
Pre-Clinical Assessment of SAR442257, a CD38/CD3xCD28 Trispecific T Cell Engager in Treatment of Relapsed/Refractory Multiple Myeloma

 , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Binding Avidity Assays
2.3. Cytotoxicity Assays and TC Proliferation Assays
2.4. Cytokine Analysis
2.5. FACS Analysis
2.6. Quantification and Statistical Analysis
3. Results
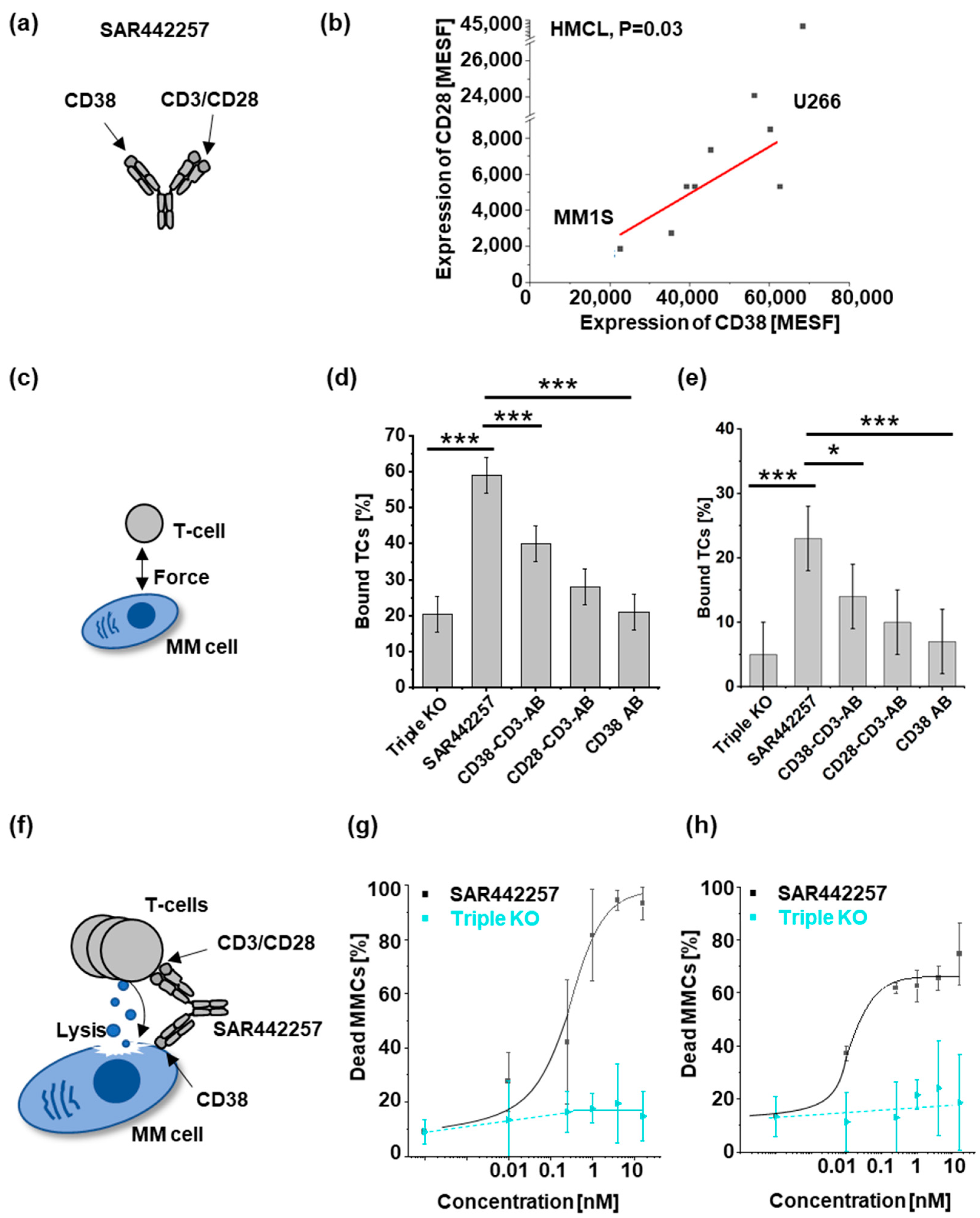
3.1. SAR442257 Enhances Cell Avidity and Tumor Killing in Human Myeloma Cell Lines
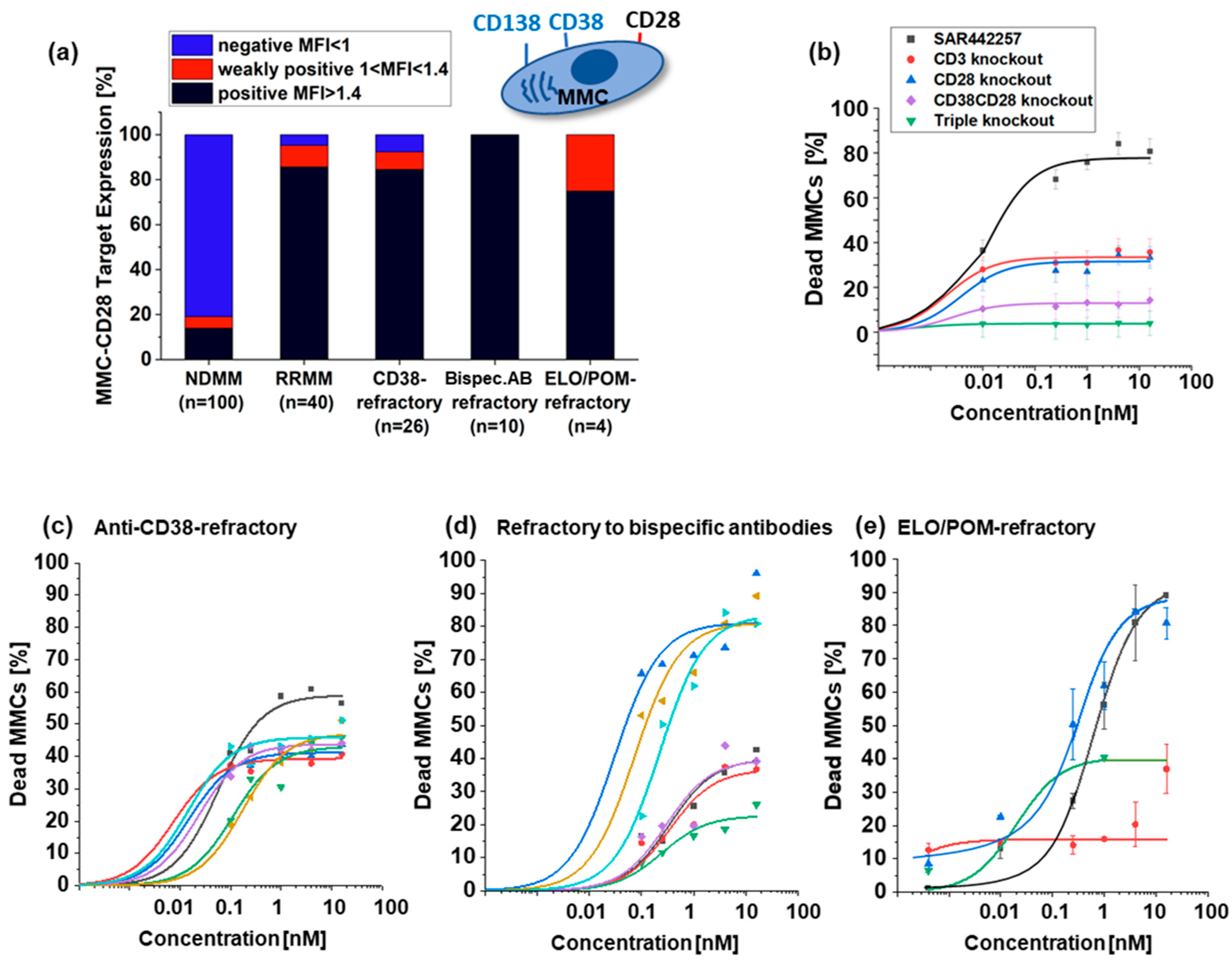
3.2. Activity of SAR442257 in RRMM Patient Cohorts
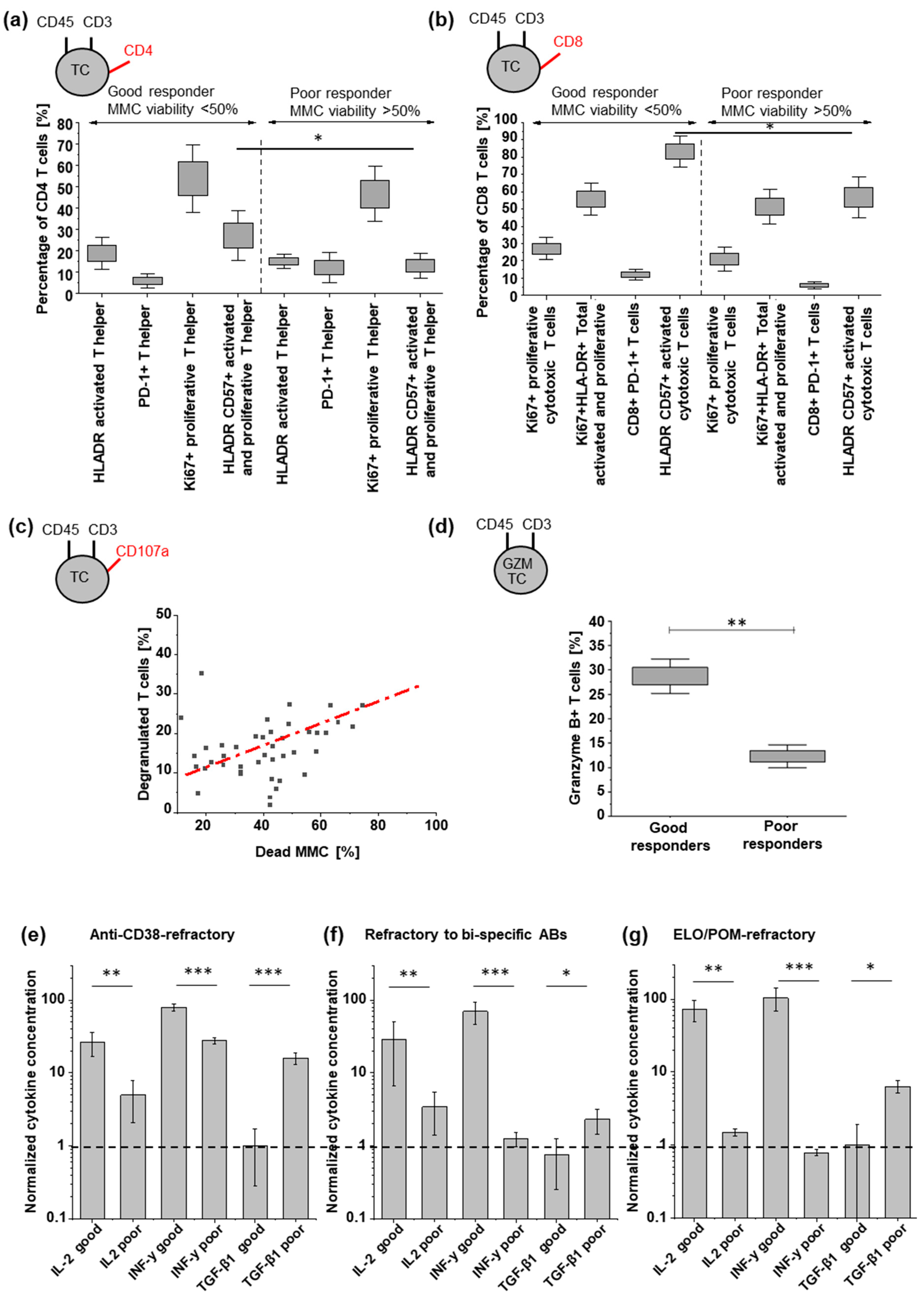
3.3. Tumor Cell Killing Correlates with Induced TC Activation
3.4. TGF-β Can Reduce TC Engager Efficiency by Reducing T Cell Activation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raje, N.; Mateos, M.-V.; Iida, S.; Reece, D. Clinical evidence for immune-based strategies in early-line multiple myeloma: Current challenges in decision-making for subsequent therapy. Blood Cancer J. 2023, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Mateo, G.; Castellanos, M.; Rasillo, A.; Gutierrez, N.C.; Montalban, M.A.; Martin, M.L.; Hernandez, J.M.; Lopez-Berges, M.C.; Montejano, L.; Blade, J.; et al. Genetic abnormalities and patterns of antigenic expression in multiple myeloma. Clin. Cancer Res. 2005, 11, 3661–3667. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.; Orfao, A.; Ocqueteau, M.; Mateo, G.; Corral, M.; Caballero, M.D.; Blade, J.; Moro, M.J.; Hernandez, J.; San Miguel, J.F. High-sensitive immunophenotyping and DNA ploidy studies for the investigation of minimal residual disease in multiple myeloma. Br. J. Haematol. 1999, 107, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Bahlis, N.J.; King, A.M.; Kolonias, D.; Carlson, L.M.; Liu, H.Y.; Hussein, M.A.; Terebelo, H.R.; Byrne, G.E., Jr.; Levine, B.L.; Boise, L.H.; et al. CD28-mediated regulation of multiple myeloma cell proliferation and survival. Blood 2007, 109, 5002–5010. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.J.; Pronschinske, K.B.; Shyer, J.A.; Sharma, S.; Leung, S.; Curran, S.A.; Lesokhin, A.M.; Devlin, S.M.; Giralt, S.A.; Young, J.W. T-cell Exhaustion in Multiple Myeloma Relapse after Autotransplant: Optimal Timing of Immunotherapy. Cancer Immunol. Res. 2016, 4, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Pilcher, W.; Thomas, B.E.; Bhasin, S.S.; Jayasinghe, R.G.; Rahman, A.H.; Kim-Schulze, S.; Gonzalez-Kozlova, E.; Kourelis, T.; Dhodapkar, M.V.; Vij, R.; et al. Characterization of T-Cell Exhaustion in Rapid Progressing Multiple Myeloma Using Cross Center Scrna-Seq Study. Blood 2021, 138, 401. [Google Scholar] [CrossRef]
- Friedrich, M.J.; Neri, P.; Kehl, N.; Michel, J.; Steiger, S.; Kilian, M.; Leblay, N.; Maity, R.; Sankowski, R.; Lee, H.; et al. The pre-existing T cell landscape determines the response to bispecific T cell engagers in multiple myeloma patients. Cancer Cell 2023, 41, 711–725.e6. [Google Scholar] [CrossRef] [PubMed]
- Awwad, M.H.S.; Mahmoud, A.; Bruns, H.; Echchannaoui, H.; Kriegsmann, K.; Lutz, R.; Raab, M.S.; Bertsch, U.; Munder, M.; Jauch, A.; et al. Selective elimination of immunosuppressive T cells in patients with multiple myeloma. Leukemia 2021, 35, 2602–2615. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, M.A.; Richardson, P.; Lonial, S. Treatment Options for Patients With Heavily Pretreated Relapsed and Refractory Multiple Myeloma. Clin. Lymphoma Myeloma Leuk. 2022, 22, 460–473. [Google Scholar] [CrossRef]
- Ailawadhi, S.; Parrondo, R.D.; Laplant, B.; Alegria, V.R.; Elliott, J.B.; Sher, T.; Paulus, A.; Chapin, D.; Heslop, K.; Chanan-Khan, A.; et al. Phase II Trial of Elotuzumab with Pomalidomide and Dexamethasone for Relapsed/Refractory Multiple Myeloma (RRMM) in the Post-Daratumumab Progression Setting. Blood 2022, 140, 4450–4451. [Google Scholar] [CrossRef]
- Bryant, A. Elotuzumab. In Resistance to Targeted Therapies in Multiple Myeloma; Ling, S.C.W., Trieu, S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 73–82. [Google Scholar]
- Saltarella, I.; Desantis, V.; Melaccio, A.; Solimando, A.G.; Lamanuzzi, A.; Ria, R.; Storlazzi, C.T.; Mariggiò, M.A.; Vacca, A.; Frassanito, M.A. Mechanisms of Resistance to Anti-CD38 Daratumumab in Multiple Myeloma. Cells 2020, 9, 167. [Google Scholar] [CrossRef]
- Caraccio, C.; Krishna, S.; Phillips, D.J.; Schürch, C.M. Bispecific Antibodies for Multiple Myeloma: A Review of Targets, Drugs, Clinical Trials, and Future Directions. Front. Immunol. 2020, 11, 501. [Google Scholar] [CrossRef] [PubMed]
- Usmani, S.Z.; Garfall, A.L.; van de Donk, N.; Nahi, H.; San-Miguel, J.F.; Oriol, A.; Rosinol, L.; Chari, A.; Bhutani, M.; Karlin, L.; et al. Teclistamab, a B-cell maturation antigen x CD3 bispecific antibody, in patients with relapsed or refractory multiple myeloma (MajesTEC-1): A multicentre, open-label, single-arm, phase 1 study. Lancet 2021, 398, 665–674. [Google Scholar] [PubMed]
- Sanchez, L.; Dardac, A.; Madduri, D.; Richard, S.; Richter, J. B-cell maturation antigen (BCMA) in multiple myeloma: The new frontier of targeted therapies. Ther. Adv. Hematol. 2021, 12, 2040620721989585. [Google Scholar] [CrossRef]
- Seckinger, A.; Delgado, J.A.; Moser, S.; Moreno, L.; Neuber, B.; Grab, A.; Li, J.; Stagg, N.J.; Johnston, J.; Harris, M.J.; et al. T-Cell Bispecific Antibodies Suppress Multiple Myeloma. Cancer Discov. 2017, 7, 452. [Google Scholar]
- Raab, M.S.; Cohen, Y.C.; Schjesvold, F.; Aardalen, K.; Oka, A.; Spencer, A.; Wermke, M.; Souza, A.D.; Kaufman, J.L.; Cafro, A.M.; et al. Preclinical discovery and initial clinical data of WVT078, a BCMA × CD3 bispecific antibody. Leukemia 2023, 37, 1349–1360. [Google Scholar] [CrossRef]
- Kararoudi, M.N.; Nagai, Y.; Elmas, E.; Pereira, M.D.F.; Ali, S.A.; Imus, P.H.; Wethington, D.; Borrello, I.M.; Lee, D.A.; Ghiaur, G. CD38 deletion of human primary NK cells eliminates daratumumab-induced fratricide and boosts their effector activity. Blood 2020, 136, 2416–2427. [Google Scholar] [CrossRef]
- Chari, A.; Minnema, M.C.; Berdeja, J.G.; Oriol, A.; van de Donk, N.; Rodríguez-Otero, P.; Askari, E.; Mateos, M.V.; Costa, L.J.; Caers, J.; et al. Talquetamab, a T-Cell-Redirecting GPRC5D Bispecific Antibody for Multiple Myeloma. N. Engl. J. Med. 2022, 387, 2232–2244. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Durante, M.; Ahn, S.; Leblay, N.; Poorebrahim, M.; Maity, R.; Tilmont, R.; Barakat, E.; Jung, D.; Ziccheddu, B.; et al. The Impact of Soluble BCMA and BCMA Gain on Anti-BCMA Immunotherapies in Multiple Myeloma. Blood 2023, 142, 4688. [Google Scholar] [CrossRef]
- Laurent, S.A.; Hoffmann, F.S.; Kuhn, P.H.; Cheng, Q.; Chu, Y.; Schmidt-Supprian, M.; Hauck, S.M.; Schuh, E.; Krumbholz, M.; Rubsamen, H.; et al. gamma-Secretase directly sheds the survival receptor BCMA from plasma cells. Nat. Commun. 2015, 6, 7333. [Google Scholar]
- Sade-Feldman, M.; Yizhak, K.; Bjorgaard, S.L.; Ray, J.P.; de Boer, C.G.; Jenkins, R.W.; Lieb, D.J.; Chen, J.H.; Frederick, D.T.; Barzily-Rokni, M.; et al. Defining T Cell States Associated with Response to Checkpoint Immunotherapy in Melanoma. Cell 2019, 176, 404. [Google Scholar] [CrossRef]
- Li, H.; van der Leun, A.M.; Yofe, I.; Lubling, Y.; Gelbard-Solodkin, D.; van Akkooi, A.C.J.; van den Braber, M.; Rozeman, E.A.; Haanen, J.; Blank, C.U.; et al. Dysfunctional CD8 T Cells Form a Proliferative, Dynamically Regulated Compartment within Human Melanoma. Cell 2019, 176, 775–789.e18. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Seung, E.; Xu, L.; Rao, E.; Lord, D.M.; Wei, R.R.; Cortez-Retamozo, V.; Ospina, B.; Posternak, V.; Ulinski, G.; et al. Trispecific antibodies enhance the therapeutic efficacy of tumor-directed T cells through T cell receptor co-stimulation. Nat. Cancer 2020, 1, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Beyersdorf, N.; Kerkau, T.; Hünig, T. CD28 co-stimulation in T-cell homeostasis: A recent perspective. Immunotargets Ther. 2015, 4, 111–122. [Google Scholar]
- Lee, H.; Ahn, S.; Maity, R.; Leblay, N.; Ziccheddu, B.; Truger, M.; Chojnacka, M.; Cirrincione, A.; Durante, M.; Tilmont, R.; et al. Mechanisms of antigen escape from BCMA- or GPRC5D-targeted immunotherapies in multiple myeloma. Nat. Med. 2023, 29, 2295–2306. [Google Scholar] [CrossRef]
- Samur, M.K.; Fulciniti, M.; Aktas Samur, A.; Bazarbachi, A.H.; Tai, Y.-T.; Prabhala, R.; Alonso, A.; Sperling, A.S.; Campbell, T.; Petrocca, F.; et al. Biallelic loss of BCMA as a resistance mechanism to CAR T cell therapy in a patient with multiple myeloma. Nat. Commun. 2021, 12, 868. [Google Scholar] [CrossRef]
- Pellat-Deceunynck, C.; Bataille, R.; Robillard, N.; Harousseau, J.L.; Rapp, M.J.; Juge-Morineau, N.; Wijdenes, J.; Amiot, M. Expression of CD28 and CD40 in human myeloma cells: A comparative study with normal plasma cells. Blood 1994, 84, 2597–2603. [Google Scholar] [CrossRef]
- Robillard, N.; Jego, G.; Pellat-Deceunynck, C.; Pineau, D.; Puthier, D.; Mellerin, M.P.; Barille, S.; Rapp, M.J.; Harousseau, J.L.; Amiot, M.; et al. CD28, a marker associated with tumoral expansion in multiple myeloma. Clin. Cancer Res. 1998, 4, 1521–1526. [Google Scholar] [PubMed]
- Shapiro, V.S.; Mollenauer, M.N.; Weiss, A. Endogenous CD28 expressed on myeloma cells up-regulates interleukin-8 production: Implications for multiple myeloma progression. Blood 2001, 98, 187–193. [Google Scholar] [CrossRef]
- Utley, A.; Peng, P.; Liu, W.S.; Ventrone, D.; Lee, K.P. CD28 Induces Metabolic Fitness in Multiple Myeloma for ROS-Dependent Survival. Blood 2018, 132, 3220. [Google Scholar] [CrossRef]
- Nair, J.R.; Carlson, L.M.; Koorella, C.; Rozanski, C.H.; Byrne, G.E.; Bergsagel, P.L.; Shaughnessy, J.P., Jr.; Boise, L.H.; Chanan-Khan, A.; Lee, K.P. CD28 expressed on malignant plasma cells induces a prosurvival and immunosuppressive microenvironment. J. Immunol. 2011, 187, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Garfall, A.L.; June, C.H. Trispecific antibodies offer a third way forward for anticancer immunotherapy. Nature 2019, 575, 450–451. [Google Scholar] [CrossRef] [PubMed]
- Gantke, T.; Weichel, M.; Herbrecht, C.; Reusch, U.; Ellwanger, K.; Fucek, I.; Eser, M.; Muller, T.; Griep, R.; Molkenthin, V.; et al. Trispecific antibodies for CD16A-directed NK cell engagement and dual-targeting of tumor cells. Protein Eng. Des. Sel. 2017, 30, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Abrams, R.E.; Pierre, K.; El-Murr, N.; Seung, E.; Wu, L.; Luna, E.; Mehta, R.; Li, J.; Larabi, K.; Ahmed, M.; et al. Quantitative systems pharmacology modeling sheds light into the dose response relationship of a trispecific T cell engager in multiple myeloma. Sci. Rep. 2022, 12, 10976. [Google Scholar] [CrossRef] [PubMed]
- Malek, E.; Rana, P.S.; Swamydas, M.; Daunov, M.; Miyagi, M.; Murphy, E.; Ignatz-Hoover, J.J.; Metheny, L.; Seong Jin, K.; Driscoll, J.J. Vactosertib, a novel TGF-β1 type I receptor kinase inhibitor, improves T-cell fitness: A single-arm, phase 1b trial in relapsed/refractory multiple myeloma. Blood 2023, 142, 4749. [Google Scholar] [CrossRef]
- Grab, A.L.; Seckinger, A.; Horn, P.; Hose, D.; Cavalcanti-Adam, E.A. Hyaluronan hydrogels delivering BMP-6 for local targeting of malignant plasma cells and osteogenic differentiation of mesenchymal stromal cells. Acta Biomater. 2019, 96, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Larson, R.C.; Kann, M.C.; Bailey, S.R.; Haradhvala, N.J.; Llopis, P.M.; Bouffard, A.A.; Scarfó, I.; Leick, M.B.; Grauwet, K.; Berger, T.R.; et al. CAR T cell killing requires the IFNγR pathway in solid but not liquid tumours. Nature 2022, 604, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Urlaub, D.; Watzl, C. Coated Latex Beads as Artificial Cells for Quantitative Investigations of Receptor/Ligand Interactions. Curr. Protoc. Immunol. 2020, 131, e111. [Google Scholar] [CrossRef]
- Herbst, S.A.; Kim, V.; Roider, T.; Schitter, E.C.; Bruch, P.M.; Liebers, N.; Kolb, C.; Knoll, M.; Lu, J.; Dreger, P.; et al. Comparing the value of mono- vs coculture for high-throughput compound screening in hematological malignancies. Blood Adv. 2023, 7, 5925–5936. [Google Scholar] [CrossRef]
- Bazarbachi, A.H.; Al Hamed, R.; Malard, F.; Harousseau, J.-L.; Mohty, M. Relapsed refractory multiple myeloma: A comprehensive overview. Leukemia 2019, 33, 2343–2357. [Google Scholar] [CrossRef]
- Moreau, P.; Touzeau, C. T-cell-redirecting bispecific antibodies in multiple myeloma: A revolution? Blood 2022, 139, 3681–3687. [Google Scholar] [CrossRef] [PubMed]
- Esensten, J.H.; Helou, Y.A.; Chopra, G.; Weiss, A.; Bluestone, J.A. CD28 Costimulation: From Mechanism to Therapy. Immunity 2016, 44, 973–988. [Google Scholar] [CrossRef] [PubMed]
- Hashim, L.; Faisal, M.S.; Iqbal, M.A.; Saeed, H.; Samhouri, Y.; Shahzad, M.; Khattak, Z.E.; Anwer, F. Bispecific T-Cell Engager Antibodies in Multiple Myeloma- a Systematic Review of Phase 1 Clinical Trials. Blood 2021, 138, 4775. [Google Scholar] [CrossRef]
- Buie, L.W.; Pecoraro, J.J.; Horvat, T.Z.; Daley, R.J. Blinatumomab: A First-in-Class Bispecific T-Cell Engager for Precursor B-Cell Acute Lymphoblastic Leukemia. Ann. Pharmacother. 2015, 49, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Seckinger, A.; Delgado, J.A.; Moser, S.; Moreno, L.; Neuber, B.; Grab, A.; Lipp, S.; Merino, J.; Prosper, F.; Emde, M.; et al. Target Expression, Generation, Preclinical Activity, and Pharmacokinetics of the BCMA-T Cell Bispecific Antibody EM801 for Multiple Myeloma Treatment. Cancer Cell 2017, 31, 396–410. [Google Scholar] [CrossRef]
- Alhallak, K.; Sun, J.; Jeske, A.; Park, C.; Yavner, J.; Bash, H.; Lubben, B.; Adebayo, O.; Khaskiah, A.; Azab, A.K. Bispecific T Cell Engagers for the Treatment of Multiple Myeloma: Achievements and Challenges. Cancers 2021, 13, 2853. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, A.; Pelletier, L.; Giustiniani, J.; Kim, P.; Bisht, K.; Wang, H.; Van de Velde, H.J.; Haioun, C.; Gaulard, P.; Ortonne, N. The CD38/CD3xCD28 Trispecific Antibody (SAR442257) Potentially Represents a Novel Therapeutic Strategy for Peripheral T-Cell Lymphomas. Blood 2023, 142, 4384. [Google Scholar] [CrossRef]
- Green, M.R.; Reville, P.K.; Dai, E.; Sheikh, I.; Deng, Q.; Henderson, J.; Le, C.; Rojas, E.; Okwuchi, C.; Wilson, A.; et al. SAR442257, a CD38/CD28/CD3 trispecific antibody, potentiates CAR T-cell activity against large B-cell lymphoma. Hematol. Oncol. 2023, 41, 275–276. [Google Scholar] [CrossRef]
- Viola, D.; Dona, A.; Caserta, E.; Troadec, E.; Besi, F.; McDonald, T.; Ghoda, L.; Gunes, E.G.; Sanchez, J.F.; Khalife, J.; et al. Daratumumab induces mechanisms of immune activation through CD38+ NK cell targeting. Leukemia 2021, 35, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.G.; Facon, T.; Bensinger, W.I.; Leleu, X.; Campana, F.; Macé, S.; Chiron, M.; van de Velde, H.; Mikhael, J. Predictive biomarkers with isatuximab plus pomalidomide and dexamethasone in relapsed/refractory multiple myeloma. Blood Cancer J. 2021, 11, 55. [Google Scholar] [CrossRef]
- Chen, L.; Qian, W.; Pan, F.; Li, D.; Yu, W.; Tong, L.; Yang, Y.; Xu, Q.; Ding, J.; Dai, R.; et al. A trispecific antibody induces potent tumor-directed T-cell activation and antitumor activity by CD3/CD28 co-engagement. Immunotherapy 2024, 16, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Krejcik, J.; Casneuf, T.; Nijhof, I.S.; Verbist, B.; Bald, J.; Plesner, T.; Syed, K.; Liu, K.; van de Donk, N.W.; Weiss, B.M.; et al. Daratumumab depletes CD38+ immune regulatory cells, promotes T-cell expansion, and skews T-cell repertoire in multiple myeloma. Blood 2016, 128, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Alrasheed, N.; Lee, L.; Ghorani, E.; Henry, J.Y.; Conde, L.; Chin, M.; Galas-Filipowicz, D.; Furness, A.J.S.; Chavda, S.J.; Richards, H.; et al. Marrow-Infiltrating Regulatory T Cells Correlate with the Presence of Dysfunctional CD4+PD-1+ Cells and Inferior Survival in Patients with Newly Diagnosed Multiple Myeloma. Clin. Cancer Res. 2020, 26, 3443–3454. [Google Scholar] [CrossRef] [PubMed]
- Rana, P.S.; Soler, D.C.; Kort, J.; Driscoll, J.J. Targeting TGF-beta signaling in the multiple myeloma microenvironment: Steering CARs and T cells in the right direction. Front. Cell Dev. Biol. 2022, 10, 1059715. [Google Scholar] [CrossRef] [PubMed]
- David, C.J.; Massagué, J. Contextual determinants of TGFβ action in development, immunity and cancer. Nat. Rev. Mol. Cell Biol. 2018, 19, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Park, J.; Cho, I.; Sheen, Y. Co-treatment with vactosertib, a novel, orally bioavailable activin receptor-like kinase 5 inhibitor, suppresses radiotherapy-induced epithelial-to-mesenchymal transition, cancer cell stemness, and lung metastasis of breast cancer. Radiol. Oncol. 2022, 56, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Russ, A.; Hua, A.B.; Montfort, W.R.; Rahman, B.; Riaz, I.B.; Khalid, M.U.; Carew, J.S.; Nawrocki, S.T.; Persky, D.; Anwer, F. Blocking “don’t eat me” signal of CD47-SIRPalpha in hematological malignancies, an in-depth review. Blood Rev. 2018, 32, 480–489. [Google Scholar] [CrossRef] [PubMed]
- You, T.; Hu, W.; Ge, X.; Shen, J.; Qin, X. Application of a novel inhibitor of human CD59 for the enhancement of complement-dependent cytolysis on cancer cells. Cell Mol. Immunol. 2011, 8, 157–163. [Google Scholar] [CrossRef]
- van de Donk, N.; Usmani, S.Z. CD38 Antibodies in Multiple Myeloma: Mechanisms of Action and Modes of Resistance. Front. Immunol. 2018, 9, 2134. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Hughes, T.; Zhang, J.; Caligiuri, M.A.; Benson, D.M.; Yu, J. Fratricide of NK Cells in Daratumumab Therapy for Multiple Myeloma Overcome by Ex Vivo-Expanded Autologous NK Cells. Clin. Cancer Res. 2018, 24, 4006–4017. [Google Scholar] [CrossRef]
- Tang, Z.; Wang, X.; Tang, M.; Wu, J.; Zhang, J.; Liu, X.; Gao, F.; Fu, Y.; Tang, P.; Li, C. Overcoming the On-Target Toxicity in Antibody-Mediated Therapies via an Indirect Active Targeting Strategy. Adv. Sci. 2023, 10, 2206912. [Google Scholar] [CrossRef] [PubMed]
- Tyrsin, D.; Chuvpilo, S.; Matskevich, A.; Nemenov, D.; Römer, P.S.; Tabares, P.; Hünig, T. From TGN1412 to TAB08: The return of CD28 superagonist therapy to clinical development for the treatment of rheumatoid arthritis. Clin. Exp. Rheumatol. 2016, 34, 45–48. [Google Scholar] [PubMed]
- Gogesch, P.; Dudek, S.; van Zandbergen, G.; Waibler, Z.; Anzaghe, M. The Role of Fc Receptors on the Effectiveness of Therapeutic Monoclonal Antibodies. Int. J. Mol. Sci. 2021, 22, 8947. [Google Scholar] [CrossRef] [PubMed]
- Ben Mkaddem, S.; Benhamou, M.; Monteiro, R.C. Understanding Fc Receptor Involvement in Inflammatory Diseases: From Mechanisms to New Therapeutic Tools. Front. Immunol. 2019, 10, 445798. [Google Scholar] [CrossRef]
- Alegre, M.L.; Peterson, L.J.; Xu, D.; Sattar, H.A.; Jeyarajah, D.R.; Kowalkowski, K.; Thistlethwaite, J.R.; Zivin, R.A.; Jolliffe, L.; Bluestone, J.A. A non-activating “humanized” anti-CD3 monoclonal antibody retains immunosuppressive properties in vivo. Transplantation 1994, 57, 1537–1543. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Company | Cat. No. |
|---|---|---|
| SAR442257 | Sanofi (MA, USA) | Not applicable (N/A) |
| CD45 | Invitrogen (Basel, Switzerland) | MHCD4528 |
| PD-1 (=CD279) | BD (Heidelberg, Germany) | 560,652 |
| CD3 | BD (Heidelberg, Germany) | 641,397 |
| CD4 | BD (Heidelberg, Germany) | 562,970 |
| CD8 | BD (Heidelberg, Germany) | 341,050 |
| HLA-DR | BD (Heidelberg, Germany) | 556,643 |
| CD57 | BD (Heidelberg, Germany) | 560,845 |
| KI67 | BD (Heidelberg, Germany) | 567,120 |
| TIGIT | BioLegend (Amsterdam, The Netherlands) | 372,709 |
| CD25 | BD (Heidelberg, Germany) | 557,741 |
| FOXP3 | Bioscience/Affimetrix (Heidelberg, Germany) | 11-4776-42 |
| CD69 | BD (Heidelberg, Germany) | 560,711 |
| CD127 | ThermoFisher (Darmstadt, Germany) | BDB-557938 |
| CD45RA | BD (Heidelberg, Germany) | 740,083 |
| CD27 | BD (Heidelberg, Germany) | 567,289 |
| Granzyme | BD (Heidelberg, Germany) | 558,905 |
| CCR7 | BD (Heidelberg, Germany) | 566,762 |
| Perforin | BD (Heidelberg, Germany) | 556,437 |
| 4-1BB Cd137 | BD (Heidelberg, Germany) | 741,000 |
| CD14 | BD (Heidelberg, Germany) | 740,163 |
| CD16 | BD (Heidelberg, Germany) | 338,426 |
| CD56 | BD (Heidelberg, Germany) | 555,518 |
| CD28 | BD (Heidelberg, Germany) | 556,621 |
| CD138 | BD (Heidelberg, Germany) | 650,660 |
| CD20 | BD (Heidelberg, Germany) | 560,736 |
| CD319 | Biomol (Hamburg, Germany) | ABD-131901C1 |
| CD28 | BD (Heidelberg, Germany) | 556,621 |
| CD19 | BD (Heidelberg, Germany) | 557,835 |
| CD107a | BD (Heidelberg, Germany) | 555,800 |
| anti-CD138 | Miltenyi Biotec (Bergisch Gladbach, Germany) | 130-051-301 |
| Buffy coat | DRK Baden–Württemberg–Hessen GmbH (Mannheim, Germany) | N/A |
| Patient material | University Hospital Heidelberg (Heidelberg, Germany) | N/A |
| QiFIKIT beads | Dako (Glostrup, Denmark) | K007811 |
| PE/R-Phycoerythrin Conjugation Kit | Abcam (Amsterdam, The Netherlands) | ab102918 |
| Transcription Factor Buffer Set | BD (Heidelberg, Germany) | 562,574 |
| Brilliant Stain Buffer Plus | BD (Heidelberg, Germany) | 566,385 |
| Legendplex | BioLegend (Amsterdam, The Netherlands) | 741,157 |
| autoMACS Pro purification | Miltenyi Biotec (Bergisch Gladbach, Germany) | N/A |
| MM1S | DSMZ (Braunschweig, Germany) | N/A |
| U266 | DSMZ (Braunschweig, Germany) | N/A |
| AMO | DSMZ (Braunschweig, Germany) | N/A |
| AMO-TP53 | DSMZ (Braunschweig, Germany) | N/A |
| RPMI 8226 | DSMZ (Braunschweig, Germany) | N/A |
| INA-6 | DSMZ (Braunschweig, Germany) | N/A |
| OPM-2 | DSMZ (Braunschweig, Germany) | N/A |
| LP-1 | DSMZ (Braunschweig, Germany) | N/A |
| KMS-11BM | DSMZ (Braunschweig, Germany) | N/A |
| KMS-11 | DSMZ (Braunschweig, Germany) | N/A |
| Origin | ADDITIVE GmbH (Friedrichsdorf, Germany) | N/A |
| Ocean software | LUMICKS (Amsterdam, The Netherlands) | N/A |
| RPMI 1640 | Gibco (ThermoFisher, Darmstadt, Germany) | 11875-093 |
| FBS | Gibco (ThermoFisher, Darmstadt, Germany) | 10270-106 |
| Vactoserib | Selleckchem (Cologne, Germany) | S7530 |
| TGF-β | Selleckchem (Cologne, Germany) | A2113 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grab, A.L.; Kim, P.S.; John, L.; Bisht, K.; Wang, H.; Baumann, A.; Van de Velde, H.; Sarkar, I.; Shome, D.; Reichert, P.; et al. Pre-Clinical Assessment of SAR442257, a CD38/CD3xCD28 Trispecific T Cell Engager in Treatment of Relapsed/Refractory Multiple Myeloma. Cells 2024, 13, 879. https://doi.org/10.3390/cells13100879
Grab AL, Kim PS, John L, Bisht K, Wang H, Baumann A, Van de Velde H, Sarkar I, Shome D, Reichert P, et al. Pre-Clinical Assessment of SAR442257, a CD38/CD3xCD28 Trispecific T Cell Engager in Treatment of Relapsed/Refractory Multiple Myeloma. Cells. 2024; 13(10):879. https://doi.org/10.3390/cells13100879
Chicago/Turabian StyleGrab, Anna Luise, Peter S. Kim, Lukas John, Kamlesh Bisht, Hongfang Wang, Anja Baumann, Helgi Van de Velde, Irene Sarkar, Debarati Shome, Philipp Reichert, and et al. 2024. "Pre-Clinical Assessment of SAR442257, a CD38/CD3xCD28 Trispecific T Cell Engager in Treatment of Relapsed/Refractory Multiple Myeloma" Cells 13, no. 10: 879. https://doi.org/10.3390/cells13100879
APA StyleGrab, A. L., Kim, P. S., John, L., Bisht, K., Wang, H., Baumann, A., Van de Velde, H., Sarkar, I., Shome, D., Reichert, P., Manta, C., Gryzik, S., Reijmers, R. M., Weinhold, N., & Raab, M. S. (2024). Pre-Clinical Assessment of SAR442257, a CD38/CD3xCD28 Trispecific T Cell Engager in Treatment of Relapsed/Refractory Multiple Myeloma. Cells, 13(10), 879. https://doi.org/10.3390/cells13100879