
Cannabidiol Exerts Anticonvulsant Effects Alone and in Combination with Δ9-THC through the 5-HT1A Receptor in the Neocortex of Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Preparation
2.2. Cortical Slice Preparation
2.3. Electrophysiology
2.4. Materials
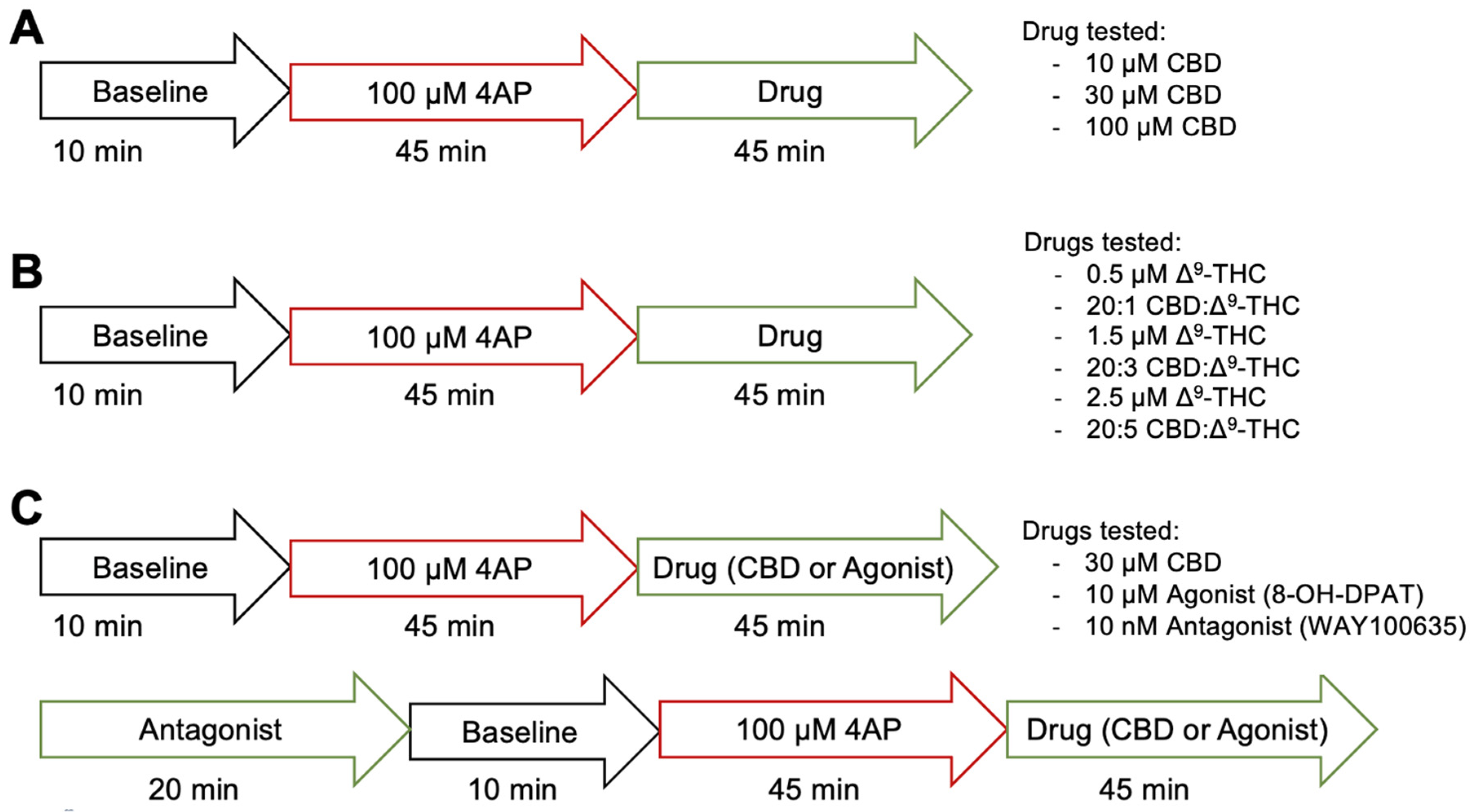
2.5. Protocol
2.6. Data and Statistical Analysis
3. Results
3.1. AIM #1—Anticonvulsant Effect of CBD Alone
Extracellular Effect of CBD on Seizure-Like Events (SLE) and Inter-Ictal Bursting Events
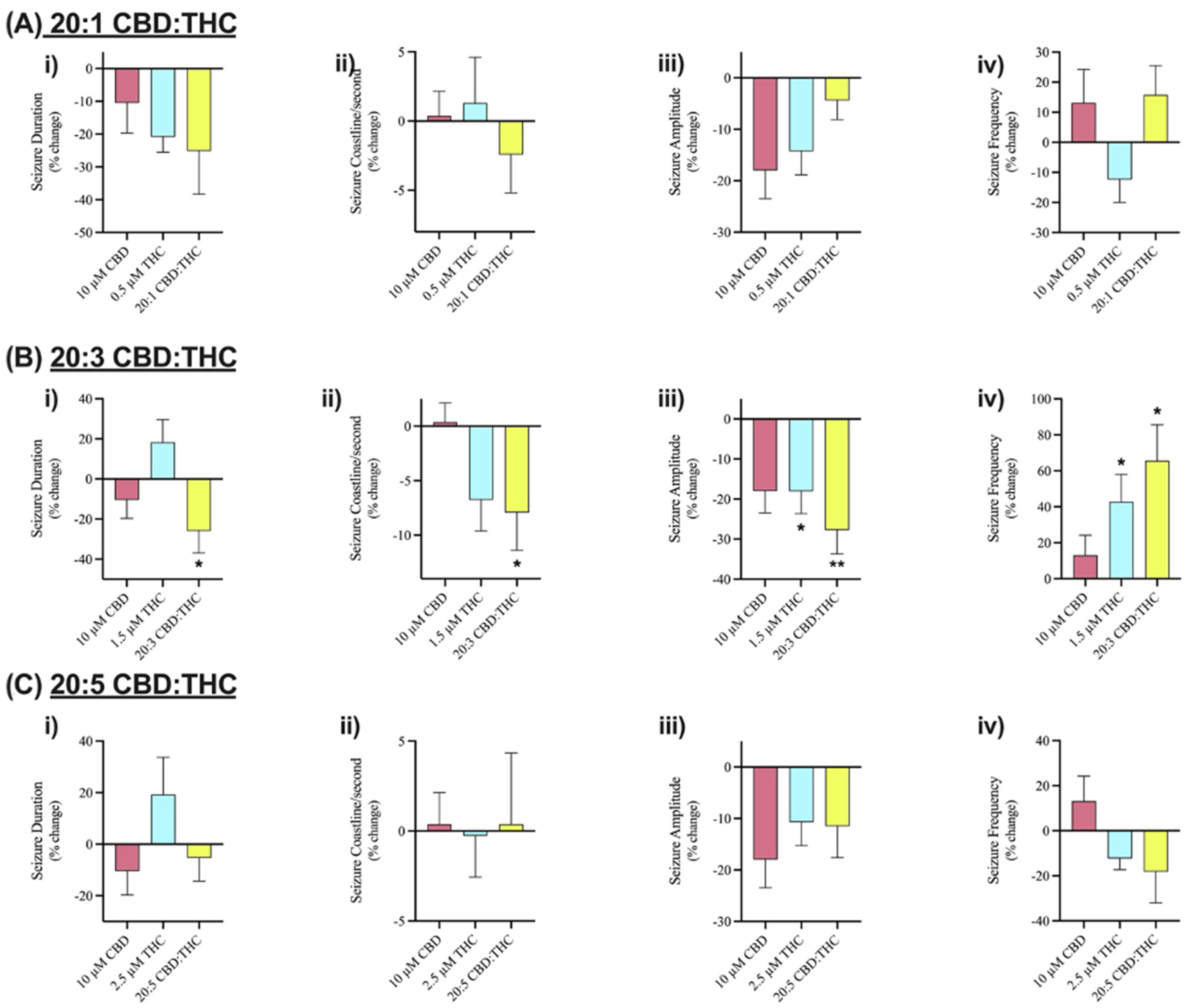
3.2. AIM #2—Anticonvulsant Effect of CBD Combined with Δ9-THC
Extracellular Effect of CBD Combined with Δ9-THC on SLE
3.3. AIM #3—CBD’s Mechanism of Action
Extracellular Effect of Blocking 5-HT1A Receptor on SLE
4. Discussion
4.1. The Anticonvulsant Effect of CBD Alone
4.2. The Anticonvulsant Effect of CBD Combined with Δ9-THC
4.3. The Role of 5HT1A Receptors in CBD’s Mechanism of Action
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kwan, P.; Brodie, M.J. Early Identification of Refractory Epilepsy. N. Engl. J. Med. 2000, 342, 314–319. [Google Scholar] [CrossRef] [PubMed]
- DeGiorgio, C.M.; Markovic, D.; Mazumder, R.; Moseley, B.D. Ranking the Leading Risk Factors for Sudden Unexpected Death in Epilepsy. Front. Neurol. 2017, 8, 473. [Google Scholar] [CrossRef]
- Mechoulam, R.; Shani, A.; Edery, H.; Grunfeld, Y. Chemical Basis of Hashish Activity. Science (1979) 1970, 169, 611–612. [Google Scholar] [CrossRef]
- Hill, A.; Mercier, M.; Hill, T.; Glyn, S.; Jones, N.; Yamasaki, Y.; Futamura, T.; Duncan, M.; Stott, C.; Stephens, G.; et al. Cannabidivarin Is Anticonvulsant in Mouse and Rat. Br. J. Pharmacol. 2012, 167, 1629. [Google Scholar] [CrossRef]
- Hill, A.J.; Jones, N.A.; Smith, I.; Hill, C.L.; Williams, C.M.; Stephens, G.J.; Whalley, B.J. Voltage-Gated Sodium (Na V) Channel Blockade by Plant Cannabinoids Does Not Confer Anticonvulsant Effects per Se. Neurosci. Lett. 2014, 566, 269–274. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Hill, C.L.; Leo, A.; Alhusaini, A.; Soubrane, C.; Mazzarella, E.; Russo, E.; Whalley, B.J.; Marzo, V.D.; Stephens, G.J. Nonpsychotropic Plant Cannabinoids, Cannabidivarin (CBDV) and Cannabidiol (CBD), Activate and Desensitize Transient Receptor Potential Vanilloid 1 (TRPV1) Channels In Vitro: Potential for the Treatment of Neuronal Hyperexcitability. CS Chem. Neurosci. 2014, 5, 1131–1141. [Google Scholar] [CrossRef]
- Jones, N.A.; Hill, A.J.; Smith, I.; Bevan, S.A.; Williams, C.M.; Whalley, B.J.; Stephens, G.J. Cannabidiol Displays Antiepileptiform and Antiseizure Properties In Vitro and In Vivo. J. Pharmacol. Exp. Ther. 2010, 332, 569–577. [Google Scholar] [CrossRef]
- Jones, N.A.; Glyn, S.E.; Akiyama, S.; Hill, T.D.M.; Hill, A.J.; Weston, S.E.; Burnett, M.D.A.; Yamasaki, Y.; Stephens, G.J.; Whalley, B.J.; et al. Cannabidiol Exerts Anti-Convulsant Effects in Animal Models of Temporal Lobe and Partial Seizures. Seizure 2012, 21, 344–352. [Google Scholar] [CrossRef]
- Vieira, I.; Lima, A.; Quaglio Bellozi, P.M.; Marques Batista, E.; Rezende Vilela, L.; Lucas Brandão, I.; Ribeiro, M.; Flávio, M.; Moraes, D.; Moreira, F.A.; et al. Cannabidiol Anticonvulsant Effect Is Mediated by the PI3Kγ Pathway. Neuropharmacology 2020, 176, 108156. [Google Scholar] [CrossRef]
- Devinsky, O.; Cross, J.H.; Laux, L.; Marsh, E.; Miller, I.; Nabbout, R.; Scheffer, I.E.; Thiele, E.A.; Wright, S. Trial of Cannabidiol for Drug-Resistant Seizures in the Dravet Syndrome. N. Engl. J. Med. 2017, 376, 2011–2020. [Google Scholar] [CrossRef]
- Devinsky, O.; Marsh, E.; Friedman, D.; Thiele, E.; Laux, L.; Sullivan, J.; Miller, I.; Flamini, R.; Wilfong, A.; Filloux, F.; et al. Cannabidiol in Patients with Treatment-Resistant Epilepsy: An Open-Label Interventional Trial. Lancet Neurol. 2016, 15, 270–278. [Google Scholar] [CrossRef]
- Devinsky, O.; Patel, A.D.; Cross, J.H.; Villanueva, V.; Wirrell, E.C.; Privitera, M.; Greenwood, S.M.; Roberts, C.; Checketts, D.; VanLandingham, K.E.; et al. Effect of Cannabidiol on Drop Seizures in the Lennox–Gastaut Syndrome. N. Engl. J. Med. 2018, 378, 1888–1897. [Google Scholar] [CrossRef]
- Miller, I.; Scheffer, I.E.; Gunning, B.; Sanchez-Carpintero, R.; Gil-Nagel, A.; Perry, M.S.; Saneto, R.P.; Checketts, D.; Dunayevich, E.; Knappertz, V. Dose-Ranging Effect of Adjunctive Oral Cannabidiol vs Placebo on Convulsive Seizure Frequency in Dravet Syndrome: A Randomized Clinical Trial. JAMA Neurol. 2020, 77, 613–621. [Google Scholar] [CrossRef]
- Crippa, J.A.S.; Crippa, A.C.S.; Hallak, J.E.C.; Martín-Santos, R.; Zuardi, A.W. Δ9-THC Intoxication by Cannabidiol-Enriched Cannabis Extract in Two Children with Refractory Epilepsy: Full Remission after Switching to Purified Cannabidiol. Front. Pharmacol. 2016, 7, 359. [Google Scholar] [CrossRef]
- Sekar, K.; Pack, A. Epidiolex as Adjunct Therapy for Treatment of Refractory Epilepsy: A Comprehensive Review with a Focus on Adverse Effects. F1000Research 2019, 8, 234. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Sinha, J.K.; Ghosh, S.; Sharma, H.; Bhaskar, R.; Narayanan, K.B. A Comprehensive Review of Emerging Trends and Innovative Therapies in Epilepsy Management. Brain Sci. 2023, 13, 1305. [Google Scholar] [CrossRef]
- Devinsky, O.; Cilio, M.R.; Cross, H.; Fernandez-Ruiz, J.; French, J.; Hill, C.; Katz, R.; Di Marzo, V.; Jutras-Aswad, D.; Notcutt, W.G.; et al. Cannabidiol: Pharmacology and Potential Therapeutic Role in Epilepsy and Other Neuropsychiatric Disorders. Epilepsia 2014, 55, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Machado Bergamaschi, M.; Helena Costa Queiroz, R.; Waldo Zuardi, A.; Alexandre, S.; Crippa, J. Safety and Side Effects of Cannabidiol, a Cannabis Sativa Constituent. Curr. Drug Saf. 2011, 6, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Ham, M.T.; Loskota, W.J.; Lomax, P. Acute and Chronic Effects of Δ9-Tetrahydrocannabinol on Seizures in the Gerbil. Eur. J. Pharmacol. 1975, 31, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.J.; Weston, S.E.; Jones, N.A.; Smith, I.; Bevan, S.A.; Williamson, E.M.; Stephens, G.J.; Williams, C.M.; Whalley, B.J. 9-Tetrahydrocannabivarin Suppresses in Vitro Epileptiform and in Vivo Seizure Activity in Adult Rats. Epilepsia 2010, 51, 1522–1532. [Google Scholar] [CrossRef]
- Wallace, M.J.; Wiley, J.L.; Martin, B.R.; Delorenzo, R.J. Assessment of the Role of CB Receptors in Cannabinoid 1 Anticonvulsant Effects. Eur. J. Pharmacol. 2001, 428, 51–57. [Google Scholar] [CrossRef]
- Kujtan, P.W.; Carlen, P.L.; Kapur, B.M. Δ9-Tetrahydrocannabinol and Cannabinol: Dose-Dependent Effects on Evoked Potentials in the Hippocampal Slice. Can. J. Physiol. Pharmacol. 1983, 61, 420–426. [Google Scholar] [CrossRef]
- Ebbert, J.O.; Scharf, E.L.; Hurt, R.T. Medical Cannabis. Mayo Clin. Proc. 2018, 93, 1842–1847. [Google Scholar] [CrossRef]
- Maa, E.; Figi, P. The Case for Medical Marijuana in Epilepsy. Epilepsia 2014, 55, 783–786. [Google Scholar] [CrossRef]
- Press, C.A.; Knupp, K.G.; Chapman, K.E. Parental Reporting of Response to Oral Cannabis Extracts for Treatment of Refractory Epilepsy. Epilepsy Behav. 2015, 45, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Porter, B.E.; Jacobson, C. Report of a Parent Survey of Cannabidiol-Enriched Cannabis Use in Pediatric Treatment-Resistant Epilepsy. Epilepsy Behav. 2013, 29, 574–577. [Google Scholar] [CrossRef]
- Hussain, S.A.; Zhou, R.; Jacobson, C.; Weng, J.; Cheng, E.; Lay, J.; Hung, P.; Lerner, J.T.; Sankar, R. Perceived Efficacy of Cannabidiol-Enriched Cannabis Extracts for Treatment of Pediatric Epilepsy: A Potential Role for Infantile Spasms and Lennox-Gastaut Syndrome. Epilepsy Behav. 2015, 47, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Fallah, M.S.; Dlugosz, L.; Scott, B.W.; Thompson, M.D.; Burnham, W.M.I. Antiseizure Effects of the Cannabinoids in the Amygdala-Kindling Model. Epilepsia 2021, 62, 2274–2282. [Google Scholar] [CrossRef] [PubMed]
- Dlugosz, L.; Zhou, H.Z.; Scott, B.W.; Burnham, M. The Effects of Cannabidiol and Δ9-Tetrahydrocannabinol, Alone and in Combination, in the Maximal Electroshock Seizure Model. Epilepsy Res. 2023, 190, 107087. [Google Scholar] [CrossRef] [PubMed]
- Hausman-Kedem, M.; Menascu, S.; Kramer, U. Efficacy of CBD-Enriched Medical Cannabis for Treatment of Refractory Epilepsy in Children and Adolescents—An Observational, Longitudinal Study. Brain Dev. 2018, 40, 544–551. [Google Scholar] [CrossRef]
- Athnaid Mccoy, B.; Wang, L.; Zak, M.; Al-Mehmadi, S.; Kabir, N.; Alhadid, K.; Mcdonald, K.; Zhang, G.; Sharma, R.; Whitney, R.; et al. A Prospective Open-Label Trial of a CBD/THC Cannabis Oil in Dravet Syndrome. Ann. Clin. Transl. Neurol. 2018, 5, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Wade, D.T.; Makela, P.M.; House, H.; Bateman, C.; Robson, P. Long-Term Use of a Cannabis-Based Medicine in the Treatment of Spasticity and Other Symptoms in Multiple Sclerosis. Mult. Scler. 2006, 12, 639–645. [Google Scholar] [CrossRef]
- Morano, A.; Fanella, M.; Albini, M.; Cifelli, P.; Palma, E.; Giallonardo, A.T.; Di Bonaventura, C. Cannabinoids in the Treatment of Epilepsy: Current Status and Future Prospects. Neuropsychiatr. Dis. Treat. 2020, 16, 381–396. [Google Scholar] [CrossRef]
- Devinsky, O.; Elder, C.; Sivathamboo, S.; Scheffer, I.E.; Koepp, M.J. Idiopathic Generalized Epilepsy: Misunderstandings, Challenges, and Opportunities. Neurology 2024, 102, e208076. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Arellano, J.; Canseco-Alba, A.; Cutler, S.J.; León, F. The Polypharmacological Effects of Cannabidiol. Molecules 2023, 28, 3271. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.X.B.; Heckman, L.; Niday, Z.; Jo, S.; Fujita, A.; Shim, J.; Pandey, R.; Al Jandal, H.; Jayakar, S.; Barrett, L.B.; et al. Cannabidiol Activates Neuronal Kv7 Channels. Elife 2022, 11, 3271. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.A.; Whalley, B.J. The Proposed Mechanisms of Action of CBD in Epilepsy. Epileptic Disord. 2020, 22, 10–15. [Google Scholar] [CrossRef]
- Reddy, D.S. Therapeutic and Clinical Foundations of Cannabidiol Therapy for Difficult-to-Treat Seizures in Children and Adults with Refractory Epilepsies. Exp. Neurol. 2023, 359, 114237. [Google Scholar] [CrossRef]
- Kaplan, J.S.; Stella, N.; Catterall, W.A.; Westenbroek, R.E. Cannabidiol Attenuates Seizures and Social Deficits in a Mouse Model of Dravet Syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, 11229–11234. [Google Scholar] [CrossRef]
- Gray, R.A.; Stott, C.G.; Jones, N.A.; Di Marzo, V.; Whalley, B.J.; Whalley, B.J. Anticonvulsive Properties of Cannabidiol in a Model of Generalized Seizure Are Transient Receptor Potential Vanilloid 1 Dependent. Cannabis Cannabinoid Res. 2020, 5, 145. [Google Scholar] [CrossRef]
- Hernandez, E.J.; Williams, P.A.; Dudek, F.E. Effects of Fluoxetine and TFMPP on Spontaneous Seizures in Rats with Pilocarpine-Induced Epilepsy. Epilepsia 2002, 43, 1337–1345. [Google Scholar] [CrossRef]
- Tokarski, K.; Zahorodna, A.; Bobula, B.; Hess, G. Comparison of the Effects of 5-HT 1A and 5-HT 4 Receptor Activation on Field Potentials and Epileptiform Activity in Rat Hippocampus. Exp. Brain Res. 2002, 147, 505–510. [Google Scholar] [CrossRef]
- Gariboldi, M.; Tutka, P.; Samanin, R.; Vezzani, A. Stimulation of 5-HTlA Receptors in the Dorsal Hippocampus and Inhibition of Limbic Seizures Induced by Kainic Acid in Rats; Stockton Press: Stockton, CA, USA, 1996. [Google Scholar]
- Sourbron, J.; Schneider, H.; Kecske, A.; Liu, Y.; Buening, E.M.; Lagae, L.; Smolders, I.; de Witte, P. Serotonergic Modulation as Effective Treatment for Dravet Syndrome in a Zebrafish Mutant Model. ACS Chem. Neurosci. 2016, 7, 35. [Google Scholar] [CrossRef]
- Russo, E.B.; Burnett, A.; Hall, B.; Parker, K.K. Agonistic Properties of Cannabidiol at 5-HT1a Receptors. Neurochem. Res. 2005, 30, 1037–1043. [Google Scholar] [CrossRef]
- Maggio, N.; Stein, S.; Segal, M.; Alzheimer, C.; Nugent, F.S.; Hebert, F.E.; Wagner, J.J. Cannabidiol Regulates Long Term Potentiation Following Status Epilepticus: Mediation by Calcium Stores and Serotonin. Front. Mol. Neurosci. 2018, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Muñoz, M.; Onetti, Y.; Cortés-Montero, E.; Garzón, J.; Sánchez-Blázquez, P. Cannabidiol Enhances Morphine Antinociception, Diminishes NMDA-Mediated Seizures and Reduces Stroke Damage via the Sigma 1 Receptor. Mol. Brain 2018, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Ledgerwood, C.J.; Greenwood, S.M.; Brett, R.R.; Pratt, J.A.; Bushell, T.J.; Bushell, T.J. Cannabidiol Inhibits Synaptic Transmission in Rat Hippocampal Cultures and Slices via Multiple Receptor Pathways. Br. J. Pharmacol. 2010, 162, 286–294. [Google Scholar] [CrossRef]
- Pelz, M.C.; Schoolcraft, K.D.; Larson, C.; Spring, M.G.; López, H.H. Assessing the Role of Serotonergic Receptors in Cannabidiol’s Anticonvulsant Efficacy. Epilepsy Behav. 2017, 73, 111–118. [Google Scholar] [CrossRef]
- Raimondo, J.V.; Heinemann, U.; de Curtis, M.; Goodkin, H.P.; Dulla, C.G.; Janigro, D.; Ikeda, A.; Lin, C.C.K.; Jiruska, P.; Galanopoulou, A.S.; et al. Methodological Standards for In Vitro Models of Epilepsy and Epileptic Seizures. A TASK1-WG4 Report of the AES/ILAE Translational Task Force of the ILAE. Epilepsia 2017, 58 (Suppl. S4), 40–52. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y.; Holmes, G.L. Effects of Seizures on Developmental Processes in the Immature Brain. Lancet Neurol. 2006, 5, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Luk, W.P.; Gillis, J.; Skinner, F.; Zhang, L. Size Does Matter: Generation of Intrinsic Network Rhythms in Thick Mouse Hippocampal Slices. J. Neurophysiol. 2005, 93, 2302–2317. [Google Scholar] [CrossRef][Green Version]
- Huntsman, R.J.; Tang-Wai, R.; Shackelford, A.E. Cannabis for Pediatric Epilepsy. J. Clin. Neurophysiol. 2020, 37, 2–8. [Google Scholar] [CrossRef]
- Kabat, J.; Król, P. Focal Cortical Dysplasia—Review. Pol. J. Radiol. 2012, 77, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Shekh-Ahmad, T.; Khalil, A.; Walker, M.C.; Ali, A.B. Cannabidiol Exerts Antiepileptic Effects by Restoring Hippocampal Interneuron Functions in a Temporal Lobe Epilepsy Model Background and Purpose. Br. J. Pharmacol. 2018, 175, 2097. [Google Scholar] [CrossRef]
- Avoli, M.; Perreault, P.; Olivier, A.; Villemure, J.G. 4-Aminopyridine Induces a Long-Lasting Depolarizing GABA-Ergic Potential in Human Neocortical and Hippocampal Neurons Maintained In Vitro. Neurosci. Lett. 1988, 94, 327–332. [Google Scholar] [CrossRef]
- Avoli, M.; de Curtis, M. GABAergic Synchronization in the Limbic System and Its Role in the Generation of Epileptiform Activity. Prog. Neurobiol. 2011, 95, 104–132. [Google Scholar] [CrossRef]
- Anderson, L.L.; Low, I.K.; McGregor, I.S.; Arnold, J.C. Interactions between Cannabidiol and Δ9-Tetrahydrocannabinol in Modulating Seizure Susceptibility and Survival in a Mouse Model of Dravet Syndrome. Br. J. Pharmacol. 2020, 177, 4261–4274. [Google Scholar] [CrossRef]
- Hosseinzadeh, M.; Nikseresht, S.; Khodagholi, F.; Naderi, N.; Maghsoudi, N. Cannabidiol Post-Treatment Alleviates Rat Epileptic-Related Behaviors and Activates Hippocampal Cell Autophagy Pathway Along with Antioxidant Defense in Chronic Phase of Pilocarpine-Induced Seizure. J. Mol. Neurosci. 2016, 58, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.J.; Koifman, J.; Shin, D.S.; Ye, H.; Florez, C.M.; Zhang, L.; Valiante, T.A.; Carlen, P.L. Transition to Seizure: Ictal Discharge Is Preceded by Exhausted Presynaptic GABA Release in the Hippocampal CA3 Region. J. Neurosci. 2012, 32, 2499–2512. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.D.; Whalley, B.J.; Baker, D.; Pryce, G.; Constanti, A.; Gibbons, S.; Williamson, E.M. Medicinal Cannabis: Is Δ9–Tetrahydrocannabinol Necessary for All Its Effects? J. Pharm. Pharmacol. 2003, 55, 1687–1694. [Google Scholar] [CrossRef] [PubMed]
- Samarut, É.; Nixon, J.; Kundap, U.P.; Drapeau, P.; Ellis, L.D. Single and Synergistic Effects of Cannabidiol and δ-9-Tetrahydrocannabinol on Zebrafish Models of Neuro-Hyperactivity. Front. Pharmacol. 2019, 10, 226. [Google Scholar] [CrossRef]
- Aguirre-Velázquez, C.G. Report from a Survey of Parents Regarding the Use of Cannabidiol (Medicinal Cannabis) in Mexican Children with Refractory Epilepsy. Neurol. Res. Int. 2017, 2017, 2985729. [Google Scholar] [CrossRef]
- Hall, W.; Solowij, N. Adverse Effects of Cannabis. Lancet 1998, 352, 1611–1616. [Google Scholar] [CrossRef]
- Volkow, N.D.; Baler, R.D.; Compton, W.M.; Weiss, S.R.B. Adverse Health Effects of Marijuana Use. N. Engl. J. Med. 2014, 370, 2219–2227. [Google Scholar] [CrossRef]
- Klotz, K.A.; Grob, D.; Hirsch, M.; Metternich, B.; Schulze-Bonhage, A.; Jacobs, J. Efficacy and Tolerance of Synthetic Cannabidiol for Treatment of Drug Resistant Epilepsy. Front. Neurol. 2019, 10, 1313. [Google Scholar] [CrossRef]
- Boggs, D.L.; Peckham, A.; Boggs, A.A.; Ranganathan, M. Delta-9-Tetrahydrocannabinol and Cannabidiol: Separating the Chemicals from the “Weed”, a Pharmacodynamic Discussion. Ment. Health Clin. 2016, 6, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Britch, S.C.; Wiley, J.L.; Yu, Z.; Clowers, B.H.; Craft, R.M. Cannabidiol-Δ9-Tetrahydrocannabinol Interactions on Acute Pain and Locomotor Activity. Drug Alcohol. Depend. 2017, 175, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Hložek, T.; Uttl, L.; Kadeřábek, L.; Balíková, M.; Lhotková, E.; Horsley, R.R.; Nováková, P.; Šíchová, K.; Štefková, K.; Tylš, F.; et al. Pharmacokinetic and Behavioural Profile of THC, CBD, and THC+CBD Combination after Pulmonary, Oral, and Subcutaneous Administration in Rats and Confirmation of Conversion in Vivo of CBD to THC. Eur. Neuropsychopharmacol. 2017, 27, 1223–1237. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Karanges, E.; Spiro, A.; Wong, A.; Spencer, J.; Huynh, T.; Gunasekaran, N.; Karl, T.; Long, L.E.; Huang, X.F.; et al. Cannabidiol Potentiates Δ 9-Tetrahydrocannabinol (THC) Behavioural Effects and Alters THC Pharmacokinetics during Acute and Chronic Treatment in Adolescent Rats. Psychopharmacology 2011, 218, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Nadulski, T.; Pragst, F.; Weinberg, G.; Roser, P.; Schnelle, M.; Fronk, E.M.; Stadelmann, A.M. Randomized, Double-Blind, Placebo-Controlled Study about the Effects of Cannabidiol (CBD) on the Pharmacokinetics of Δ9-Tetrahydrocannabinol (THC) after Oral Application of Thc Verses Standardized Cannabis Extract. Ther. Drug Monit. 2005, 27, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.; Guy, G.W. A Tale of Two Cannabinoids: The Therapeutic Rationale for Combining Tetrahydrocannabinol and Cannabidiol. Med. Hypotheses 2006, 66, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Theodore, W.H.; Wiggs, E.A.; Martinez, A.R.; Dustin, I.H.; Khan, O.I.; Appel, S.; Reeves-Tyer, P.; Sato, S. Serotonin 1A Receptors, Depression, and Memory in Temporal Lobe Epilepsy. Epilepsia 2012, 53, 129–133. [Google Scholar] [CrossRef]
- Theodore, W.H.; Hasler, G.; Giovacchini, G.; Kelley, K.; Reeves-Tyer, P.; Herscovitch, P.; Drevets, W. Reduced Hippocampal 5HT1A PET Receptor Binding and Depression in Temporal Lobe Epilepsy. Epilepsia 2007, 48, 1526–1530. [Google Scholar] [CrossRef]
- Lattanzi, S.; Trinka, E.; Russo, E.; Del Giovane, C.; Matricardi, S.; Meletti, S.; Striano, P.; Damavandi, P.T.; Silvestrini, M.; Brigo, F. Pharmacotherapy for Dravet Syndrome: A Systematic Review and Network Meta-Analysis of Randomized Controlled Trials. Drugs 2023, 83, 1409–1424. [Google Scholar] [CrossRef] [PubMed]
- Dini, G.; Di Cara, G.; Ferrara, P.; Striano, P.; Verrotti, A. Reintroducing Fenfluramine as a Treatment for Seizures: Current Knowledge, Recommendations and Gaps in Understanding. Neuropsychiatr. Dis. Treat. 2023, 19, 2013–2025. [Google Scholar] [CrossRef] [PubMed]
- Peričić, D.; Lazić, J.; Jazvinšćak Jembrek, M.; Švob Štrac, D. Stimulation of 5-HT1A Receptors Increases the Seizure Threshold for Picrotoxin in Mice. Eur. J. Pharmacol. 2005, 527, 105–110. [Google Scholar] [CrossRef]
- Espejo-Porras, F.; Fernández-Ruiz, J.; Pertwee, R.G.; Mechoulam, R.; García, C. Motor Effects of the Non-Psychotropic Phytocannabinoid Cannabidiol That Are Mediated by 5-HT1A Receptors. Neuropharmacology 2013, 75, 155–163. [Google Scholar] [CrossRef]
- Rodrigues da Silva, N.; Gomes, F.V.; Sonego, A.B.; da Silva, N.R.; Guimarães, F.S. Cannabidiol Attenuates Behavioral Changes in a Rodent Model of Schizophrenia through 5-HT1A, but Not CB1 and CB2 Receptors. Pharmacol. Res. 2020, 156, 104749. [Google Scholar] [CrossRef]
- Martínez-Aguirre, C.; Carmona-Cruz, F.; Velasco, A.L.; Velasco, F.; Aguado-Carrillo, G.; Cuéllar-Herrera, M.; Rocha, L. Cannabidiol Acts at 5-HT1A Receptors in the Human Brain: Relevance for Treating Temporal Lobe Epilepsy. Front. Behav. Neurosci. 2020, 14, 233. [Google Scholar] [CrossRef]
- Grabenstatter, H.L. A Clue to Seizure-Induced SUDEP Risk: Loss of Brainstem Serotonergic Control of Cardiorespiratory Function. Epilepsy Curr. 2016, 16, 411–413. [Google Scholar] [CrossRef]
- Richerson, G.B.; Buchanan, G.F. The Serotonin Axis: Shared Mechanisms in Seizures, Depression, and SUDEP. Epilepsia 2011, 52 (Suppl. S1), 28–38. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Ma, H.; Lian, X.; Gu, L.; Yu, Q.; Zhang, H. Activation of Central 5-HT2A Receptor in Brain Inhibited the Seizure-Induced 1 Respiratory Arrest in the DBA/1 Mouse SUDEP Model. bioRxiv bioRxiv:2020.12.04.410969. 2020. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javadzadeh, Y.; Santos, A.; Aquilino, M.S.; Mylvaganam, S.; Urban, K.; Carlen, P.L. Cannabidiol Exerts Anticonvulsant Effects Alone and in Combination with Δ9-THC through the 5-HT1A Receptor in the Neocortex of Mice. Cells 2024, 13, 466. https://doi.org/10.3390/cells13060466
Javadzadeh Y, Santos A, Aquilino MS, Mylvaganam S, Urban K, Carlen PL. Cannabidiol Exerts Anticonvulsant Effects Alone and in Combination with Δ9-THC through the 5-HT1A Receptor in the Neocortex of Mice. Cells. 2024; 13(6):466. https://doi.org/10.3390/cells13060466
Chicago/Turabian StyleJavadzadeh, Yasaman, Alexandra Santos, Mark S. Aquilino, Shanthini Mylvaganam, Karolina Urban, and Peter L. Carlen. 2024. "Cannabidiol Exerts Anticonvulsant Effects Alone and in Combination with Δ9-THC through the 5-HT1A Receptor in the Neocortex of Mice" Cells 13, no. 6: 466. https://doi.org/10.3390/cells13060466
APA StyleJavadzadeh, Y., Santos, A., Aquilino, M. S., Mylvaganam, S., Urban, K., & Carlen, P. L. (2024). Cannabidiol Exerts Anticonvulsant Effects Alone and in Combination with Δ9-THC through the 5-HT1A Receptor in the Neocortex of Mice. Cells, 13(6), 466. https://doi.org/10.3390/cells13060466