
Exploring Extracellular Matrix Crosslinking as a Therapeutic Approach to Fibrosis


Abstract
1. Introduction
2. ECM Crosslinking Biochemical Pathways
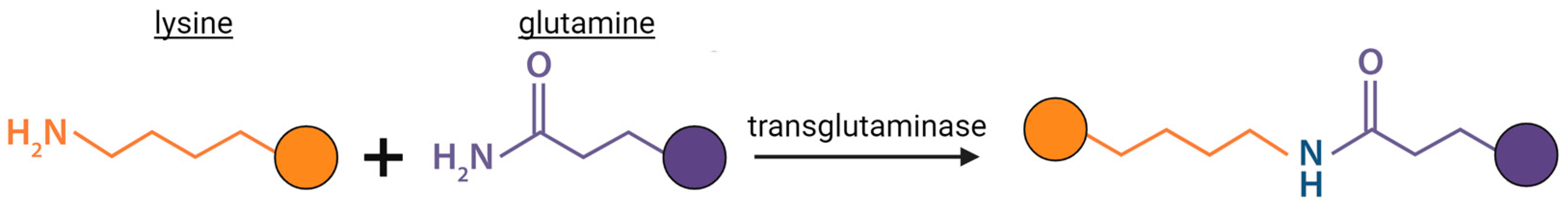
2.1. Transglutaminase (TG) Crosslinking
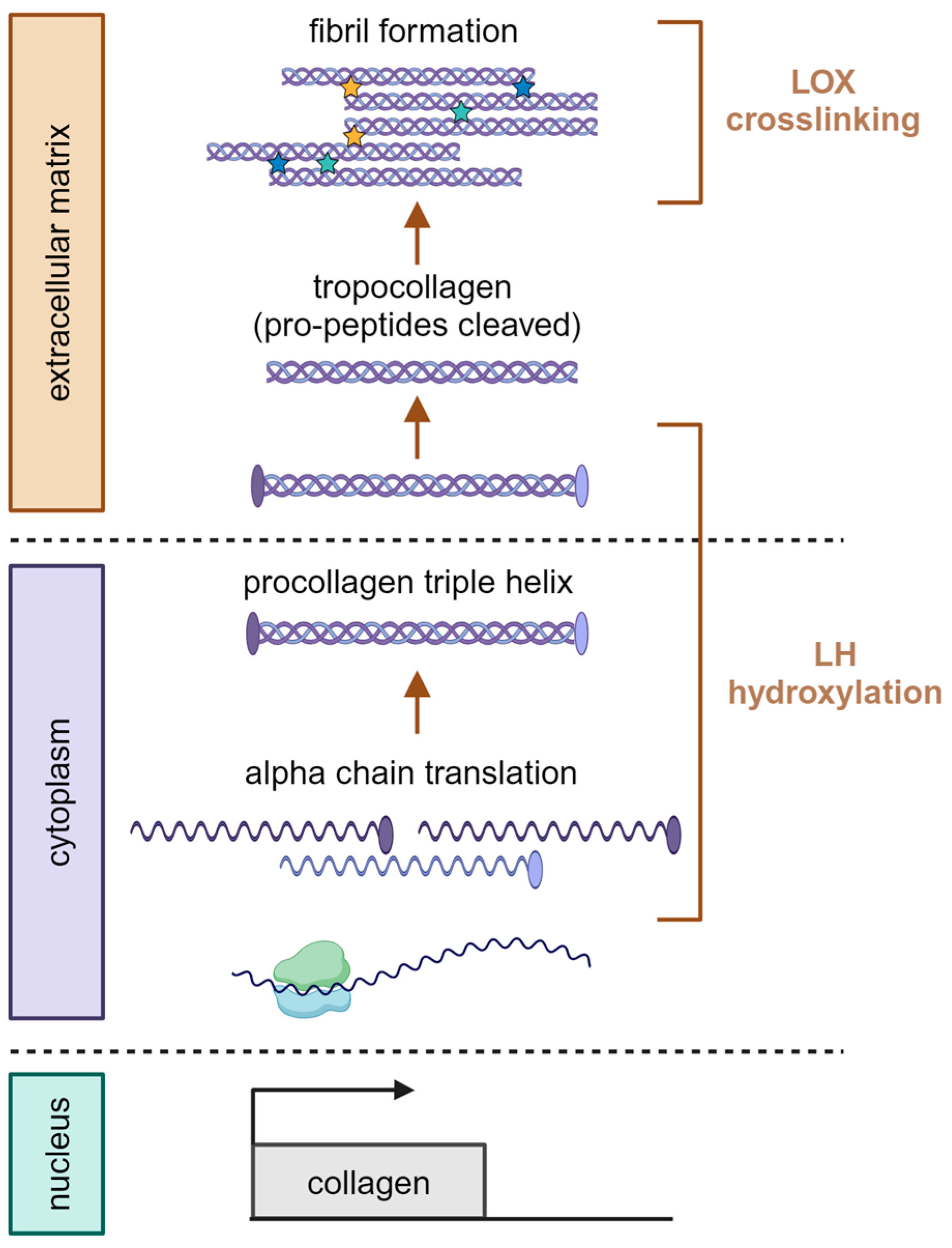
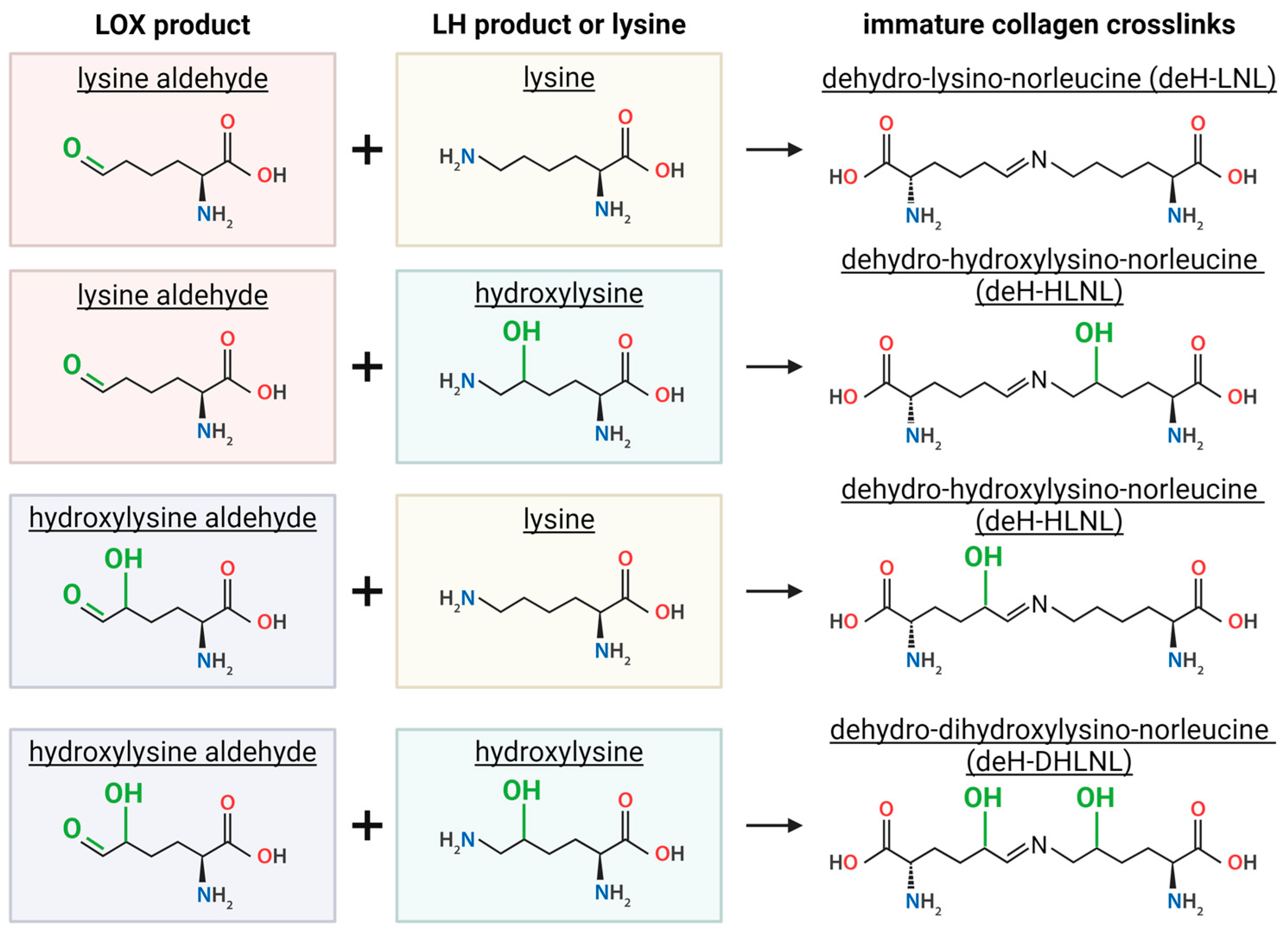
2.2. Lysine Hydroxylase (LH)
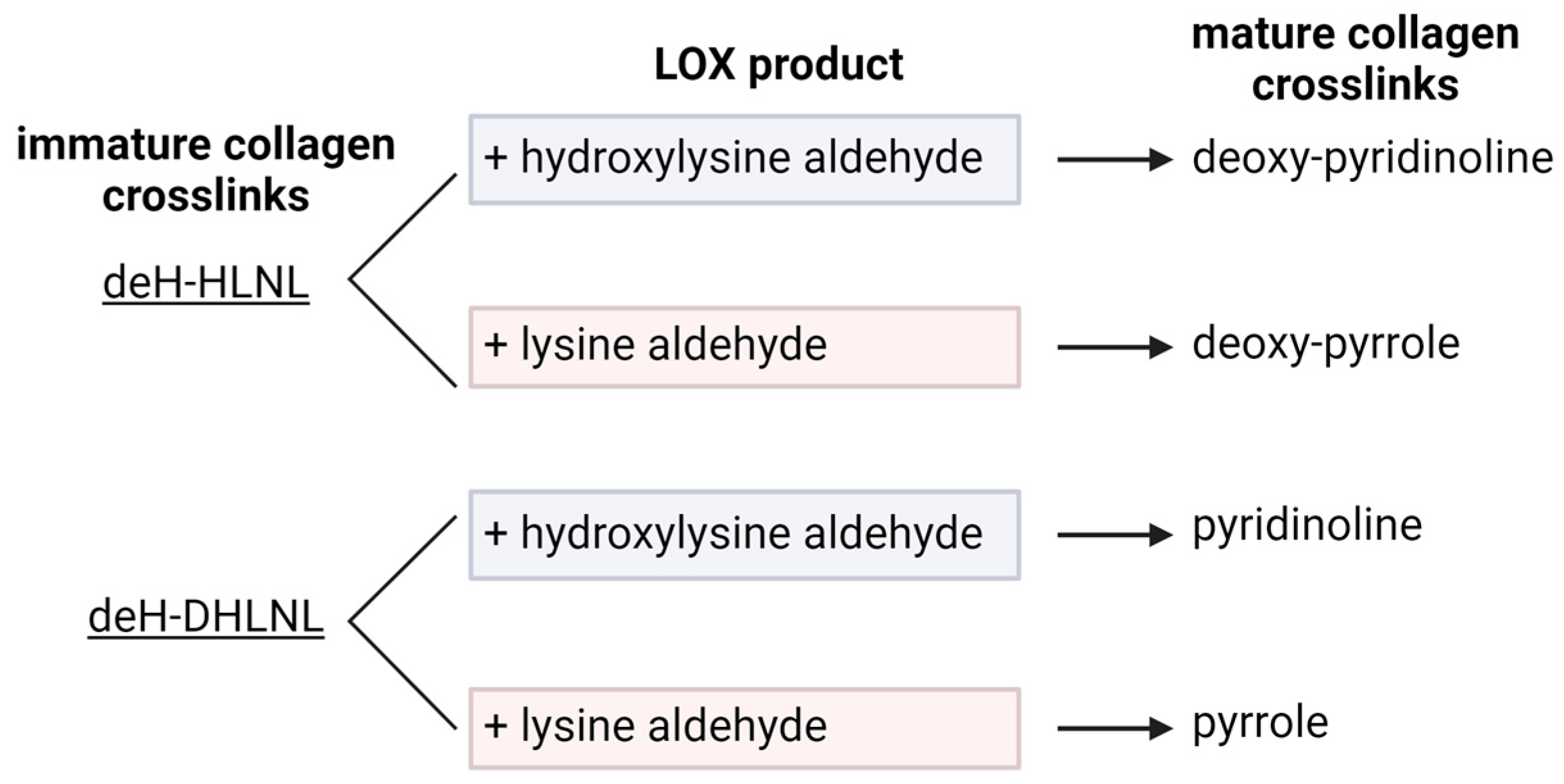
2.3. Lysyl Oxidase (LOX) Crosslinking
2.4. LOX-Mediated Collagen Crosslinking
2.5. LOX-Mediated Elastin Crosslinking
2.6. LOX in Fibrosis
2.7. Summary of Crosslinking Biochemistry
3. ECM Crosslinking Analysis Methods
4. Dysregulation of ECM Crosslinking in Fibrotic Disease
4.1. Lung Fibrosis
4.2. Liver Fibrosis
4.3. Skin Fibrosis
4.4. Insights into Crosslinking Dysregulation in Fibrotic Disease
5. ECM Crosslink-Based Therapeutic Strategies, Drug Targets, and Molecules
6. Discussion
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lurje, I.; Gaisa, N.T.; Weiskirchen, R.; Tacke, F. Mechanisms of Organ Fibrosis: Emerging Concepts and Implications for Novel Treatment Strategies. Mol. Asp. Med. 2023, 92, 101191. [Google Scholar] [CrossRef]
- Henderson, N.C.; Rieder, F.; Wynn, T.A. Fibrosis: From Mechanisms to Medicines. Nature 2020, 587, 555–566. [Google Scholar] [CrossRef]
- Distler, J.H.W.; Györfi, A.-H.; Ramanujam, M.; Whitfield, M.L.; Königshoff, M.; Lafyatis, R. Shared and Distinct Mechanisms of Fibrosis. Nat. Rev. Rheumatol. 2019, 15, 705–730. [Google Scholar] [CrossRef]
- Weiskirchen, R.; Weiskirchen, S.; Tacke, F. Organ and Tissue Fibrosis: Molecular Signals, Cellular Mechanisms and Translational Implications. Mol. Asp. Med. 2019, 65, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Pakshir, P.; Hinz, B. The Big Five in Fibrosis: Macrophages, Myofibroblasts, Matrix, Mechanics, and Miscommunication. Matrix Biol. 2018, 68, 81–93. [Google Scholar] [CrossRef]
- Wynn, T.A. Fibrotic Disease and the TH1/TH2 Paradigm. Nat. Rev. Immunol. 2004, 4, 583–594. [Google Scholar] [CrossRef]
- Talbott, H.E.; Mascharak, S.; Griffin, M.; Wan, D.C.; Longaker, M.T. Wound Healing, Fibroblast Heterogeneity, and Fibrosis. Cell Stem Cell 2022, 29, 1161–1180. [Google Scholar] [CrossRef]
- Karsdal, M.A.; Nielsen, S.H.; Leeming, D.J.; Langholm, L.L.; Nielsen, M.J.; Manon-Jensen, T.; Siebuhr, A.; Gudmann, N.S.; Rønnow, S.; Sand, J.M.; et al. The Good and the Bad Collagens of Fibrosis—Their Role in Signaling and Organ Function. Adv. Drug Deliv. Rev. 2017, 121, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Herrera, J.; Henke, C.A.; Bitterman, P.B. Extracellular Matrix as a Driver of Progressive Fibrosis. J. Clin. Investig. 2018, 128, 45–53. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, J.; Sun, H.; Zhang, Y.; Zou, D. New Insights into Fibrosis from the ECM Degradation Perspective: The Macrophage-MMP-ECM Interaction. Cell Biosci. 2022, 12, 117. [Google Scholar] [CrossRef]
- Hynes, R.O.; Naba, A. Overview of the Matrisome—An Inventory of Extracellular Matrix Constituents and Functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the Extracellular Matrix in Development and Disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Piersma, B.; Bank, R.A. Collagen Cross-Linking Mediated by Lysyl Hydroxylase 2: An Enzymatic Battlefield to Combat Fibrosis. Essays Biochem. 2019, 63, 377–387. [Google Scholar] [CrossRef]
- Kong, W.; Lyu, C.; Liao, H.; Du, Y. Collagen Crosslinking: Effect on Structure, Mechanics and Fibrosis Progression. Biomed. Mater. 2021, 16, 062005. [Google Scholar] [CrossRef]
- Vallet, S.D.; Ricard-Blum, S. Lysyl Oxidases: From Enzyme Activity to Extracellular Matrix Cross-Links. Essays Biochem. 2019, 63, 349–364. [Google Scholar] [CrossRef]
- Beninati, S.; Piacentini, M. The Transglutaminase Family: An Overview: Minireview Article. Amino Acids 2004, 26, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Schmelzer, C.E.H.; Hedtke, T.; Heinz, A. Unique Molecular Networks: Formation and Role of Elastin Cross-links. IUBMB Life 2020, 72, 842–854. [Google Scholar] [CrossRef]
- Muiznieks, L.D.; Keeley, F.W. Molecular Assembly and Mechanical Properties of the Extracellular Matrix: A Fibrous Protein Perspective. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 866–875. [Google Scholar] [CrossRef] [PubMed]
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef]
- Huizen, N.A.; Ijzermans, J.N.M.; Burgers, P.C.; Luider, T.M. Collagen Analysis with Mass Spectrometry. Mass Spectrom. Rev. 2020, 39, 309–335. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Sricholpech, M. Lysine Post-Translational Modifications of Collagen. Essays Biochem. 2012, 52, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Gjaltema, R.A.F.; Bank, R.A. Molecular Insights into Prolyl and Lysyl Hydroxylation of Fibrillar Collagens in Health and Disease. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 74–95. [Google Scholar] [CrossRef] [PubMed]
- Pehrsson, M.; Mortensen, J.H.; Manon-Jensen, T.; Bay-Jensen, A.-C.; Karsdal, M.A.; Davies, M.J. Enzymatic Cross-Linking of Collagens in Organ Fibrosis—Resolution and Assessment. Expert Rev. Mol. Diagn. 2021, 21, 1049–1064. [Google Scholar] [CrossRef] [PubMed]
- Salo, A.M.; Myllyharju, J. Prolyl and Lysyl Hydroxylases in Collagen Synthesis. Exp. Dermatol. 2021, 30, 38–49. [Google Scholar] [CrossRef]
- Lamande, S.R.; Bateman, J.F. Procollagen Folding and Assembly: The Role of Endoplasmic Reticulum Enzymes and Molecular Chaperones. Semin. Cell Dev. Biol. 1999, 10, 455–464. [Google Scholar] [CrossRef]
- Eckert, R.L.; Kaartinen, M.T.; Nurminskaya, M.; Belkin, A.M.; Colak, G.; Johnson, G.V.W.; Mehta, K. Transglutaminase Regulation of Cell Function. Physiol. Rev. 2014, 94, 383–417. [Google Scholar] [CrossRef]
- Tatsukawa, H.; Takeuchi, T.; Shinoda, Y.; Hitomi, K. Identification and Characterization of Substrates Crosslinked by Transglutaminases in Liver and Kidney Fibrosis. Anal. Biochem. 2020, 604, 113629. [Google Scholar] [CrossRef]
- Benn, M.C.; Weber, W.; Klotzsch, E.; Vogel, V.; Pot, S.A. Tissue Transglutaminase in Fibrosis—More than an Extracellular Matrix Cross-Linker. Curr. Opin. Biomed. Eng. 2019, 10, 156–164. [Google Scholar] [CrossRef]
- Lee, Z.-W.; Kwon, S.-M.; Kim, S.-W.; Yi, S.-J.; Kim, Y.-M.; Ha, K.-S. Activation of in Situ Tissue Transglutaminase by Intracellular Reactive Oxygen Species. Biochem. Biophys. Res. Commun. 2003, 305, 633–640. [Google Scholar] [CrossRef]
- Siegel, M.; Strnad, P.; Watts, R.E.; Choi, K.; Jabri, B.; Omary, M.B.; Khosla, C. Extracellular Transglutaminase 2 Is Catalytically Inactive, but Is Transiently Activated upon Tissue Injury. PLoS ONE 2008, 3, e1861. [Google Scholar] [CrossRef]
- Soltani, F.; Kaartinen, M.T. Transglutaminases in Fibrosis—Overview and Recent Advances. Am. J. Physiol.-Cell Physiol. 2023, 325, C885–C894. [Google Scholar] [CrossRef]
- Troilo, H.; Steer, R.; Collins, R.F.; Kielty, C.M.; Baldock, C. Independent Multimerization of Latent TGFβ Binding Protein-1 Stabilized by Cross-Linking and Enhanced by Heparan Sulfate. Sci. Rep. 2016, 6, 34347. [Google Scholar] [CrossRef]
- van der Slot, A.J.; Zuurmond, A.-M.; van den Bogaerdt, A.J.; Ulrich, M.M.W.; Middelkoop, E.; Boers, W.; Ronday, H.K.; DeGroot, J.; Huizinga, T.W.J.; Bank, R.A. Increased Formation of Pyridinoline Cross-Links Due to Higher Telopeptide Lysyl Hydroxylase Levels Is a General Fibrotic Phenomenon. Matrix Biol. 2004, 23, 251–257. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, H.; Terajima, M.; Banerjee, P.; Liu, X.; Yu, J.; Momin, A.A.; Katayama, H.; Hanash, S.M.; Burns, A.R.; et al. Lysyl Hydroxylase 2 Is Secreted by Tumor Cells and Can Modify Collagen in the Extracellular Space. J. Biol. Chem. 2016, 291, 25799–25808. [Google Scholar] [CrossRef]
- Jessen, H.; Hoyer, N.; Prior, T.S.; Frederiksen, P.; Karsdal, M.A.; Leeming, D.J.; Bendstrup, E.; Sand, J.M.B.; Shaker, S.B. Turnover of Type I and III Collagen Predicts Progression of Idiopathic Pulmonary Fibrosis. Respir. Res. 2021, 22, 205. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, M.; Goheen, M.; Jia, S.; Ling, S.; White, E.S.; Kim, K.K. Type I Collagen Signaling Regulates Opposing Fibrotic Pathways through α 2 β 1 Integrin. Am. J. Respir. Cell Mol. Biol. 2020, 63, 613–622. [Google Scholar] [CrossRef] [PubMed]
- van der Slot, A.J.; Zuurmond, A.-M.; Bardoel, A.F.J.; Wijmenga, C.; Pruijs, H.E.H.; Sillence, D.O.; Brinckmann, J.; Abraham, D.J.; Black, C.M.; Verzijl, N.; et al. Identification of PLOD2 as Telopeptide Lysyl Hydroxylase, an Important Enzyme in Fibrosis*. J. Biol. Chem. 2003, 278, 40967–40972. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Zhang, X.; Duan, L.; Fang, H.; Rao, S.; Liu, W.; Guo, B.; Zhang, X. Lysyl Hydroxylase Inhibition by Minoxidil Blocks Collagen Deposition and Prevents Pulmonary Fibrosis via TGF-Β1/Smad3 Signaling Pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 8592–8601. [Google Scholar] [CrossRef] [PubMed]
- Brereton, C.J.; Yao, L.; Davies, E.R.; Zhou, Y.; Vukmirovic, M.; Bell, J.A.; Wang, S.; Ridley, R.A.; Dean, L.S.; Andriotis, O.G.; et al. Pseudohypoxic HIF Pathway Activation Dysregulates Collagen Structure-Function in Human Lung Fibrosis. eLife 2022, 11, e69348. [Google Scholar] [CrossRef] [PubMed]
- Zaffryar-Eilot, S.; Hasson, P. Lysyl Oxidases: Orchestrators of Cellular Behavior and ECM Remodeling and Homeostasis. Int. J. Mol. Sci. 2022, 23, 11378. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Barker, T.H.; Gibbons, D.L.; Kurie, J.M. The Fibrotic Tumor Stroma. J. Clin. Investig. 2018, 128, 16–25. [Google Scholar] [CrossRef]
- Knott, L.; Bailey, A.J. Collagen Cross-Links in Mineralizing Tissues: A Review of Their Chemistry, Function, and Clinical Relevance. Bone 1998, 22, 181–187. [Google Scholar] [CrossRef]
- Svensson, R.B.; Mulder, H.; Kovanen, V.; Magnusson, S.P. Fracture Mechanics of Collagen Fibrils: Influence of Natural Cross-Links. Biophys. J. 2013, 104, 2476–2484. [Google Scholar] [CrossRef]
- Shetty, S.S.; Sharma, M.; Kabekkodu, S.P.; Kumar, N.A.; Satyamoorthy, K.; Radhakrishnan, R. Understanding the Molecular Mechanism Associated with Reversal of Oral Submucous Fibrosis Targeting Hydroxylysine Aldehyde-Derived Collagen Cross-Links. J. Carcinog. 2021, 20, 9. [Google Scholar] [CrossRef]
- Yamauchi, M.; Taga, Y.; Terajima, M. Analyses of Lysine Aldehyde Cross-Linking in Collagen Reveal That the Mature Cross-Link Histidinohydroxylysinonorleucine Is an Artifact. J. Biol. Chem. 2019, 294, 14163. [Google Scholar] [CrossRef]
- Eyre, D.R.; Weis, M.; Rai, J. Analyses of Lysine Aldehyde Cross-Linking in Collagen Reveal That the Mature Cross-Link Histidinohydroxylysinonorleucine Is an Artifact. J. Biol. Chem. 2019, 294, 6578–6590. [Google Scholar] [CrossRef] [PubMed]
- Puente, A.; Fortea, J.I.; Cabezas, J.; Loste, M.T.A.; Iruzubieta, P.; Llerena, S.; Huelin, P.; Fábrega, E.; Crespo, J. LOXL2—A New Target in Antifibrogenic Therapy? Int. J. Mol. Sci. 2019, 20, 1634. [Google Scholar] [CrossRef] [PubMed]
- Rosenbloom, J.; Ren, S.; Macarak, E. New Frontiers in Fibrotic Disease Therapies: The Focus of the Joan and Joel Rosenbloom Center for Fibrotic Diseases at Thomas Jefferson University. Matrix Biol. 2016, 51, 14–25. [Google Scholar] [CrossRef]
- Ma, H.-Y.; Li, Q.; Wong, W.R.; N’Diaye, E.-N.; Caplazi, P.; Bender, H.; Huang, Z.; Arlantico, A.; Jeet, S.; Wong, A.; et al. LOXL4, but Not LOXL2, Is the Critical Determinant of Pathological Collagen Cross-Linking and Fibrosis in the Lung. Sci. Adv. 2023, 9, eadf0133. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.G.; Andriotis, O.G.; Roberts, J.J.; Lunn, K.; Tear, V.J.; Cao, L.; Ask, K.; Smart, D.E.; Bonfanti, A.; Johnson, P.; et al. Nanoscale Dysregulation of Collagen Structure-Function Disrupts Mechano-Homeostasis and Mediates Pulmonary Fibrosis. eLife 2018, 7, e36354. [Google Scholar] [CrossRef] [PubMed]
- Istok, R.; Bély, M.; Stancíková, M.; Rovenský, J. Evidence for Increased Pyridinoline Concentration in Fibrotic Tissues in Diffuse Systemic Sclerosis. Exp. Dermatol. 2001, 26, 545–547. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Lemus, E.; Hägglund, P.; López-Alarcón, C.; Davies, M.J. Oxidative Crosslinking of Peptides and Proteins: Mechanisms of Formation, Detection, Characterization and Quantification. Molecules 2021, 27, 15. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Davies, M.J. Detection, Identification, and Quantification of Oxidative Protein Modifications. J. Biol. Chem. 2019, 294, 19683–19708. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, T.; Seyama, Y.; Yamashita, S. Comparison of ELISA and HPLC for the Determination of Desmosine or Isodesmosine in Aortic Tissue Elastin. J. Clin. Lab. Anal. 1995, 9, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Stoilov, I.; Starcher, B.C.; Mecham, R.P.; Broekelmann, T.J. Chapter 7 Measurement of Elastin, Collagen, and Total Protein Levels in Tissues. Methods Cell Biol. 2018, 143, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Taga, Y.; Hattori, S.; Shiiba, M.; Terajima, M. Chapter 6 Analysis of Collagen and Elastin Cross-Links. Methods Cell Biol. 2018, 143, 115–132. [Google Scholar] [CrossRef]
- Gineyts, E.; Borel, O.; Chapurlat, R.; Garnero, P. Quantification of Immature and Mature Collagen Crosslinks by Liquid Chromatography–Electrospray Ionization Mass Spectrometry in Connective Tissues. J. Chromatogr. B 2010, 878, 1449–1454. [Google Scholar] [CrossRef]
- Zork, N.M.; Myers, K.M.; Yoshida, K.; Cremers, S.; Jiang, H.; Ananth, C.V.; Wapner, R.J.; Kitajewski, J.; Vink, J. A Systematic Evaluation of Collagen Cross-Links in the Human Cervix. Am. J. Obstet. Gynecol. 2015, 212, 321.e1–321.e8. [Google Scholar] [CrossRef]
- Glass, D.S.; Grossfeld, D.; Renna, H.A.; Agarwala, P.; Spiegler, P.; DeLeon, J.; Reiss, A.B. Idiopathic Pulmonary Fibrosis: Current and Future Treatment. Clin. Respir. J. 2022, 16, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Tjin, G.; White, E.S.; Faiz, A.; Sicard, D.; Tschumperlin, D.J.; Mahar, A.; Kable, E.P.W.; Burgess, J.K. Lysyl Oxidases Regulate Fibrillar Collagen Remodelling in Idiopathic Pulmonary Fibrosis. Dis. Model. Mech. 2017, 10, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Afratis, N.A.; Klepfish, M.; Karamanos, N.K.; Sagi, I. The Apparent Competitive Action of ECM Proteases and Cross-Linking Enzymes during Fibrosis: Applications to Drug Discovery. Adv. Drug Deliv. Rev. 2018, 129, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Stalnikowitz, D.K.; Weissbrod, A.B. Liver Fibrosis and Inflammation. A Review. Ann. Hepatol. 2003, 2, 159–163. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, A.; Chen, W.; Wang, P.; Liu, T.; Cong, M.; Xu, A.; Yan, X.; Jia, J.; You, H. Inhibition of Lysyl Oxidase-like 1 (LOXL1) Expression Arrests Liver Fibrosis Progression in Cirrhosis by Reducing Elastin Crosslinking. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2018, 1864, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Hayasaka, A.; Ilda, S.; Suzuki, N.; Kondo, F.; Miyazaki, M.; Yonemitsu, H. Pyridinoline Collagen Cross-Links in Patients with Chronic Viral Hepatitis and Cirrhosis. J. Hepatol. 1996, 24, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Lyu, C.; Kong, W.; Liu, Z.; Wang, S.; Zhao, P.; Liang, K.; Niu, Y.; Yang, W.; Xiang, C.; Hu, X.; et al. Advanced Glycation End-Products as Mediators of the Aberrant Crosslinking of Extracellular Matrix in Scarred Liver Tissue. Nat. Biomed. Eng. 2023, 7, 1437–1454. [Google Scholar] [CrossRef]
- van der Slot-Verhoeven, A.J.; van Dura, E.A.; Attema, J.; Blauw, B.; DeGroot, J.; Huizinga, T.W.J.; Zuurmond, A.-M.; Bank, R.A. The Type of Collagen Cross-Link Determines the Reversibility of Experimental Skin Fibrosis. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2005, 1740, 60–67. [Google Scholar] [CrossRef]
- Huang, M.; Cai, G.; Baugh, L.M.; Liu, Z.; Smith, A.; Watson, M.; Popovich, D.; Zhang, T.; Stawski, L.S.; Trojanowska, M.; et al. Systemic Sclerosis Dermal Fibroblasts Induce Cutaneous Fibrosis Through Lysyl Oxidase–like 4: New Evidence from Three-Dimensional Skin-like Tissues. Arthritis Rheumatol. 2020, 72, 791–801. [Google Scholar] [CrossRef]
- Brinckmann, J.; Açil, Y.; Tronnier, M.; Notbohm, H.; Bätge, B.; Schmeller, W.; Koch, M.H.J.; Müller, P.K.; Wolff, H.H. Altered X-Ray Diffraction Pattern Is Accompanied by a Change in the Mode of Cross-Link Formation in Lipodermatosclerosis. J. Investig. Dermatol. 1996, 107, 589–592. [Google Scholar] [CrossRef]
- Harrison, S.A.; Abdelmalek, M.F.; Caldwell, S.; Shiffman, M.L.; Diehl, A.M.; Ghalib, R.; Lawitz, E.J.; Rockey, D.C.; Schall, R.A.; Jia, C.; et al. Simtuzumab Is Ineffective for Patients with Bridging Fibrosis or Compensated Cirrhosis Caused by Nonalcoholic Steatohepatitis. Gastroenterology 2018, 155, 1140–1153. [Google Scholar] [CrossRef]
- Muir, A.J.; Levy, C.; Janssen, H.L.A.; Montano-Loza, A.J.; Shiffman, M.L.; Caldwell, S.; Luketic, V.; Ding, D.; Jia, C.; McColgan, B.J.; et al. Simtuzumab for Primary Sclerosing Cholangitis: Phase 2 Study Results with Insights on the Natural History of the Disease. Hepatology 2019, 69, 684–698. [Google Scholar] [CrossRef]
- Verstovsek, S.; Savona, M.R.; Mesa, R.A.; Dong, H.; Maltzman, J.D.; Sharma, S.; Silverman, J.; Oh, S.T.; Gotlib, J. A Phase 2 Study of Simtuzumab in Patients with Primary, Post-polycythaemia Vera or Post-essential Thrombocythaemia Myelofibrosis. Br. J. Haematol. 2017, 176, 939–949. [Google Scholar] [CrossRef]
- Raghu, G.; Brown, K.K.; Collard, H.R.; Cottin, V.; Gibson, K.F.; Kaner, R.J.; Lederer, D.J.; Martinez, F.J.; Noble, P.W.; Song, J.W.; et al. Efficacy of Simtuzumab versus Placebo in Patients with Idiopathic Pulmonary Fibrosis: A Randomised, Double-Blind, Controlled, Phase 2 Trial. Lancet Respir. Med. 2016, 5, 22–32. [Google Scholar] [CrossRef]
- Chitty, J.L.; Yam, M.; Perryman, L.; Parker, A.L.; Skhinas, J.N.; Setargew, Y.F.I.; Mok, E.T.Y.; Tran, E.; Grant, R.D.; Latham, S.L.; et al. A First-in-Class Pan-Lysyl Oxidase Inhibitor Impairs Stromal Remodeling and Enhances Gemcitabine Response and Survival in Pancreatic Cancer. Nat. Cancer 2023, 4, 1326–1344. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Findlay, A.; Stolp, J.; Rayner, B.; Ask, K.; Jarolimek, W. Pan-Lysyl Oxidase Inhibitor PXS-5505 Ameliorates Multiple-Organ Fibrosis by Inhibiting Collagen Crosslinks in Rodent Models of Systemic Sclerosis. Int. J. Mol. Sci. 2022, 23, 5533. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Saad, S.; Shi, Y.; Wang, R.; Chou, A.S.Y.; Gill, A.; Yao, Y.; Jarolimek, W.; Pollock, C.A. Lysyl Oxidase Inhibitors Attenuate Cyclosporin A-Induced Nephropathy in Mouse. Sci. Rep. 2021, 11, 12437. [Google Scholar] [CrossRef] [PubMed]
- Rowbottom, M.W.; Bain, G.; Calderon, I.; Lasof, T.; Lonergan, D.; Lai, A.; Huang, F.; Darlington, J.; Prodanovich, P.; Santini, A.M.; et al. Identification of 4-(Aminomethyl)-6-(Trifluoromethyl)-2-(Phenoxy)Pyridine Derivatives as Potent, Selective, and Orally Efficacious Inhibitors of the Copper-Dependent Amine Oxidase, Lysyl Oxidase-Like 2 (LOXL2). J. Med. Chem. 2017, 60, 4403–4423. [Google Scholar] [CrossRef] [PubMed]
- Harrison, C.; Mascarenhas, J.; Cilloni, D.; Schlenk, R.; Jacoby, B.; Slack, R.J.; Aslanis, V.; Singh, B.; Lindmark, B.; Verstovsek, S.; et al. P1024: MYLOX-1: A PHASE II STUDY EVALUATING THE SAFETY, TOLERABILITY, PHARMACOKINETICS AND PHARMACODYNAMICS OF ORAL LOXL2 INHIBITOR GB2064 (WITH FOCUS ON BONE MARROW COLLAGEN) IN PATIENTS WITH MYELOFIBROSIS. HemaSphere 2023, 7, e87048d4. [Google Scholar] [CrossRef]
- Findlay, A.; Turner, C.; Schilter, H.; Deodhar, M.; Zhou, W.; Perryman, L.; Foot, J.; Zahoor, A.; Yao, Y.; Hamilton, R.; et al. An Activity-based Bioprobe Differentiates a Novel Small Molecule Inhibitor from a LOXL2 Antibody and Provides Renewed Promise for Anti-fibrotic Therapeutic Strategies. Clin. Transl. Med. 2021, 11, e572. [Google Scholar] [CrossRef]
- Chaudhari, N.; Findlay, A.D.; Stevenson, A.W.; Clemons, T.D.; Yao, Y.; Joshi, A.; Sayyar, S.; Wallace, G.; Rea, S.; Toshniwal, P.; et al. Topical Application of an Irreversible Small Molecule Inhibitor of Lysyl Oxidases Ameliorates Skin Scarring and Fibrosis. Nat. Commun. 2022, 13, 5555. [Google Scholar] [CrossRef]
- Schilter, H.; Findlay, A.D.; Perryman, L.; Yow, T.T.; Moses, J.; Zahoor, A.; Turner, C.I.; Deodhar, M.; Foot, J.S.; Zhou, W.; et al. The Lysyl Oxidase like 2/3 Enzymatic Inhibitor, PXS-5153A, Reduces Crosslinks and Ameliorates Fibrosis. J. Cell. Mol. Med. 2019, 23, 1759–1770. [Google Scholar] [CrossRef]
- Leiva, O.; Ng, S.K.; Matsuura, S.; Chitalia, V.; Lucero, H.; Findlay, A.; Turner, C.; Jarolimek, W.; Ravid, K. Novel Lysyl Oxidase Inhibitors Attenuate Hallmarks of Primary Myelofibrosis in Mice. Int. J. Hematol. 2019, 110, 699–708. [Google Scholar] [CrossRef]
- Büchold, C.; Hils, M.; Gerlach, U.; Weber, J.; Pelzer, C.; Heil, A.; Aeschlimann, D.; Pasternack, R. Features of ZED1227: The First-In-Class Tissue Transglutaminase Inhibitor Undergoing Clinical Evaluation for the Treatment of Celiac Disease. Cells 2022, 11, 1667. [Google Scholar] [CrossRef]
- Schuppan, D.; Mäki, M.; Lundin, K.E.A.; Isola, J.; Friesing-Sosnik, T.; Taavela, J.; Popp, A.; Koskenpato, J.; Langhorst, J.; Hovde, Ø.; et al. A Randomized Trial of Transglutaminase 2 Inhibitor for Celiac Disease. N. Engl. J. Med. 2021, 385, 35–45. [Google Scholar] [CrossRef]
- Maamra, M.; Benayad, O.M.; Matthews, D.; Kettleborough, C.; Atkinson, J.; Cain, K.; Bon, H.; Brand, H.; Parkinson, M.; Watson, P.F.; et al. Transglutaminase 2: Development of Therapeutic Antibodies Reveals Four Inhibitory Epitopes and Confirms Extracellular Function in Fibrotic Remodelling. Br. J. Pharmacol. 2022, 179, 2697–2712. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Trinh-Minh, T.; Matei, A.; Györfi, A.; Hong, X.; Bergmann, C.; Schett, G.; Atkinson, J.; Bowcutt, R.; Patel, J.; et al. Amelioration of Fibrotic Remodeling of Human 3-Dimensional Full-Thickness Skin by Transglutamase 2 Inhibition. Arthritis Rheumatol. 2023, 75, 1619–1627. [Google Scholar] [CrossRef]
- Fell, S.; Wang, Z.; Blanchard, A.; Nanthakumar, C.; Griffin, M. Transglutaminase 2: A Novel Therapeutic Target for Idiopathic Pulmonary Fibrosis Using Selective Small Molecule Inhibitors. Amino Acids 2021, 53, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Stuckey, D.J.; Murdoch, C.E.; Camelliti, P.; Lip, G.Y.H.; Griffin, M. Cardiac Fibrosis Can Be Attenuated by Blocking the Activity of Transglutaminase 2 Using a Selective Small-Molecule Inhibitor. Cell Death Dis. 2018, 9, 613. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zu, C.; Zhang, Y.; Wang, X.; Huan, X.; Wang, L. Blocking TG2 Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice through Inhibiting EMT. Respir. Physiol. Neurobiol. 2020, 276, 103402. [Google Scholar] [CrossRef] [PubMed]
- Badarau, E.; Wang, Z.; Rathbone, D.L.; Costanzi, A.; Thibault, T.; Murdoch, C.E.; Alaoui, S.E.; Bartkeviciute, M.; Griffin, M. Development of Potent and Selective Tissue Transglutaminase Inhibitors: Their Effect on TG2 Function and Application in Pathological Conditions. Chem. Biol. 2015, 22, 1347–1361. [Google Scholar] [CrossRef]
- Lee, J.; Guo, H.; Wang, S.; Maghsoud, Y.; Vázquez-Montelongo, E.A.; Jing, Z.; Sammons, R.M.; Cho, E.J.; Ren, P.; Cisneros, G.A.; et al. Unleashing the Potential of 1,3-Diketone Analogues as Selective LH2 Inhibitors. ACS Med. Chem. Lett. 2023, 14, 1396–1403. [Google Scholar] [CrossRef] [PubMed]
- Maghsoud, Y.; Vázquez-Montelongo, E.A.; Yang, X.; Liu, C.; Jing, Z.; Lee, J.; Harger, M.; Smith, A.K.; Espinoza, M.; Guo, H.-F.; et al. Computational Investigation of a Series of Small Molecules as Potential Compounds for Lysyl Hydroxylase-2 (LH2) Inhibition. J. Chem. Inf. Model. 2023, 63, 986–1001. [Google Scholar] [CrossRef]
- Devkota, A.K.; Veloria, J.R.; Guo, H.-F.; Kurie, J.M.; Cho, E.J.; Dalby, K.N. Development of a High-Throughput Lysyl Hydroxylase (LH) Assay and Identification of Small-Molecule Inhibitors against LH2. SLAS Discov. 2018, 24, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Meissner, E.G.; McLaughlin, M.; Matthews, L.; Gharib, A.M.; Wood, B.J.; Levy, E.; Sinkus, R.; Virtaneva, K.; Sturdevant, D.; Martens, C.; et al. Simtuzumab Treatment of Advanced Liver Fibrosis in HIV and HCV-infected Adults: Results of a 6-month Open-label Safety Trial. Liver Int. 2016, 36, 1783–1792. [Google Scholar] [CrossRef] [PubMed]
- Vachhani, P.; Baskar, J.; Charlton, B.; Cheung, S.; Jarolimek, W.; Lee, S.-E.; Tan, P.; Watson, A.M.; Wu, S.-J. PXS5505-MF-101: A Phase 1/2a Study to Evaluate Safety, Pharmacokinetics and Pharmacodynamics of Pxs-5505 in Patients with Primary, Post-Polycythemia Vera or Post-Essential Thrombocythemia Myelofibrosis. Blood 2023, 142, 625. [Google Scholar] [CrossRef]
- Zhao, P.; Sun, T.; Lyu, C.; Liang, K.; Du, Y. Cell Mediated ECM-Degradation as an Emerging Tool for Anti-Fibrotic Strategy. Cell Regen. 2023, 12, 29. [Google Scholar] [CrossRef]
- Rozario, T.; DeSimone, D.W. The Extracellular Matrix in Development and Morphogenesis: A Dynamic View. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Phogat, S.; Thiam, F.; Yazeedi, S.A.; Abokor, F.A.; Osei, E.T. 3D in Vitro Hydrogel Models to Study the Human Lung Extracellular Matrix and Fibroblast Function. Respir. Res. 2023, 24, 242. [Google Scholar] [CrossRef] [PubMed]
- Urciuolo, F.; Imparato, G.; Netti, P.A. In Vitro Strategies for Mimicking Dynamic Cell–ECM Reciprocity in 3D Culture Models. Front. Bioeng. Biotechnol. 2023, 11, 1197075. [Google Scholar] [CrossRef]
- Perez-Castillejos, R. Replication of the 3D Architecture of Tissues. Mater. Today 2010, 13, 32–41. [Google Scholar] [CrossRef]
- Qin, L.; Liu, N.; Bao, C.; Yang, D.; Ma, G.; Yi, W.; Xiao, G.; Cao, H. Mesenchymal Stem Cells in Fibrotic Diseases—The Two Sides of the Same Coin. Acta Pharmacol. Sin. 2023, 44, 268–287. [Google Scholar] [CrossRef]
- Brennan, P.N.; MacMillan, M.; Manship, T.; Moroni, F.; Glover, A.; Graham, C.; Semple, S.; Morris, D.M.; Fraser, A.R.; Pass, C.; et al. Study Protocol: A Multicentre, Open-Label, Parallel-Group, Phase 2, Randomised Controlled Trial of Autologous Macrophage Therapy for Liver Cirrhosis (MATCH). BMJ Open 2021, 11, e053190. [Google Scholar] [CrossRef] [PubMed]
- Moroni, F.; Dwyer, B.J.; Graham, C.; Pass, C.; Bailey, L.; Ritchie, L.; Mitchell, D.; Glover, A.; Laurie, A.; Doig, S.; et al. Safety Profile of Autologous Macrophage Therapy for Liver Cirrhosis. Nat. Med. 2019, 25, 1560–1565. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Target | Drug Molecule | Drug Type | Indication | Status | Reference |
|---|---|---|---|---|---|---|
| LOX | LOXL2 | Simtuzumab/GS-6624/AB0024 | antibody | Cancer, IPF and liver fibrosis, myelofibrosis, PSC, NASH | Phase II—Discontinued | [69,70,71,72] |
| Pan-LOX | PXS-5505/SNT-5505 | small molecule | Myelofibrosis, liver and pancreatic cancer | Phase II | [73,74,75] NCT05109052, NCT04676529 | |
| LOXL2 | PAT-1251 | small molecule | IPF and other fibrotic diseases, myelofibrosis | Phase II | [76], NCT04054245, NCT02852551 | |
| LOXL2 | GB2064 | small molecule | Myelofibrosis | Phase II | [77], NCT04679870 | |
| LOXL2 | PXS-5338 | small molecule | NASH, IPF, liver and kidney fibrosis | Phase I | [78] | |
| LOXL2 | PXS-5382/SNT-5382 | small molecule | Anti-fibrotic IPF/CKD/NASH | Phase I | [75] | |
| Pan-LOX | PXS-6302 | small molecule | Anti-scarring; burns, established scars | Phase I | [79] | |
| LOXL2/LOXL3 | PXS-5153A | small molecule | Liver fibrosis, myocardial infarction | Preclinical | [80] | |
| LOX | PXS-LOX_1 and PXS-LOX_2 | small molecule | Primary myelofibrosis (PMF) | Preclinical | [81] | |
| TG | TG2 | ZED1227/TAK-227 | small molecule | NAFLD with significant fibrosis | Phase II | [82,83], NCT05305599 |
| TG2 | Zampilimab | antibody | Adult kidney transplant recipients with chronic allograft injury | Phase I/II | NCT04705350, NCT04335578 | |
| TG2 | AB1, DC1, and BB7 | antibody | Fibrosis and auto-immune disease | Preclinical | [84,95] | |
| TG2 | 1–155 | small molecule | IPF, cardiac fibrosis | Preclinical | [86,87] | |
| TG2 | R281 | small molecule | IPF | Preclinical | [86] | |
| TG2 | GK921 | small molecule | Pulmonary fibrosis | Preclinical | [88] | |
| TG2 | Compound 3h | small molecule | Hypertensive nephrosclerosis | Preclinical | [89] | |
| LH | LH2 | 1,3-Diketone analogs | small molecule | Cancer metastasis | Preclinical | [90,91,92] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lloyd, S.M.; He, Y. Exploring Extracellular Matrix Crosslinking as a Therapeutic Approach to Fibrosis. Cells 2024, 13, 438. https://doi.org/10.3390/cells13050438
Lloyd SM, He Y. Exploring Extracellular Matrix Crosslinking as a Therapeutic Approach to Fibrosis. Cells. 2024; 13(5):438. https://doi.org/10.3390/cells13050438
Chicago/Turabian StyleLloyd, Sarah M., and Yupeng He. 2024. "Exploring Extracellular Matrix Crosslinking as a Therapeutic Approach to Fibrosis" Cells 13, no. 5: 438. https://doi.org/10.3390/cells13050438
APA StyleLloyd, S. M., & He, Y. (2024). Exploring Extracellular Matrix Crosslinking as a Therapeutic Approach to Fibrosis. Cells, 13(5), 438. https://doi.org/10.3390/cells13050438