

Pllans−II: Unveiling the Action Mechanism of a Promising Chemotherapeutic Agent Targeting Cervical Cancer Cell Adhesion and Survival Pathways
 ,
,  , , , , , , and
, , , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Venom Collection and Pllans−II Purification
2.2. Cell Culture and IC50 Determination
2.3. Proliferation Assay
2.4. Visualization of HeLa Morphological Alterations Induced by Pllans−II
2.5. Mitochondrial Membrane Potential Analysis
2.6. Lactate Dehydrogenase (LDH) Assay
2.7. Three-Dimensional Structure of Pllans−II and Molecular Docking with α5β1 Integrin
2.8. Pllans−II Derivatization and Visualization of Interactions with HeLa Cells
2.9. Adhesion Inhibition Evaluation
2.10. Transcriptomic Analysis
2.11. Statistical Analysis
3. Results
3.1. Pllans−II Affects HeLa Cell Proliferation and Triggers Apoptosis
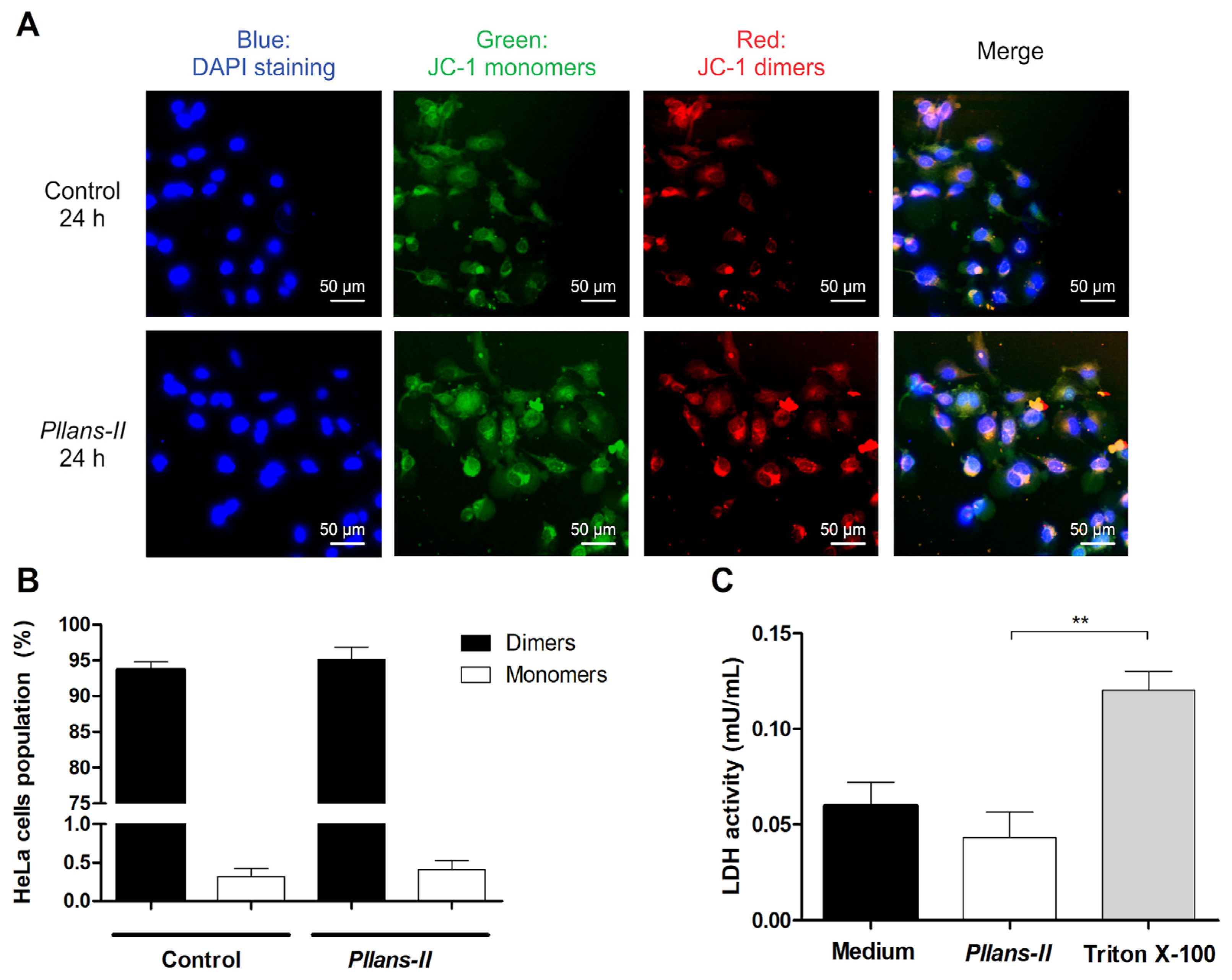
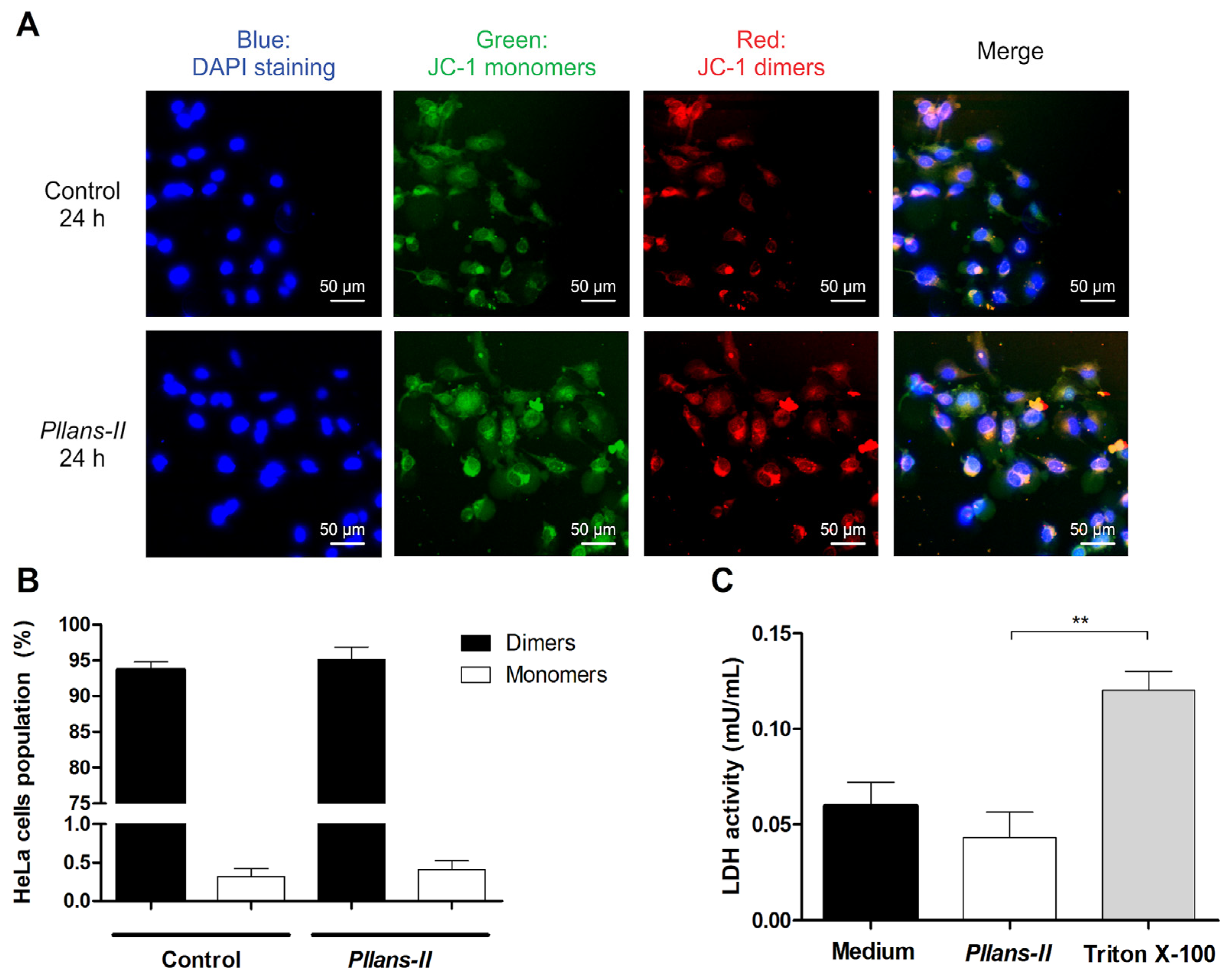
3.2. Pllans−II Does Not Affect the Cytoplasmic Membrane or Mitochondrial Integrity
3.3. Pllans−II Interacts with HeLa Cells at the Cytoplasmic Membrane Level, Disrupting Adhesion Interactions with ECM Proteins
3.4. Pllans−II Interaction with HeLa Cells Affects the Expression of Genes Mainly Related to Adhesion to the ECM and Proliferative Processes
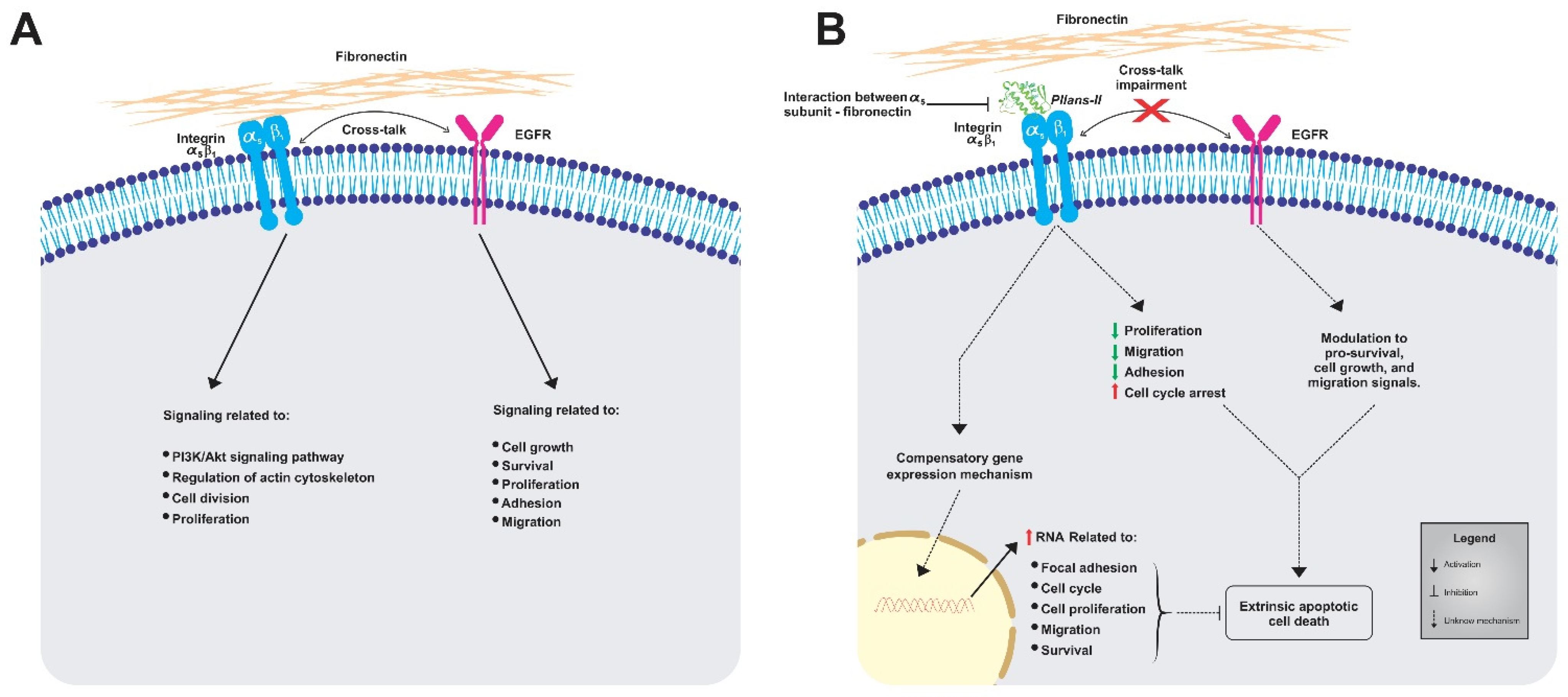
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arbyn, M.; Weiderpass, E.; Bruni, L.; de Sanjosé, S.; Saraiya, M.; Ferlay, J.; Bray, F. Estimates of Incidence and Mortality of Cervical Cancer in 2018: A Worldwide Analysis. Lancet Glob. Health 2020, 8, e191–e203. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bouvard, V.; Wentzensen, N.; Mackie, A.; Berkhof, J.; Brotherton, J.; Giorgi-Rossi, P.; Kupets, R.; Smith, R.; Arrossi, S.; Bendahhou, K. The IARC Perspective on Cervical Cancer Screening. N. Engl. J. Med. 2021, 385, 1908–1918. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.A.; James, D.; Marzan, A.; Armaos, M. Cervical Cancer: An Overview of Pathophysiology and Management. In Seminars in Oncology Nursing; Elsevier: Amsterdam, The Netherlands, 2019; Volume 35, pp. 166–174. [Google Scholar] [CrossRef]
- Rogers, L.; Siu, S.S.N.; Luesley, D.; Bryant, A.; Dickinson, H.O. Radiotherapy and Chemoradiation after Surgery for Early Cervical Cancer. Cochrane Database Syst. Rev. 2012, 5. [Google Scholar] [CrossRef]
- Huang, H.; Feng, Y.-L.; Wan, T.; Zhang, Y.-N.; Cao, X.-P.; Huang, Y.-W.; Xiong, Y.; Huang, X.; Zheng, M.; Li, Y.-F. Effectiveness of Sequential Chemoradiation vs Concurrent Chemoradiation or Radiation Alone in Adjuvant Treatment after Hysterectomy for Cervical Cancer: The STARS Phase 3 Randomized Clinical Trial. JAMA Oncol. 2021, 7, 361–369. [Google Scholar] [CrossRef] [PubMed]
- McSweeney, K.R.; Gadanec, L.K.; Qaradakhi, T.; Ali, B.A.; Zulli, A.; Apostolopoulos, V. Mechanisms of Cisplatin-Induced Acute Kidney Injury: Pathological Mechanisms, Pharmacological Interventions, and Genetic Mitigations. Cancers 2021, 13, 1572. [Google Scholar] [CrossRef] [PubMed]
- Donato, N.J.; Martin, C.A.; Perez, M.; Newman, R.A.; Vidal, J.C.; Etcheverry, M. Regulation of Epidermal Growth Factor Receptor Activity by Crotoxin, a Snake Venom Phospholipase A2 Toxin: A Novel Growth Inhibitory Mechanism. Biochem. Pharmacol. 1996, 51, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Gómez, A.; Montealegre-Sánchez, L.; García-Perdomo, H.A.; Jiménez-Charris, E. Cervical Cancer and Potential Pharmacological Treatment with Snake Venoms. Mol. Biol. Rep. 2020, 47, 4709–4721. [Google Scholar] [CrossRef]
- Jiménez–Charris, E.; Lopes, D.S.; Gimenes, S.N.C.; Teixeira, S.C.; Montealegre–Sánchez, L.; Solano–Redondo, L.; Fierro–Pérez, L.; Ávila, V.d.M.R. Antitumor Potential of Pllans–II, an Acidic Asp49–PLA2 from Porthidium lansbergii lansbergii Snake Venom on Human Cervical Carcinoma HeLa Cells. Int. J. Biol. Macromol. 2019, 122, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Suhr, S.-M.; Kim, D.-S. Identification of the Snake Venom Substance That Induces Apoptosis. Biochem. Biophys. Res. Commun. 1996, 224, 134–139. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, L.-J. ACTX-8, a Cytotoxic L-Amino Acid Oxidase Isolated from Agkistrodon Acutus Snake Venom, Induces Apoptosis in Hela Cervical Cancer Cells. Life Sci. 2007, 80, 1189–1197. [Google Scholar] [CrossRef]
- Berg, O.G.; Gelb, M.H.; Tsai, M.-D.; Jain, M.K. Interfacial Enzymology: The Secreted Phospholipase A2-Paradigm. Chem. Rev. 2001, 101, 2613–2654. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Gómez, A.; Franco, N.R.; Montealegre-Sanchez, L.I.; Solano-Redondo, L.M.; Castillo, A.; Mosquera-Escudero, M.; Jiménez-Charris, E. Pllans–II Induces Cell Death in Cervical Cancer Squamous Epithelial Cells via Unfolded Protein Accumulation and Endoplasmic Reticulum Stress. Molecules 2022, 27, 6491. [Google Scholar] [CrossRef]
- Silva, M.A.; Lopes, D.S.; Teixeira, S.C.; Gimenes, S.N.C.; Azevedo, F.V.P.V.; Polloni, L.; Borges, B.C.; da Silva, M.S.; Barbosa, M.J.; de Oliveira Junior, R.J. Genotoxic Effects of BnSP-6, a Lys-49 Phospholipase A2 (PLA2) Homologue from Bothrops pauloensis Snake Venom, on MDA-MB-231 Breast Cancer Cells. Int. J. Biol. Macromol. 2018, 118, 311–319. [Google Scholar] [CrossRef] [PubMed]
- de Vasconcelos Azevedo, F.V.P.; Zóia, M.A.P.; Lopes, D.S.; Gimenes, S.N.; Vecchi, L.; Alves, P.T.; Rodrigues, R.S.; Silva, A.C.A.; Yoneyama, K.A.G.; Goulart, L.R. Antitumor and Antimetastatic Effects of PLA2-BthTX-II from Bothrops jararacussu Venom on Human Breast Cancer Cells. Int. J. Biol. Macromol. 2019, 135, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Araya, C.; Lomonte, B. Antitumor Effects of Cationic Synthetic Peptides Derived from Lys49 Phospholipase A2 Homologues of Snake Venoms. Cell Biol. Int. 2007, 31, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.R.; Menaldo, D.L.; Oliveira, C.Z.; Santos-Filho, N.A.; Teixeira, S.S.; Nomizo, A.; Fuly, A.L.; Monteiro, M.C.; de Souza, B.M.; Palma, M.S. Myotoxic Phospholipases A2 Isolated from Bothrops brazili Snake Venom and Synthetic Peptides Derived from Their C-Terminal Region: Cytotoxic Effect on Microorganism and Tumor Cells. Peptides 2008, 29, 1645–1656. [Google Scholar] [CrossRef]
- Fujisawa, D.; Yamazaki, Y.; Lomonte, B.; Morita, T. Catalytically Inactive Phospholipase A2 Homologue Binds to Vascular Endothelial Growth Factor Receptor-2 via a C-Terminal Loop Region. Biochem. J. 2008, 411, 515–522. [Google Scholar] [CrossRef]
- Gebrim, L.C.; Marcussi, S.; Menaldo, D.L.; de Menezes, C.S.R.; Nomizo, A.; Hamaguchi, A.; Silveira-Lacerda, E.P.; Homsi-Brandeburgo, M.I.; Sampaio, S.V.; Soares, A.M. Antitumor Effects of Snake Venom Chemically Modified Lys49 Phospholipase A2-like BthTX-I and a Synthetic Peptide Derived from Its C-Terminal Region. Biologicals 2009, 37, 222–229. [Google Scholar] [CrossRef]
- Osipov, A.V.; Utkin, Y.N. Antiproliferative Effects of Snake Venom Phospholipases A2 and Their Perspectives for Cancer Treatment. In Toxins and Drug Discovery; Springer: Dordrecht, The Netherlands, 2015; pp. 129–146. [Google Scholar] [CrossRef]
- Lomonte, B.; Angulo, Y.; Moreno, E. Synthetic Peptides Derived from the C-Terminal Region of Lys49 Phospholipase A2 Homologues from Viperidae Snake Venoms: Biomimetic Activities and Potential Applications. Curr. Pharm. Des. 2010, 16, 3224–3230. [Google Scholar] [CrossRef]
- Massimino, M.L.; Simonato, M.; Spolaore, B.; Franchin, C.; Arrigoni, G.; Marin, O.; Monturiol-Gross, L.; Fernández, J.; Lomonte, B.; Tonello, F. Cell Surface Nucleolin Interacts with and Internalizes Bothrops asper Lys49 Phospholipase A2 and Mediates Its Toxic Activity. Sci. Rep. 2018, 8, 10619. [Google Scholar] [CrossRef] [PubMed]
- Berger, C.M.; Gaume, X.; Bouvet, P. The Roles of Nucleolin Subcellular Localization in Cancer. Biochimie 2015, 113, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.S.; Izidoro, L.F.M.; De Oliveira, J.R.; Robson, J.; Sampaio, S.V.; Soares, A.M.; Rodrigues, V.M. Snake Venom Phospholipases A2: A New Class of Antitumor Agents. Protein Pept. Lett. 2009, 16, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Charris, E.; Montealegre-Sánchez, L.; Solano-Redondo, L.; Castro-Herrera, F.; Fierro-Pérez, L.; Lomonte, B. Divergent Functional Profiles of Acidic and Basic Phospholipases A2 in the Venom of the Snake Porthidium lansbergii lansbergii. Toxicon 2016, 119, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Di Veroli, G.Y.; Fornari, C.; Goldlust, I.; Mills, G.; Koh, S.B.; Bramhall, J.L.; Richards, F.M.; Jodrell, D.I. An Automated Fitting Procedure and Software for Dose-Response Curves with Multiphasic Features. Sci. Rep. 2015, 5, 14701. [Google Scholar] [CrossRef]
- Möbius, W.; Cooper, B.; Kaufmann, W.A.; Imig, C.; Ruhwedel, T.; Snaidero, N.; Saab, A.S.; Varoqueaux, F. Electron Microscopy of the Mouse Central Nervous System. Methods Cell Biol. 2010, 96, 475–512. [Google Scholar] [CrossRef]
- Cossarizza, A.; Baccaranicontri, M.; Kalashnikova, G.; Franceschi, C. A New Method for the Cytofluorometric Analysis of Mitochondrial Membrane Potential Using the J-Aggregate Forming Lipophilic Cation 5, 5′, 6, 6′-Tetrachloro-1, 1′, 3, 3′-Tetraethylbenzimidazolcarbocyanine Iodide (JC-1). Biochem. Biophys. Res. Commun. 1993, 197, 40–45. [Google Scholar] [CrossRef]
- Reers, M.; Smith, T.W.; Chen, L.B. J-Aggregate Formation of a Carbocyanine as a Quantitative Fluorescent Indicator of Membrane Potential. Biochemistry 1991, 30, 4480–4486. [Google Scholar] [CrossRef]
- Smiley, S.T.; Reers, M.; Mottola-Hartshorn, C.; Lin, M.; Chen, A.; Smith, T.W.; Steele, G.D., Jr.; Chen, L.B. Intracellular Heterogeneity in Mitochondrial Membrane Potentials Revealed by a J-Aggregate-Forming Lipophilic Cation JC-1. Proc. Natl. Acad. Sci. USA 1991, 88, 3671–3675. [Google Scholar] [CrossRef]
- Gu, L.; Zhang, H.; Song, S.; Zhou, Y.; Lin, Z. Structure of an Acidic Phospholipase A2 from the Venom of Deinagkistrodon Acutus. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002, 58, 104–110. [Google Scholar] [CrossRef]
- Benkert, P.; Künzli, M.; Schwede, T. QMEAN Server for Protein Model Quality Estimation. Nucleic Acids Res. 2009, 37 (Suppl. 2), W510–W514. [Google Scholar] [CrossRef] [PubMed]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro Web Server for Protein–Protein Docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Graille, M.; Pagano, M.; Rose, T.; Ravaux, M.R.; Van Tilbeurgh, H. Zinc Induces Structural Reorganization of Gelatin Binding Domain from Human Fibronectin and Affects Collagen Binding. Structure 2010, 18, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Sinha, A.; Dasgupta, S.; Mukherjee, A.K. Binding of a Naja Naja Venom Acidic Phospholipase A2 Cognate Complex to Membrane-Bound Vimentin of Rat L6 Cells: Implications in Cobra Venom-Induced Cytotoxicity. Biochim. Biophys. Acta (BBA)-Biomembr. 2019, 1861, 958–977. [Google Scholar] [CrossRef]
- Holzer, M.; Mackessy, S.P. An Aqueous Endpoint Assay of Snake Venom Phospholipase A2. Toxicon 1996, 34, 1149–1155. [Google Scholar] [CrossRef] [PubMed]
- Varol, M. Cell-Extracellular Matrix Adhesion Assay. In Epidermal Cells Methods and Protocols; Humana Press: New York, NY, USA, 2020; pp. 209–217. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. Omi. A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Spolaore, B.; Fernández, J.; Lomonte, B.; Massimino, M.L.; Tonello, F. Enzymatic Labelling of Snake Venom Phospholipase A2 Toxins. Toxicon 2019, 170, 99–107. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Chen, Z.; Wang, Z.-H.; Wang, H.; Huang, L.-M. Prognostic Significance of α5β1-Integrin Expression in Cervical Cancer. Asian Pac. J. Cancer Prev. 2013, 14, 3891–3895. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, A.; Li, S.; Tao, X.; Sheng, B.; Chetry, M.; Zhu, X. Predictive Role of Galectin-1 and Integrin α5β1 in Cisplatin-Based Neoadjuvant Chemotherapy of Bulky Squamous Cervical Cancer. Biosci. Rep. 2017, 37, BSR20170958. [Google Scholar] [CrossRef]
- Nagae, M.; Re, S.; Mihara, E.; Nogi, T.; Sugita, Y.; Takagi, J. Crystal Structure of α5β1 Integrin Ectodomain: Atomic Details of the Fibronectin Receptor. J. Cell Biol. 2012, 197, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, F.; Ray, A.M.; Dontenwill, M. Integrin α5β1, the Fibronectin Receptor, as a Pertinent Therapeutic Target in Solid Tumors. Cancers 2013, 5, 27–47. [Google Scholar] [CrossRef]
- Qian, F.; Zhang, Z.-C.; Wu, X.-F.; Li, Y.-P.; Xu, Q. Interaction between Integrin α5 and Fibronectin Is Required for Metastasis of B16F10 Melanoma Cells. Biochem. Biophys. Res. Commun. 2005, 333, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Murillo, C.A.; Rychahou, P.G.; Evers, B.M. Inhibition of α5 Integrin Decreases PI3K Activation and Cell Adhesion of Human Colon Cancers. Surgery 2004, 136, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Vuori, K.; Reed, J.C.; Ruoslahti, E. The Alpha 5 Beta 1 Integrin Supports Survival of Cells on Fibronectin and Up-Regulates Bcl-2 Expression. Proc. Natl. Acad. Sci. USA 1995, 92, 6161–6165. [Google Scholar] [CrossRef] [PubMed]
- Han, S.W.; Roman, J. Fibronectin Induces Cell Proliferation and Inhibits Apoptosis in Human Bronchial Epithelial Cells: Pro-Oncogenic Effects Mediated by PI3-Kinase and NF-kB. Oncogene 2006, 25, 4341–4349. [Google Scholar] [CrossRef] [PubMed]
- Guha, D.; Saha, T.; Bose, S.; Chakraborty, S.; Dhar, S.; Khan, P.; Adhikary, A.; Das, T.; Sa, G. Integrin-EGFR Interaction Regulates Anoikis Resistance in Colon Cancer Cells. Apoptosis 2019, 24, 958–971. [Google Scholar] [CrossRef]
- Zhou, X.; Zhai, Y.; Liu, C.; Yang, G.; Guo, J.; Li, G.; Sun, C.; Qi, X.; Li, X.; Guan, F. Sialidase NEU1 Suppresses Progression of Human Bladder Cancer Cells by Inhibiting Fibronectin-Integrin α5β1 Interaction and Akt Signaling Pathway. Cell Commun. Signal. 2020, 18, 44. [Google Scholar] [CrossRef] [PubMed]
- El-Brolosy, M.A.; Stainier, D.Y.R. Genetic Compensation: A Phenomenon in Search of Mechanisms. PLoS Genet. 2017, 13, e1006780. [Google Scholar] [CrossRef]
- Trusolino, L.; Bertotti, A. Compensatory Pathways in Oncogenic Kinase Signaling and Resistance to Targeted Therapies: Six Degrees of Separation. Cancer Discov. 2012, 2, 876–880. [Google Scholar] [CrossRef]
- Wang, J.; Deng, L.; Huang, J.; Cai, R.; Zhu, X.; Liu, F.; Wang, Q.; Zhang, J.; Zheng, Y. High Expression of Fibronectin 1 Suppresses Apoptosis through the NF-kB Pathway and Is Associated with Migration in Nasopharyngeal Carcinoma. Am. J. Transl. Res. 2017, 9, 4502. [Google Scholar] [PubMed]
- Deng, Z.; Wang, H.; Liu, J.; Deng, Y.; Zhang, N. Comprehensive Understanding of Anchorage-Independent Survival and Its Implication in Cancer Metastasis. Cell Death Dis. 2021, 12, 629. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; Zhang, X. Upregulation of CCT3 Promotes Cervical Cancer Progression through FN1. Mol. Med. Rep. 2021, 24, 856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, F.; Fu, Y.; Chen, X.; Zhang, D. MiR-520d-5p Functions as a Tumor-Suppressor Gene in Cervical Cancer through Targeting PTK2. Life Sci. 2020, 254, 117558. [Google Scholar] [CrossRef]
- Bossler, F.; Hoppe-Seyler, K.; Hoppe-Seyler, F. PI3K/AKT/MTOR Signaling Regulates the Virus/Host Cell Crosstalk in HPV-Positive Cervical Cancer Cells. Int. J. Mol. Sci. 2019, 20, 2188. [Google Scholar] [CrossRef]
- Hoppe-Seyler, K.; Bossler, F.; Lohrey, C.; Bulkescher, J.; Rösl, F.; Jansen, L.; Mayer, A.; Vaupel, P.; Dürst, M.; Hoppe-Seyler, F. Induction of Dormancy in Hypoxic Human Papillomavirus-Positive Cancer Cells. Proc. Natl. Acad. Sci. USA 2017, 114, E990–E998. [Google Scholar] [CrossRef]
- Hang, Q.; Isaji, T.; Hou, S.; Im, S.; Fukuda, T.; Gu, J. Integrin α5 Suppresses the Phosphorylation of Epidermal Growth Factor Receptor and Its Cellular Signaling of Cell Proliferation via N-Glycosylation. J. Biol. Chem. 2015, 290, 29345–29360. [Google Scholar] [CrossRef]
- Kuwada, S.K.; Li, X. Integrin α5/β1 Mediates Fibronectin-Dependent Epithelial Cell Proliferation through Epidermal Growth Factor Receptor Activation. Mol. Biol. Cell 2000, 11, 2485–2496. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montoya-Gómez, A.; Tonello, F.; Spolaore, B.; Massimino, M.L.; Montealegre-Sánchez, L.; Castillo, A.; Rivera Franco, N.; Sevilla-Sánchez, M.J.; Solano-Redondo, L.M.; Mosquera-Escudero, M.; et al. Pllans−II: Unveiling the Action Mechanism of a Promising Chemotherapeutic Agent Targeting Cervical Cancer Cell Adhesion and Survival Pathways. Cells 2023, 12, 2715. https://doi.org/10.3390/cells12232715
Montoya-Gómez A, Tonello F, Spolaore B, Massimino ML, Montealegre-Sánchez L, Castillo A, Rivera Franco N, Sevilla-Sánchez MJ, Solano-Redondo LM, Mosquera-Escudero M, et al. Pllans−II: Unveiling the Action Mechanism of a Promising Chemotherapeutic Agent Targeting Cervical Cancer Cell Adhesion and Survival Pathways. Cells. 2023; 12(23):2715. https://doi.org/10.3390/cells12232715
Chicago/Turabian StyleMontoya-Gómez, Alejandro, Fiorella Tonello, Barbara Spolaore, Maria Lina Massimino, Leonel Montealegre-Sánchez, Andrés Castillo, Nelson Rivera Franco, María José Sevilla-Sánchez, Luis Manuel Solano-Redondo, Mildrey Mosquera-Escudero, and et al. 2023. "Pllans−II: Unveiling the Action Mechanism of a Promising Chemotherapeutic Agent Targeting Cervical Cancer Cell Adhesion and Survival Pathways" Cells 12, no. 23: 2715. https://doi.org/10.3390/cells12232715
APA StyleMontoya-Gómez, A., Tonello, F., Spolaore, B., Massimino, M. L., Montealegre-Sánchez, L., Castillo, A., Rivera Franco, N., Sevilla-Sánchez, M. J., Solano-Redondo, L. M., Mosquera-Escudero, M., & Jiménez-Charris, E. (2023). Pllans−II: Unveiling the Action Mechanism of a Promising Chemotherapeutic Agent Targeting Cervical Cancer Cell Adhesion and Survival Pathways. Cells, 12(23), 2715. https://doi.org/10.3390/cells12232715