
Zebrafish: A Model Deciphering the Impact of Flavonoids on Neurodegenerative Disorders

 ,
, 
 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Flavonoids
2.1. Chemical Structure and Subclasses of Flavonoids
2.1.1. Isoflavones
2.1.2. Flavonols
2.1.3. Flavones
2.1.4. Flavanones
2.1.5. Flavanols
2.1.6. Anthocyanidins
2.2. Bioavailability of Flavonoid in the Brain
3. The Roles of Flavonoids in Neurodegenerative Diseases
3.1. Neuroinflammation in Neurodegenerative Disorders
3.2. Neuroinflammation: Modulation by Flavonoids
3.3. Oxidative Stress in Neurodegenerative Diseases
3.4. Antioxidant Effect of Flavonoids in Neurodegenerative Diseases
3.5. Neuroplasticity: A Combined Process of Neurogenesis and Synaptogenesis
3.6. Neuroplasticity and Effects of Flavonoids
3.7. Reduction of Neuropathological Protein Accumulation
4. Zebrafish as Neurodegenerative Model in Translational Research
5. Effects of Flavonoids against Neurodegenerative Disorders in Zebrafish Model
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mathur, S.; Hoskins, C. Drug Development: Lessons from Nature. Biomed. Rep. 2017, 6, 612–614. [Google Scholar] [CrossRef] [PubMed]
- Nisar, B.; Sultan, A.; Rubab, S.L. Comparison of Medicinally Important Natural Products versus Synthetic Drugs-A Short Commentary. Nat. Prod. Chem. Res. 2018, 6, 308. [Google Scholar] [CrossRef]
- Farooq, S.; Ngaini, Z. Natural and Synthetic Drugs as Potential Treatment for Coronavirus Disease 2019 (COVID-2019). Chem. Afr. 2021, 4, 1–13. [Google Scholar] [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and Human Health: Prevention of Disease and Mechanisms of Action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yang, S.; Wang, K.; Bao, X.; Liu, Y.; Zhou, S.; Liu, H.; Qiu, Y.; Wang, T.; Yu, H. Alkaloids from Traditional Chinese Medicine against Hepatocellular Carcinoma. Biomed. Pharmacother. 2019, 120, 109543. [Google Scholar] [CrossRef] [PubMed]
- Masyita, A.; Mustika Sari, R.; Dwi Astuti, A.; Yasir, B.; Rahma Rumata, N.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and Terpenoids as Main Bioactive Compounds of Essential Oils, Their Roles in Human Health and Potential Application as Natural Food Preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef]
- Aquila, S.; Giner, R.M.; Recio, M.C.; Spegazzini, E.D.; Ríos, J.L. Anti-Inflammatory Activity of Flavonoids from Cayaponia Tayuya Roots. J. Ethnopharmacol. 2009, 121, 333–337. [Google Scholar] [CrossRef]
- Ginwala, R.; Bhavsar, R.; Chigbu, D.I.; Jain, P.; Khan, Z.K. Potential Role of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin. Antioxidants 2019, 8, 35. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, H.; Guan, X.; Zhou, Z. The Anti-Aging Potential of Neohesperidin and Its Synergistic Effects with Other Citrus Flavonoids in Extending Chronological Lifespan of Saccharomyces Cerevisiae BY4742. Molecules 2019, 24, 4093. [Google Scholar] [CrossRef]
- Elangovan, V.; Sekar, N.; Govindasamy, S. Chemopreventive Potential of Dietary Bioflavonoids against 20-Methylcholanthrene-Induced Tumorigenesis. Cancer Lett. 1994, 87, 107–113. [Google Scholar] [CrossRef]
- Ponzo, V.; Goitre, I.; Fadda, M.; Gambino, R.; De Francesco, A.; Soldati, L.; Gentile, L.; Magistroni, P.; Cassader, M.; Bo, S. Dietary Flavonoid Intake and Cardiovascular Risk: A Population-Based Cohort Study. J. Transl. Med. 2015, 13, 218. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Pandey, V.; Vadnere, G.P.; Lodhi, S. Role of Flavonoids in Management of Inflammatory Disorders. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases; Elsevier: Amsterdam, The Netherlands, 2019; pp. 293–322. [Google Scholar]
- Putteeraj, M.; Lim, W.L.; Teoh, S.L.; Yahaya, M.F. Flavonoids and Its Neuroprotective Effects on Brain Ischemia and Neurodegenerative Diseases. Curr. Drug Targets 2018, 19, 1710–1720. [Google Scholar] [CrossRef] [PubMed]
- Socci, V.; Tempesta, D.; Desideri, G.; De Gennaro, L.; Ferrara, M. Enhancing Human Cognition with Cocoa Flavonoids. Front. Nutr. 2017, 4, 19. [Google Scholar] [CrossRef]
- Godos, J.; Caraci, F.; Castellano, S.; Currenti, W.; Galvano, F.; Ferri, R.; Grosso, G. Association between Dietary Flavonoids Intake and Cognitive Function in an Italian Cohort. Biomolecules 2020, 10, 1300. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.M.; Abd El Mohsen, M.; Vauzour, D.; Rendeiro, C.; Butler, L.T.; Ellis, J.A.; Whiteman, M.; Spencer, J.P. Blueberry-Induced Changes in Spatial Working Memory Correlate with Changes in Hippocampal CREB Phosphorylation and Brain-Derived Neurotrophic Factor (BDNF) Levels. Free Radic. Biol. Med. 2008, 45, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Horwood, J.M.; Dufour, F.; Laroche, S.; Davis, S. Signalling Mechanisms Mediated by the Phosphoinositide 3-kinase/Akt Cascade in Synaptic Plasticity and Memory in the Rat. Eur. J. Neurosci. 2006, 23, 3375–3384. [Google Scholar] [CrossRef]
- Stahr, P.-L.; Grewal, R.; Eckert, G.P.; Keck, C.M. Investigating Hesperetin Nanocrystals with Tailor-Made Sizes for the Prevention and Treatment of Alzheimer’s Disease. Drug Deliv. Transl. Res. 2021, 11, 659–674. [Google Scholar] [CrossRef]
- Kheradmand, E.; Hajizadeh Moghaddam, A.; Zare, M. Neuroprotective Effect of Hesperetin and Nano-Hesperetin on Recognition Memory Impairment and the Elevated Oxygen Stress in Rat Model of Alzheimer’s Disease. Biomed Pharm. 2018, 97, 1096–1101. [Google Scholar] [CrossRef]
- Boyina, H.K.; Geethakhrishnan, S.L.; Panuganti, S.; Gangarapu, K.; Devarakonda, K.P.; Bakshi, V.; Guggilla, S.R. In Silico and in Vivo Studies on Quercetin as Potential Anti-Parkinson Agent. In GeNeDis 2018; Springer: Cham, Switzerland, 2020; pp. 1–11. [Google Scholar]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.-M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the Adult Human Hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef]
- Curtis, M.A.; Penney, E.B.; Pearson, A.G.; van Roon-Mom, W.M.; Butterworth, N.J.; Dragunow, M.; Connor, B.; Faull, R.L. Increased Cell Proliferation and Neurogenesis in the Adult Human Huntington’s Disease Brain. Proc. Natl. Acad. Sci. USA 2003, 100, 9023–9027. [Google Scholar] [CrossRef]
- Colucci-D’Amato, L.; Bonavita, V.; di Porzio, U. The End of the Central Dogma of Neurobiology: Stem Cells and Neurogenesis in Adult CNS. Neurol. Sci. 2006, 27, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Beylin, A.; Tanapat, P.; Reeves, A.; Shors, T.J. Learning Enhances Adult Neurogenesis in the Hippocampal Formation. Nat. Neurosci. 1999, 2, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Reznikov, K.Y. Cell Proliferation and Cytogenesis in the Mouse Hippocampus; Springer: Berlin/Heidelberg, Germany, 1991; Volume 122, ISBN 3-642-76447-9. [Google Scholar]
- Luskin, M.B. Restricted Proliferation and Migration of Postnatally Generated Neurons Derived from the Forebrain Subventricular Zone. Neuron 1993, 11, 173–189. [Google Scholar] [CrossRef]
- Gonzalez-Perez, O. Neural Stem Cells in the Adult Human Brain. Biol. Biomed. Rep. 2012, 2, 59–69. [Google Scholar] [PubMed]
- Lois, C.; Alvarez-Buylla, A. Long-Distance Neuronal Migration in the Adult Mammalian Brain. Science 1994, 264, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Haughey, N.J.; Nath, A.; Chan, S.L.; Borchard, A.; Rao, M.S.; Mattson, M.P. Disruption of Neurogenesis by Amyloid Β-peptide, and Perturbed Neural Progenitor Cell Homeostasis, in Models of Alzheimer’s Disease. J. Neurochem. 2002, 83, 1509–1524. [Google Scholar] [CrossRef]
- Thompson, A.; Boekhoorn, K.; Van Dam, A.-M.; Lucassen, P.J. Changes in Adult Neurogenesis in Neurodegenerative Diseases: Cause or Consequence? Genes Brain Behav. 2008, 7, 28–42. [Google Scholar] [CrossRef]
- Hua, F.-Z.; Ying, J.; Zhang, J.; Wang, X.-F.; Hu, Y.-H.; Liang, Y.-P.; Liu, Q.; Xu, G.-H. Naringenin Pre-Treatment Inhibits Neuroapoptosis and Ameliorates Cognitive Impairment in Rats Exposed to Isoflurane Anesthesia by Regulating the PI3/Akt/PTEN Signalling Pathway and Suppressing NF-ΚB-Mediated Inflammation. Int. J. Mol. Med. 2016, 38, 1271–1280. [Google Scholar] [CrossRef]
- Karimipour, M.; Rahbarghazi, R.; Tayefi, H.; Shimia, M.; Ghanadian, M.; Mahmoudi, J.; Bagheri, H.S. Quercetin Promotes Learning and Memory Performance Concomitantly with Neural Stem/Progenitor Cell Proliferation and Neurogenesis in the Adult Rat Dentate Gyrus. Int. J. Dev. Neurosci. 2019, 74, 18–26. [Google Scholar] [CrossRef]
- Calis, Z.; Mogulkoc, R.; Baltaci, A.K. The Roles of Flavonols/Flavonoids in Neurodegeneration and Neuroinflammation. Mini Rev. Med. Chem. 2020, 20, 1475–1488. [Google Scholar] [CrossRef]
- Vafeiadou, K.; Vauzour, D.; Lee, H.Y.; Rodriguez-Mateos, A.; Williams, R.J.; Spencer, J.P. The Citrus Flavanone Naringenin Inhibits Inflammatory Signalling in Glial Cells and Protects against Neuroinflammatory Injury. Arch. Biochem. Biophys. 2009, 484, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Park, C.M.; Song, Y.-S. Luteolin and Luteolin-7-O-Glucoside Protect against Acute Liver Injury through Regulation of Inflammatory Mediators and Antioxidative Enzymes in GalN/LPS-Induced Hepatitic ICR Mice. Nutr. Res. Pract. 2019, 13, 473–479. [Google Scholar] [CrossRef]
- Ali, F.; Jyoti, S.; Naz, F.; Ashafaq, M.; Shahid, M.; Siddique, Y.H. Therapeutic Potential of Luteolin in Transgenic Drosophila Model of Alzheimer’s Disease. Neurosci. Lett. 2019, 692, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Wang, Y.; Sang, J.; Cui, W.; Zhao, W.; Wei, W.; Chen, B.; Lu, F.; Liu, F. Dihydromyricetin Inhibits α-Synuclein Aggregation, Disrupts Preformed Fibrils, and Protects Neuronal Cells in Culture against Amyloid-Induced Cytotoxicity. J. Agric. Food Chem. 2019, 67, 3946–3955. [Google Scholar] [CrossRef] [PubMed]
- Montalbano, G.; Mhalhel, K.; Briglia, M.; Levanti, M.; Abbate, F.; Guerrera, M.C.; D’Alessandro, E.; Laurà, R.; Germanà, A. Zebrafish and Flavonoids: Adjuvants against Obesity. Molecules 2021, 26, 3014. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.-B.; He, K.-J.; Wang, F.; Liu, C.-F. Advances of Zebrafish in Neurodegenerative Disease: From Models to Drug Discovery. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Saleem, S.; Kannan, R.R. Zebrafish: An Emerging Real-Time Model System to Study Alzheimer’s Disease and Neurospecific Drug Discovery. Cell Death Discov. 2018, 4, 45. [Google Scholar] [CrossRef]
- Tanwar, B.; Modgil, R. Flavonoids: Dietary Occurrence and Health Benefits. Spatula Dd 2012, 2, 59–68. [Google Scholar] [CrossRef]
- Havsteen, B.H. The Biochemistry and Medical Significance of the Flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Lois, R. Accumulation of UV-Absorbing Flavonoids Induced by UV-B Radiation in Arabidopsis Thaliana L. Planta 1994, 194, 498–503. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.-J.; Reddy, M.S.S.; Wang, L. The Phenylpropanoid Pathway and Plant Defence—A Genomics Perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, J.-L.; Austin, M.B.; Stewart, C.; Noel, J.P. Structure and Function of Enzymes Involved in the Biosynthesis of Phenylpropanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Brodowska, K.M. Natural flavonoids: Classification, potential role, and application of flavonoid analogues. Eur. J. Biol. Res. 2017, 7, 108–123. [Google Scholar]
- Dixon, R.A.; Sumner, L.W. Legume Natural Products: Understanding and Manipulating Complex Pathways for Human and Animal Health. Plant Physiol. 2003, 131, 878–885. [Google Scholar] [CrossRef]
- Rípodas, C.; Via, V.D.; Aguilar, O.M.; Zanetti, M.E.; Blanco, F.A. Knock-down of a Member of the Isoflavone Reductase Gene Family Impairs Plant Growth and Nodulation in Phaseolus Vulgaris. Plant Physiol. Biochem. 2013, 68, 81–89. [Google Scholar] [CrossRef]
- Dixon, R.A.; Ferreira, D. Genistein. Phytochemistry 2002, 60, 205–211. [Google Scholar] [CrossRef]
- Manas, E.S.; Xu, Z.B.; Unwalla, R.J.; Somers, W.S. Understanding the Selectivity of Genistein for Human Estrogen Receptor-β Using X-Ray Crystallography and Computational Methods. Structure 2004, 12, 2197–2207. [Google Scholar] [CrossRef]
- Dang, Z.-C.; Audinot, V.; Papapoulos, S.E.; Boutin, J.A.; Löwik, C.W.G.M. Peroxisome Proliferator-Activated Receptor γ (PPARγ) as a Molecular Target for the Soy Phytoestrogen Genistein. J. Biol. Chem. 2003, 278, 962–967. [Google Scholar] [CrossRef]
- Liang, F.; Cao, W.; Huang, Y.; Fang, Y.; Cheng, Y.; Pan, S.; Xu, X. Isoflavone Biochanin A, a Novel Nuclear Factor Erythroid 2-Related Factor 2 (Nrf2)-Antioxidant Response Element Activator, Protects against Oxidative Damage in HepG2 Cells. BioFactors 2019, 45, 563–574. [Google Scholar] [CrossRef]
- Suraweera, T.L.; Rupasinghe, H.P.V.; Dellaire, G.; Xu, Z. Regulation of Nrf2/ARE Pathway by Dietary Flavonoids: A Friend or Foe for Cancer Management? Antioxidants 2020, 9, 973. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef]
- Tapas, A.; Sakarkar, D.; Kakde, R. A Review of Flavonoids as Nutraceuticals. Trop. J. Pharm. Res. 2008, 7, 1089–1099. [Google Scholar] [CrossRef]
- Holland, T.M.; Agarwal, P.; Wang, Y.; Leurgans, S.E.; Bennett, D.A.; Booth, S.L.; Morris, M.C. Dietary Flavonols and Risk of Alzheimer Dementia. Neurology 2020, 94, e1749. [Google Scholar] [CrossRef]
- Makris, D.P.; Kallithraka, S.; Kefalas, P. Flavonols in Grapes, Grape Products and Wines: Burden, Profile and Influential Parameters. J. Food Compos. Anal. 2006, 19, 396–404. [Google Scholar] [CrossRef]
- Singh, M.; Kaur, M.; Silakari, O. Flavones: An Important Scaffold for Medicinal Chemistry. Eur. J. Med. Chem. 2014, 84, 206–239. [Google Scholar] [CrossRef]
- Lopez-Lazaro, M. Distribution and Biological Activities of the Flavonoid Luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Samy, R.P.; Gopalakrishnakone, P.; Ignacimuthu, S. Anti-Tumor Promoting Potential of Luteolin against 7,12-Dimethylbenz(a)Anthracene-Induced Mammary Tumors in Rats. Chem. -Biol. Interact. 2006, 164, 1–14. [Google Scholar] [CrossRef]
- Ahmed, S.; Khan, H.; Fratantonio, D.; Hasan, M.M.; Sharifi, S.; Fathi, N.; Ullah, H.; Rastrelli, L. Apoptosis Induced by Luteolin in Breast Cancer: Mechanistic and Therapeutic Perspectives. Phytomedicine 2019, 59, 152883. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Braidy, N.; Gortzi, O.; Sobarzo-Sanchez, E.; Daglia, M.; Skalicka-Woźniak, K.; Nabavi, S.M. Luteolin as an Anti-Inflammatory and Neuroprotective Agent: A Brief Review. Brain Res. Bull. 2015, 119, 1–11. [Google Scholar] [CrossRef]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. IJMS 2019, 20, 1305. [Google Scholar] [CrossRef]
- Shankar, E.; Goel, A.; Gupta, K.; Gupta, S. Plant Flavone Apigenin: An Emerging Anticancer Agent. Curr. Pharmacol. Rep. 2017, 3, 423–446. [Google Scholar] [CrossRef]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in Cancer Therapy: Anti-Cancer Effects and Mechanisms of Action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. Aoac. Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef]
- Khan, A.; Ikram, M.; Hahm, J.R.; Kim, M.O. Antioxidant and Anti-Inflammatory Effects of Citrus Flavonoid Hesperetin: Special Focus on Neurological Disorders. Antioxidants 2020, 9, 609. [Google Scholar] [CrossRef]
- Cavia-Saiz, M.; Busto, M.D.; Pilar-Izquierdo, M.C.; Ortega, N.; Perez-Mateos, M.; Muñiz, P. Antioxidant Properties, Radical Scavenging Activity and Biomolecule Protection Capacity of Flavonoid Naringenin and Its Glycoside Naringin: A Comparative Study. J. Sci. Food Agric. 2010, 90, 1238–1244. [Google Scholar] [CrossRef]
- Wadhwa, R.; Paudel, K.R.; Chin, L.H.; Hon, C.M.; Madheswaran, T.; Gupta, G.; Panneerselvam, J.; Lakshmi, T.; Singh, S.K.; Gulati, M.; et al. Anti-Inflammatory and Anticancer Activities of Naringenin-Loaded Liquid Crystalline Nanoparticles in Vitro. J. Food Biochem. 2021, 45, e13572. [Google Scholar] [CrossRef]
- Shi, X.; Luo, X.; Chen, T.; Guo, W.; Liang, C.; Tang, S.; Mo, J. Naringenin Inhibits Migration, Invasion, Induces Apoptosis in Human Lung Cancer Cells and Arrests Tumour Progression in Vitro. J. Cell. Mol. Med. 2021, 25, 2563–2571. [Google Scholar] [CrossRef]
- Patel, K.; Watson, R.R. Chapter 21-Chocolate and Its Component’s Effect on Cardiovascular Disease. In Lifestyle in Heart Health and Disease; Watson, R.R., Zibadi, S., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 255–266. ISBN 978-0-12-811279-3. [Google Scholar]
- Krittanawong, C.; Narasimhan, B.; Wang, Z.; Hahn, J.; Virk, H.U.H.; Farrell, A.M.; Zhang, H.; Tang, W.W. Association between Chocolate Consumption and Risk of Coronary Artery Disease: A Systematic Review and Meta-Analysis. Eur. J. Prev. Cardiol. 2020, 2047487320936787. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.Á.; Goya, L.; Ramos, S. Antidiabetic Actions of Cocoa Flavanols. Mol. Nutr. Food Res. 2016, 60, 1756–1769. [Google Scholar] [CrossRef]
- Varela-Castillo, O.; Cordero, P.; Gutiérrez-Iglesias, G.; Palma, I.; Rubio-Gayosso, I.; Meaney, E.; Ramirez-Sanchez, I.; Villarreal, F.; Ceballos, G.; Nájera, N. Characterization of the Cytotoxic Effects of the Combination of Cisplatin and Flavanol (-)-Epicatechin on Human Lung Cancer Cell Line A549. An Isobolographic Approach. Exp. Oncol. 2018, 40, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.-Y.; Sang, L.-X.; Jiang, M. Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease. Molecules 2017, 22, 484. [Google Scholar] [CrossRef]
- Yonekura, L.; Martins, C.A.; Sampaio, G.R.; Monteiro, M.P.; César, L.A.M.; Mioto, B.M.; Mori, C.S.; Mendes, T.M.N.; Ribeiro, M.L.; Arçari, D.P. Bioavailability of Catechins from Guaraná (Paullinia Cupana) and Its Effect on Antioxidant Enzymes and Other Oxidative Stress Markers in Healthy Human Subjects. Food Funct. 2016, 7, 2970–2978. [Google Scholar] [CrossRef]
- Iacobucci, G.A.; Sweeny, J.G. The Chemistry of Anthocyanins, Anthocyanidins and Related Flavylium Salts. Tetrahedron 1983, 39, 3005–3038. [Google Scholar] [CrossRef]
- Tuladhar, P.; Sasidharan, S.; Saudagar, P. 17 - Role of Phenols and Polyphenols in Plant Defense Response to Biotic and Abiotic Stresses. In Biocontrol Agents and Secondary Metabolites; Jogaiah, S., Ed.; Woodhead Publishing: Sawston, United Kingdom, 2021; pp. 419–441. ISBN 978-0-12-822919-4. [Google Scholar]
- Teng, H.; Mi, Y.; Cao, H.; Chen, L. Enzymatic Acylation of Raspberry Anthocyanin: Evaluations on Its Stability and Oxidative Stress Prevention. Food Chem. 2022, 372, 130766. [Google Scholar] [CrossRef] [PubMed]
- Laganà, G.; Barreca, D.; Smeriglio, A.; Germanò, M.P.; D’Angelo, V.; Calderaro, A.; Bellocco, E.; Trombetta, D. Evaluation of Anthocyanin Profile, Antioxidant, Cytoprotective, and Anti-Angiogenic Properties of Callistemon Citrinus Flowers. Plants 2020, 9, 1045. [Google Scholar] [CrossRef]
- Si, X.; Bi, J.; Chen, Q.; Cui, H.; Bao, Y.; Tian, J.; Shu, C.; Wang, Y.; Tan, H.; Zhang, W. Effect of Blueberry Anthocyanin-Rich Extracts on Peripheral and Hippocampal Antioxidant Defensiveness: The Analysis of the Serum Fatty Acid Species and Gut Microbiota Profile. J. Agric. Food Chem. 2021, 69, 3658–3666. [Google Scholar] [CrossRef]
- Faria, A.; Mateus, N.; Calhau, C. Flavonoid Transport across Blood-Brain Barrier: Implication for Their Direct Neuroprotective Actions. Nutr. Aging 2012, 1, 89–97. [Google Scholar] [CrossRef]
- Daneman, R.; Prat, A. The Blood-Brain Barrier. Cold Spring Harb. Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef] [PubMed]
- Milbury, P.E.; Kalt, W. Xenobiotic Metabolism and Berry Flavonoid Transport across the Blood-Brain Barrier. J. Agric. Food Chem. 2010, 58, 3950–3956. [Google Scholar] [CrossRef] [PubMed]
- Carbonel, A.A.F.; Cecyn, M.N.; Girão, J.H.R.C.; da Silva Sasso, G.R.; de Mello Ponteciano, B.; Vellozo, E.P.; Simões, R.S.; de Jesus Simões, M.; Girão, M.J.B.C.; de Oliveira, D.R. Flavonoids as Modulators of Synaptic Plasticity: Implications for the Development of Novel Therapeutic Strategies for Healthy Lifestyle. In Flavonoids—A Coloring Model for Cheering up Life; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar]
- Babcock, K.R.; Page, J.S.; Fallon, J.R.; Webb, A.E. Adult Hippocampal Neurogenesis in Aging and Alzheimer’s Disease. Stem Cell Rep. 2021, 16, 681–693. [Google Scholar] [CrossRef] [PubMed]
- King, R.A.; Bursill, D.B. Plasma and Urinary Kinetics of the Isoflavones Daidzein and Genistein after a Single Soy Meal in Humans. Am. J. Clin. Nutr. 1998, 67, 867–872. [Google Scholar] [CrossRef]
- Hollman, P.C.; van Trijp, J.M.; Buysman, M.N.; van der Gaag, M.S.; Mengelers, M.J.; de Vries, J.H.; Katan, M.B. Relative Bioavailability of the Antioxidant Flavonoid Quercetin from Various Foods in Man. FEBS Lett. 1997, 418, 152–156. [Google Scholar] [CrossRef]
- McAnlis, G.T.; McEneny, J.; Pearce, J.; Young, I.S. Absorption and Antioxidant Effects of Quercetin from Onions, in Man. Eur. J. Clin. Nutr. 1999, 53, 92–96. [Google Scholar] [CrossRef]
- Hollman, P.C.; Bijsman, M.N.; van Gameren, Y.; Cnossen, E.P.; de Vries, J.H.; Katan, M.B. The Sugar Moiety Is a Major Determinant of the Absorption of Dietary Flavonoid Glycosides in Man. Free Radic. Res. 1999, 31, 569–573. [Google Scholar] [CrossRef]
- Graefe, E.U.; Wittig, J.; Mueller, S.; Riethling, A.K.; Uehleke, B.; Drewelow, B.; Pforte, H.; Jacobasch, G.; Derendorf, H.; Veit, M. Pharmacokinetics and Bioavailability of Quercetin Glycosides in Humans. J. Clin. Pharm. 2001, 41, 492–499. [Google Scholar] [CrossRef]
- Manach, C.; Morand, C.; Gil-Izquierdo, A.; Bouteloup-Demange, C.; Rémésy, C. Bioavailability in Humans of the Flavanones Hesperidin and Narirutin after the Ingestion of Two Doses of Orange Juice. Eur. J. Clin. Nutr. 2003, 57, 235–242. [Google Scholar] [CrossRef]
- Erlund, I.; Meririnne, E.; Alfthan, G.; Aro, A. Plasma Kinetics and Urinary Excretion of the Flavanones Naringenin and Hesperetin in Humans after Ingestion of Orange Juice and Grapefruit Juice. J. Nutr. 2001, 131, 235–241. [Google Scholar] [CrossRef]
- Bugianesi, R.; Catasta, G.; Spigno, P.; D’Uva, A.; Maiani, G. Naringenin from Cooked Tomato Paste Is Bioavailable in Men. J. Nutr. 2002, 132, 3349–3352. [Google Scholar] [CrossRef] [PubMed]
- Fuhr, U.; Kummert, A.L. The Fate of Naringin in Humans: A Key to Grapefruit Juice-Drug Interactions? Clin. Pharm. 1995, 58, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and Bioefficacy of Polyphenols in Humans. I. Review of 97 Bioavailability Studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosi, N.; Cozzolino, M.; Apolloni, S. The Contribution of Non-Neuronal Cells in Neurodegeneration: From Molecular Pathogenesis to Therapeutic Challenges. Cells 2022, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, V.; Sharma, S. Role of Mitochondrial Dysfunction, Oxidative Stress and Autophagy in Progression of Alzheimer’s Disease. J. Neurol. Sci. 2021, 421, 117253. [Google Scholar] [CrossRef]
- Sharma, C.; Kim, S.R. Linking Oxidative Stress and Proteinopathy in Alzheimer’s Disease. Antioxidants 2021, 10, 1231. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, X.; Wu, T.; Zhang, W.; Shu, J.; He, Y.; Tang, S.-J. Quercetin Attenuates AZT-Induced Neuroinflammation in the CNS. Scientific Reports 2018, 8, 6194. [Google Scholar] [CrossRef]
- Zeng, Y.; Song, J.; Zhang, M.; Wang, H.; Zhang, Y.; Suo, H. Comparison of In Vitro and In Vivo Antioxidant Activities of Six Flavonoids with Similar Structures. Antioxidants 2020, 9, 732. [Google Scholar] [CrossRef]
- Anusha, C.; Sumathi, T.; Joseph, L.D. Protective role of apigenin on rotenone induced rat model of Parkinson’s disease: Suppression of neuroinflammation and oxidative stress mediated apoptosis. Chem. Interactions 2017, 269, 67–79. [Google Scholar] [CrossRef]
- Siddiqui, T.; Bhattarai, P.; Popova, S.; Cosacak, M.I.; Sariya, S.; Zhang, Y.; Mayeux, R.; Tosto, G.; Kizil, C. KYNA/Ahr Signaling Suppresses Neural Stem Cell Plasticity and Neurogenesis in Adult Zebrafish Model of Alzheimer’s Disease. Cells 2021, 10, 2748. [Google Scholar] [CrossRef]
- Zhu, C.; Dong, Y.; Liu, H.; Ren, H.; Cui, Z. Hesperetin Protects against H2O2-Triggered Oxidative Damage via Upregulation of the Keap1-Nrf2/HO-1 Signal Pathway in ARPE-19 Cells. Biomedicine & Pharmacotherapy 2017, 88, 124–133. [Google Scholar]
- Vauzour, D.; Rendeiro, C.; D’amato, A.; Waffo-Téguo, P.; Richard, T.; Mérillon, J.M.; Pontifex, M.G.; Connell, E.; Müller, M.; Butler, L.T. Anthocyanins Promote Learning through Modulation of Synaptic Plasticity Related Proteins in an Animal Model of Ageing. Antioxidants 2021, 10, 1235. [Google Scholar] [CrossRef] [PubMed]
- Maher, P.; Akaishi, T.; Abe, K. Flavonoid fisetin promotes ERK-dependent long-term potentiation and enhances memory. Proc. Natl. Acad. Sci. 2006, 103, 16568–16573. [Google Scholar] [CrossRef] [PubMed]
- Sawamoto, A.; Okuyama, S.; Nakajima, M.; Furukawa, Y. Citrus Flavonoid 3, 5, 6, 7, 8, 3′, 4′-Heptamethoxyflavone Induces BDNF via CAMP/ERK/CREB Signaling and Reduces Phosphodiesterase Activity in C6 Cells. Pharmacol Rep. 2019, 71, 653–658. [Google Scholar] [CrossRef]
- Jia, Z.; Yang, J.; Cao, Z.; Zhao, J.; Zhang, J.; Lu, Y.; Chu, L.; Zhang, S.; Chen, Y.; Pei, L. Baicalin ameliorates chronic unpredictable mild stress-induced depression through the BDNF/ERK/CREB signaling pathway. Behav. Brain Res. 2021, 414, 113463. [Google Scholar] [CrossRef]
- Messaoudi, M.; Bisson, J.-F.; Nejdi, A.; Rozan, P.; Javelot, H. Antidepressant-like Effects of a Cocoa Polyphenolic Extract in Wistar–Unilever Rats. Nutr. Neurosci. 2008, 11, 269–276. [Google Scholar] [CrossRef]
- Yang, X.; Song, D.; Chen, L.; Xiao, H.; Ma, X.; Jiang, Q.; Cheng, O. Curcumin Promotes Neurogenesis of Hippocampal Dentate Gyrus via Wnt/β-Catenin Signal Pathway Following Cerebral Ischemia in Mice. Brain Res. 2021, 1751, 147197. [Google Scholar] [CrossRef]
- Meyer, E.; Mori, M.A.; Campos, A.C.; Andreatini, R.; Guimarães, F.S.; Milani, H.; de Oliveira, R.M.W. Myricitrin Induces Antidepressant-like Effects and Facilitates Adult Neurogenesis in Mice. Behav. Brain Res. 2017, 316, 59–65. [Google Scholar] [CrossRef]
- Jiang, T.; Xu, S.; Shen, Y.; Xu, Y.; Li, Y. Genistein Attenuates Isoflurane-Induced Neuroinflammation by Inhibiting TLR4-Mediated Microglial-Polarization in vivo and in vitro. J. Inflamm. Res. 2021, 14, 2587–2600. [Google Scholar] [CrossRef]
- Yang, Y.; Tan, X.; Xu, J.; Wang, T.; Liang, T.; Xu, X.; Ma, C.; Xu, Z.; Wang, W.; Li, H.; et al. Luteolin Alleviates Neuroinflammation via Downregulating the TLR4/TRAF6/NF-ΚB Pathway after Intracerebral Hemorrhage. Biomed. Pharmacother 2020, 126, 110044. [Google Scholar] [CrossRef]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms Underlying Inflammation in Neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef] [PubMed]
- Sonninen, T.-M.; Goldsteins, G.; Laham-Karam, N.; Koistinaho, J.; Lehtonen, Š. Proteostasis Disturbances and Inflammation in Neurodegenerative Diseases. Cells 2020, 9, 2183. [Google Scholar] [CrossRef]
- Rajendran, L.; Paolicelli, R.C. Microglia-Mediated Synapse Loss in Alzheimer’s Disease. J. Neurosci. 2018, 38, 2911–2919. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-Y.; Tan, M.-S.; Yu, J.-T.; Tan, L. Role of Pro-Inflammatory Cytokines Released from Microglia in Alzheimer’s Disease. Ann. Transl. Med. 2015, 3, 136. [Google Scholar] [CrossRef]
- Di Benedetto, G.; Burgaletto, C.; Bellanca, C.M.; Munafò, A.; Bernardini, R.; Cantarella, G. Role of Microglia and Astrocytes in Alzheimer’s Disease: From Neuroinflammation to Ca2+ Homeostasis Dysregulation. Cells 2022, 11, 2728. [Google Scholar] [CrossRef]
- More, S.V.; Kumar, H.; Kim, I.S.; Song, S.-Y.; Choi, D.-K. Cellular and Molecular Mediators of Neuroinflammation in the Pathogenesis of Parkinson’s Disease. Mediat. Inflamm. 2013, 2013, 952375. [Google Scholar] [CrossRef]
- Du, Y.; Brennan, F.H.; Popovich, P.G.; Zhou, M. Microglia Maintain the Normal Structure and Function of the Hippocampal Astrocyte Network. Glia 2022, 70, 1359–1379. [Google Scholar] [CrossRef]
- Torres-Ceja, B.; Olsen, M.L. A Closer Look at Astrocyte Morphology: Development, Heterogeneity, and Plasticity at Astrocyte Leaflets. Curr. Opin. Neurobiol. 2022, 74, 102550. [Google Scholar] [CrossRef]
- Lee, H.-G.; Wheeler, M.A.; Quintana, F.J. Function and Therapeutic Value of Astrocytes in Neurological Diseases. Nat. Rev. Drug Discov. 2022, 21, 339–358. [Google Scholar] [CrossRef] [PubMed]
- Kozuka, N.; Itofusa, R.; Kudo, Y.; Morita, M. Lipopolysaccharide and Proinflammatory Cytokines Require Different Astrocyte States to Induce Nitric Oxide Production. J. Neurosci. Res. 2005, 82, 717–728. [Google Scholar] [CrossRef]
- Sekiyama, K.; Sugama, S.; Fujita, M.; Sekigawa, A.; Takamatsu, Y.; Waragai, M.; Takenouchi, T.; Hashimoto, M. Neuroinflammation in Parkinson’s Disease and Related Disorders: A Lesson from Genetically Manipulated Mouse Models of -Synucleinopathies. Park. Dis. 2012, 2012, 271732. [Google Scholar] [CrossRef]
- Lee, H.-J.; Suk, J.-E.; Patrick, C.; Bae, E.-J.; Cho, J.-H.; Rho, S.; Hwang, D.; Masliah, E.; Lee, S.-J. Direct Transfer of α-Synuclein from Neuron to Astroglia Causes Inflammatory Responses in Synucleinopathies. J. Biol. Chem. 2010, 285, 9262–9272. [Google Scholar] [CrossRef]
- Hirsch, E.C.; Hunot, S. Neuroinflammation in Parkinson’s Disease: A Target for Neuroprotection? Lancet Neurol. 2009, 8, 382–397. [Google Scholar] [CrossRef]
- Kempuraj, D.; Thangavel, R.; Natteru, P.; Selvakumar, G.; Saeed, D.; Zahoor, H.; Zaheer, S.; Iyer, S.; Zaheer, A. Neuroinflammation Induces Neurodegeneration. J. Neurol. Neurosurg. Spine 2016, 1, 1003. [Google Scholar]
- Chen, W.; Zhang, X.; Huang, W. Role of Neuroinflammation in Neurodegenerative Diseases. Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef]
- Ellwardt, E.; Zipp, F. Molecular Mechanisms Linking Neuroinflammation and Neurodegeneration in MS. Exp. Neurol. 2014, 262, 8–17. [Google Scholar] [CrossRef]
- Gelders, G.; Baekelandt, V.; Van der Perren, A. Linking Neuroinflammation and Neurodegeneration in Parkinson’s Disease. J. Immunol. Res. 2018, 2018, 4784268. [Google Scholar] [CrossRef]
- Whitton, P. Inflammation as a Causative Factor in the Aetiology of Parkinson’s Disease. Br. J. Pharmacol. 2007, 150, 963–976. [Google Scholar] [CrossRef]
- Gerhard, A.; Pavese, N.; Hotton, G.; Turkheimer, F.; Es, M.; Hammers, A.; Eggert, K.; Oertel, W.; Banati, R.B.; Brooks, D.J. In Vivo Imaging of Microglial Activation with [11C](R)-PK11195 PET in Idiopathic Parkinson’s Disease. Neurobiol. Dis. 2006, 21, 404–412. [Google Scholar] [CrossRef]
- Breitzig, M.; Bhimineni, C.; Lockey, R.; Kolliputi, N. 4-Hydroxy-2-Nonenal: A Critical Target in Oxidative Stress? Am J Physiol Cell Physiol. 2016, 311, C537–C543. [Google Scholar] [CrossRef]
- Markesbery, W.R.; Lovell, M.A. Four-Hydroxynonenal, a Product of Lipid Peroxidation, Is Increased in the Brain in Alzheimer’s Disease. Neurobiol. Aging 1998, 19, 33–36. [Google Scholar] [CrossRef]
- Sultana, R.; Boyd-Kimball, D.; Cai, J.; Pierce, W.M.; Klein, J.B.; Merchant, M.; Butterfield, D.A. Proteomics Analysis of the Alzheimer’s Disease Hippocampal Proteome. J. Alzheimers Dis. 2007, 11, 153–164. [Google Scholar] [CrossRef]
- Sultana, R.; Perluigi, M.; Butterfield, D.A. Lipid Peroxidation Triggers Neurodegeneration: A Redox Proteomics View into the Alzheimer Disease Brain. Free Radic. Biol. Med. 2013, 62, 157–169. [Google Scholar] [CrossRef]
- Borowiec, K.; Michalak, A. Flavonoids from Edible Fruits as Therapeutic Agents in Neuroinflammation–a Comprehensive Review and Update. Crit. Rev. Food Sci. Nutr. 2021, 62, 6742–6760. [Google Scholar] [CrossRef]
- Bayat, P.; Farshchi, M.; Yousefian, M.; Mahmoudi, M.; Yazdian-Robati, R. Flavonoids, the Compounds with Anti-Inflammatory and Immunomodulatory Properties, as Promising Tools in Multiple Sclerosis (MS) Therapy: A Systematic Review of Preclinical Evidence. Int. Immunopharmacol. 2021, 95, 107562. [Google Scholar] [CrossRef]
- Owona, B.A.; Abia, W.A.; Moundipa, P.F. Natural Compounds Flavonoids as Modulators of Inflammasomes in Chronic Diseases. Int. Immunopharmacol. 2020, 84, 106498. [Google Scholar] [CrossRef]
- Yousefian, M.; Shakour, N.; Hosseinzadeh, H.; Hayes, A.W.; Hadizadeh, F.; Karimi, G. The Natural Phenolic Compounds as Modulators of NADPH Oxidases in Hypertension. Phytomedicine 2019, 55, 200–213. [Google Scholar] [CrossRef]
- González Mosquera, D.; Hernández Ortega, Y.; Fernández, P.; González, Y.; Doens, D.; Vander Heyden, Y.; Foubert, K.; Pieters, L. Flavonoids from Boldoa Purpurascens Inhibit Proinflammatory Cytokines (TNF-α and IL-6) and the Expression of COX-2. Phytother. Res. 2018, 32, 1750–1754. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Y.; Cheng, X.; Qiao, X. The Alleviative Effects of Quercetin on Cadmium-Induced Necroptosis via Inhibition ROS/INOS/NF-ΚB Pathway in the Chicken Brain. Biol. Trace Elem. Res. 2021, 199, 1584–1594. [Google Scholar] [CrossRef]
- Brown, C.M.; Mulcahey, T.A.; Filipek, N.C.; Wise, P.M. Production of Proinflammatory Cytokines and Chemokines during Neuroinflammation: Novel Roles for Estrogen Receptors α and β. Endocrinology 2010, 151, 4916–4925. [Google Scholar] [CrossRef]
- Vegeto, E.; Belcredito, S.; Ghisletti, S.; Meda, C.; Etteri, S.; Maggi, A. The Endogenous Estrogen Status Regulates Microglia Reactivity in Animal Models of Neuroinflammation. Endocrinology 2006, 147, 2263–2272. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, M.; Haque, A.; Banik, N.L.; Nagarkatti, P.; Nagarkatti, M.; Ray, S.K. Estrogen Receptor Agonists for Attenuation of Neuroinflammation and Neurodegeneration. Brain Res. Bull. 2014, 109, 22–31. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed]
- Shibata, N.; Kobayashi, M. The role for oxidative stress in neurodegenerative diseases. Brain Nerve 2008, 60, 157–170. [Google Scholar] [PubMed]
- Golden, T.R.; Hinerfeld, D.A.; Melov, S. Oxidative Stress and Aging: Beyond Correlation. Aging Cell 2002, 1, 117–123. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial Dysfunction and Oxidative Stress in Neurodegenerative Diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Kehrer, J.P. The Haber–Weiss Reaction and Mechanisms of Toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of Apoptosis Signalling Pathways by Reactive Oxygen Species. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Rego, A.C.; Oliveira, C.R. Mitochondrial Dysfunction and Reactive Oxygen Species in Excitotoxicity and Apoptosis: Implications for the Pathogenesis of Neurodegenerative Diseases. Neurochem. Res. 2003, 28, 1563–1574. [Google Scholar] [CrossRef]
- Tamás, M.J.; Sharma, S.K.; Ibstedt, S.; Jacobson, T.; Christen, P. Heavy Metals and Metalloids as a Cause for Protein Misfolding and Aggregation. Biomolecules 2014, 4, 252–267. [Google Scholar] [CrossRef]
- Devi, S.; Chaturvedi, M.; Fatima, S.; Priya, S. Environmental Factors Modulating Protein Conformations and Their Role in Protein Aggregation Diseases. Toxicology 2022, 465, 153049. [Google Scholar] [CrossRef] [PubMed]
- Chaves, R.S.; Melo, T.Q.; Martins, S.A.; Ferrari, M.F. Protein Aggregation Containing Beta-Amyloid, Alpha-Synuclein and Hyperphosphorylated Tau in Cultured Cells of Hippocampus, Substantia Nigra and Locus Coeruleus after Rotenone Exposure. BMC Neurosci. 2010, 11, 144. [Google Scholar] [CrossRef] [PubMed]
- Husain, S.R.; Cillard, J.; Cillard, P. Hydroxyl Radical Scavenging Activity of Flavonoids. Phytochemistry 1987, 26, 2489–2491. [Google Scholar] [CrossRef]
- Tian, Y.; Wen, Z.; Lei, L.; Li, F.; Zhao, J.; Zhi, Q.; Li, F.; Yin, R.; Ming, J. Coreopsis Tinctoria Flowers Extract Ameliorates D-Galactose Induced Aging in Mice via Regulation of Sirt1-Nrf2 Signaling Pathway. J. Funct. Foods 2019, 60, 103464. [Google Scholar] [CrossRef]
- Cichon, N.; Saluk-Bijak, J.; Gorniak, L.; Przyslo, L.; Bijak, M. Flavonoids as a Natural Enhancer of Neuroplasticity-An Overview of the Mechanism of Neurorestorative Action. Antioxidants 2020, 9, 1035. [Google Scholar] [CrossRef]
- Sampaio-Baptista, C.; Khrapitchev, A.A.; Foxley, S.; Schlagheck, T.; Scholz, J.; Jbabdi, S.; DeLuca, G.C.; Miller, K.L.; Taylor, A.; Thomas, N.; et al. Motor Skill Learning Induces Changes in White Matter Microstructure and Myelination. J. Neurosci. 2013, 33, 19499–19503. [Google Scholar] [CrossRef]
- Sampaio-Baptista, C.; Johansen-Berg, H. White Matter Plasticity in the Adult Brain. Neuron 2017, 96, 1239–1251. [Google Scholar] [CrossRef]
- Irmen, F.; Karabanov, A.N.; Bögemann, S.A.; Andersen, K.W.; Madsen, K.H.; Bisgaard, T.; Dyrby, T.B.; Siebner, H.R. Functional and Structural Plasticity Co-Express in a Left Premotor Region During Early Bimanual Skill Learning. Front. Hum. Neurosci. 2020, 14, 310. [Google Scholar] [CrossRef]
- Bahr-Hosseini, M.; Saver, J.L. Mechanisms of Action of Acute and Subacute Sphenopalatine Ganglion Stimulation for Ischemic Stroke. Int. J. Stroke 2020, 15, 839–848. [Google Scholar] [CrossRef]
- Kempermann, G.; Krebs, J.; Fabel, K. The Contribution of Failing Adult Hippocampal Neurogenesis to Psychiatric Disorders. Curr. Opin. Psychiatry 2008, 21, 290–295. [Google Scholar] [CrossRef]
- Dranovsky, A.; Hen, R. Hippocampal Neurogenesis: Regulation by Stress and Antidepressants. Biol. Psychiatry 2006, 59, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The Role of Adult Hippocampal Neurogenesis in Brain Health and Disease. Mol. Psychiatry 2019, 24, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.J.; Umemori, H. The Best-Laid Plans Go Oft Awry: Synaptogenic Growth Factor Signaling in Neuropsychiatric Disease. Front. Synaptic Neurosci. 2014, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Meeker, R.B.; Williams, K.S. The P75 Neurotrophin Receptor: At the Crossroad of Neural Repair and Death. Neural Regen. Res. 2015, 10, 721–725. [Google Scholar] [CrossRef]
- Thomaz, A.; Jaeger, M.; Brunetto, A.L.; Brunetto, A.T.; Gregianin, L.; de Farias, C.B.; Ramaswamy, V.; Nör, C.; Taylor, M.D.; Roesler, R. Neurotrophin Signaling in Medulloblastoma. Cancers 2020, 12, 2542. [Google Scholar] [CrossRef]
- Bruel-Jungerman, E.; Veyrac, A.; Dufour, F.; Horwood, J.; Laroche, S.; Davis, S. Inhibition of PI3K-Akt Signaling Blocks Exercise-Mediated Enhancement of Adult Neurogenesis and Synaptic Plasticity in the Dentate Gyrus. PloS One 2009, 4, e7901. [Google Scholar]
- Hettige, N.C.; Ernst, C. FOXG1 Dose in Brain Development. Front. Pediatr. 2019, 7, 482. [Google Scholar] [CrossRef]
- Chiola, S.; Do, M.D.; Centrone, L.; Mallamaci, A. Foxg1 Overexpression in Neocortical Pyramids Stimulates Dendrite Elongation Via Hes1 and PCreb1 Upregulation. Cerebral Cortex 2019, 29, 1006–1019. [Google Scholar] [CrossRef]
- Brivio, P.; Sbrini, G.; Corsini, G.; Paladini, M.S.; Racagni, G.; Molteni, R.; Calabrese, F. Chronic Restraint Stress Inhibits the Response to a Second Hit in Adult Male Rats: A Role for BDNF Signaling. Int. J. Mol. Sci. 2020, 21, 6261. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef]
- Letenneur, L.; Proust-Lima, C.; Le Gouge, A.; Dartigues, J.; Barberger-Gateau, P. Flavonoid Intake and Cognitive Decline over a 10-Year Period. Am. J. Epidemiol. 2007, 165, 1364–1371. [Google Scholar] [CrossRef] [PubMed]
- Alberini, C.M. Transcription Factors in Synaptic Plasticity and Learning and Memory. In Encyclopedia of Neuroscience; Squire, L.R., Ed.; Academic Press: Oxford, UK, 2009; pp. 1081–1092. ISBN 978-0-08-045046-9. [Google Scholar]
- Spencer, J.P.E. Flavonoids and Brain Health: Multiple Effects Underpinned by Common Mechanisms. Genes Nutr. 2009, 4, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Vafeiadou, K.; Rice-Evans, C.; Williams, R.J.; Spencer, J.P. Activation of Pro-survival Akt and ERK1/2 Signalling Pathways Underlie the Anti-apoptotic Effects of Flavanones in Cortical Neurons. J. Neurochem. 2007, 103, 1355–1367. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, Q.; Yu, Q. Quercetin Enrich Diet during the Early-Middle Not Middle-Late Stage of Alzheimer’s Disease Ameliorates Cognitive Dysfunction. Am. J. Transl. Res. 2018, 10, 1237. [Google Scholar] [PubMed]
- Yu, J.-J.; Pei, L.-B.; Zhang, Y.; Wen, Z.-Y.; Yang, J.-L. Chronic Supplementation of Curcumin Enhances the Efficacy of Antidepressants in Major Depressive Disorder: A Randomized, Double-Blind, Placebo-Controlled Pilot Study. J. Clin. Psychopharmacol. 2015, 35, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Valente, T.; Hidalgo, J.; Bolea, I.; Ramirez, B.; Anglés, N.; Reguant, J.; Morelló, J.R.; Gutiérrez, C.; Boada, M.; Unzeta, M. A Diet Enriched in Polyphenols and Polyunsaturated Fatty Acids, LMN Diet, Induces Neurogenesis in the Subventricular Zone and Hippocampus of Adult Mouse Brain. J. Alzheimer’s Dis. 2009, 18, 849–865. [Google Scholar] [CrossRef]
- Fernández-Fernández, L.; Comes, G.; Bolea, I.; Valente, T.; Ruiz, J.; Murtra, P.; Ramirez, B.; Anglés, N.; Reguant, J.; Morelló, J.R. LMN Diet, Rich in Polyphenols and Polyunsaturated Fatty Acids, Improves Mouse Cognitive Decline Associated with Aging and Alzheimer’s Disease. Behav. Brain Res. 2012, 228, 261–271. [Google Scholar] [CrossRef]
- Ma, Z.-X.; Zhang, R.-Y.; Rui, W.-J.; Wang, Z.-Q.; Feng, X. Quercetin Alleviates Chronic Unpredictable Mild Stress-Induced Depressive-like Behaviors by Promoting Adult Hippocampal Neurogenesis via FoxG1/CREB/BDNF Signaling Pathway. Behav. Brain Res. 2021, 406, 113245. [Google Scholar] [CrossRef]
- Baba, M.; Nakajo, S.; Tu, P.-H.; Tomita, T.; Nakaya, K.; Lee, V.; Trojanowski, J.Q.; Iwatsubo, T. Aggregation of Alpha-Synuclein in Lewy Bodies of Sporadic Parkinson’s Disease and Dementia with Lewy Bodies. Am. J. Pathol. 1998, 152, 879. [Google Scholar]
- Diao, J.; Burré, J.; Vivona, S.; Cipriano, D.J.; Sharma, M.; Kyoung, M.; Südhof, T.C.; Brunger, A.T. Native α-Synuclein Induces Clustering of Synaptic-Vesicle Mimics via Binding to Phospholipids and Synaptobrevin-2/VAMP2. elife 2013, 2, e00592. [Google Scholar] [CrossRef]
- Jensen, P.H.; Nielsen, M.S.; Jakes, R.; Dotti, C.G.; Goedert, M. Binding of α-Synuclein to Brain Vesicles Is Abolished by Familial Parkinson’s Disease Mutation. J. Biol. Chem. 1998, 273, 26292–26294. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Schlachetzki, J.C.M.; Helling, S.; Bussmann, J.C.; Berlinghof, M.; Schäffer, T.E.; Marcus, K.; Winkler, J.; Klucken, J.; Becker, C.-M. Oxidative Stress-Induced Posttranslational Modifications of Alpha-Synuclein: Specific Modification of Alpha-Synuclein by 4-Hydroxy-2-Nonenal Increases Dopaminergic Toxicity. Mol. Cell. Neurosci. 2013, 54, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Selkoe, D.J. The Amyloid Hypothesis of Alzheimer’s Disease: Progress and Problems on the Road to Therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Tsuji, M. Protofibrils of Amyloid-β Are Important Targets of a Disease-Modifying Approach for Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 952. [Google Scholar] [CrossRef] [PubMed]
- Koffie, R.M.; Meyer-Luehmann, M.; Hashimoto, T.; Adams, K.W.; Mielke, M.L.; Garcia-Alloza, M.; Micheva, K.D.; Smith, S.J.; Kim, M.L.; Lee, V.M. Oligomeric Amyloid β Associates with Postsynaptic Densities and Correlates with Excitatory Synapse Loss near Senile Plaques. Proc. Natl. Acad. Sci. USA 2009, 106, 4012–4017. [Google Scholar] [CrossRef]
- Wei, W.; Nguyen, L.N.; Kessels, H.W.; Hagiwara, H.; Sisodia, S.; Malinow, R. Amyloid Beta from Axons and Dendrites Reduces Local Spine Number and Plasticity. Nat. Neurosci. 2010, 13, 190–196. [Google Scholar] [CrossRef]
- Walsh, D.M.; Klyubin, I.; Fadeeva, J.V.; Cullen, W.K.; Anwyl, R.; Wolfe, M.S.; Rowan, M.J.; Selkoe, D.J. Naturally Secreted Oligomers of Amyloid β Protein Potently Inhibit Hippocampal Long-Term Potentiation in Vivo. Nature 2002, 416, 535–539. [Google Scholar] [CrossRef]
- Joshi, V.; Mishra, R.; Upadhyay, A.; Amanullah, A.; Poluri, K.M.; Singh, S.; Kumar, A.; Mishra, A. Polyphenolic Flavonoid (Myricetin) Upregulated Proteasomal Degradation Mechanisms: Eliminates Neurodegenerative Proteins Aggregation. J. Cell Physiol. 2019, 234, 20900–20914. [Google Scholar] [CrossRef]
- Freyssin, A.; Page, G.; Fauconneau, B.; Rioux Bilan, A. Natural Polyphenols Effects on Protein Aggregates in Alzheimer’s and Parkinson’s Prion-like Diseases. Neural Regen. Res. 2018, 13, 955–961. [Google Scholar] [CrossRef]
- Baul, H.S.; Rajiniraja, M. Favorable Binding of Quercetin to α-Synuclein as Potential Target in Parkinson Disease: An Insilico Approach. Res. J. Pharm. Technol. 2018, 11, 203–206. [Google Scholar] [CrossRef]
- Bieschke, J.; Russ, J.; Friedrich, R.P.; Ehrnhoefer, D.E.; Wobst, H.; Neugebauer, K.; Wanker, E.E. EGCG Remodels Mature α-Synuclein and Amyloid-β Fibrils and Reduces Cellular Toxicity. Proc. Natl. Acad. Sci. USA 2010, 107, 7710–7715. [Google Scholar] [CrossRef] [PubMed]
- Gauci, A.J.; Caruana, M.; Giese, A.; Scerri, C.; Vassallo, N. Identification of Polyphenolic Compounds and Black Tea Extract as Potent Inhibitors of Lipid Membrane Destabilization by Aβ 42 Aggregates. J. Alzheimer’s Dis. 2011, 27, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Li, L.; Takamura, Y.; Yoshiike, Y.; Zhu, L.; Han, F.; Mao, X.; Ikeda, T.; Takasaki, J.; Nishijo, H.; et al. Phenolic Compounds Prevent Amyloid β-Protein Oligomerization and Synaptic Dysfunction by Site-Specific Binding. J. Biol. Chem. 2012, 287, 14631–14643. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.Y.; Kang, S.S.; Lee, S.K.; Han, B.H. Polyphenolic Biflavonoids Inhibit Amyloid-Beta Fibrillation and Disaggregate Preformed Amyloid-Beta Fibrils. Biomol. Ther. 2020, 28, 145. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Rezai-Zadeh, K.; Koyama, N.; Arendash, G.W.; Yamaguchi, H.; Kakuda, N.; Horikoshi-Sakuraba, Y.; Tan, J.; Town, T. Tannic Acid Is a Natural β-Secretase Inhibitor That Prevents Cognitive Impairment and Mitigates Alzheimer-like Pathology in Transgenic Mice. J. Biol. Chem. 2012, 287, 6912–6927. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Zhao, W.; Sang, J.; Wang, W.; Wei, W.; Wang, Y.; Zhao, F.; Lu, F.; Liu, F. Inhibitory Effect of a Flavonoid Dihydromyricetin against Aβ40 Amyloidogenesis and Its Associated Cytotoxicity. ACS Chem. Neurosci. 2019, 10, 4696–4703. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.S.; Mona, M.M.; Elsisy, R.A.; Hantash, E.M. Hesperidin Preserves Cognitive Functions and Hippocampus Histological Architecture in Albino Wistar Rats Subjected to Stress Through Enhancement of Brain-Derived Neurotrophic Factor. Neurotox. Res. 2022, 40, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Caruana, M.; Neuner, J.; Högen, T.; Schmidt, F.; Kamp, F.; Scerri, C.; Giese, A.; Vassallo, N. Polyphenolic Compounds Are Novel Protective Agents against Lipid Membrane Damage by α-Synuclein Aggregates in Vitro. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2012, 1818, 2502–2510. [Google Scholar] [CrossRef] [PubMed]
- Thapa, A.; Chi, E.Y. Biflavonoids as Potential Small Molecule Therapeutics for Alzheimer’s Disease. In Natural Compounds as Therapeutic Agents for Amyloidogenic Diseases; Springer: Cham Switzerland, 2015; pp. 55–77. ISBN 978-3-319-18365-7. [Google Scholar]
- Germanà, A.; Guerrera, M.C.; Laurà, R.; Levanti, M.; Aragona, M.; Mhalhel, K.; Germanà, G.; Montalbano, G.; Abbate, F. Expression and Localization of BDNF/TrkB System in the Zebrafish Inner Ear. Int. J. Mol. Sci. 2020, 21, 5787. [Google Scholar] [CrossRef]
- Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Laurà, R.; Cometa, M.; Levanti, M.; Abbate, F.; Cobo, T.; Capitelli, G.; et al. The BDNF/TrkB Neurotrophin System in the Sensory Organs of Zebrafish. Int. J. Mol. Sci. 2022, 23, 2621. [Google Scholar] [CrossRef]
- Whitfield, T.T. Zebrafish as a Model for Hearing and Deafness. J. Neurobiol. 2002, 53, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Laurà, R.; Levanti, M.; Abbate, F.; Cobo, T.; Capitelli, G.; Calapai, F.; et al. Localization of BDNF and Calretinin in Olfactory Epithelium and Taste Buds of Zebrafish (Danio Rerio). Int. J. Mol. Sci. 2022, 23, 4696. [Google Scholar] [CrossRef] [PubMed]
- Roper, C.; Tanguay, R.L. Chapter 12-Zebrafish as a Model for Developmental Biology and Toxicology. In Handbook of Developmental Neurotoxicology (Second Edition); Slikker, W., Paule, M.G., Wang, C., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 143–151. ISBN 978-0-12-809405-1. [Google Scholar]
- Montalbano, G.; Maugeri, A.; Guerrera, M.C.; Miceli, N.; Navarra, M.; Barreca, D.; Cirmi, S.; Germanà, A. A White Grape Juice Extract Reduces Fat Accumulation through the Modulation of Ghrelin and Leptin Expression in an In Vivo Model of Overfed Zebrafish. Molecules 2021, 26, 1119. [Google Scholar] [CrossRef]
- Abbate, F.; Germana, G.P.; De Carlos, F.; Montalbano, G.; Laurà, R.; Levanti, M.B.; Germana, A. The Oral Cavity of the Adult Zebrafish (Danio Rerio). Anat. Histol. Embryol. 2006, 35, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Montalbano, G.; Mania, M.; Guerrera, M.C.; Laurà, R.; Abbate, F.; Levanti, M.; Maugeri, A.; Germanà, A.; Navarra, M. Effects of a Flavonoid-Rich Extract from Citrus Sinensis Juice on a Diet-Induced Obese Zebrafish. Int. J. Mol. Sci. 2019, 20, 5116. [Google Scholar] [CrossRef]
- Kim, G.-H.J.; Mo, H.; Liu, H.; Okorie, M.; Chen, S.; Zheng, J.; Li, H.; Arkin, M.; Huang, B.; Guo, S. In Vivo Dopamine Neuron Imaging-Based Small Molecule Screen Identifies Novel Neuroprotective Compounds and Targets. Front Pharm. 2022, 13, 837756. [Google Scholar] [CrossRef] [PubMed]
- Yaniv, K.; Isogai, S.; Castranova, D.; Dye, L.; Hitomi, J.; Weinstein, B.M. Live Imaging of Lymphatic Development in the Zebrafish. Nat. Med. 2006, 12, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.L.; Cross, S.J.; Lu, Y.; Kadler, K.E.; Lu, Y.; Dallas, S.L.; Martin, P. Live Imaging of Collagen Deposition during Skin Development and Repair in a Collagen I–GFP Fusion Transgenic Zebrafish Line. Dev. Biol. 2018, 441, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Vicente, M.; Salgado-Almario, J.; Collins, M.M.; Martínez-Sielva, A.; Minoshima, M.; Kikuchi, K.; Domingo, B.; Llopis, J. Cardioluminescence in Transgenic Zebrafish Larvae: A Calcium Imaging Tool to Study Drug Effects and Pathological Modeling. Biomedicines 2021, 9, 1294. [Google Scholar] [CrossRef]
- Markin, P.A.; Brito, A.; Moskaleva, N.E.; Tagliaro, F.; Tarasov, V.V.; La Frano, M.R.; Savitskii, M.V.; Appolonova, S.A. Short- and Medium-Term Exposures of Diazepam Induce Metabolomic Alterations Associated with the Serotonergic, Dopaminergic, Adrenergic and Aspartic Acid Neurotransmitter Systems in Zebrafish (Danio Rerio) Embryos/Larvae. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 38, 100816. [Google Scholar] [CrossRef]
- Ton, C.; Parng, C. The Use of Zebrafish for Assessing Ototoxic and Otoprotective Agents. Hear. Res. 2005, 208, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Bang, P.I.; Sewell, W.F.; Malicki, J.J. Morphology and Cell Type Heterogeneities of the Inner Ear Epithelia in Adult and Juvenile Zebrafish (Danio Rerio). J. Comp. Neurol. 2001, 438, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Tsang, B.; Gerlai, R. Breeding and Larviculture of Zebrafish (Danio Rerio). In Laboratory Fish in Biomedical Research; D’Angelo, L., de Girolamo, P., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 63–80. [Google Scholar]
- Balcombe, J.P.; Barnard, N.D.; Sandusky, C. Laboratory Routines Cause Animal Stress. J. Am. Assoc. Lab. Anim. Sci. 2004, 43, 42–51. [Google Scholar]
- Nasiadka, A.; Clark, M.D. Zebrafish Breeding in the Laboratory Environment. ILAR J. 2012, 53, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Berghmans, S.; Jette, C.; Langenau, D.; Hsu, K.; Stewart, R.; Look, T.; Kanki, J.P. Making Waves in Cancer Research: New Models in the Zebrafish. BioTechniques 2005, 39, 227–237. [Google Scholar] [CrossRef]
- OECD. Test No. 236: Fish Embryo Acute Toxicity (FET) Test 2013. Available online: https://www.oecd-ilibrary.org/environment/test-no-236-fish-embryo-acute-toxicity-fet-test_9789264203709-en (accessed on 29 September 2022).
- Vilella, A.J.; Severin, J.; Ureta-Vidal, A.; Heng, L.; Durbin, R.; Birney, E. EnsemblCompara GeneTrees: Complete, Duplication-Aware Phylogenetic Trees in Vertebrates. Genome Res. 2009, 19, 327–335. [Google Scholar] [CrossRef]
- Golzio, C.; Willer, J.; Talkowski, M.E.; Oh, E.C.; Taniguchi, Y.; Jacquemont, S.; Reymond, A.; Sun, M.; Sawa, A.; Gusella, J.F. KCTD13 Is a Major Driver of Mirrored Neuroanatomical Phenotypes of the 16p11. 2 Copy Number Variant. Nature 2012, 485, 363–367. [Google Scholar] [CrossRef]
- Panizzi, J.R.; Becker-Heck, A.; Castleman, V.H.; Al-Mutairi, D.A.; Liu, Y.; Loges, N.T.; Pathak, N.; Austin-Tse, C.; Sheridan, E.; Schmidts, M. CCDC103 Mutations Cause Primary Ciliary Dyskinesia by Disrupting Assembly of Ciliary Dynein Arms. Nat. Genet. 2012, 44, 714–719. [Google Scholar] [CrossRef]
- Roscioli, T.; Kamsteeg, E.-J.; Buysse, K.; Maystadt, I.; van Reeuwijk, J.; van den Elzen, C.; van Beusekom, E.; Riemersma, M.; Pfundt, R.; Vissers, L.E. Mutations in ISPD Cause Walker-Warburg Syndrome and Defective Glycosylation of α-Dystroglycan. Nat. Genet. 2012, 44, 581–585. [Google Scholar] [CrossRef]
- Leimer, U.; Lun, K.; Romig, H.; Walter, J.; Grünberg, J.; Brand, M.; Haass, C. Zebrafish (Danio Rerio) Presenilin Promotes Aberrant Amyloid β-Peptide Production and Requires a Critical Aspartate Residue for Its Function in Amyloidogenesis. Biochemistry 1999, 38, 13602–13609. [Google Scholar] [CrossRef]
- Groth, C.; Nornes, S.; McCarty, R.; Tamme, R.; Lardelli, M. Identification of a Second Presenilin Gene in Zebrafish with Similarity to the Human Alzheimer’s Disease Presenilin2. Dev. Genes Evol. 2002, 212, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.; McGrath, G.; Zhang, J.; Ruddy, D.A.; Sym, M.; Apfeld, J.; Nicoll, M.; Maxwell, M.; Hai, B.; Ellis, M.C.; et al. Aph-1 and Pen-2 Are Required for Notch Pathway Signaling, γ-Secretase Cleavage of ΒAPP, and Presenilin Protein Accumulation. Dev. Cell 2002, 3, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Musa, A.; Lehrach, H.; Russo, V.E.A. Distinct Expression Patterns of Two Zebrafish Homologues of the Human APP Gene during Embryonic Development. Dev. Genes Evol. 2001, 211, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Moussavi Nik, S.H.; Wilson, L.; Newman, M.; Croft, K.; Mori, T.A.; Musgrave, I.; Lardelli, M. The BACE1-PSEN-AβPP Regulatory Axis Has an Ancient Role in Response to Low Oxygen/Oxidative Stress. J. Alzheimer’s Dis. 2012, 28, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Van Bebber, F.; Hruscha, A.; Willem, M.; Schmid, B.; Haass, C. Loss of Bace2 in Zebrafish Affects Melanocyte Migration and Is Distinct from Bace1 Knock out Phenotypes. J. Neurochem. 2013, 127, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Martins, R.N.; Lardelli, M. Complex Splicing and Neural Expression of Duplicated Tau Genes in Zebrafish Embryos. J. Alzheimer’s Dis. 2009, 18, 305–317. [Google Scholar] [CrossRef]
- Babin, P.J.; Thisse, C.; Durliat, M.; Andre, M.; Akimenko, M.-A.; Thisse, B. Both Apolipoprotein E and A-I Genes Are Present in a Nonmammalian Vertebrate and Are Highly Expressed during Embryonic Development. Proc. Natl. Acad. Sci. USA 1997, 94, 8622–8627. [Google Scholar] [CrossRef]
- Razali, K.; Othman, N.; Mohd Nasir, M.H.; Doolaanea, A.A.; Kumar, J.; Ibrahim, W.N.; Mohamed Ibrahim, N.; Mohamed, W.M.Y. The Promise of the Zebrafish Model for Parkinson’s Disease: Today’s Science and Tomorrow’s Treatment. Front. Genet. 2021, 12, 655550. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, C.; Singh, A. Zebrafish an Experimental Model of Huntington’s Disease: Molecular Aspects, Therapeutic Targets and Current Challenges. Mol. Biol. Rep. 2021, 48, 8181–8194. [Google Scholar] [CrossRef]
- Ebrahimie, E.; Fruzangohar, M.; Moussavi Nik, S.H.; Newman, M. Gene Ontology-Based Analysis of Zebrafish Omics Data Using the Web Tool Comparative Gene Ontology. Zebrafish 2017, 14, 492–494. [Google Scholar] [CrossRef]
- Lacour, M.; Quenez, O.; Rovelet-Lecrux, A.; Salomon, B.; Rousseau, S.; Richard, A.-C.; Quillard-Muraine, M.; Pasquier, F.; Rollin-Sillaire, A.; Martinaud, O.; et al. Causative Mutations and Genetic Risk Factors in Sporadic Early Onset Alzheimer’s Disease Before 51 Years. J. Alzheimer’s Dis. 2019, 71, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Ryan, N.S.; Nicholas, J.M.; Weston, P.S.J.; Liang, Y.; Lashley, T.; Guerreiro, R.; Adamson, G.; Kenny, J.; Beck, J.; Chavez-Gutierrez, L.; et al. Clinical Phenotype and Genetic Associations in Autosomal Dominant Familial Alzheimer’s Disease: A Case Series. Lancet Neurol. 2016, 15, 1326–1335. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; An, S.S.A.; Kim, S. Mutations in Presenilin 2 and Its Implications in Alzheimer’s Disease and Other Dementia-Associated Disorders. Clin. Interv. Aging 2015, 10, 1163–1172. [Google Scholar] [CrossRef]
- Lanoiselée, H.-M.; Nicolas, G.; Wallon, D.; Rovelet-Lecrux, A.; Lacour, M.; Rousseau, S.; Richard, A.-C.; Pasquier, F.; Rollin-Sillaire, A.; Martinaud, O.; et al. APP, PSEN1, and PSEN2 Mutations in Early-Onset Alzheimer Disease: A Genetic Screening Study of Familial and Sporadic Cases. PLOS Med. 2017, 14, e1002270. [Google Scholar] [CrossRef]
- Kim, J.; Basak, J.M.; Holtzman, D.M. The Role of Apolipoprotein E in Alzheimer’s Disease. Neuron 2009, 63, 287–303. [Google Scholar] [CrossRef]
- Van den Bossche, T.; Sleegers, K.; Cuyvers, E.; Engelborghs, S.; Sieben, A.; De Roeck, A.; Van Cauwenberghe, C.; Vermeulen, S.; Van den Broeck, M.; Laureys, A.; et al. Phenotypic Characteristics of Alzheimer Patients Carrying an ABCA7 Mutation. Neurology 2016, 86, 2126. [Google Scholar] [CrossRef]
- Marelli, F.; Persani, L. Zebrafish Come Modello per Lo Studio Di Malattie Della Tiroide. L’Endocrinologo 2021, 22, 42–49. [Google Scholar] [CrossRef]
- Lemmens, R.; Van Hoecke, A.; Hersmus, N.; Geelen, V.; D’Hollander, I.; Thijs, V.; Van Den Bosch, L.; Carmeliet, P.; Robberecht, W. Overexpression of Mutant Superoxide Dismutase 1 Causes a Motor Axonopathy in the Zebrafish. Hum. Mol. Genet. 2007, 16, 2359–2365. [Google Scholar] [CrossRef] [PubMed]
- Zoghbi, H.Y.; Orr, H.T. Pathogenic Mechanisms of a Polyglutamine-Mediated Neurodegenerative Disease, Spinocerebellar Ataxia Type 1. J. Biol. Chem. 2009, 284, 7425–7429. [Google Scholar] [CrossRef]
- Bai, Q.; Wei, X.; Burton, E.A. Expression of a 12-Kb Promoter Element Derived from the Zebrafish Enolase-2 Gene in the Zebrafish Visual System. Neurosci. Lett. 2009, 449, 252–257. [Google Scholar] [CrossRef][Green Version]
- Paquet, D.; Bhat, R.; Sydow, A.; Mandelkow, E.-M.; Berg, S.; Hellberg, S.; Fälting, J.; Distel, M.; Köster, R.W.; Schmid, B. A Zebrafish Model of Tauopathy Allows in Vivo Imaging of Neuronal Cell Death and Drug Evaluation. J. Clin. Investig. 2009, 119, 1382–1395. [Google Scholar] [CrossRef]
- Bashirzade, A.A.; Zabegalov, K.N.; Volgin, A.D.; Belova, A.S.; Demin, K.A.; de Abreu, M.S.; Babchenko, V.Y.; Bashirzade, K.A.; Yenkoyan, K.B.; Tikhonova, M.A.; et al. Modeling Neurodegenerative Disorders in Zebrafish. Neurosci. Biobehav. Rev. 2022, 138, 104679. [Google Scholar] [CrossRef]
- Luna, S.; Cameron, D.J.; Ethell, D.W. Amyloid-β and APP Deficiencies Cause Severe Cerebrovascular Defects: Important Work for an Old Villain. PLoS ONE 2013, 8, e75052. [Google Scholar] [CrossRef]
- Pu, Y.-Z.; Liang, L.; Fu, A.-L.; Liu, Y.; Sun, L.; Li, Q.; Wu, D.; Sun, M.-J.; Zhang, Y.-G.; Zhao, B.-Q. Generation of Alzheimer’s Disease Transgenic Zebrafish Expressing Human APP Mutation Under Control of Zebrafish Appb Promotor. Curr. Alzheimer’s. Res. 2017, 14, 668–679. [Google Scholar] [CrossRef]
- Hogl, S.; Van Bebber, F.; Dislich, B.; Kuhn, P.-H.; Haass, C.; Schmid, B.; Lichtenthaler, S.F. Label-Free Quantitative Analysis of the Membrane Proteome of Bace1 Protease Knock-out Zebrafish Brains. Proteomics 2013, 13, 1519–1527. [Google Scholar] [CrossRef]
- Sundvik, M.; Chen, Y.-C.; Panula, P. Presenilin1 Regulates Histamine Neuron Development and Behavior in Zebrafish, Danio Rerio. J. Neurosci. 2013, 33, 1589–1597. [Google Scholar] [CrossRef]
- Newman, M.; Tucker, B.; Nornes, S.; Ward, A.; Lardelli, M. Altering Presenilin Gene Activity in Zebrafish Embryos Causes Changes in Expression of Genes with Potential Involvement in Alzheimer’s Disease Pathogenesis. J. Alzheimer’s Dis. 2009, 16, 133–147. [Google Scholar] [CrossRef]
- Nornes, S.; Newman, M.; Verdile, G.; Wells, S.; Stoick-Cooper, C.L.; Tucker, B.; Frederich-Sleptsova, I.; Martins, R.; Lardelli, M. Interference with Splicing of Presenilin Transcripts Has Potent Dominant Negative Effects on Presenilin Activity. Hum. Mol. Genet 2008, 17, 402–412. [Google Scholar] [CrossRef]
- Campbell, W.A.; Yang, H.; Zetterberg, H.; Baulac, S.; Sears, J.A.; Liu, T.; Wong, S.T.C.; Zhong, T.P.; Xia, W. Zebrafish Lacking Alzheimer Presenilin Enhancer 2 (Pen-2) Demonstrate Excessive P53-Dependent Apoptosis and Neuronal Loss. J. Neurochem. 2006, 96, 1423–1440. [Google Scholar] [CrossRef]
- Milanese, C.; Sager, J.J.; Bais, Q.; Farrells, T.C.; Cannons, J.R.; Greenamyre, J.T.; Burtons, E.A. Hypokinesia and Reduced Dopamine Levels in Zebrafish Lacking β- and Γ1-Synucleins. J. Biol. Chem. 2012, 287, 2971–2983. [Google Scholar] [CrossRef]
- Vaz, R.L.; Outeiro, T.F.; Ferreira, J.J. Zebrafish as an Animal Model for Drug Discovery in Parkinson’s Disease and Other Movement Disorders: A Systematic Review. Front. Neurol. 2018, 9, 347. [Google Scholar] [CrossRef] [PubMed]
- Quinn, P.M.J.; Moreira, P.I.; Ambrósio, A.F.; Alves, C.H. PINK1/PARKIN Signalling in Neurodegeneration and Neuroinflammation. Acta Neuropathol. Commun. 2020, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Anichtchik, O.; Diekmann, H.; Fleming, A.; Roach, A.; Goldsmith, P.; Rubinsztein, D.C. Loss of PINK1 Function Affects Development and Results in Neurodegeneration in Zebrafish. J. Neurosci. 2008, 28, 8199–8207. [Google Scholar] [CrossRef] [PubMed]
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.K.; Harvey, K.; Gispert, S.; Ali, Z.; Del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary Early-Onset Parkinson’s Disease Caused by Mutations in PINK1. Science 2004, 304, 1158–1160. [Google Scholar] [CrossRef] [PubMed]
- Flinn, L.; Mortiboys, H.; Volkmann, K.; Kster, R.W.; Ingham, P.W.; Bandmann, O. Complex i Deficiency and Dopaminergic Neuronal Cell Loss in Parkin-Deficient Zebrafish (Danio Rerio). Brain 2009, 132, 1613–1623. [Google Scholar] [CrossRef]
- Pap, D.; Veres-Székely, A.; Szebeni, B.; Vannay, Á. PARK7/DJ-1 as a Therapeutic Target in Gut-Brain Axis Diseases. Int. J. Mol. Sci. 2022, 23, 6626. [Google Scholar] [CrossRef]
- Antipova, D.; Bandopadhyay, R. Expression of DJ-1 in Neurodegenerative Disorders. In DJ-1/PARK7 Protein: Parkinson’s Disease, Cancer and Oxidative Stress-Induced Diseases; Ariga, H., Iguchi-Ariga, S.M.M., Eds.; Springer: Singapore, 2017; pp. 25–43. ISBN 978-981-10-6583-5. [Google Scholar]
- Bretaud, S.; Allen, C.; Ingham, P.W.; Bandmann, O. P53-Dependent Neuronal Cell Death in a DJ-1-Deficient Zebrafish Model of Parkinson’s Disease. J. Neurochem. 2007, 100, 1626–1635. [Google Scholar] [CrossRef]
- Ghosh, R.; Tabrizi, S.J. Clinical Features of Huntington’s Disease. In Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2018; Volume 1049, pp. 1–28. ISBN 978-3-319-71778-4. [Google Scholar]
- Karlovich, C.A.; John, R.M.; Ramirez, L.; Stainier, D.Y.R.; Myers, R.M. Characterization of the Huntington’s Disease (HD) Gene Homolog in the Zebrafish Danio Rerio. Gene 1998, 217, 117–125. [Google Scholar] [CrossRef]
- Sidik, H.; Ang, C.J.; Pouladi, M.A. Huntingtin Confers Fitness but Is Not Embryonically Essential in Zebrafish Development. Dev. Biol. 2020, 458, 98–105. [Google Scholar] [CrossRef]
- Veldman, M.B.; Rios-Galdamez, Y.; Lu, X.-H.; Gu, X.; Qin, W.; Li, S.; Yang, X.W.; Lin, S. The N17 Domain Mitigates Nuclear Toxicity in a Novel Zebrafish Huntington’s Disease Model. Mol. Neurodegener. 2015, 10, 67. [Google Scholar] [CrossRef]
- Braems, E.; Tziortzouda, P.; Van Den Bosch, L. Exploring the Alternative: Fish, Flies and Worms as Preclinical Models for ALS. Neurosci. Lett. 2021, 759, 136041. [Google Scholar] [CrossRef]
- Paez-Colasante, X.; Figueroa-Romero, C.; Sakowski, S.A.; Goutman, S.A.; Feldman, E.L. Amyotrophic Lateral Sclerosis: Mechanisms and Therapeutics in the Epigenomic Era. Nat. Rev. Neurol. 2015, 11, 266–279. [Google Scholar] [CrossRef]
- Ramesh, T.; Lyon, A.N.; Pineda, R.H.; Wang, C.; Janssen, P.M.L.; Canan, B.D.; Burghes, A.H.M.; Beattie, C.E. A Genetic Model of Amyotrophic Lateral Sclerosis in Zebrafish Displays Phenotypic Hallmarks of Motoneuron Disease. DMM Dis. Model. Mech. 2010, 3, 652–662. [Google Scholar] [CrossRef]
- Da Costa, M.M.J.; Allen, C.E.; Higginbottom, A.; Ramesh, T.; Shaw, P.J.; McDermott, C.J. A New Zebrafish Model Produced by TILLING of SOD1-Related Amyotrophic Lateral Sclerosis Replicates Key Features of the Disease and Represents a Tool for in Vivo Therapeutic Screening. DMM Dis. Model. Mech. 2014, 7, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Lissouba, A.; Liao, M.; Kabashi, E.; Drapeau, P. Transcriptomic Analysis of Zebrafish TDP-43 Transgenic Lines. Front. Mol. Neurosci. 2018, 11, 463. [Google Scholar] [CrossRef]
- Marrone, L.; Drexler, H.C.A.; Wang, J.; Tripathi, P.; Distler, T.; Heisterkamp, P.; Anderson, E.N.; Kour, S.; Moraiti, A.; Maharana, S.; et al. FUS Pathology in ALS Is Linked to Alterations in Multiple ALS-Associated Proteins and Rescued by Drugs Stimulating Autophagy. Acta Neuropathol. 2019, 138, 67–84. [Google Scholar] [CrossRef]
- Bourefis, A.-R.; Campanari, M.-L.; Buee-Scherrer, V.; Kabashi, E. Functional Characterization of a FUS Mutant Zebrafish Line as a Novel Genetic Model for ALS. Neurobiol. Dis. 2020, 142, 104935. [Google Scholar] [CrossRef]
- Kustermann, M.; Manta, L.; Paone, C.; Kustermann, J.; Lausser, L.; Wiesner, C.; Eichinger, L.; Clemen, C.S.; Schröder, R.; Kestler, H.A.; et al. Loss of the Novel Vcp (Valosin Containing Protein) Interactor Washc4 Interferes with Autophagy-Mediated Proteostasis in Striated Muscle and Leads to Myopathy in Vivo. Autophagy 2018, 14, 1911–1927. [Google Scholar] [CrossRef]
- Ciura, S.; Lattante, S.; Le Ber, I.; Latouche, M.; Tostivint, H.; Brice, A.; Kabashi, E. Loss of Function of C9orf72 Causes Motor Deficits in a Zebrafish Model of Amyotrophic Lateral Sclerosis. Ann. Neurol. 2013, 74, 180–187. [Google Scholar] [CrossRef]
- Shaw, M.P.; Higginbottom, A.; McGown, A.; Castelli, L.M.; James, E.; Hautbergue, G.M.; Shaw, P.J.; Ramesh, T.M. Stable Transgenic C9orf72 Zebrafish Model Key Aspects of the ALS/FTD Phenotype and Reveal Novel Pathological Features. Acta Neuropathol. Commun. 2018, 6, 125. [Google Scholar] [CrossRef]
- Swinnen, B.; Bento-Abreu, A.; Gendron, T.F.; Boeynaems, S.; Bogaert, E.; Nuyts, R.; Timmers, M.; Scheveneels, W.; Hersmus, N.; Wang, J.; et al. A Zebrafish Model for C9orf72 ALS Reveals RNA Toxicity as a Pathogenic Mechanism. Acta Neuropathol. 2018, 135, 427–443. [Google Scholar] [CrossRef] [PubMed]
- Guo, S. Linking Genes to Brain, Behavior and Neurological Diseases: What Can We Learn from Zebrafish? Genes Brain Behav. 2004, 3, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Hutton, M. Missense and Splice Site Mutations in Tau Associated with FTDP-17: Multiple Pathogenic Mechanisms. Neurology 2001, 56, S21–S25. [Google Scholar] [CrossRef] [PubMed]
- Tomasiewicz, H.G.; Flaherty, D.B.; Soria, J.P.; Wood, J.G. Transgenic Zebrafish Model of Neurodegeneration. J. Neurosci. Res. 2002, 70, 734–745. [Google Scholar] [CrossRef]
- Mazanetz, M.P.; Fischer, P.M. Untangling Tau Hyperphosphorylation in Drug Design for Neurodegenerative Diseases. Nat. Rev. Drug Discov. 2007, 6, 464–479. [Google Scholar] [CrossRef]
- Jeong, J.-Y.; Kwon, H.-B.; Ahn, J.-C.; Kang, D.; Kwon, S.-H.; Park, J.A.; Kim, K.-W. Functional and Developmental Analysis of the Blood–Brain Barrier in Zebrafish. Brain Res. Bull. 2008, 75, 619–628. [Google Scholar] [CrossRef]
- Xie, J.; Farage, E.; Sugimoto, M.; Anand-Apte, B. A Novel Transgenic Zebrafish Model for Blood-Brain and Blood-Retinal Barrier Development. BMC Dev. Biol. 2010, 10, 76. [Google Scholar] [CrossRef]
- Jurisch-Yaksi, N.; Yaksi, E.; Kizil, C. Radial Glia in the Zebrafish Brain: Functional, Structural, and Physiological Comparison with the Mammalian Glia. Glia 2020, 68, 2451–2470. [Google Scholar] [CrossRef]
- Schmidt, R.; Strähle, U.; Scholpp, S. Neurogenesis in Zebrafish–from Embryo to Adult. Neural Dev. 2013, 8, 3. [Google Scholar] [CrossRef]
- Wullimann, M.F.; Mueller, T. Teleostean and Mammalian Forebrains Contrasted: Evidence from Genes to Behavior. J. Comp. Neurol. 2004, 475, 143–162. [Google Scholar] [CrossRef]
- Mueller, T.; Wullimann, M.F.; Guo, S. Early Teleostean Basal Ganglia Development Visualized by Zebrafish Dlx2a, Lhx6, Lhx7, Tbr2 (Eomesa), and GAD67 Gene Expression. J. Comp. Neurol. 2008, 507, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Rink, E.; Wullimann, M.F. Connections of the Ventral Telencephalon (Subpallium) in the Zebrafish (Danio Rerio). Brain Res. 2004, 1011, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Diotel, N.; Lübke, L.; Strähle, U.; Rastegar, S. Common and Distinct Features of Adult Neurogenesis and Regeneration in the Telencephalon of Zebrafish and Mammals. Front. Neurosci. 2020, 14, 568930. [Google Scholar] [CrossRef] [PubMed]
- Ghaddar, B.; Lübke, L.; Couret, D.; Rastegar, S.; Diotel, N. Cellular Mechanisms Participating in Brain Repair of Adult Zebrafish and Mammals after Injury. Cells 2021, 10, 391. [Google Scholar] [CrossRef] [PubMed]
- Doetsch, F.; Scharff, C. Challenges for Brain Repair: Insights from Adult Neurogenesis in Birds and Mammals. Brain Behav. Evol. 2001, 58, 306–322. [Google Scholar] [CrossRef]
- Chapouton, P.; Jagasia, R.; Bally-Cuif, L. Adult Neurogenesis in Non-mammalian Vertebrates. Bioessays 2007, 29, 745–757. [Google Scholar] [CrossRef]
- Kaslin, J.; Ganz, J.; Brand, M. Proliferation, Neurogenesis and Regeneration in the Non-Mammalian Vertebrate Brain. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 101–122. [Google Scholar] [CrossRef]
- Grandel, H.; Kaslin, J.; Ganz, J.; Wenzel, I.; Brand, M. Neural Stem Cells and Neurogenesis in the Adult Zebrafish Brain: Origin, Proliferation Dynamics, Migration and Cell Fate. Dev. Biol. 2006, 295, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Altman, J. Autoradiographic and Histological Studies of Postnatal Neurogenesis. IV. Cell Proliferation and Migration in the Anterior Forebrain, with Special Reference to Persisting Neurogenesis in the Olfactory Bulb. J. Comp. Neurol. 1969, 137, 433–457. [Google Scholar] [CrossRef]
- Hack, M.A.; Saghatelyan, A.; de Chevigny, A.; Pfeifer, A.; Ashery-Padan, R.; Lledo, P.-M.; Götz, M. Neuronal Fate Determinants of Adult Olfactory Bulb Neurogenesis. Nat. Neurosci. 2005, 8, 865–872. [Google Scholar] [CrossRef]
- Seri, B.; Garcıa-Verdugo, J.M.; McEwen, B.S.; Alvarez-Buylla, A. Astrocytes Give Rise to New Neurons in the Adult Mammalian Hippocampus. J. Neurosci. 2001, 21, 7153–7160. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.S.; Bell, D.H. Mitotic Neuroblasts in the 9-Day-Old and 11-Month-Old Rodent Hippocampus. J. Neurosci. 1984, 4, 1429–1441. [Google Scholar] [CrossRef] [PubMed]
- Gould, E. How Widespread Is Adult Neurogenesis in Mammals? Nat. Rev. Neurosci. 2007, 8, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Palmer, T.D.; Ray, J.; Gage, F.H. FGF-2-Responsive Neuronal Progenitors Reside in Proliferative and Quiescent Regions of the Adult Rodent Brain. Mol. Cell. Neurosci. 1995, 6, 474–486. [Google Scholar] [CrossRef]
- Pencea, V.; Bingaman, K.D.; Wiegand, S.J.; Luskin, M.B. Infusion of Brain-Derived Neurotrophic Factor into the Lateral Ventricle of the Adult Rat Leads to New Neurons in the Parenchyma of the Striatum, Septum, Thalamus, and Hypothalamus. J. Neurosci. 2001, 21, 6706–6717. [Google Scholar] [CrossRef]
- Reynolds, B.A.; Weiss, S. Generation of Neurons and Astrocytes from Isolated Cells of the Adult Mammalian Central Nervous System. Science 1992, 255, 1707–1710. [Google Scholar] [CrossRef]
- Rivers, L.E.; Young, K.M.; Rizzi, M.; Jamen, F.; Psachoulia, K.; Wade, A.; Kessaris, N.; Richardson, W.D. PDGFRA/NG2 Glia Generate Myelinating Oligodendrocytes and Piriform Projection Neurons in Adult Mice. Nat. Neurosci. 2008, 11, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Lie, D.C.; Dziewczapolski, G.; Willhoite, A.R.; Kaspar, B.K.; Shults, C.W.; Gage, F.H. The Adult Substantia Nigra Contains Progenitor Cells with Neurogenic Potential. J. Neurosci. 2002, 22, 6639–6649. [Google Scholar] [CrossRef]
- Nunes, M.C.; Roy, N.S.; Keyoung, H.M.; Goodman, R.R.; McKhann, G.; Jiang, L.; Kang, J.; Nedergaard, M.; Goldman, S.A. Identification and Isolation of Multipotential Neural Progenitor Cells from the Subcortical White Matter of the Adult Human Brain. Nat. Med. 2003, 9, 439–447. [Google Scholar] [CrossRef]
- Kokoeva, M.V.; Yin, H.; Flier, J.S. Neurogenesis in the Hypothalamus of Adult Mice: Potential Role in Energy Balance. Science 2005, 310, 679–683. [Google Scholar] [CrossRef]
- Migaud, M.; Batailler, M.; Segura, S.; Duittoz, A.; Franceschini, I.; Pillon, D. Emerging New Sites for Adult Neurogenesis in the Mammalian Brain: A Comparative Study between the Hypothalamus and the Classical Neurogenic Zones. Eur. J. Neurosci. 2010, 32, 2042–2052. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.A.; Bedont, J.L.; Pak, T.; Wang, H.; Song, J.; Miranda-Angulo, A.; Takiar, V.; Charubhumi, V.; Balordi, F.; Takebayashi, H. Tanycytes of the Hypothalamic Median Eminence Form a Diet-Responsive Neurogenic Niche. Nat. Neurosci. 2012, 15, 700–702. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Sallinen, V.; Sundvik, M.; Kolehmainen, J.; Torkko, V.; Tiittula, A.; Moshnyakov, M.; Podlasz, P. Modulatory Neurotransmitter Systems and Behavior: Towards Zebrafish Models of Neurodegenerative Diseases. Zebrafish 2006, 3, 235–247. [Google Scholar] [CrossRef]
- Zupanc, G.K. Adult Neurogenesis and Neuronal Regeneration in the Brain of Teleost Fish. J. Physiol. -Paris 2008, 102, 357–373. [Google Scholar] [CrossRef]
- Pellegrini, E.; Mouriec, K.; Anglade, I.; Menuet, A.; Le Page, Y.; Gueguen, M.-M.; Marmignon, M.-H.; Brion, F.; Pakdel, F.; Kah, O. Identification of Aromatase-positive Radial Glial Cells as Progenitor Cells in the Ventricular Layer of the Forebrain in Zebrafish. J. Comp. Neurol. 2007, 501, 150–167. [Google Scholar] [CrossRef]
- Pinto, L.; Götz, M. Radial Glial Cell Heterogeneity—The Source of Diverse Progeny in the CNS. Prog. Neurobiol. 2007, 83, 2–23. [Google Scholar] [CrossRef]
- Rowitch, D.H.; Kriegstein, A.R. Developmental Genetics of Vertebrate Glial–Cell Specification. Nature 2010, 468, 214–222. [Google Scholar] [CrossRef]
- Malatesta, P.; Hartfuss, E.; Gotz, M. Isolation of Radial Glial Cells by Fluorescent-Activated Cell Sorting Reveals a Neuronal Lineage. Development 2000, 127, 5253–5263. [Google Scholar] [CrossRef]
- Noctor, S.C.; Flint, A.C.; Weissman, T.A.; Dammerman, R.S.; Kriegstein, A.R. Neurons Derived from Radial Glial Cells Establish Radial Units in Neocortex. Nature 2001, 409, 714–720. [Google Scholar] [CrossRef]
- Fares, J.; Bou Diab, Z.; Nabha, S.; Fares, Y. Neurogenesis in the Adult Hippocampus: History, Regulation, and Prospective Roles. Int. J. Neurosci. 2019, 129, 598–611. [Google Scholar] [CrossRef]
- Rothenaigner, I.; Krecsmarik, M.; Hayes, J.A.; Bahn, B.; Lepier, A.; Fortin, G.; Götz, M.; Jagasia, R.; Bally-Cuif, L. Clonal Analysis by Distinct Viral Vectors Identifies Bona Fide Neural Stem Cells in the Adult Zebrafish Telencephalon and Characterizes Their Division Properties and Fate. Development 2011, 138, 1459–1469. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.S.; März, M.; Strähle, U. GFAP and Nestin Reporter Lines Reveal Characteristics of Neural Progenitors in the Adult Zebrafish Brain. Dev. Dyn. 2009, 238, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Adolf, B.; Chapouton, P.; Lam, C.S.; Topp, S.; Tannhäuser, B.; Strähle, U.; Götz, M.; Bally-Cuif, L. Conserved and Acquired Features of Adult Neurogenesis in the Zebrafish Telencephalon. Dev. Biol. 2006, 295, 278–293. [Google Scholar] [CrossRef]
- Zupanc, G.K.; Clint, S.C.; Takimoto, N.; Hughes, A.T.; Wellbrock, U.M.; Meissner, D. Spatio-Temporal Distribution of Microglia/Macrophages during Regeneration in the Cerebellum of Adult Teleost Fish, Apteronotus Leptorhynchus: A Quantitative Analysis. Brain Behav. Evol. 2003, 62, 31–42. [Google Scholar] [CrossRef]
- Diotel, N.; Vaillant, C.; Kah, O.; Pellegrini, E. Mapping of Brain Lipid Binding Protein (Blbp) in the Brain of Adult Zebrafish, Co-Expression with Aromatase B and Links with Proliferation. Gene Expr Patterns 2016, 20, 42–54. [Google Scholar] [CrossRef]
- März, M.; Chapouton, P.; Diotel, N.; Vaillant, C.; Hesl, B.; Takamiya, M.; Lam, C.S.; Kah, O.; Bally-Cuif, L.; Strähle, U. Heterogeneity in Progenitor Cell Subtypes in the Ventricular Zone of the Zebrafish Adult Telencephalon. Glia 2010, 58, 870–888. [Google Scholar] [CrossRef]
- Zupanc, G.K.; Zupanc, M.M. New Neurons for the Injured Brain: Mechanisms of Neuronal Regeneration in Adult Teleost Fish. 2006. Available online: https://www.futuremedicine.com/doi/abs/10.2217/17460751.1.2.207 (accessed on 5 October 2022).
- Gerhard, G.S.; Kauffman, E.J.; Wang, X.; Stewart, R.; Moore, J.L.; Kasales, C.J.; Demidenko, E.; Cheng, K.C. Life Spans and Senescent Phenotypes in Two Strains of Zebrafish (Danio Rerio). Exp. Gerontol. 2002, 37, 1055–1068. [Google Scholar] [CrossRef]
- Hinsch, K.; Zupanc, G.K.H. Generation and Long-Term Persistence of New Neurons in the Adult Zebrafish Brain: A Quantitative Analysis. Neuroscience 2007, 146, 679–696. [Google Scholar] [CrossRef]
- Kizil, C.; Kaslin, J.; Kroehne, V.; Brand, M. Adult Neurogenesis and Brain Regeneration in Zebrafish. Dev. Neurobiol. 2012, 72, 429–461. [Google Scholar] [CrossRef]
- Zupanc, G.K.; Horschke, I.; Ott, R.; Rascher, G.B. Postembryonic Development of the Cerebellum in Gymnotiform Fish. J. Comp. Neurol. 1996, 370, 443–464. [Google Scholar] [CrossRef]
- Ott, R.; Zupanc, G.K.; Horschke, I. Long-Term Survival of Postembryonically Born Cells in the Cerebellum of Gymnotiform Fish, Apteronotus Leptorhynchus. Neurosci. Lett. 1997, 221, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Edelmann, K.; Glashauser, L.; Sprungala, S.; Hesl, B.; Fritschle, M.; Ninkovic, J.; Godinho, L.; Chapouton, P. Increased Radial Glia Quiescence, Decreased Reactivation upon Injury and Unaltered Neuroblast Behavior Underlie Decreased Neurogenesis in the Aging Zebrafish Telencephalon. J. Comp. Neurol. 2013, 521, 3099–3115. [Google Scholar] [CrossRef] [PubMed]
- Cosacak, M.I.; Bhattarai, P.; De Jager, P.L.; Menon, V.; Tosto, G.; Kizil, C. Single Cell/Nucleus Transcriptomics Comparison in Zebrafish and Humans Reveals Common and Distinct Molecular Responses to Alzheimer’s Disease. Cells 2022, 11, 1807. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, P.; Thomas, A.K.; Cosacak, M.I.; Papadimitriou, C.; Mashkaryan, V.; Froc, C.; Reinhardt, S.; Kurth, T.; Dahl, A.; Zhang, Y.; et al. IL4/STAT6 Signaling Activates Neural Stem Cell Proliferation and Neurogenesis upon Amyloid-Β42 Aggregation in Adult Zebrafish Brain. Cell Rep. 2016, 17, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Duan, W.; Mattson, M.P. Evidence That Brain-Derived Neurotrophic Factor Is Required for Basal Neurogenesis and Mediates, in Part, the Enhancement of Neurogenesis by Dietary Restriction in the Hippocampus of Adult Mice. J Neurochem 2002, 82, 1367–1375. [Google Scholar] [CrossRef]
- Cacialli, P.; D’angelo, L.; Kah, O.; Coumailleau, P.; Gueguen, M.; Pellegrini, E.; Lucini, C. Neuronal Expression of Brain Derived Neurotrophic Factor in the Injured Telencephalon of Adult Zebrafish. J. Comp. Neurol. 2018, 526, 569–582. [Google Scholar] [CrossRef]
- Bhattarai, P.; Cosacak, M.I.; Mashkaryan, V.; Demir, S.; Popova, S.D.; Govindarajan, N.; Brandt, K.; Zhang, Y.; Chang, W.; Ampatzis, K.; et al. Neuron-Glia Interaction through Serotonin-BDNF-NGFR Axis Enables Regenerative Neurogenesis in Alzheimer’s Model of Adult Zebrafish Brain. PLOS Biol. 2020, 18, e3000585. [Google Scholar] [CrossRef]
- Kovacs, G.G. Molecular Pathological Classification of Neurodegenerative Diseases: Turning towards Precision Medicine. Int. J. Mol. Sci. 2016, 17, 189. [Google Scholar] [CrossRef]
- Jellinger, K.A. Recent Advances in Our Understanding of Neurodegeneration. J. Neural Transm. 2009, 116, 1111–1162. [Google Scholar]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a Risk Factor for Neurodegenerative Disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef]
- Gupta, J.; Fatima, M.T.; Islam, Z.; Khan, R.H.; Uversky, V.N.; Salahuddin, P. Nanoparticle Formulations in the Diagnosis and Therapy of Alzheimer’s Disease. Int. J. Biol. Macromol. 2019, 130, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Martínez, J.D.; Valdés, A.; Gallego, R.; Suárez-Montenegro, Z.J.; Alarcón, M.; Ibañez, E.; Alvarez-Rivera, G.; Cifuentes, A. Blood-Brain Barrier Permeability Study of Potential Neuroprotective Compounds Recovered From Plants and Agri-Food by-Products. Front. Nutr. 2022, 9, 924596. [Google Scholar] [CrossRef] [PubMed]
- Gence, L.; Fernezelian, D.; Bringart, M.; Veeren, B.; Christophe, A.; Brion, F.; Meilhac, O.; Bascands, J.-L.; Diotel, N. Hypericum Lanceolatum Lam. Medicinal Plant: Potential Toxicity and Therapeutic Effects Based on a Zebrafish Model. Front. Pharm. 2022, 13, 832928. [Google Scholar] [CrossRef] [PubMed]
- Ghaddar, B.; Gence, L.; Veeren, B.; Bringart, M.; Bascands, J.-L.; Meilhac, O.; Diotel, N. Aqueous Extract of Psiloxylon Mauritianum, Rich in Gallic Acid, Prevents Obesity and Associated Deleterious Effects in Zebrafish. Antioxidants 2022, 11, 1309. [Google Scholar] [CrossRef]
- Ghaddar, B.; Veeren, B.; Rondeau, P.; Bringart, M.; Lefebvre d’Hellencourt, C.; Meilhac, O.; Bascands, J.-L.; Diotel, N. Impaired Brain Homeostasis and Neurogenesis in Diet-Induced Overweight Zebrafish: A Preventive Role from A. Borbonica Extract. Sci. Rep. 2020, 10, 14496. [Google Scholar] [CrossRef]
- D’Onofrio, G.; Sancarlo, D.; Ruan, Q.; Yu, Z.; Panza, F.; Daniele, A.; Greco, A.; Seripa, D. Phytochemicals in the Treatment of Alzheimer’s Disease: A Systematic Review. Curr. Drug Targets 2017, 18, 1487–1498. [Google Scholar] [CrossRef]
- Dajas, F.; Rivera-Megret, F.; Blasina, F.; Arredondo, F.; Abin-Carriquiry, J.; Costa, G.; Echeverry, C.; Lafon, L.; Heizen, H.; Ferreira, M. Neuroprotection by Flavonoids. Braz. J. Med. Biol. Res. 2003, 36, 1613–1620. [Google Scholar] [CrossRef]
- Youdim, K.A.; Qaiser, M.Z.; Begley, D.J.; Rice-Evans, C.A.; Abbott, N.J. Flavonoid Permeability across an in Situ Model of the Blood–Brain Barrier. Free Radic. Biol. Med. 2004, 36, 592–604. [Google Scholar] [CrossRef]
- Talavéra, S.; Felgines, C.; Texier, O.; Besson, C.; Gil-Izquierdo, A.; Lamaison, J.-L.; Rémésy, C. Anthocyanin Metabolism in Rats and Their Distribution to Digestive Area, Kidney, and Brain. J. Agric. Food Chem. 2005, 53, 3902–3908. [Google Scholar] [CrossRef] [PubMed]
- Faal, M.; Manouchehri, H.; Changizi, R.; Bootorabi, F.; Khorramizadeh, M.R. Assessment of Resveratrol on Diabetes of Zebrafish (Danio Rerio). J. Diabetes Metab. Disord. 2022, 21, 823–833. [Google Scholar] [CrossRef]
- Ran, G.; Ying, L.; Li, L.; Yan, Q.; Yi, W.; Ying, C.; Wu, H.; Ye, X. Resveratrol Ameliorates Diet-Induced Dysregulation of Lipid Metabolism in Zebrafish (Danio Rerio). PLoS One 2017, 12, e0180865. [Google Scholar] [CrossRef] [PubMed]
- Meguro, S.; Hasumura, T.; Hase, T. Coffee Polyphenols Exert Hypocholesterolemic Effects in Zebrafish Fed a High-Cholesterol Diet. Nutr. Metab. 2013, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Abbate, F.; Maugeri, A.; Laurà, R.; Levanti, M.; Navarra, M.; Cirmi, S.; Germanà, A. Zebrafish as a Useful Model to Study Oxidative Stress-Linked Disorders: Focus on Flavonoids. Antioxidants 2021, 10, 668. [Google Scholar] [CrossRef] [PubMed]
- Solanki, I.; Parihar, P.; Mansuri, M.L.; Parihar, M.S. Flavonoid-Based Therapies in the Early Management of Neurodegenerative Diseases. Adv. Nutr. 2015, 6, 64–72. [Google Scholar] [CrossRef]
- Wang, J.; Cao, H. Zebrafish and Medaka: Important Animal Models for Human Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 10766. [Google Scholar] [CrossRef]
- Kim, S.; Kim, M.; Kang, M.-C.; Lee, H.H.L.; Cho, C.H.; Choi, I.; Park, Y.; Lee, S.-H. Antioxidant Effects of Turmeric Leaf Extract against Hydrogen Peroxide-Induced Oxidative Stress In Vitro in Vero Cells and In Vivo in Zebrafish. Antioxidants 2021, 10, 112. [Google Scholar] [CrossRef]
- Issac, P.K.; Guru, A.; Velayutham, M.; Pachaiappan, R.; Arasu, M.V.; Al-Dhabi, N.A.; Choi, K.C.; Harikrishnan, R.; Arockiaraj, J. Oxidative Stress Induced Antioxidant and Neurotoxicity Demonstrated in Vivo Zebrafish Embryo or Larval Model and Their Normalization Due to Morin Showing Therapeutic Implications. Life Sci. 2021, 283, 119864. [Google Scholar] [CrossRef]
- Horvathova, K.; Novotny, L.; Vachalkova, A. The Free Radical Scavenging Activity of Four Flavonoids Determined by the Comet Assay. Neoplasma 2003, 50, 291–295. [Google Scholar]
- Spencer, J.P.; Vauzour, D.; Rendeiro, C. Flavonoids and Cognition: The Molecular Mechanisms Underlying Their Behavioural Effects. Arch. Biochem. Biophys. 2009, 492, 1–9. [Google Scholar] [CrossRef]
- Braganhol, E.; Tamajusuku, A.S.; Bernardi, A.; Wink, M.R.; Battastini, A.M. Ecto-5′-Nucleotidase/CD73 Inhibition by Quercetin in the Human U138MG Glioma Cell Line. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2007, 1770, 1352–1359. [Google Scholar] [CrossRef]
- Mohebali, N.; Shahzadeh Fazeli, S.A.; Ghafoori, H.; Farahmand, Z.; MohammadKhani, E.; Vakhshiteh, F.; Ghamarian, A.; Farhangniya, M.; Sanati, M.H. Effect of Flavonoids Rich Extract of Capparis Spinosa on Inflammatory Involved Genes in Amyloid-Beta Peptide Injected Rat Model of Alzheimer’s Disease. Nutr. Neurosci. 2018, 21, 143–150. [Google Scholar] [CrossRef]
- Tongjaroenbuangam, W.; Ruksee, N.; Chantiratikul, P.; Pakdeenarong, N.; Kongbuntad, W.; Govitrapong, P. Neuroprotective Effects of Quercetin, Rutin and Okra (Abelmoschus Esculentus Linn.) in Dexamethasone-Treated Mice. Neurochem. Int. 2011, 59, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Richetti, S.K.; Blank, M.; Capiotti, K.M.; Piato, A.L.; Bogo, M.R.; Vianna, M.R.; Bonan, C.D. Quercetin and Rutin Prevent Scopolamine-Induced Memory Impairment in Zebrafish. Behav. Brain Res. 2011, 217, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Thirugnanam, T.; Madhusoodanan, S.; Santhakumar, K. Quercetin Rescues MPP+–Induced Parkinsonian-like Locomotor Impairment in Zebrafish Larvae. J. Appl. Biol. Biotechnol. 2022, 10, 1. [Google Scholar] [CrossRef]
- Pan, H.; Zhang, J.; Wang, Y.; Cui, K.; Cao, Y.; Wang, L.; Wu, Y. Linarin Improves the Dyskinesia Recovery in Alzheimer’s Disease Zebrafish by Inhibiting the Acetylcholinesterase Activity. Life Sci. 2019, 222, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yang, L.; Liu, Q.; Zhu, Z.; Yang, Q.; Liu, J.; Beta, T. Protective Effects of Cyanidin-3-O-Glucoside on BPA-Induced Neurodevelopmental Toxicity in Zebrafish Embryo Model. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2023, 264, 109525. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, J.; Liu, F.; Tong, L.; Chen, Z.; Chen, J.; He, H.; Xu, R.; Ma, Y.; Huang, C. Neuroprotective Effects of Anthocyanins and Its Major Component Cyanidin-3-O-Glucoside (C3G) in the Central Nervous System: An Outlined Review. Eur. J. Pharmacol. 2019, 858, 172500. [Google Scholar] [CrossRef]
- Thayumanavan, G.; Jeyabalan, S.; Fuloria, S.; Sekar, M.; Ravi, M.; Selvaraj, L.K.; Bala, L.; Chidambaram, K.; Gan, S.H.; Rani, N.N.I.M.; et al. Silibinin and Naringenin against Bisphenol A-Induced Neurotoxicity in Zebrafish Model—Potential Flavonoid Molecules for New Drug Design, Development, and Therapy for Neurological Disorders. Molecules 2022, 27, 2572. [Google Scholar] [CrossRef]
- Lee, D.; Kim, N.; Jeon, S.H.; Gee, M.S.; Ju, Y.-J.; Jung, M.-J.; Cho, J.S.; Lee, Y.; Lee, S.; Lee, J.K. Hesperidin Improves Memory Function by Enhancing Neurogenesis in a Mouse Model of Alzheimer’s Disease. Nutrients 2022, 14, 3125. [Google Scholar] [CrossRef]
- Sharma, P.; Kumari, S.; Sharma, J.; Purohit, R.; Singh, D. Hesperidin Interacts with CREB-BDNF Signaling Pathway to Suppress Pentylenetetrazole-Induced Convulsions in Zebrafish. Front. Pharmacol. 2021, 11, 607797. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, S.; Ba, S.; Dang, J.; Ren, Q.; Zhu, Y.; Liu, K.; Jin, M. Eucommia Ulmoides Olive Male Flower Extracts Ameliorate Alzheimer’s Disease-Like Pathology in Zebrafish via Regulating Autophagy, Acetylcholinesterase, and the Dopamine Transporter. Front. Mol. Neurosci. 2022, 15, 901953. [Google Scholar] [CrossRef] [PubMed]




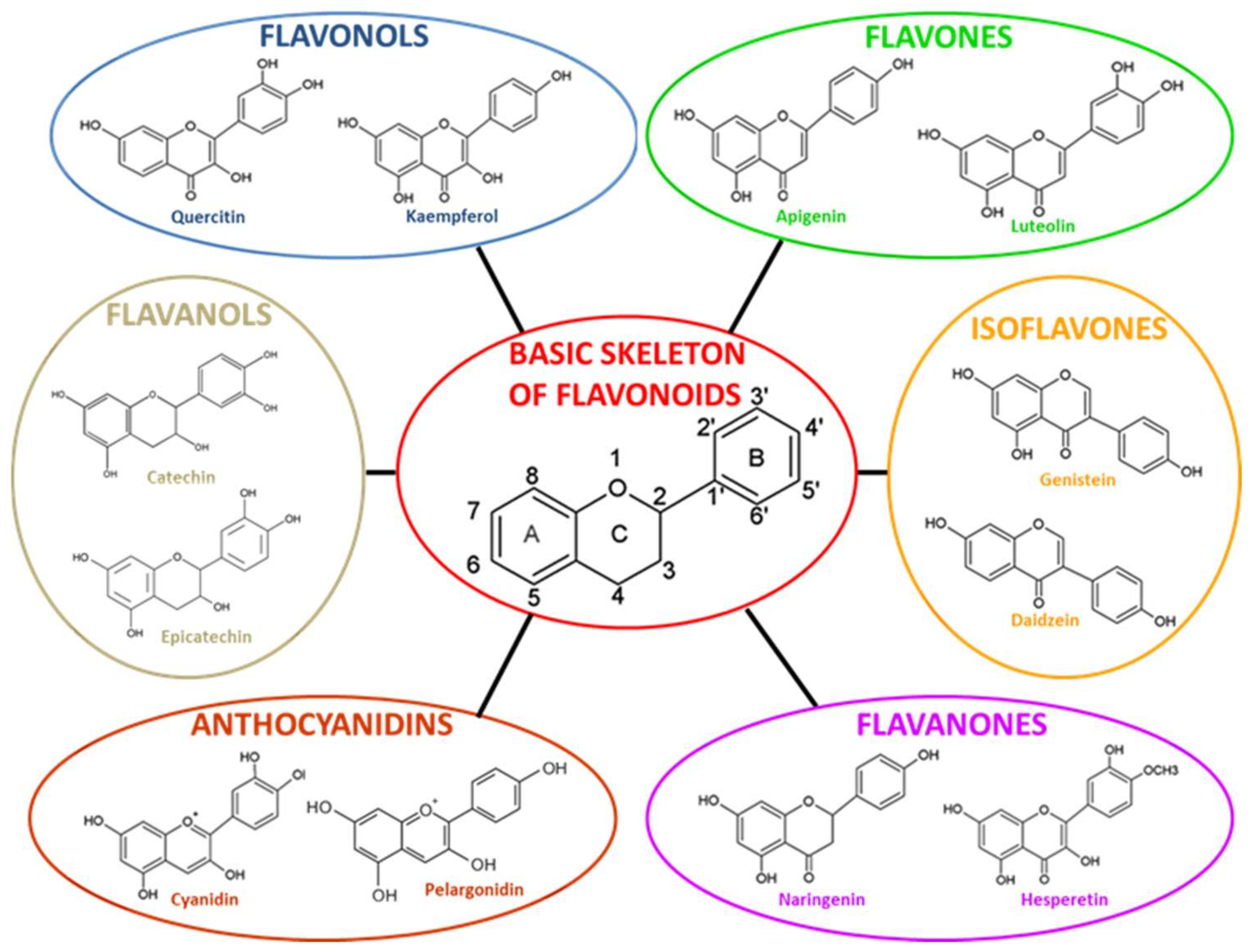{kind=link}
{kind=link}
{kind=link}
| Flavonoid Subclasses | Cmax | Time to Reach Cmax (h) | The Elimination Half-Life (h) |
|---|---|---|---|
| Isoflavones | 0.21–4.05 | 7.4–8.4 | 4.7–8.4 |
| Flavonols | <0.33–7.6 | <0.5–9.3 | 10.3–28.3 |
| Flavanones | 0.06–5.99 | 2–5.8 | 1.3–2.9 |
| Flavanols | 0.077–7.8 | 0.5–4.1 | 1–6.9 |
| Anthocyanidins | 0.011–0.041 | 2 | - |
| Flavonoid Subclass | Flavonoid | Antioxidant Potential | Anti-Inflammatory Potential | Neuroplasticity Potential | Anti- Neuropathological Protein Accumulation Potential | References |
|---|---|---|---|---|---|---|
| Flavonol | Quercetin | High capacity to scavenge ROS | Reduce the expression of inflammatory cytokines such as IL-6 and IL-1α | Increase of FoxG1, p-CREB and BDNF expression in DG promoting AHN | High affinity binding α-synuclein | [103,104] |
| Flavone | Apigenin | - | - | - | Reduced the expression and aggregation of α-synuclein | [105] |
| Flavanone | Hesperidin | Antioxidant action | - | - | Reduction of hippocampal Aβ levels | [70,106] |
| Flavonone | Hesperitin | - | - | Prevent neuronal apoptosis (inhibition of ASK1, caspases 3 and 9); Activation/phosphorylation of both Akt/PKB and ERK1/2 | Affect the expression of antioxidant enzymes such as GSH and SOD by modulating the nuclear factor erythroid 2-related factor 2 (Nrf2)-ARE pathway | [107,108] |
| Flavonol | Fisetin | Phosphorylation of ERK and 3-fold in CREB | - | - | - | [109] |
| Flavones | 3,5,6,7,8,3′,4′-heptamethoxyflavone | - | - | Inducing BDNF expression; Activation of cAMP/ERK/CREB signaling pathway | - | [110] |
| Flavone | Baicalin | - | - | Regulating the ERK/CREB/BDNF signaling pathway | - | [111] |
| Flavanols | Catechins | - | - | Increasing AHN | - | [112] |
| Curcumin flavonoids | - | - | Increasing AHN; Promotes hippocampal neurogenesis via the Wnt-β catenin pathway; Increasing the expression of Ngn2 and NeuroD1 (involved in neurogenesis); Promoted differentiation and maturation of new neurons | - | [113] | |
| Flavonol | Myricetin | Increasing AHN | [114] | |||
| Isoflavones | Genistein | - | Decrease the expression of Toll-Like 4 (TLR-4), inhibits microglial polarization and promoting the phenotypic switch from M1 to M2. | - | - | [115] |
| Flavone | Luteolin | - | Inhibit microglia activation by; Reducing the release of inflammatory mediators; Down-regulating TLR-4/TNF receptor-associated factor 6 (TRAF6)/NF-κB signaling | - | - | [116] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mhalhel, K.; Sicari, M.; Pansera, L.; Chen, J.; Levanti, M.; Diotel, N.; Rastegar, S.; Germanà, A.; Montalbano, G. Zebrafish: A Model Deciphering the Impact of Flavonoids on Neurodegenerative Disorders. Cells 2023, 12, 252. https://doi.org/10.3390/cells12020252
Mhalhel K, Sicari M, Pansera L, Chen J, Levanti M, Diotel N, Rastegar S, Germanà A, Montalbano G. Zebrafish: A Model Deciphering the Impact of Flavonoids on Neurodegenerative Disorders. Cells. 2023; 12(2):252. https://doi.org/10.3390/cells12020252
Chicago/Turabian StyleMhalhel, Kamel, Mirea Sicari, Lidia Pansera, Jincan Chen, Maria Levanti, Nicolas Diotel, Sepand Rastegar, Antonino Germanà, and Giuseppe Montalbano. 2023. "Zebrafish: A Model Deciphering the Impact of Flavonoids on Neurodegenerative Disorders" Cells 12, no. 2: 252. https://doi.org/10.3390/cells12020252
APA StyleMhalhel, K., Sicari, M., Pansera, L., Chen, J., Levanti, M., Diotel, N., Rastegar, S., Germanà, A., & Montalbano, G. (2023). Zebrafish: A Model Deciphering the Impact of Flavonoids on Neurodegenerative Disorders. Cells, 12(2), 252. https://doi.org/10.3390/cells12020252