



Hepatic Nampt Deficiency Aggravates Dyslipidemia and Fatty Liver in High Fat Diet Fed Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of Hepatocyte-Specific Nampt Knockout Mice
2.2. Animal Treatment and Sample Collection
2.3. Genomic DNA Extraction and Genotyping by Polymerase Chain Reaction
2.4. Serum Lipids Assys
2.5. RNA Isolation and Real-Time PCR Analysis
2.6. Atherosclerotic Lesion Analysis
2.7. Histopathological Analysis and Oil Red O Staining
2.8. Measurement of Fasting Blood Glucose
2.9. Serum Biochemical Analysis
2.10. Statistical Analysis
3. Results
3.1. Identification of the Hepatocyte-Specific Nampt Knockout Mice
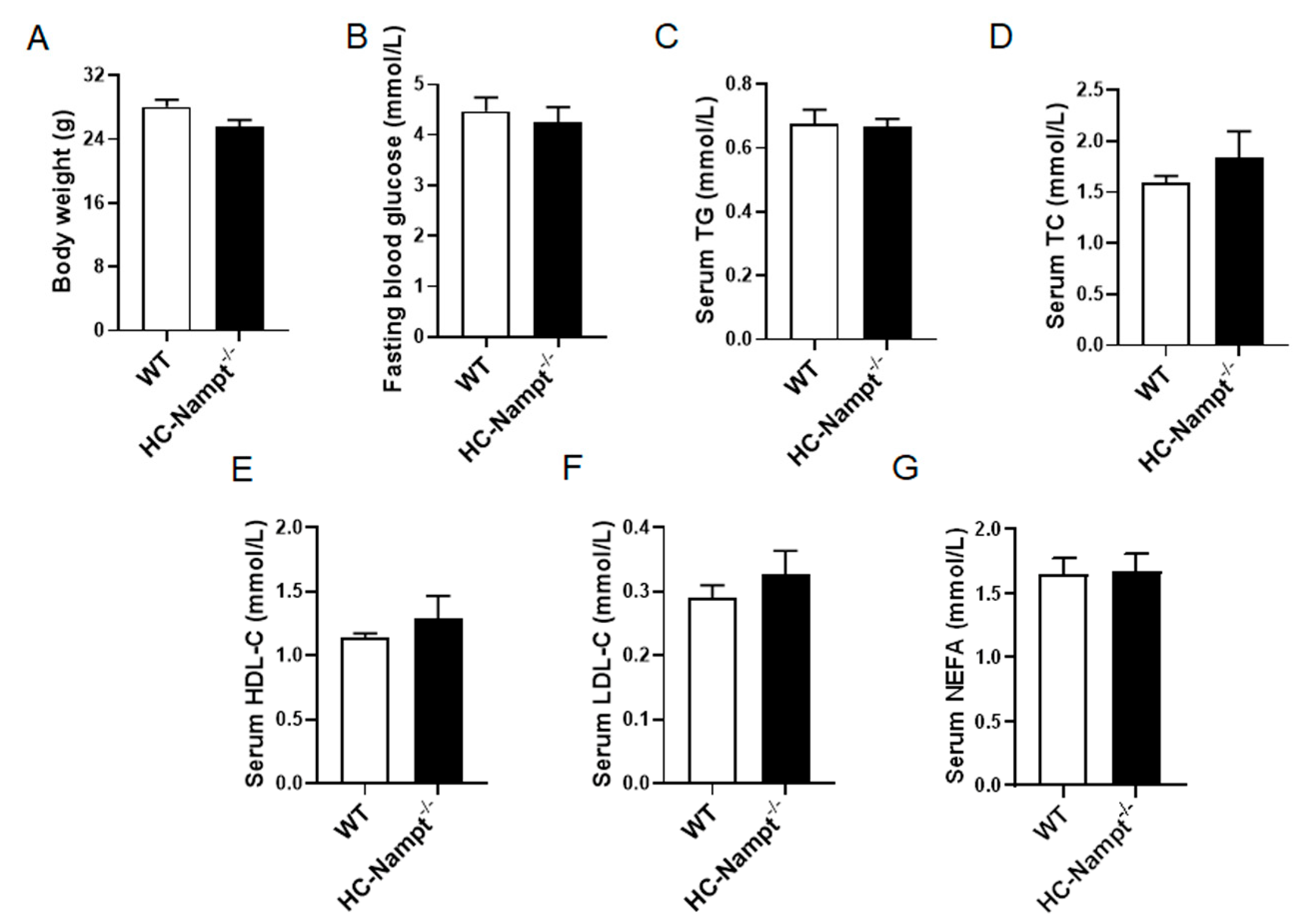
3.2. Deficiency of Hepatic Nampt Has No Effect on Body Weight, Fasting Blood Glucose, and Serum Lipids Levels under Normal Chow Diet
3.3. Deficiency of Hepatic Nampt Does Not Alter Liver Weight and Induce Atherosclerosis in Mice under Normal Chow Diet
3.4. Deficiency of Hepatic Nampt Increases Serum Lipids Levels but Does Not Affect Body Weight and Fasting Blood Glucose in Mice Fed a High-Fat Diet
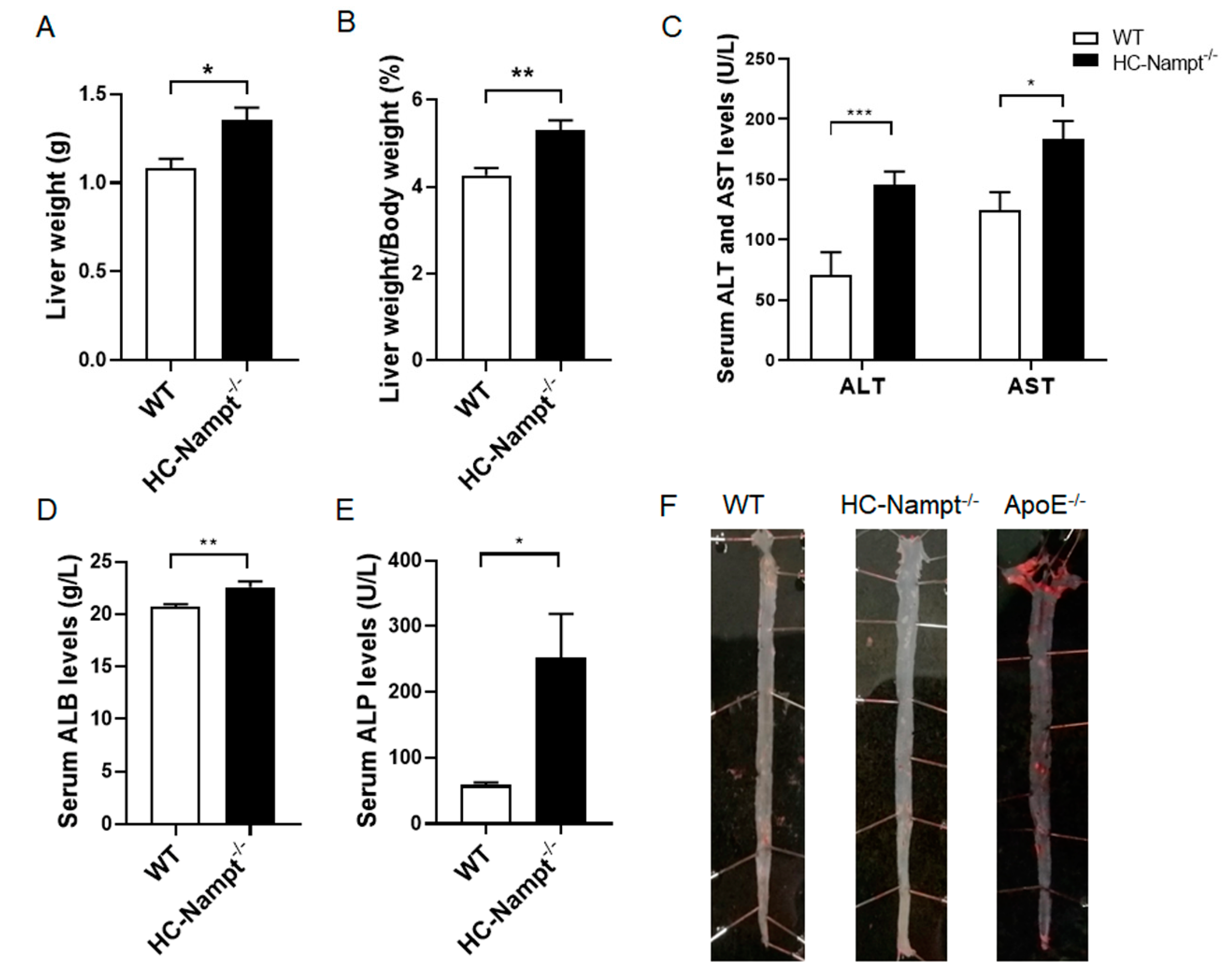
3.5. Deficiency of Hepatic Nampt Increases Liver Weight but Has No Atherosclerosis in Mice Fed a High-Fat Diet
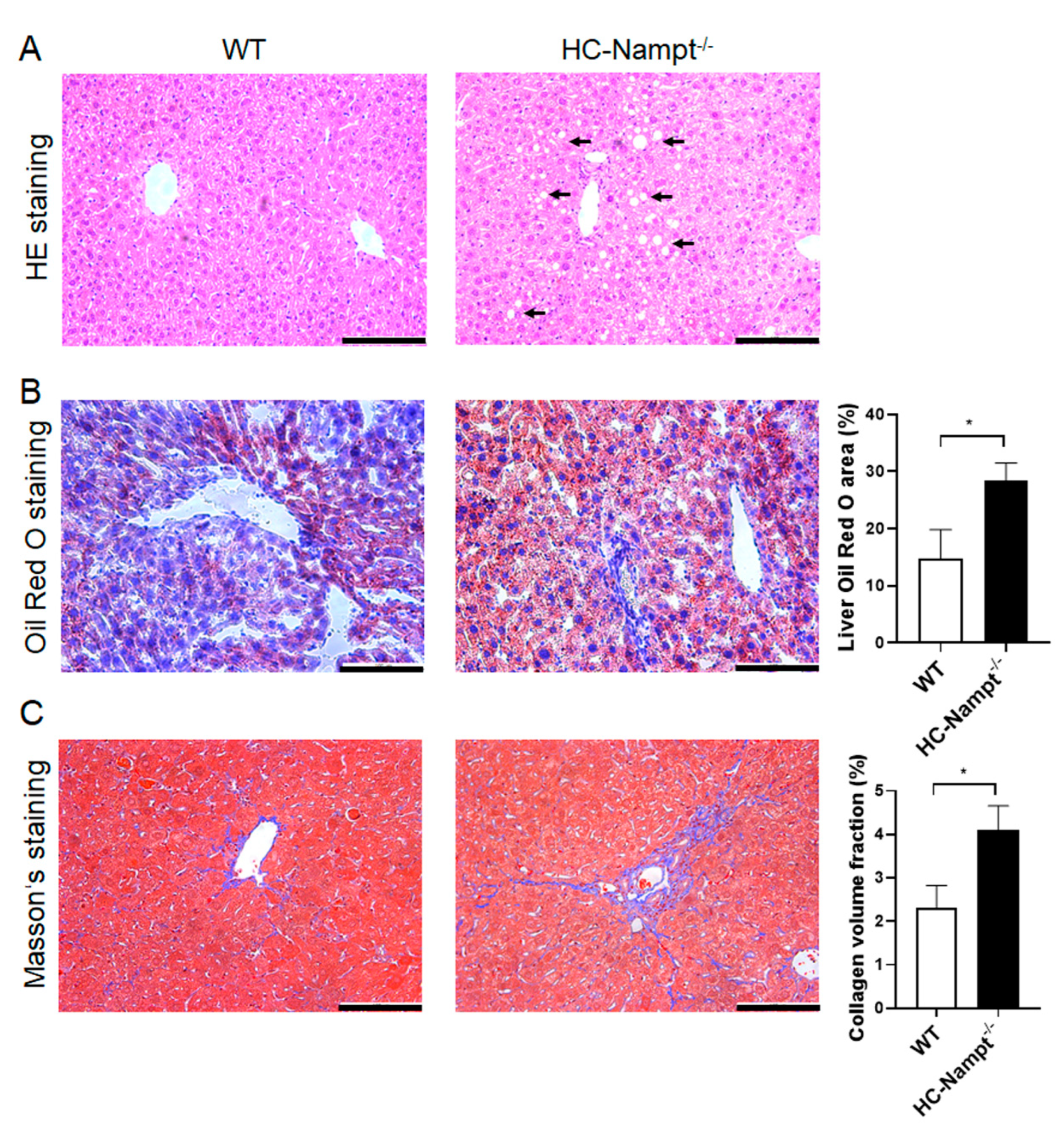
3.6. Deficiency of Hepatic Nampt Induces Liver Histopathological Changes in Mice Fed a High-Fat Diet
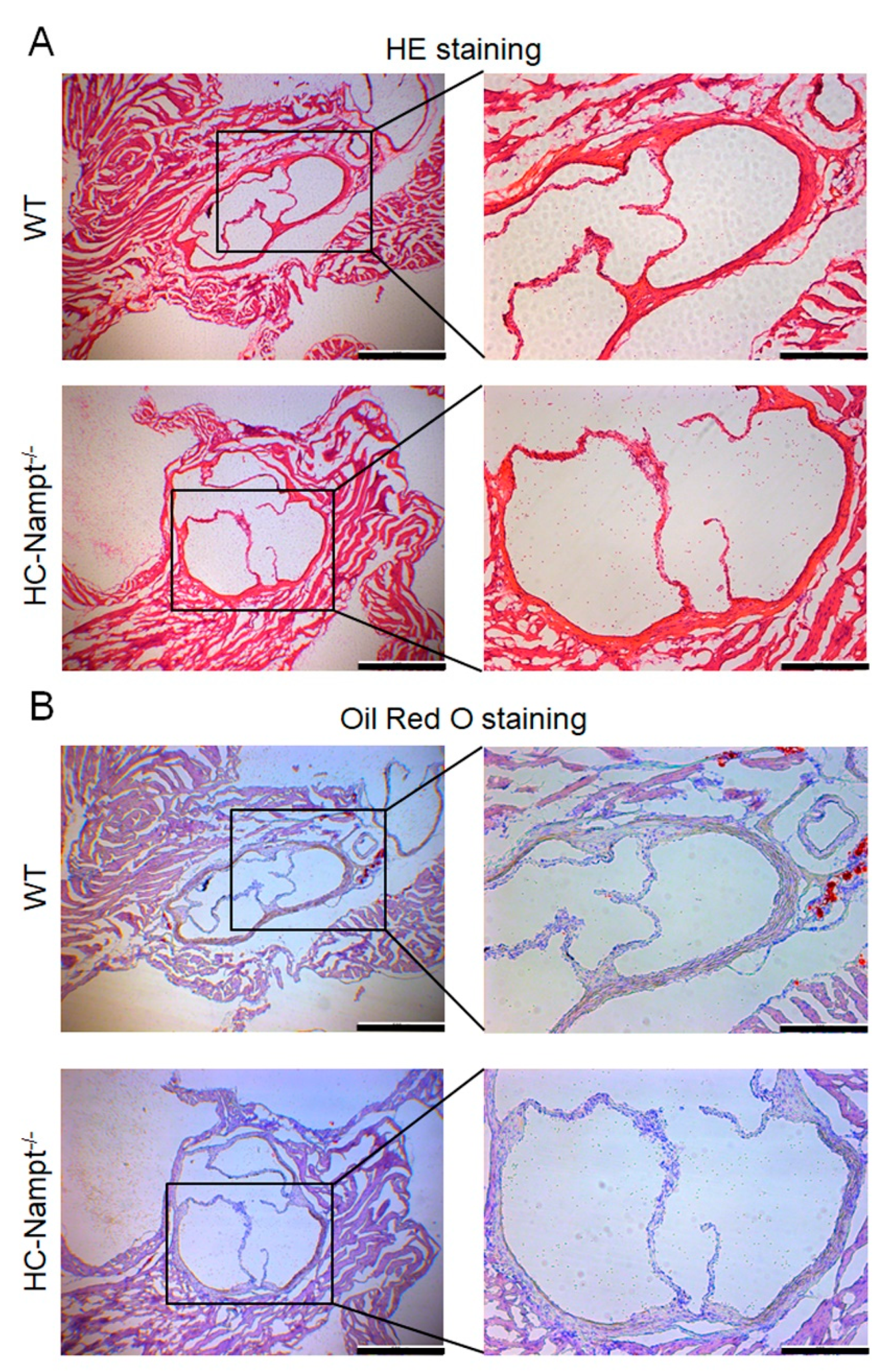
3.7. Deficiency of Hepatic Nampt Does Not Change the Aortic Root Morphology in Mice Fed a High-Fat Diet
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Revollo, J.R.; Körner, A.; Mills, K.F.; Satoh, A.; Wang, T.; Garten, A.; Dasgupta, B.; Sasaki, Y.; Wolberger, C.; Townsend, R.R.; et al. Nampt/PBEF/Visfatin regulates insulin secretion in beta cells as a systemic NAD biosynthetic enzyme. Cell Metab. 2007, 6, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L.; Yeo, D. Maintenance of NAD+ Homeostasis in Skeletal Muscle during Aging and Exercise. Cells 2022, 11, 710. [Google Scholar] [CrossRef] [PubMed]
- Navas, L.E.; Carnero, A. Nicotinamide Adenine Dinucleotide (NAD) Metabolism as a Relevant Target in Cancer. Cells 2022, 11, 2627. [Google Scholar] [CrossRef]
- Song, J.; Ke, S.F.; Zhou, C.C.; Zhang, S.L.; Guan, Y.F.; Xu, T.Y.; Sheng, C.Q.; Wang, P.; Miao, C.Y. Nicotinamide phosphoribosyltransferase is required for the calorie restriction-mediated improvements in oxidative stress, mitochondrial biogenesis, and metabolic adaptation. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 44–57. [Google Scholar] [CrossRef]
- Wang, P.; Miao, C.Y. NAMPT as a Therapeutic Target against Stroke. Trends Pharmacol. Sci. 2015, 36, 891–905. [Google Scholar] [CrossRef]
- Wang, P.; Du, H.; Zhou, C.C.; Song, J.; Liu, X.; Cao, X.; Mehta, J.L.; Shi, Y.; Su, D.F.; Miao, C.Y. Intracellular NAMPT-NAD+-SIRT1 cascade improves post-ischaemic vascular repair by modulating Notch signalling in endothelial progenitors. Cardiovasc. Res. 2014, 104, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.C.; Yang, X.; Hua, X.; Liu, J.; Fan, M.B.; Li, G.Q.; Song, J.; Xu, T.Y.; Li, Z.Y.; Guan, Y.F.; et al. Hepatic NAD(+) deficiency as a therapeutic target for non-alcoholic fatty liver disease in ageing. Br. J. Pharmacol. 2016, 173, 2352–2368. [Google Scholar] [CrossRef]
- Kong, Y.Y.; Li, G.Q.; Zhang, W.J.; Hua, X.; Zhou, C.C.; Xu, T.Y.; Li, Z.Y.; Wang, P.; Miao, C.Y. Nicotinamide phosphoribosyltransferase aggravates inflammation and promotes atherosclerosis in ApoE knockout mice. Acta Pharmacol. Sin. 2019, 40, 1184–1192. [Google Scholar] [CrossRef]
- Dahl, T.B.; Holm, S.; Aukrust, P.; Halvorsen, B. Visfatin/NAMPT: A multifaceted molecule with diverse roles in physiology and pathophysiology. Annu. Rev. Nutr. 2012, 32, 229–243. [Google Scholar] [CrossRef]
- Pouwels, S.; Sakran, N.; Graham, Y.; Leal, A.; Pintar, T.; Yang, W.; Kassir, R.; Singhal, R.; Mahawar, K.; Ramnarain, D. Non-alcoholic fatty liver disease (NAFLD): A review of pathophysiology, clinical management and effects of weight loss. BMC Endocr. Disord. 2022, 22, 63. [Google Scholar] [CrossRef]
- Heeren, J.; Scheja, L. Metabolic-associated fatty liver disease and lipoprotein metabolism. Mol. Metab. 2021, 50, 101238. [Google Scholar] [CrossRef]
- Cohen, D.E.; Fisher, E.A. Lipoprotein metabolism, dyslipidemia, and nonalcoholic fatty liver disease. Semin. Liver Dis. 2013, 33, 380–388. [Google Scholar] [CrossRef]
- Ilbeigi, D.; Nourbakhsh, M.; Pasalar, P.; Meshkani, R.; Shokri Afra, H.; Panahi, G.H.; Borji, M.; Sharifi, R. Nicotinamide Phosphoribosyltransferase Knockdown Leads to Lipid Accumulation in HepG2 Cells through The SIRT1-AMPK Pathway. Cell J. 2020, 22, 125–132. [Google Scholar] [CrossRef] [PubMed]
- McLellan, M.A.; Rosenthal, N.A.; Pinto, A.R. Cre-loxP-Mediated Recombination: General Principles and Experimental Considerations. Curr. Protoc. Mouse Biol. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Li, Y.S.; Meng, R.R.; Chen, X.; Shang, C.L.; Li, H.B.; Zhang, T.J.; Long, H.Y.; Li, H.Q.; Wang, Y.J.; Wang, F.C. Generation of H11-albumin-rtTA Transgenic Mice: A Tool for Inducible Gene Expression in the Liver. G3 (Bethesda) 2019, 9, 591–599. [Google Scholar] [CrossRef]
- Qi, Q.; Hu, W.J.; Zheng, S.L.; Zhang, S.L.; Le, Y.Y.; Li, Z.Y.; Miao, C.Y. Metrnl deficiency decreases blood HDL cholesterol and increases blood triglyceride. Acta Pharmacol. Sin. 2020, 41, 1568–1575. [Google Scholar] [CrossRef]
- Li, Z.Y.; Song, J.; Zheng, S.L.; Fan, M.B.; Guan, Y.F.; Qu, Y.; Xu, J.; Wang, P.; Miao, C.Y. Adipocyte Metrnl Antagonizes Insulin Resistance Through PPARgamma Signaling. Diabetes 2015, 64, 4011–4022. [Google Scholar] [CrossRef]
- Zhang, L.Q.; Nsumu, M.; Huang, P.; Heruth, D.P.; Riordan, S.M.; Shortt, K.; Zhang, N.; Grigoryev, D.N.; Li, D.Y.; Friesen, C.A.; et al. Novel Protective Role of Nicotinamide Phosphoribosyltransferase in Acetaminophen-Induced Acute Liver Injury in Mice. Am. J. Pathol. 2018, 188, 1640–1652. [Google Scholar] [CrossRef] [PubMed]
- Uddin, G.M.; Youngson, N.A.; Chowdhury, S.S.; Hagan, C.; Sinclair, D.A.; Morris, M.J. Administration of Nicotinamide Mononucleotide (NMN) Reduces Metabolic Impairment in Male Mouse Offspring from Obese Mothers. Cells 2020, 9, 791. [Google Scholar] [CrossRef]
- Luo, C.; Ding, W.; Zhu, S.; Chen, Y.; Liu, X.; Deng, H. Nicotinamide Mononucleotide Administration Amends Protein Acetylome of Aged Mouse Liver. Cells 2022, 11, 1654. [Google Scholar] [CrossRef] [PubMed]
- Poltronieri, P.; Celetti, A.; Palazzo, L. Mono(ADP-ribosyl)ation Enzymes and NAD(+) Metabolism: A Focus on Diseases and Therapeutic Perspectives. Cells 2021, 10, 128. [Google Scholar] [CrossRef]
- Li, S.; Wang, C.; Li, K.; Li, L.; Tian, M.; Xie, J.; Yang, M.; Jia, Y.; He, J.; Gao, L.; et al. NAMPT knockdown attenuates atherosclerosis and promotes reverse cholesterol transport in ApoE KO mice with high-fat-induced insulin resistance. Sci. Rep. 2016, 6, 26746. [Google Scholar] [CrossRef]
- Nencioni, A.; da Silva, R.F.; Fraga-Silva, R.A.; Steffens, S.; Fabre, M.; Bauer, I.; Caffa, I.; Magnone, M.; Sociali, G.; Quercioli, A.; et al. Nicotinamide phosphoribosyltransferase inhibition reduces intraplaque CXCL1 production and associated neutrophil infiltration in atherosclerotic mice. Thromb. Haemost. 2014, 111, 308–322. [Google Scholar] [CrossRef]
- Dall, M.; Trammell, S.A.J.; Asping, M.; Hassing, A.S.; Agerholm, M.; Vienberg, S.G.; Gillum, M.P.; Larsen, S.; Treebak, J.T. Mitochondrial function in liver cells is resistant to perturbations in NAD(+) salvage capacity. J. Biol. Chem. 2019, 294, 13304–13326. [Google Scholar] [CrossRef]
- Dall, M.; Hassing, A.S.; Niu, L.; Nielsen, T.S.; Ingerslev, L.R.; Sulek, K.; Trammell, S.A.J.; Gillum, M.P.; Barrès, R.; Larsen, S.; et al. Hepatocyte-specific perturbation of NAD(+) biosynthetic pathways in mice induces reversible nonalcoholic steatohepatitis-like phenotypes. J. Biol. Chem. 2021, 297, 101388. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, P.; Tang, X.; Wang, S.; Shen, J.; Zheng, Y.; Gao, C.; Mi, P.; Zhang, C.; Qu, H.; et al. Metabolic Rewiring of Kynurenine Pathway during Hepatic Ischemia-Reperfusion Injury Exacerbates Liver Damage by Impairing NAD Homeostasis. Adv. Sci. (Weinh) 2022, 9, e2204697. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Wang, X.N.; Huang, C.C.; Hu, L.; Xiao, Y.F.; Guan, X.H.; Qian, Y.S.; Deng, K.Y.; Xin, H.B. Inhibition of NAMPT aggravates high fat diet-induced hepatic steatosis in mice through regulating Sirt1/AMPKα/SREBP1 signaling pathway. Lipids Health Dis. 2017, 16, 82. [Google Scholar] [CrossRef] [PubMed]
- Garten, A.; Schuster, S.; Penke, M.; Gorski, T.; de Giorgis, T.; Kiess, W. Physiological and pathophysiological roles of NAMPT and NAD metabolism. Nat. Rev. Endocrinol. 2015, 11, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Wen, K.; Li, Q.; Huang, C.C.; Zhao, J.L.; Zhao, Q.H.; Xiao, Y.F.; Guan, X.H.; Qian, Y.S.; Gan, L.; et al. CD38 Deficiency Protects Mice from High Fat Diet-Induced Nonalcoholic Fatty Liver Disease through Activating NAD(+)/Sirtuins Signaling Pathways-Mediated Inhibition of Lipid Accumulation and Oxidative Stress in Hepatocytes. Int. J. Biol. Sci. 2021, 17, 4305–4315. [Google Scholar] [CrossRef]
- Jiang, Z.; Wang, H.; Yang, Y.; Yao, Y.; Ma, H. Genistein activated SIRT1-AMPK signaling pathway mediated by ERβ-FOXO1-Nampt to reduce fat accumulation in chicken hepatocytes. Life Sci. 2023, 312, 121259. [Google Scholar] [CrossRef]
- Morató, L.; Astori, S.; Zalachoras, I.; Rodrigues, J.; Ghosal, S.; Huang, W.; Guillot de Suduiraut, I.; Grosse, J.; Zanoletti, O.; Cao, L.; et al. eNAMPT actions through nucleus accumbens NAD(+)/SIRT1 link increased adiposity with sociability deficits programmed by peripuberty stress. Sci. Adv. 2022, 8, eabj9109. [Google Scholar] [CrossRef] [PubMed]
- Caligiuri, A.; Gentilini, A.; Pastore, M.; Gitto, S.; Marra, F. Cellular and Molecular Mechanisms Underlying Liver Fibrosis Regression. Cells 2021, 10, 2759. [Google Scholar] [CrossRef] [PubMed]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride Metabolism in the Liver. Compr. Physiol. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Temel, R.E.; Brown, J.M. A new model of reverse cholesterol transport: EnTICEing strategies to stimulate intestinal cholesterol excretion. Trends Pharmacol. Sci. 2015, 36, 440–451. [Google Scholar] [CrossRef]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef]
- Missaglia, S.; Coleman, R.A.; Mordente, A.; Tavian, D. Neutral Lipid Storage Diseases as Cellular Model to Study Lipid Droplet Function. Cells 2019, 8, 187. [Google Scholar] [CrossRef] [PubMed]
- Selen, E.S.; Choi, J.; Wolfgang, M.J. Discordant hepatic fatty acid oxidation and triglyceride hydrolysis leads to liver disease. JCI Insight 2021, 6, e135626. [Google Scholar] [CrossRef]
- Griffin, J.D.; Bejarano, E.; Wang, X.D.; Greenberg, A.S. Integrated Action of Autophagy and Adipose Tissue Triglyceride Lipase Ameliorates Diet-Induced Hepatic Steatosis in Liver-Specific PLIN2 Knockout Mice. Cells 2021, 10, 1016. [Google Scholar] [CrossRef]
- Xue, Y.; Xu, J.; Li, M.; Gao, Y. Potential screening indicators for early diagnosis of NAFLD/MAFLD and liver fibrosis: Triglyceride glucose index-related parameters. Front. Endocrinol. (Lausanne) 2022, 13, 951689. [Google Scholar] [CrossRef]
- Amzolini, A.M.; Forţofoiu, M.C.; Barău Abu-Alhija, A.; Vladu, I.M.; Clenciu, D.; Mitrea, A.; Forţofoiu, M.; Matei, D.; Enăchescu, V.; Predescu, O.I.; et al. Triglyceride and glucose index: A useful tool for non-alcoholic liver disease assessed by liver biopsy in patients with metabolic syndrome? Rom. J. Morphol. Embryol. 2021, 62, 475–480. [Google Scholar] [CrossRef]
- Yao, Y.; Li, Q.; Gao, P.; Wang, W.; Chen, L.; Zhang, J.; Xu, Y. Glucagon-like peptide-1 contributes to increases ABCA1 expression by downregulating miR-758 to regulate cholesterol homeostasis. Biochem. Biophys. Res. Commun. 2018, 497, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, J.; Zhao, K.; Gao, L.; Zhao, J. Cholesterol-induced toxicity: An integrated view of the role of cholesterol in multiple diseases. Cell Metab. 2021, 33, 1911–1925. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yu, X.H.; Ou, X.; Ouyang, X.P.; Tang, C.K. Hepatic cholesterol transport and its role in non-alcoholic fatty liver disease and atherosclerosis. Prog. Lipid Res. 2021, 83, 101109. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, G.J.; Xiong, X.; Yu, J.; Fan, R.; Zhang, C.; Xu, L.; Sun, X.; Huang, Y.; Wang, Q.; Ruan, H.-B.; et al. NAMPT overexpression alleviates alcohol-induced hepatic steatosis in mice. PLoS ONE 2019, 14, e0212523. [Google Scholar] [CrossRef]
- Brown, G.T.; Kleiner, D.E. Histopathology of nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Metab. Clin. Exp. 2016, 65, 1080–1086. [Google Scholar] [CrossRef]
- Wree, A.; Schlattjan, M.; Bechmann, L.P.; Claudel, T.; Sowa, J.P.; Stojakovic, T.; Scharnagl, H.; Köfeler, H.; Baba, H.A.; Gerken, G.; et al. Adipocyte cell size, free fatty acids and apolipoproteins are associated with non-alcoholic liver injury progression in severely obese patients. Metab. Clin. Exp. 2014, 63, 1542–1552. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse Genes | Accession NCBI | Forward Primer 5′–3′ | Reverse Primer 5′–3′ |
|---|---|---|---|
| Nampt | 59,027 | GCAGAAGCCGAGTTCAACATC | TTTTCACGGCATTCAAAGTAGGA |
| GAPDH | 14,433 | AGGTCGGTGTGAACGGATTTG | TGTAGACCATGTAGTTGAGGTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.-X.; Qing, S.-L.; Miao, Z.-W.; Luo, H.-Y.; Tian, J.-S.; Zhang, X.-P.; Wang, S.-N.; Zhang, T.-G.; Miao, C.-Y. Hepatic Nampt Deficiency Aggravates Dyslipidemia and Fatty Liver in High Fat Diet Fed Mice. Cells 2023, 12, 568. https://doi.org/10.3390/cells12040568
Wang D-X, Qing S-L, Miao Z-W, Luo H-Y, Tian J-S, Zhang X-P, Wang S-N, Zhang T-G, Miao C-Y. Hepatic Nampt Deficiency Aggravates Dyslipidemia and Fatty Liver in High Fat Diet Fed Mice. Cells. 2023; 12(4):568. https://doi.org/10.3390/cells12040568
Chicago/Turabian StyleWang, Dao-Xin, Sheng-Li Qing, Zhu-Wei Miao, Heng-Yu Luo, Jia-Sheng Tian, Xiu-Ping Zhang, Shu-Na Wang, Tian-Guang Zhang, and Chao-Yu Miao. 2023. "Hepatic Nampt Deficiency Aggravates Dyslipidemia and Fatty Liver in High Fat Diet Fed Mice" Cells 12, no. 4: 568. https://doi.org/10.3390/cells12040568
APA StyleWang, D.-X., Qing, S.-L., Miao, Z.-W., Luo, H.-Y., Tian, J.-S., Zhang, X.-P., Wang, S.-N., Zhang, T.-G., & Miao, C.-Y. (2023). Hepatic Nampt Deficiency Aggravates Dyslipidemia and Fatty Liver in High Fat Diet Fed Mice. Cells, 12(4), 568. https://doi.org/10.3390/cells12040568