
The TIP60-CD44 Axis Modulates Colorectal Cancer Stemness



Abstract

1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Generation of Stable Cell Line
2.3. CD44 Overexpression
2.4. RNA Isolation and Quantitative PCR (qPCR)
2.5. Immunofluorescence
2.6. Cell Invasion Assay
2.7. Spheroid Formation Assay (3D Culture)
2.8. Colony Formation Assay and Absorbance Assay
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results and Discussion
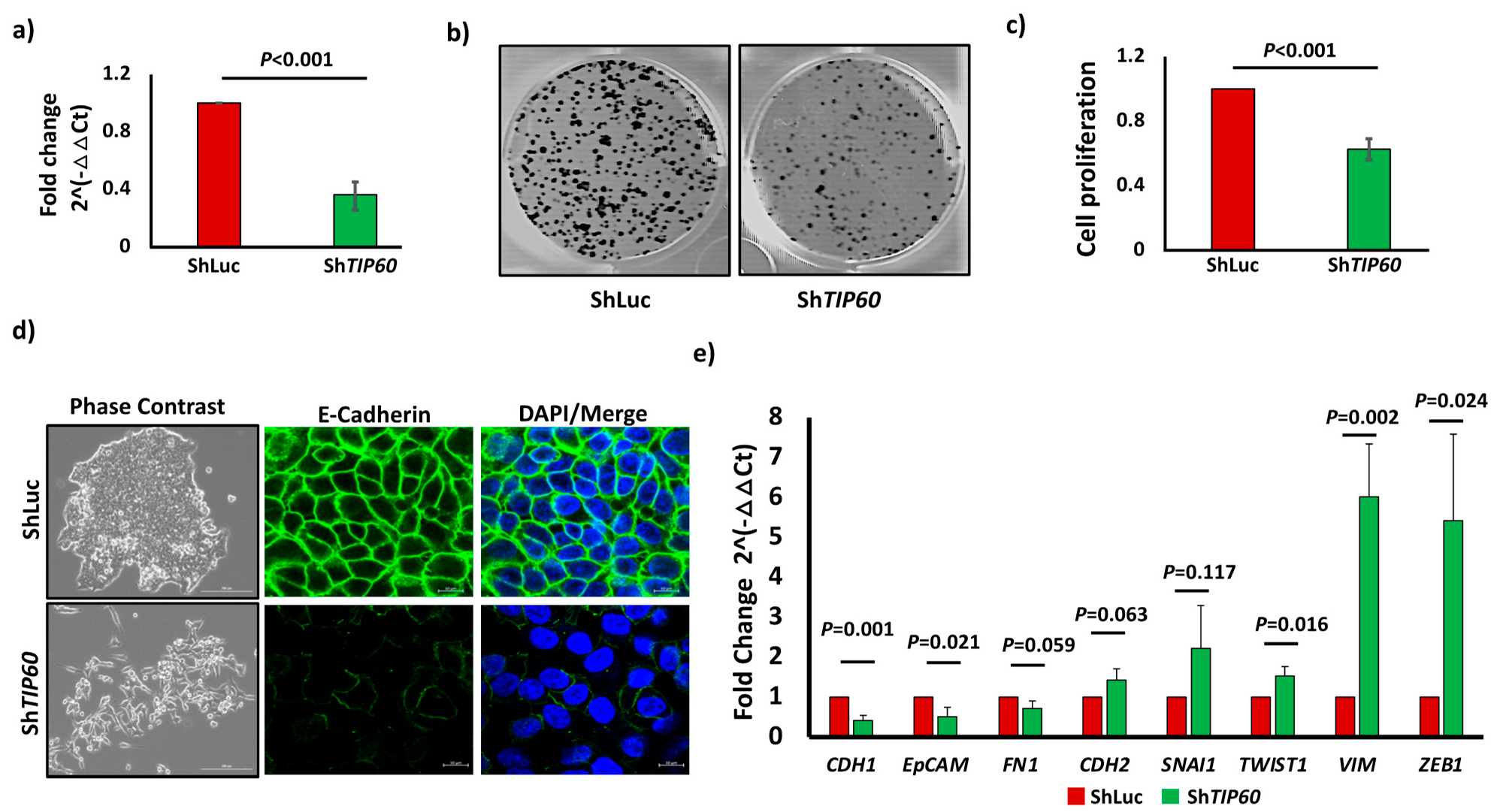
3.1. TIP60 Drives Cell Proliferation and Negatively Regulates EMT in CRC
3.2. TIP60 Modulates Cancer Stem Cell Characteristics in CRC
3.3. The TIP60-CD44 Axis Regulates Cancer Stem Cell Properties
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BME | Basement membrane extract |
| CalceinAM | Calcein Acetoxymethyl Ester |
| CAMs | Cell adhesion molecules |
| CRC | Colorectal cancer |
| CRCSCs | Colorectal cancer stem cells |
| CSCs | Cancer stem cells |
| Ct | Threshold cycle |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMEM | Dulbecco’s Modified Eagle Medium |
| ECL | Enhanced chemiluminescence |
| ECM | Extracellular matrix |
| EMT | Epithelial–mesenchymal transition |
| ESCs | Embryonic stem cells |
| EthD1 | Ethidium Homodimer-1 |
| FBS | Fetal Bovine Serum |
| HRP | Horseradish peroxidase |
| HSCs | Hematopoietic stem cells |
| KAT5 | Lysine acetyltransferase 5 |
| MYST | Moz, Ybf2/Sas3, Sas2, and TIP60 |
| PVDF | Polyvinylidene difluoride |
| RT-qPCR | Quantitative real-time PCR |
| SDS | Sodium dodecyl sulfate |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| TIP60 | HIV-1 Tat-interactive protein |
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Gmeiner, W.H. Recent Advances in Therapeutic Strategies to Improve Colorectal Cancer Treatment. Cancers 2024, 16, 1029. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hammond, W.A.; Swaika, A.; Mody, K. Pharmacologic resistance in colorectal cancer: A review. Ther. Adv. Med. Oncol. 2016, 8, 57–84. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lin, C.C.; Liao, T.T.; Yang, M.H. Immune Adaptation of Colorectal Cancer Stem Cells and Their Interaction with the Tumor Microenvironment. Front. Oncol. 2020, 10, 588542. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yu, Z.; Pestell, T.G.; Lisanti, M.P.; Pestell, R.G. Cancer stem cells. Int. J. Biochem. Cell Biol. 2012, 44, 2144–2151. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Phi, L.T.H.; Sari, I.N.; Yang, Y.G.; Lee, S.H.; Jun, N.; Kim, K.S.; Lee, Y.K.; Kwon, H.Y. Cancer Stem Cells (CSCs) in Drug Resistance and their Therapeutic Implications in Cancer Treatment. Stem Cells Int. 2018, 2018, 5416923. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Marzagalli, M.; Fontana, F.; Raimondi, M.; Limonta, P. Cancer Stem Cells-Key Players in Tumor Relapse. Cancers 2021, 13, 376. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yamamoto, T.; Horikoshi, M. Novel substrate specificity of the histone acetyltransferase activity of HIV-1-Tat interactive protein Tip60. J. Biol. Chem. 1997, 272, 30595–30598. [Google Scholar] [CrossRef] [PubMed]
- Ikura, T.; Ogryzko, V.V.; Grigoriev, M.; Groisman, R.; Wang, J.; Horikoshi, M.; Scully, R.; Qin, J.; Nakatani, Y. Involvement of the TIP60 histone acetylase complex in DNA repair and apoptosis. Cell 2000, 102, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Luo, J.; Zhang, W.; Gu, W. Tip60-dependent acetylation of p53 modulates the decision between cell-cycle arrest and apoptosis. Mol. Cell 2006, 24, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Sykes, S.M.; Mellert, H.S.; Holbert, M.A.; Li, K.; Marmorstein, R.; Lane, W.S.; McMahon, S.B. Acetylation of the p53 DNA-binding domain regulates apoptosis induction. Mol. Cell 2006, 24, 841–851. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sun, Y.; Jiang, X.; Chen, S.; Fernandes, N.; Price, B.D. A role for the Tip60 histone acetyltransferase in the acetylation and activation of ATM. Proc. Natl. Acad. Sci. USA 2005, 102, 13182–13187. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ghobashi, A.H.; Kamel, M.A. Tip60: Updates. J. Appl. Genet. 2018, 59, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Cheng, Y.; Tang, Y.; Martinka, M.; Li, G. Role of Tip60 in human melanoma cell migration, metastasis, and patient survival. J. Investig. Dermatol. 2012, 132, 2632–2641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Subbaiah, V.K.; Rajagopalan, D.; Tham, C.Y.; Abdullah, L.N.; Toh, T.B.; Gong, M.; Tan, T.Z.; Jadhav, S.P.; Pandey, A.K.; et al. TIP60 inhibits metastasis by ablating DNMT1-SNAIL2-driven epithelial-mesenchymal transition program. J. Mol. Cell Biol. 2016, 8, 384–399. [Google Scholar] [CrossRef] [PubMed]
- Numata, A.; Kwok, H.S.; Zhou, Q.L.; Li, J.; Tirado-Magallanes, R.; Angarica, V.E.; Hannah, R.; Park, J.; Wang, C.Q.; Krishnan, V.; et al. Lysine acetyltransferase Tip60 is required for hematopoietic stem cell maintenance. Blood 2020, 136, 1735–1747. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, P.B.; Hung, J.H.; Hickman, T.L.; Coles, A.H.; Carey, J.F.; Weng, Z.; Chu, F.; Fazzio, T.G. Hdac6 regulates Tip60-p400 function in stem cells. Elife 2013, 2, e01557. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hassn Mesrati, M.; Syafruddin, S.E.; Mohtar, M.A.; Syahir, A. CD44: A Multifunctional Mediator of Cancer Progression. Biomolecules 2021, 11, 1850. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, L.; Zuo, X.; Xie, K.; Wei, D. The Role of CD44 and Cancer Stem Cells. Methods Mol. Biol. 2018, 1692, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Stacy, A.J.; Zhang, J.; Craig, M.P.; Hira, A.; Dole, N.; Kadakia, M.P. TIP60 upregulates ΔNp63α to promote cellular proliferation. J. Biol. Chem. 2019, 294, 17007–17016. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shiota, M.; Yokomizo, A.; Masubuchi, D.; Tada, Y.; Inokuchi, J.; Eto, M.; Uchiumi, T.; Fujimoto, N.; Naito, S. Tip60 promotes prostate cancer cell proliferation by translocation of androgen receptor into the nucleus. Prostate 2010, 70, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, D.; Barker, N.; McNeil, N.; Hu, Y.; Camps, J.; McKinnon, K.; Clevers, H.; Ried, T.; Gaiser, T. LGR5 positivity defines stem-like cells in colorectal cancer. Carcinogenesis 2014, 35, 849–858. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Elbadawy, M.; Usui, T.; Yamawaki, H.; Sasaki, K. Emerging Roles of C-Myc in Cancer Stem Cell-Related Signaling and Resistance to Cancer Chemotherapy: A Potential Therapeutic Target Against Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 2340. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhou, H.; Hu, Y.U.; Wang, W.; Mao, Y.; Zhu, J.; Zhou, B.; Sun, J.; Zhang, X. Expression of Oct-4 is significantly associated with the development and prognosis of colorectal cancer. Oncol Lett. 2015, 10, 691–696. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, J.; Espinoza, L.A.; Kinders, R.J.; Lawrence, S.M.; Pfister, T.D.; Zhou, M.; Veenstra, T.D.; Thorgeirsson, S.S.; Jessup, J.M. NANOG modulates stemness in human colorectal cancer. Oncogene 2013, 32, 4397–4405. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Frank, S.R.; Parisi, T.; Taubert, S.; Fernandez, P.; Fuchs, M.; Chan, H.M.; Livingston, D.M.; Amati, B. MYC recruits the TIP60 histone acetyltransferase complex to chromatin. EMBO Rep. 2003, 4, 575–580. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shibahara, D.; Akanuma, N.; Kobayashi, I.S.; Heo, E.; Ando, M.; Fujii, M.; Jiang, F.; Prin, P.N.; Pan, G.; Wong, K.K.; et al. TIP60 is required for tumorigenesis in non-small cell lung cancer. Cancer Sci. 2023, 114, 2400–2413. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sakuraba, K.; Yasuda, T.; Sakata, M.; Kitamura, Y.H.; Shirahata, A.; Goto, T.; Mizukami, H.; Saito, M.; Ishibashi, K.; Kigawa, G.; et al. Down-regulation of Tip60 gene as a potential marker for the malignancy of colorectal cancer. Anticancer Res. 2009, 29, 3953–3955. [Google Scholar] [PubMed]
- Sakuraba, K.; Yokomizo, K.; Shirahata, A.; Goto, T.; Saito, M.; Ishibashi, K.; Kigawa, G.; Nemoto, H.; Hibi, K. TIP60 as a potential marker for the malignancy of gastric cancer. Anticancer Res. 2011, 31, 77–79. [Google Scholar] [PubMed]
- Bassi, C.; Li, Y.T.; Khu, K.; Mateo, F.; Baniasadi, P.S.; Elia, A.; Mason, J.; Stambolic, V.; Pujana, M.A.; Mak, T.W.; et al. The acetyltransferase Tip60 contributes to mammary tumorigenesis by modulating DNA repair. Cell Death Differ. 2016, 23, 1198–1208. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward 5′ to 3′ | Reverse 5′ to 3′ |
|---|---|---|
| TIP60 | AATGTGGCCTGCATCCTAAC | TGTTTTCCCTTCCACTTTGG |
| CD44 | CACATATTGCTTCAATGCCTCAG | CCATCACGGTTGACAATAGTTATG |
| MYC | TCAAGAGGTGCCACGTCTCC | TCTTGGCAGCAGGATAGTCCTT |
| OCT4 | GGAGTCTGGAGACCATGTTTCTG | GAACCATACTCGAACCACATCCTT |
| LGR5 | TTCAATCCCTGCGCCTAGAT | TGCAGGCCGCTGAAACA |
| NANOG | TGGAAGCCACTAGGGAAAGC | TGGAGTCACACACTAGTTCACGAATAA |
| CDH1 | TTACTGCCCCCAGAGGATGA | TGCAACGTCGTTACGAGTCA |
| EpCAM | GCTGGCCGTAAACTGCTTTG | ACATTTGGCAGCCAGCTTTG |
| FN1 | AACCCTTCCACACCCCAATC | ACTGGGTTGCTGACCAGAAG |
| CDH2 | CCGGTTTCATTTGAGGGCAC | TCCCTCAGGAACTGTCCCAT |
| SNAI1 | TCTTTCCTCGTCAGGAAGCC | GATCTCCGGAGGTGGGATGG |
| SNAI2 | CTCCTCATCTTTGGGGCGAG | CTTCAATGGCATGGGGGTCT |
| TWIST1 | TCGGACAAGCTGAGCAAGATT | GCAGCTTGCCATCTTGGAGT |
| VIM | CTGCCAACCGGAACAATGAC | CATTTCACGCATCTGGCGTT |
| ZEB1 | AGGATGACCTGCCAACAGAC | CTTCAGGCCCCAGGATTTCTT |
| ACTB | CCAGATCATGTTTGAGACCTTCAAC | CCAGAGGCGTACAGGGATAGC |
| GAPDH | CAGCCTCAAGATCATCAGCA | TGTGGTCATGAGTCCTTCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammad, A.; Jha, S. The TIP60-CD44 Axis Modulates Colorectal Cancer Stemness. Cells 2025, 14, 686. https://doi.org/10.3390/cells14100686
Mohammad A, Jha S. The TIP60-CD44 Axis Modulates Colorectal Cancer Stemness. Cells. 2025; 14(10):686. https://doi.org/10.3390/cells14100686
Chicago/Turabian StyleMohammad, Asad, and Sudhakar Jha. 2025. "The TIP60-CD44 Axis Modulates Colorectal Cancer Stemness" Cells 14, no. 10: 686. https://doi.org/10.3390/cells14100686
APA StyleMohammad, A., & Jha, S. (2025). The TIP60-CD44 Axis Modulates Colorectal Cancer Stemness. Cells, 14(10), 686. https://doi.org/10.3390/cells14100686