 [−1.04; 1.05] | ||
| ZEM12.5 | 6 | 74.65 ± 6.55 | >0.9999 | 0.12 [−0.86; 1.1] | ||
| ZEM25 | 6 | 56.92 ± 9.73 | 0.3050 | 0.8 [−1.85; 0.19] | ||
| MES | 6 | 90.07 ± 5.55 | 0.1817 | 0.95 [−0.06; 2.02] | ||
| GSSG (ng/g) | X2(4) = 9.25. p = 0.0551 | UCMS | 12 | 161.3 ± 41.10 | ||
| ESC | 5 | 124.3 ± 30.73 | 0.3737 | 0.91 [−2.04; 0.15] | ||
| ZEM12.5 | 6 | 154.2 ± 44.30 | >0.9999 | 0.16 [−1.15; 0.81] | ||
| ZEM25 | 6 | 204.1 ± 67.90 | 0.5909 | 0.8 [−0.2; 1.85] | ||
| MES | 6 | 141.7 ± 21.43 | 0.7953 | 0.52 [−1.53; 0.46] | ||
| GSSG/GSH | X2(4) = 12.21, p = 0.0159 | UCMS | 12 | 2.48 ± 1.11 | ||
| ESC | 5 | 1.72 ± 0.34 | >0.9999 | 0.75 [−1.85; 0.31] | ||
| ZEM12.5 | 6O | 2.05 ± 0.46 | >0.9999 | 0.43 [−1.44; 0.55] | ||
| ZEM25 | 6 | 3.56 ± 0.72 | 0.1494 | 1.02 [0.01; 2.1] | ||
| MES | 6 | 1.57 ± 0.17 | 0.3884 | 0.93 [−2.0; 0.07] | ||
| Plasma Analyses | ||||||
| IL-10 (pg/g) | X2(4) = 17.24, p = 0.0017 | UCMS | 12 | 18.66 ± 10.22 | ||
| ESC | 5 | 24.37 ± 10.50 | >0.9999 | 0.52 [−0.55; 1.64] | ||
| ZEM12.5 | 6 | 50.02 ± 10.05 | 0.0007 | 2.9 [1.47; 4.45] | ||
| ZEM25 | 6 | 35.93 ± 2.39 | 0.0271 | 1.89 [0.66; 3.3] | ||
| MES | 6 | 26.13 ± 3.50 | >0.9999 | 0.81 [−0.28; 1.96] | ||
| TNF-α (pg/g) | X2(4) = 10.37, p = 0.0346 | UCMS | 12 | 8.14 ± 3.30 | ||
| ESC | 5 | 3.28 ± 2.96 | 0.1451 | 1.43 [−2.72; −0.28] | ||
| ZEM12.5 | 6 | 8.51 ± 1.44 | >0.9999 | 0.12 [−0.95; 1.2] | ||
| ZEM25 | 6 | 10.91 ± 1.82 | 0.4629 | 0.89 [−0.2; 2.06] | ||
| MES | 6 | 7.6 ± 4.17 | >0.9999 | 0.14 [−1.22; 0.93] | ||
References
- Albert, P.R. Why is depression more prevalent in women? J. Psychiatry Neurosci. 2015, 40, 219–221. [Google Scholar] [CrossRef]
- Kessler, R.C.; Bromet, E.J. The epidemiology of depression across cultures. Annu. Rev. Public Health 2013, 34, 119–138. [Google Scholar] [CrossRef]
- Brand, S.J.; Moller, M.; Harvey, B.H. A review of biomarkers in mood and psychotic disorders: A dissection of clinical vs. preclinical correlates. Curr. Neuropharmacol. 2015, 13, 324–368. [Google Scholar] [CrossRef]
- Thomas, S.J.; Shin, M.; McInnis, M.G.; Bostwick, J.R. Combination therapy with monoamine oxidase inhibitors and other antidepressants or stimulants: Strategies for the management of treatment-resistant depression. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2015, 35, 433–449. [Google Scholar] [CrossRef]
- Rush, A.J.; Zimmerman, M.; Wisniewski, S.R.; Fava, M.; Hollon, S.D.; Warden, D.; Biggs, M.M.; Shores-Wilson, K.; Shelton, R.C.; Luther, J.F.; et al. Comorbid psychiatric disorders in depressed outpatients: Demographic and clinical features. J. Affect. Disord. 2005, 87, 43–55. [Google Scholar] [CrossRef]
- Willner, P. Chronic mild stress (CMS) revisited: Consistency and behavioral -neurobiological concordance in the effects of CMS. Neuropsychobiology 2005, 52, 90–110. [Google Scholar] [CrossRef]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Nollet, M.; Guisquet, A.M.L.; Belzung, C. Models of depression: Unpredictable chronic mild stress in mice. Curr. Protoc. Pharmacol. 2013, 61, 5.65. 1–5.65. 17. [Google Scholar] [CrossRef]
- Bueno-Notivol, J.; Gracia-García, P.; Olaya, B.; Lasheras, I.; López-Antón, R.; Santabárbara, J. Prevalence of depression during the COVID-19 outbreak: A meta-analysis of community-based studies. Int. J. Clin. Health Psychol. 2021, 21, 100196. [Google Scholar] [CrossRef]
- Salari, N.; Hosseinian-Far, A.; Jalali, R.; Vaisi-Raygani, A.; Rasoulpoor, S.; Mohammadi, M.; Rasoulpoor, S.; Khaledi-Paveh, B. Prevalence of stress, anxiety, depression among the general population during the COVID-19 pandemic: A systematic review and meta-analysis. Glob. Health 2020, 16, 57. [Google Scholar] [CrossRef]
- Santabárbara, J.; Lasheras, I.; Lipnicki, D.M.; Bueno-Notivol, J.; Pérez-Moreno, M.; López-Antón, R.; De la Cámara, C.; Lobo, A.; Gracia-García, P. Prevalence of anxiety in the COVID-19 pandemic: An updated meta-analysis of community-based studies. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 109, 110207. [Google Scholar] [CrossRef]
- Duval, E.R.; Javanbakht, A.; Liberzon, I. Neural circuits in anxiety and stress disorders: A focused review. Ther. Clin. Risk Manag. 2015, 11, 115–126. [Google Scholar]
- Walker, F.R.; Nilsson, M.; Jones, K. Acute and chronic stress-induced disturbances of microglial plasticity, phenotype and function. Curr. Drug Targets 2013, 14, 1262–1276. [Google Scholar] [CrossRef]
- McEwen, B.S.; Akil, H. Revisiting the stress concept: Implications for affective disorders. J. Neurosci. 2020, 40, 12–21. [Google Scholar] [CrossRef]
- Willner, P.; Scheel-Krüger, J.; Belzung, C. The neurobiology of depression and antidepressant action. Neurosci. Biobehav. Rev. 2013, 37, 2331–2371. [Google Scholar]
- Bosch, O.; Seifritz, E.; Wetter, T. Stress-related depression: Neuroendocrine, genetic, and therapeutical aspects. World J. Biol. Psychiatry 2012, 13, 556–568. [Google Scholar] [CrossRef]
- Liston, C.; Cichon, J.M.; Jeanneteau, F.; Jia, Z.; Chao, M.V.; Gan, W.-B. Circadian glucocorticoid oscillations promote learning-dependent synapse formation and maintenance. Nat. Neurosci. 2013, 16, 698–705. [Google Scholar] [CrossRef]
- Bennett, A.C. Neuro-and immunomodulatory effects of Sceletium tortuosum. Ph.D. Dissertation, Stellenbosch University, Stellenbosch, South Africa, 2018. [Google Scholar]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M.J.N. Neurobiology of depression. Precis. Med. Commun. 2002, 34, 13–25. [Google Scholar] [CrossRef]
- Jeon, S.W.; Kim, Y.-K. Inflammation-induced depression: Its pathophysiology and therapeutic implications. J. Neuroimmunol. 2017, 313, 92–98. [Google Scholar] [CrossRef]
- Miura, H.; Ozaki, N.; Sawada, M.; Isobe, K.; Ohta, T.; Nagatsu, T. A link between stress and depression: Shifts in the balance between the kynurenine and serotonin pathways of tryptophan metabolism and the etiology and pathophysiology of depression. Stress 2008, 11, 198–209. [Google Scholar] [CrossRef]
- Moller, M.; Swanepoel, T.; Harvey, B.H. Neurodevelopmental animal models reveal the convergent role of neurotransmitter systems, inflammation, and oxidative stress as biomarkers of schizophrenia: Implications for novel drug development. ACS Chem. Neurosci. 2015, 6, 987–1016. [Google Scholar] [CrossRef]
- Pearse, D.D.; Hughes, Z.A. PDE4B as a microglia target to reduce neuroinflammation. Glia 2016, 64, 1698–1709. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; Li, M.; Li, C.; Yu, H.; Zhou, D.; Chen, Z. Reducing neuroinflammation in psychiatric disorders: Novel target of phosphodiesterase 4 (PDE4) and developing of the PDE4 inhibitors. Mech. Neuroinflamm. 2017, 3–23. [Google Scholar]
- Heckman, P.; Blokland, A.; Ramaekers, J.; Prickaerts, J. PDE and cognitive processing: Beyond the memory domain. Neurobiol. Learn. Mem. 2015, 119, 108–122. [Google Scholar] [CrossRef]
- Brunoni, A.R.; Lopes, M.; Fregni, F. A systematic review and meta-analysis of clinical studies on major depression and BDNF levels: Implications for the role of neuroplasticity in depression. Int. J. Neuropsychopharmacol. 2008, 11, 1169–1180. [Google Scholar] [CrossRef]
- Olatunji, T.; Siebert, F.; Adetunji, A.; Harvey, B.; Gericke, J.; Hamman, J.; Van der Kooy, F. Sceletium tortuosum: A review on its phytochemistry, pharmacokinetics, biological and clinical activities. J. Ethnopharmacol. 2021, 280, 114476. [Google Scholar] [CrossRef]
- Harvey, A.L.; Young, L.C.; Viljoen, A.M.; Gericke, N.P. Pharmacological actions of the South African medicinal and functional food plant Sceletium tortuosum and its principal alkaloids. J. Ethnopharmacol. 2011, 137, 1124–1129. [Google Scholar] [CrossRef]
- Swart, A.C.; Smith, C. Modulation of glucocorticoid, mineralocorticoid and androgen production in H295 cells by Trimesemine™, a mesembrine-rich Sceletium extract. J. Ethnopharmacol. 2016, 177, 35–45. [Google Scholar] [CrossRef]
- Gericke, J.; Harvey, B.H.; Pretorius, L.; Ollewagen, T.; Benecke, R.M.; Smith, C. Sceletium tortuosum-derived mesembrine significantly contributes to the anxiolytic effect of Zembrin®, but its anti-depressant effect may require synergy of multiple plant constituents. J. Ethnopharmacol. 2024, 319, 117113. [Google Scholar] [CrossRef]
- Gericke, J. Evaluating the Antidepressant-Like Properties of Sceletium tortuosum, Alone and as Adjunctive Treatment. Doctoral Dissertation, North-West University, Potchefstroom, South-Africa, 2020. [Google Scholar]
- Bekris, S.; Antoniou, K.; Daskas, S.; Papadopoulou-Daifoti, Z. Behavioral and neurochemical effects induced by chronic mild stress applied to two different rat strains. Behav. Brain Res. 2005, 161, 45–59. [Google Scholar] [CrossRef]
- Jayatissa, M.N.; Bisgaard, C.; Tingström, A.; Papp, M.; Wiborg, O. Hippocampal Cytogenesis Correlates to Escitalopram-Mediated Recovery in a Chronic Mild Stress Rat Model of Depression. Neuropsychopharmacology 2006, 31, 2395. [Google Scholar] [CrossRef]
- Shansky, R.M. Are hormones a “female problem” for animal research? Science 2019, 364, 825–826. [Google Scholar] [CrossRef]
- Seney, M.L.; Sibille, E. Sex differences in mood disorders: Perspectives from humans and rodent models. Biol. Sex Differ. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Shepard, R.; Coutellier, L. Sensitivity of the prefrontal GABAergic system to chronic stress in male and female mice: Relevance for sex differences in stress-related disorders. Neuroscience 2016, 332, 1–12. [Google Scholar] [CrossRef]
- Dalla, C.; Antoniou, K.; Drossopoulou, G.; Xagoraris, M.; Kokras, N.; Sfikakis, A.; Papadopoulou-Daifoti, Z. Chronic mild stress impact: Are females more vulnerable? Neuroscience 2005, 135, 703–714. [Google Scholar] [CrossRef]
- Foglio, B.; Panzica, G. Behavioral effects of unpredictable chronic mild stress (UCMS) in a murine model of depression. In Proceedings of the 7th International Meeting Steroids and Nervous System-Abstract Book, Torino, Italy, 19–22 February 2013; p. 150. [Google Scholar]
- Dean, J.; Keshavan, M. The neurobiology of depression: An integrated view. Asian J. Psychiatry 2017, 27, 101–111. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Han, Q.-Q.; Gong, W.-Q.; Pan, D.-H.; Wang, L.-Z.; Hu, W.; Yang, M.; Li, B.; Yu, J.; Liu, Q. Microglial activation mediates chronic mild stress-induced depressive- and anxiety-like behavior in adult rats. J. Neuroinflamm. 2018, 15, 21. [Google Scholar] [CrossRef]
- Kumar, B.H.; Mishra, S.K.; Rana, P.; Singh, S.; Khushu, S. Neurodegenerative evidences during early onset of depression in CMS rats as detected by proton magnetic resonance spectroscopy at 7 T. Behav. Brain Res. 2012, 232, 53–59. [Google Scholar] [CrossRef]
- You, Z.; Luo, C.; Zhang, W.; Chen, Y.; He, J.; Zhao, Q.; Zuo, R.; Wu, Y. Pro-and anti-inflammatory cytokines expression in rat’s brain and spleen exposed to chronic mild stress: Involvement in depression. Behav. Brain Res. 2011, 225, 135–141. [Google Scholar] [CrossRef]
- SANS 10386:2008; The care and use of animals for scientific purposes. South African Bureau of Standards: Pretoria, South Africa, 2008.
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. J. Cereb. Blood Flow Metab. 2020, 40, 1769–1777. [Google Scholar] [CrossRef]
- Zamani, M.; Radahmadi, M.; Reisi, P. Therapeutic effects of exercise, escitalopram and exercise-accompanied escitalopram on brain functions in rats with depression. Physiol. Pharmacol. 2022, 26, 188–199. [Google Scholar] [CrossRef]
- Wiborg, O. Chronic mild stress for modeling anhedonia. Cell Tissue Res. 2013, 354, 155–169. [Google Scholar] [CrossRef]
- Willner, P. Reliability of the chronic mild stress model of depression: A user survey. Neurobiol. Stress 2017, 6, 68–77. [Google Scholar] [CrossRef]
- Wang, J.-M.; Yang, L.-H.; Zhang, Y.-Y.; Niu, C.-L.; Cui, Y.; Feng, W.-S.; Wang, G.-F. BDNF and COX-2 participate in anti-depressive mechanisms of catalpol in rats undergoing chronic unpredictable mild stress. Physiol. Behav. 2015, 151, 360–368. [Google Scholar] [CrossRef]
- He, L.W.; Zeng, L.; Tian, N.; Li, Y.; He, T.; Tan, D.M.; Zhang, Q.; Tan, Y. Optimization of food deprivation and sucrose preference test in SD rat model undergoing chronic unpredictable mild stress. Anim. Models Exp. Med. 2020, 3, 69–78. [Google Scholar] [CrossRef]
- Hirschfeld, R.M. The comorbidity of major depression and anxiety disorders: Recognition and management in primary care. Prim. Care Companion J. Clin. Psychiatry 2001, 3, 244. [Google Scholar] [CrossRef]
- Pawlak, C.R.; Karrenbauer, B.D.; Schneider, P.; Ho, Y.-J. The elevated plus-maze test: Differential psychopharmacology of anxiety-related behavior. Emot. Rev. 2012, 4, 98–115. [Google Scholar] [CrossRef]
- Regenass, W.; Möller, M.; Harvey, B.H. Studies into the anxiolytic actions of agomelatine in social isolation reared rats: Role of corticosterone and sex. J. Psychopharmacol. 2018, 32, 134–145. [Google Scholar] [CrossRef]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef]
- Gawel, K.; Gibula, E.; Marszalek-Grabska, M.; Filarowska, J.; Kotlinska, J.H. Assessment of spatial learning and memory in the Barnes maze task in rodents—Methodological consideration. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 1–18. [Google Scholar] [CrossRef]
- Rodríguez Peris, L.; Scheuber, M.I.; Shan, H.; Braun, M.; Schwab, M.E. Barnes maze test for spatial memory: A new, sensitive scoring system for mouse search strategies. Behav. Brain Res. 2024, 458, 114730. [Google Scholar] [CrossRef]
- Gibula-Tarlowska, E.; Wydra, K.; Kotlinska, J.H. Deleterious effects of ethanol, Δ (9)-Tetrahydrocannabinol (THC), and their combination on the spatial memory and cognitive flexibility in adolescent and adult male rats in the Barnes Maze Task. Pharmaceutics 2020, 12, 654. [Google Scholar] [CrossRef]
- Patil, S.S.; Sunyer, B.; Höger, H.; Lubec, G. Evaluation of spatial memory of C57BL/6J and CD1 mice in the Barnes maze, the Multiple T-maze and in the Morris water maze. Behav. Brain Res. 2009, 198, 58–68. [Google Scholar] [CrossRef]
- Spasojevic, N.; Stefanovic, B.; Jovanovic, P.; Dronjak, S. Anxiety and hyperlocomotion induced by chronic unpredictable mild stress can be moderated with melatonin treatment. Folia Biol. 2016, 62, 250. [Google Scholar] [CrossRef]
- Strekalova, T.; Spanagel, R.; Bartsch, D.; Henn, F.A.; Gass, P. Stress-induced anhedonia in mice is associated with deficits in forced swimming and exploration. Neuropsychopharmacology 2004, 29, 2007–2017. [Google Scholar] [CrossRef]
- Gericke, J.; Lekhooa, M.; Steyn, S.F.; Viljoen, A.M.; Harvey, B.H. An acute dose-ranging evaluation of the antidepressant properties of Sceletium tortuosum (Zembrin®) versus escitalopram in the Flinders Sensitive Line rat. J. Ethnopharmacol. 2022, 284, 114550. [Google Scholar] [CrossRef]
- Sestakova, N.; Puzserova, A.; Kluknavsky, M.; Bernatova, I. Determination of motor activity and anxiety-related behavior in rodents: Methodological aspects and role of nitric oxide. Interdiscip. Toxicol. 2013, 6, 126–135. [Google Scholar] [CrossRef]
- Gould, T.D.; Dao, D.T.; Kovacsics, C.E. The open field test in Mood and Anxiety Related Phenotypes in Mice. In Mood and Anxiety Related Phenotypes in Mice; Humana Press: New York, NY, USA, 2009; pp. 1–20. [Google Scholar]
- Bogdanova, O.V.; Kanekar, S.; D’Anci, K.E.; Renshaw, P.F. Factors influencing behavior in the forced swim test. Physiol. Behav. 2013, 118, 227–239. [Google Scholar] [CrossRef]
- Möller, M.; Du Preez, J.L.; Viljoen, F.P.; Berk, M.; Emsley, R.; Harvey, B.H. Social isolation rearing induces mitochondrial, immunological, neurochemical and behavioral deficits in rats, and is reversed by clozapine or N-acetyl cysteine. Brain Behav. Immun. 2013, 30, 156–167. [Google Scholar] [CrossRef]
- Viljoen, F.; Du Preez, J.; Wessels, J.; Aucamp, M. HPLC electrochemical detection and quantification of monoamines and their metabolites in rat brain tissue samples. Die Pharm.-Int. J. Pharm. Sci. 2018, 73, 563–569. [Google Scholar]
- Bayram, B.; Rimbach, G.; Frank, J.; Esatbeyoglu, T. Rapid method for glutathione quantitation using high-performance liquid chromatography with coulometric electrochemical detection. J. Agric. Food Chem. 2014, 62, 402–408. [Google Scholar] [CrossRef]
- Crow, J.P. [16] Measurement and significance of free and protein-bound 3-nitrotyrosine, 3-chlorotyrosine, and free 3-nitro-4-hydroxyphenylacetic acid in biologic samples: A high-performance liquid chromatography method using electrochemical detection. Methods Enzymol. 1999, 301, 151–160. [Google Scholar]
- Yilmaz, Ö.; Keser, S.; Tuzcu, M.; Güvenc, M.; Cetintas, B.; Irtegün, S.; Tastan, H.; Sahin, K. A practical HPLC method to measure reduced (GSH) and oxidized (GSSG) glutathione concentrations in animal tissues. J. Anim. Vet. Adv. 2009, 8, 343–347. [Google Scholar]
- Statsig. Understanding Significance Levels: A Key to Accurate Data Analysis. 2024. Available online: https://www.statsig.com/blog/understanding-significance-levels-a-key-to-accurate-data-analysis#:~:text=When%20deciding%20on%20a%20significance,significance%20level%20may%20be%20acceptable (accessed on 6 December 2024).
- Scribbr. What Is Effect Size and Why Does It Matter? (Examples). 2020. Available online: https://www.scribbr.com/statistics/effect-size/ (accessed on 15 May 2025).
- De Kloet, E.; Molendijk, M. Coping with the forced swim stressor: Towards understanding an adaptive mechanism. Neural Plast. 2016, 2016, 6503162. [Google Scholar] [CrossRef]
- Commons, K.G.; Cholanians, A.B.; Babb, J.A.; Ehlinger, D.G. The rodent forced swim test measures stress-coping strategy, not depression-like behavior. ACS Chem. Neurosci. 2017, 8, 955–960. [Google Scholar] [CrossRef]
- Anyan, J.; Amir, S. Too depressed to swim or too afraid to stop? A reinterpretation of the forced swim test as a measure of anxiety-like behavior. Neuropsychopharmacology 2018, 43, 931–933. [Google Scholar] [CrossRef]
- De Pablo, J.M.; Parra, A.; Segovia, S.; Guillamón, A. Learned immobility explains the behavior of rats in the forced swimming test. Physiol. Behav. 1989, 46, 229–237. [Google Scholar] [CrossRef]
- Shankman, S.A.; Klein, D.N. The relation between depression and anxiety: An evaluation of the tripartite, approach-withdrawal and valence-arousal models. Clin. Psychol. Rev. 2003, 23, 605–637. [Google Scholar] [CrossRef]
- Kim, Y.-K. Anxiety Disorders: Rethinking and Understanding Recent Discoveries; Springer Nature: Berlin/Heidelberg, Germany, 2020; Volume 1191. [Google Scholar]
- Villada, C.; Espin, L.; Hidalgo, V.; Rubagotti, S.; Sgoifo, A.; Salvador, A. The influence of coping strategies and behavior on the physiological response to social stress in women: The role of age and menstrual cycle phase. Physiol. Behav. 2017, 170, 37–46. [Google Scholar] [CrossRef]
- Franceschelli, A.; Herchick, S.; Thelen, C.; Papadopoulou-Daifoti, Z.; Pitychoutis, P.M. Sex differences in the chronic mild stress model of depression. Behav. Pharmacol. 2014, 25, 372–383. [Google Scholar] [CrossRef]
- Rantala, M.J.; Luoto, S.; Krams, I.; Karlsson, H. Depression subtyping based on evolutionary psychiatry: Proximate mechanisms and ultimate functions. Brain Behav. Immun. 2018, 69, 603–617. [Google Scholar] [CrossRef]
- Sequeira-Cordero, A.; Salas-Bastos, A.; Fornaguera, J.; Brenes, J.C. Behavioral characterisation of chronic unpredictable stress based on ethologically relevant paradigms in rats. Sci. Rep. 2019, 9, 17403. [Google Scholar] [CrossRef]
- Lu, Q.; Mouri, A.; Yang, Y.; Kunisawa, K.; Teshigawara, T.; Hirakawa, M.; Mori, Y.; Yamamoto, Y.; Libo, Z.; Nabeshima, T. Chronic unpredictable mild stress-induced behavioral changes are coupled with dopaminergic hyperfunction and serotonergic hypofunction in mouse models of depression. Behav. Brain Res. 2019, 372, 112053. [Google Scholar] [CrossRef]
- Möhler, H. The GABA system in anxiety and depression and its therapeutic potential. Neuropharmacology 2012, 62, 42–53. [Google Scholar] [CrossRef]
- Strekalova, T.; Spanagel, R.; Dolgov, O.; Bartsch, D. Stress-induced hyperlocomotion as a confounding factor in anxiety and depression models in mice. Behav. Pharmacol. 2005, 16, 171–180. [Google Scholar] [CrossRef]
- Gamaro, G.D.; Manoli, L.P.; Torres, I.L.S.; Silveira, R.; Dalmaz, C. Effects of chronic variate stress on feeding behavior and on monoamine levels in different rat brain structures. Neurochem. Int. 2003, 42, 107–114. [Google Scholar] [CrossRef]
- Heidbreder, C.A.; Weiss, I.C.; Domeney, A.M.; Pryce, C.; Homberg, J.; Hedou, G.; Feldon, J.; Moran, M.C.; Nelson, P. Behavioral, neurochemical and endocrinological characterization of the early social isolation syndrome. Neuroscience 2000, 100, 749–768. [Google Scholar] [CrossRef]
- Kamaei, A.-K.; Hosseini, S.-F.; Teimourparsaei, P.; Payamani, M.; Vaseghi, S. The effect of acute crocin on behavioral changes and BDNF expression level in socially isolated rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2024, 397, 3929–3944. [Google Scholar] [CrossRef]
- Thompson, J.B.; Conrad, S.E.; Torres, C.; Papini, M.R. Inescapable exposure to the Barnes maze increases preference for alcohol over water in rats: Implications for depression and anxiety. Learn. Motiv. 2020, 69, 101602. [Google Scholar] [CrossRef]
- Neuwirth, L.S.; Verrengia, M.T.; Harikinish-Murrary, Z.I.; Orens, J.E.; Lopez, O.E. Under or absent reporting of light stimuli in testing of anxiety-like behaviors in rodents: The need for standardization. Front. Mol. Neurosci. 2022, 15, 912146. [Google Scholar] [CrossRef]
- Hughes, R.N.; Hancock, N.J.; Henwood, G.A.; Rapley, S.A. Evidence for anxiolytic effects of acute caffeine on anxiety-related behavior in male and female rats tested with and without bright light. Behav. Brain Res. 2014, 271, 7–15. [Google Scholar] [CrossRef]
- Liu, J.; Lin, L.; Wang, D.V. Representation of fear of heights by basolateral amygdala neurons. J. Neurosci. 2021, 41, 1080–1091. [Google Scholar] [CrossRef]
- Dalla, C.; Antoniou, K.; Kokras, N.; Drossopoulou, G.; Papathanasiou, G.; Bekris, S.; Daskas, S.; Papadopoulou-Daifoti, Z. Sex differences in the effects of two stress paradigms on dopaminergic neurotransmission. Physiol. Behav. 2008, 93, 595–605. [Google Scholar] [CrossRef]
- Gouirand, A.M.; Matuszewich, L. The effects of chronic unpredictable stress on male rats in the water maze. Physiol. Behav. 2005, 86, 21–31. [Google Scholar] [CrossRef]
- Sarris, J.; Marx, W.; Ashton, M.M.; Ng, C.H.; Galvao-Coelho, N.; Ayati, Z.; Zhang, Z.-J.; Kasper, S.; Ravindran, A.; Harvey, B.H. Plant-based Medicines (Phytoceuticals) in the Treatment of Psychiatric Disorders: A Meta-review of Meta-analyses of Randomized Controlled Trials. Can. J. Psychiatry. 2021, 66, 849–862. [Google Scholar] [CrossRef]
- Kobiella, A.; Reimold, M.; Ulshöfer, D.; Ikonomidou, V.; Vollmert, C.; Vollstädt-Klein, S.; Rietschel, M.; Heinz, A.; Smolka, M.N. How the serotonin transporter 5-HTTLPR polymorphism influences amygdala function: The roles of in vivo serotonin transporter expression and amygdala structure. Transl. Psychiatry 2011, 1, e37. [Google Scholar] [CrossRef]
- Miró, X.; Pérez-Torres, S.; Artigas, F.; Puigdomènech, P.; Palacios, J.M.; Mengod, G. Regulation of cAMP phosphodiesterase mRNAs expression in rat brain by acute and chronic fluoxetine treatment. An in situ hybridization study. Neuropharmacology 2002, 43, 1148–1157. [Google Scholar] [CrossRef]
- Podda, M.V.; Grassi, C. New perspectives in cyclic nucleotide-mediated functions in the CNS: The emerging role of cyclic nucleotide-gated (CNG) channels. Pflügers Arch.-Eur. J. Physiol. 2014, 466, 1241–1257. [Google Scholar] [CrossRef]
- Tibbo, A.J.; Baillie, G.S. Phosphodiesterase 4B: Master Regulator of Brain Signaling. Cells 2020, 9, 1254. [Google Scholar] [CrossRef]
- Mori, T.; Baba, J.; Ichimaru, Y.; Suzuki, T. Effects of Rolipram, a Selective Inhibitor of Phosphodiesterase 4, on Hyperlocomotion Induced by Several Abused Drugs in Mice. Jpn. J. Pharmacol. 2000, 83, 113–118. [Google Scholar] [CrossRef]
- Zhong, Q.; Yu, H.; Huang, C.; Zhong, J.; Wang, H.; Xu, J.; Cheng, Y. FCPR16, a novel phosphodiesterase 4 inhibitor, produces an antidepressant-like effect in mice exposed to chronic unpredictable mild stress. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 90, 62–75. [Google Scholar] [CrossRef]
- Jindal, A.; Mahesh, R.; Bhatt, S. Etazolate, a phosphodiesterase 4 inhibitor reverses chronic unpredictable mild stress-induced depression-like behavior and brain oxidative damage. Pharmacol. Biochem. Behav. 2013, 105, 63–70. [Google Scholar] [CrossRef]
- Shalaby, A.M.; Kamal, S.M. Effect of rolipram, a phosphodiesterase enzyme type–4 inhibitor, on γ-amino butyric acid content of the frontal cortex in mice exposed to chronic mild stress. J. Pharmacol. Pharmacother. 2012, 3, 132–137. [Google Scholar]
- Mac Giollabhui, N.; Ng, T.H.; Ellman, L.M.; Alloy, L.B. The longitudinal associations of inflammatory biomarkers and depression revisited: Systematic review, meta-analysis, and meta-regression. Mol. Psychiatry 2021, 26, 3302–3314. [Google Scholar] [CrossRef]
- Strawbridge, R.; Arnone, D.; Danese, A.; Papadopoulos, A.; Vives, A.H.; Cleare, A.J. Inflammation and clinical response to treatment in depression: A meta-analysis. Eur. Neuropsychopharmacol. 2015, 25, 1532–1543. [Google Scholar] [CrossRef]
- Curtis, M.P.; Hicks, A.J.; Neidigh, J.W. Kinetics of 3-chlorotyrosine formation and loss due to hypochlorous acid and chloramines. Chem. Res. Toxicol. 2011, 24, 418–428. [Google Scholar] [CrossRef]
- Mohiuddin, I.; Chai, H.; Lin, P.H.; Lumsden, A.B.; Yao, Q.; Chen, C. Nitrotyrosine and Chlorotyrosine: Clinical Significance and Biological Functions in the Vascular System. J. Surg. Res. 2006, 133, 143–149. [Google Scholar] [CrossRef]
- Whiteman, M.; Spencer, J.P.E. Loss of 3-chlorotyrosine by inflammatory oxidants: Implications for the use of 3-chlorotyrosine as a bio-marker in vivo. Biochem. Biophys. Res. Commun. 2008, 371, 50–53. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Hypochlorous acid chemistry in mammalian cells—Influence on infection and role in various pathologies. Int. J. Mol. Sci. 2022, 23, 10735. [Google Scholar] [CrossRef]
- Hill, M.N.; Hellemans, K.G.; Verma, P.; Gorzalka, B.B.; Weinberg, J. Neurobiology of chronic mild stress: Parallels to major depression. Neurosci. Biobehav. Rev. 2012, 36, 2085–2117. [Google Scholar] [CrossRef]
- Grønli, J.; Fiske, E.; Murison, R.; Bjorvatn, B.; Sørensen, E.; Ursin, R.; Portas, C.M. Extracellular levels of serotonin and GABA in the hippocampus after chronic mild stress in rats. A microdialysis study in an animal model of depression. Behav. Brain Res. 2007, 181, 42–51. [Google Scholar] [CrossRef]
- Barton, D.A.; Esler, M.D.; Dawood, T.; Lambert, E.A.; Haikerwal, D.; Brenchley, C.; Socratous, F.; Hastings, J.; Guo, L.; Wiesner, G. Elevated brain serotonin turnover in patients with depression: Effect of genotype and therapy. Arch. Gen. Psychiatry 2008, 65, 38–46. [Google Scholar] [CrossRef]
- Whitney, A.J.; Lindeque, Z.; Kruger, R.; Steyn, S.F. Genetically predisposed and resilient animal models of depression reveal divergent responses to early-life adversity. Acta Neuropsychiatr. 2024, 36, 338–350. [Google Scholar] [CrossRef]
- Cao, B.; Zhu, J.; Zuckerman, H.; Rosenblat, J.D.; Brietzke, E.; Pan, Z.; Subramanieapillai, M.; Park, C.; Lee, Y.; McIntyre, R.S. Pharmacological interventions targeting anhedonia in patients with major depressive disorder: A systematic review. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 92, 109–117. [Google Scholar] [CrossRef]
- Ng, C.; Wong, S.; Loh, H.; Yee, A. Anhedonia among patients with Major Depressive Disorder: A comparison between patients on escitalopram and healthy controls. La Clin. Ter. 2014, 165, e384–e390. [Google Scholar]
- Ables, A.Z.; Baughman, O.L., III. Antidepressants: Update on new agents and indications. Am. Fam. Physician 2003, 67, 547–555. [Google Scholar]
- Andrews, P.W.; Bharwani, A.; Lee, K.R.; Fox, M.; Thomson, J.A., Jr. Is serotonin an upper or a downer? The evolution of the serotonergic system and its role in depression and the antidepressant response. Neurosci. Biobehav. Rev. 2015, 51, 164–188. [Google Scholar] [CrossRef]
- Suchting, R.; Tirumalaraju, V.; Gareeb, R.; Bockmann, T.; de Dios, C.; Aickareth, J.; Pinjari, O.; Soares, J.C.; Cowen, P.J.; Selvaraj, S. Revisiting monoamine oxidase inhibitors for the treatment of depressive disorders: A systematic review and network meta-analysis. J. Affect. Disord. 2021, 282, 1153–1160. [Google Scholar] [CrossRef]
- Bennett, A.C.; Smith, C. Immunomodulatory effects of Sceletium tortuosum (Trimesemine™) elucidated in vitro: Implications for chronic disease. J. Ethnopharmacol. 2018, 214, 134–140. [Google Scholar] [CrossRef]
- Tsao Rong, T.R.; Li HongYan, L.H. Antioxidant properties in vitro and in vivo: Realistic assessments of efficacy of plant extracts. CABI Rev. 2012, 1–9. [Google Scholar] [CrossRef]
- Harvey, B.H.; Joubert, C.; du Preez, J.L.; Berk, M. Effect of chronic N-acetyl cysteine administration on oxidative status in the presence and absence of induced oxidative stress in rat striatum. Neurochem. Res. 2008, 33, 508–517. [Google Scholar] [CrossRef]
- Harvey, B.H.; Hamer, M.; Louw, R.; van der Westhuizen, F.H.; Malan, L. Metabolic and glutathione redox markers associated with brain-derived neurotrophic factor in depressed African men and women: Evidence for counterregulation? Neuropsychobiology 2012, 67, 33–40. [Google Scholar] [CrossRef]
- Lushchak, V.I. Glutathione Homeostasis and Functions: Potential Targets for Medical Interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef]
- Rosa, J.M.; Dafre, A.L.; Rodrigues, A.L.S. Antidepressant-like responses in the forced swimming test elicited by glutathione and redox modulation. Behav. Brain Res. 2013, 253, 165–172. [Google Scholar] [CrossRef]
- Schwartz, J.B. The influence of sex on pharmacokinetics. Clin. Pharmacokinet. 2003, 42, 107–121. [Google Scholar] [CrossRef]
- Konkle, A.T.; Baker, S.L.; Kentner, A.C.; Barbagallo, L.S.-M.; Merali, Z.; Bielajew, C. Evaluation of the effects of chronic mild stressors on hedonic and physiological responses: Sex and strain compared. Brain Res. 2003, 992, 227–238. [Google Scholar] [CrossRef]
- Zarrindast, M.-R.; Khakpai, F. The Modulatory Role of Dopamine in Anxiety-like Behavior. Arch Iran Med. 2015, 18, 591–603. [Google Scholar]
- Dionisie, V.; Ciobanu, A.M.; Toma, V.A.; Manea, M.C.; Baldea, I.; Olteanu, D.; Sevastre-Berghian, A.; Clichici, S.; Manea, M.; Riga, S.; et al. Escitalopram Targets Oxidative Stress, Caspase-3, BDNF and MeCP2 in the Hippocampus and Frontal Cortex of a Rat Model of Depression Induced by Chronic Unpredictable Mild Stress. Int. J. Mol. Sci. 2021, 22, 7483. [Google Scholar] [CrossRef]
- Courtiol, E.; Menezes, E.C.; Teixeira, C.M. Serotonergic regulation of the dopaminergic system: Implications for re-ward-related functions. Neurosci. Biobehav. Rev. 2021, 128, 282–293. [Google Scholar] [CrossRef]
- Allegra, S.; Chiara, F.; Di Grazia, D.; Gaspari, M.; De Francia, S. Evaluation of Sex Differences in Preclinical Pharmacology Research: How Far Is Left to Go? Pharmaceuticals 2023, 16, 786. [Google Scholar] [CrossRef]
- Beery, A.K. Inclusion of females does not increase variability in rodent research studies. Curr. Opin. Behav. Sci. 2018, 23, 143–149. [Google Scholar] [CrossRef]
- Becker, J.B.; Prendergast, B.J.; Liang, J.W. Female rats are not more variable than male rats: A meta-analysis of neuroscience studies. Biol. Sex Differ. 2016, 7, 1–7. [Google Scholar] [CrossRef]
- Rincón-Cortés, M.; Herman, J.P.; Lupien, S.; Maguire, J.; Shansky, R.M. Stress: Influence of sex, reproductive status and gender. Neurobiol. Stress 2019, 10, 100155. [Google Scholar] [CrossRef]
- ter Horst, J.P.; de Kloet, E.R.; Schächinger, H.; Oitzl, M.S. Relevance of Stress and Female Sex Hormones for Emotion and Cognition. Cell. Mol. Neurobiol. 2012, 32, 725–735. [Google Scholar] [CrossRef]
- Foglio, B.; Panzica, G. Sexually dimorphic effects of unpredictable chronic mild stress (UCMS) in a murine model of de-pression and anxiety. in Poster Session 2 7th September 2013-poster number P255. 2013. p. 1-1. In Proceedings of the XV National Congress of the Italian Society of Neuroscience (SINS), Rome, Italy, 3–5 October 2013. [Google Scholar]
- Yap, L.-P.; Sancheti, H.; Ybanez, M.D.; Garcia, J.; Cadenas, E.; Han, D. Determination of GSH, GSSG, and GSNO using HPLC with electrochemical detection. Methods Enzymol. 2010, 473, 137–147. [Google Scholar]
- Monostori, P.; Wittmann, G.; Karg, E.; Túri, S. Determination of glutathione and glutathione disulfide in biological samples: An in-depth review. J. Chromatogr. B 2009, 877, 3331–3346. [Google Scholar] [CrossRef]
- Zitka, O.; Skalickova, S.; Gumulec, J.; Masarik, M.; Adam, V.; Hubalek, J.; Trnkova, L.; Kruseova, J.; Eckschlager, T.; Kizek, R. Redox status expressed as GSH:GSSG ratio as a marker for oxidative stress in paediatric tumour patients. Oncol. Lett. 2012, 4, 1247–1253. [Google Scholar] [CrossRef]
- Lakritz, J.; Plopper, C.G.; Buckpitt, A.R. Validated High-Performance Liquid Chromatography-Electrochemical Method for Determination of Glutathione and Glutathione Disulfide in Small Tissue Samples. Anal. Biochem. 1997, 247, 63–68. [Google Scholar] [CrossRef]
- Hensley, K.; Williamson, K.S.; Maidt, M.L.; Prasad Gabbita, S.; Grammas, P.; Floyd, R.A. Determination of biological oxidative stress using high performance liquid chromatography with electrochemical detection (HPLC-ECD). J. High Resolut. Chromatogr. 1999, 22, 429–437. [Google Scholar] [CrossRef]
- Potesil, D.; Petrlova, J.; Adam, V.; Vacek, J.; Klejdus, B.; Zehnalek, J.; Trnkova, L.; Havel, L.; Kizek, R. Simultaneous femtomole determination of cysteine, reduced and oxidized glutathione, and phytochelatin in maize (Zea mays L.) kernels using high-performance liquid chromatography with electrochemical detection. J. Chromatogr. A 2005, 1084, 134–144. [Google Scholar] [CrossRef]
- Shabir, G.A. Step-by-step analytical methods validation and protocol in the quality system compliance industry. J. Valid. Technol. 2005, 10, 314–325. [Google Scholar]
- Harvey, B.H.; Brand, L.; Jeeva, Z.; Stein, D.J. Cortical/hippocampal monoamines, HPA-axis changes and aversive behavior following stress and restress in an animal model of post-traumatic stress disorder. Physiol. Behav. 2006, 87, 881–890. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gericke, J.; Steyn, S.F.; Viljoen, F.P.; Harvey, B.H. Neuro-Inflammatory and Behavioral Changes Are Selectively Reversed by Sceletium tortuosum (Zembrin®) and Mesembrine in Male Rats Subjected to Unpredictable Chronic Mild Stress. Cells 2025, 14, 1029. https://doi.org/10.3390/cells14131029
Gericke J, Steyn SF, Viljoen FP, Harvey BH. Neuro-Inflammatory and Behavioral Changes Are Selectively Reversed by Sceletium tortuosum (Zembrin®) and Mesembrine in Male Rats Subjected to Unpredictable Chronic Mild Stress. Cells. 2025; 14(13):1029. https://doi.org/10.3390/cells14131029
Chicago/Turabian StyleGericke, Johané, Stephan F. Steyn, Francois P. Viljoen, and Brian H. Harvey. 2025. "Neuro-Inflammatory and Behavioral Changes Are Selectively Reversed by Sceletium tortuosum (Zembrin®) and Mesembrine in Male Rats Subjected to Unpredictable Chronic Mild Stress" Cells 14, no. 13: 1029. https://doi.org/10.3390/cells14131029
APA StyleGericke, J., Steyn, S. F., Viljoen, F. P., & Harvey, B. H. (2025). Neuro-Inflammatory and Behavioral Changes Are Selectively Reversed by Sceletium tortuosum (Zembrin®) and Mesembrine in Male Rats Subjected to Unpredictable Chronic Mild Stress. Cells, 14(13), 1029. https://doi.org/10.3390/cells14131029