

Concentration-Dependent Attenuation of Pro-Fibrotic Responses after Cannabigerol Exposure in Primary Rat Hepatocytes Cultured in Palmitate and Fructose Media
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Liver Perfusion Procedure
2.2. Preparation of Reagents for Cell Treatment
2.3. Cell Treatment
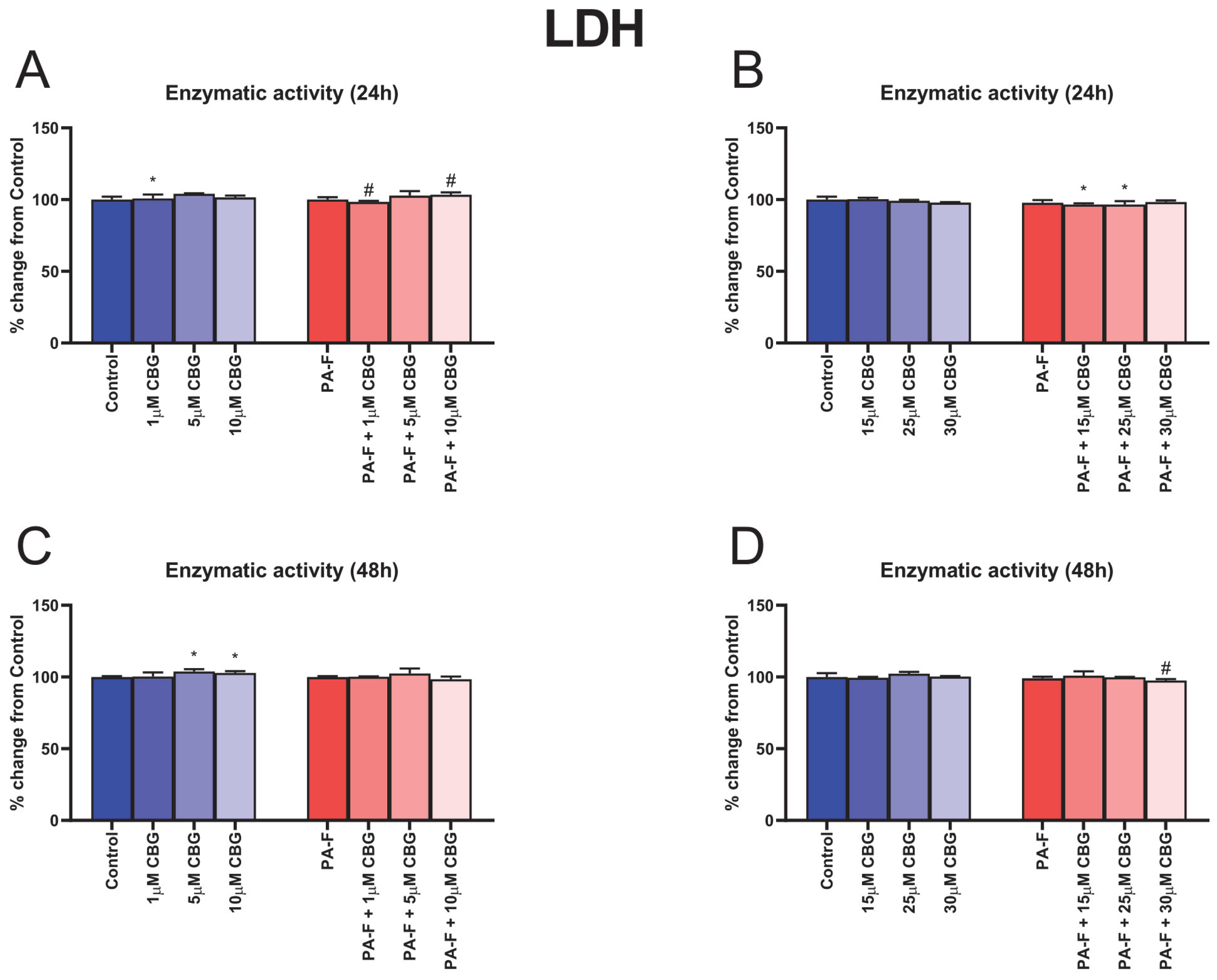
2.4. Cell Cytotoxicity
2.5. Protein Extraction and Western Blot Assay
2.6. RNA Extraction and Quantitative Real-Time PCR Assay
2.7. Immunoassay
2.8. Statistical Analysis
3. Results
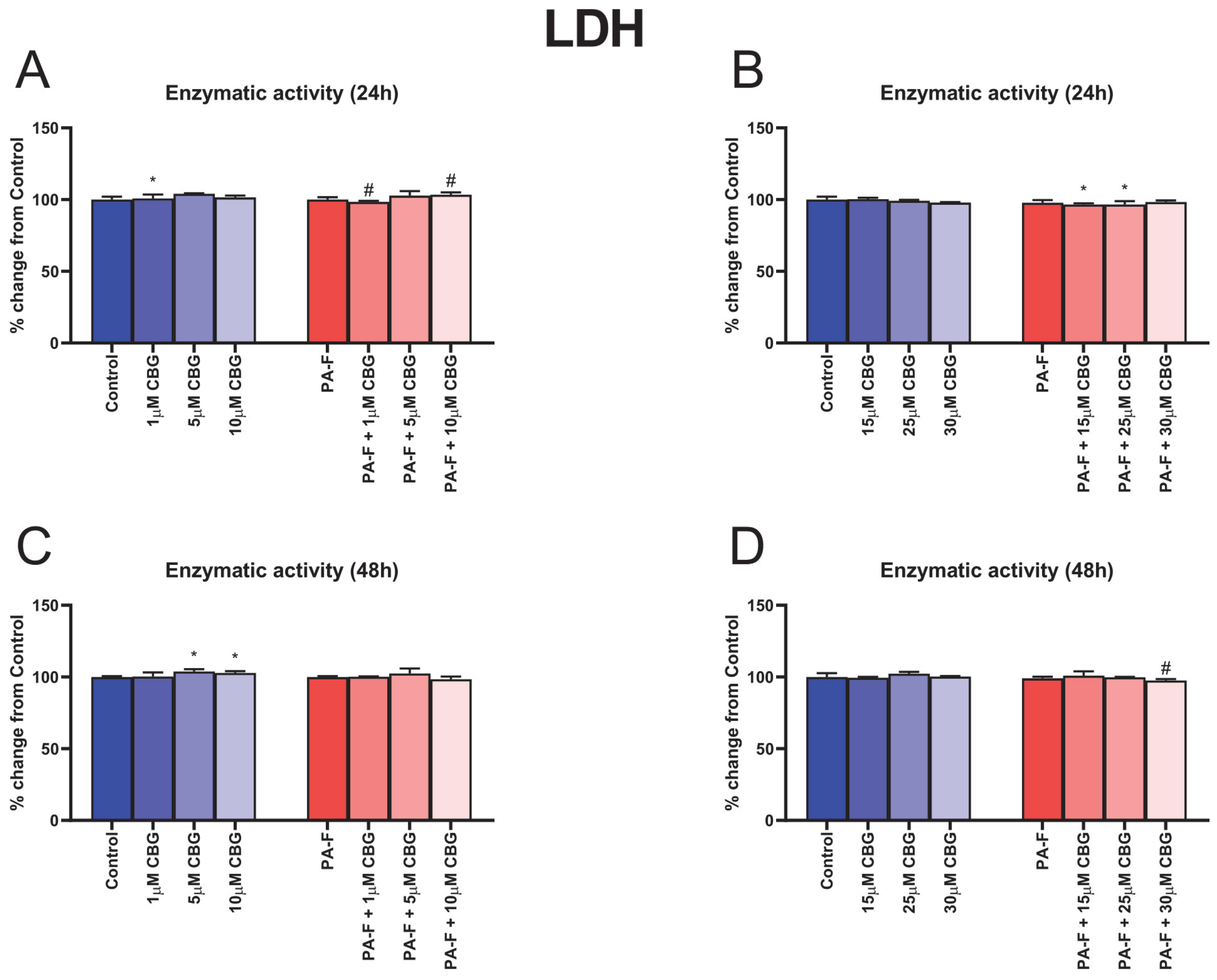
3.1. Concentration-Dependent Influence of Cannabigerol on Cytotoxicity in Post-Incubation Media from Primary Rat Hepatocytes Exposed to Palmitate in Combination with Fructose
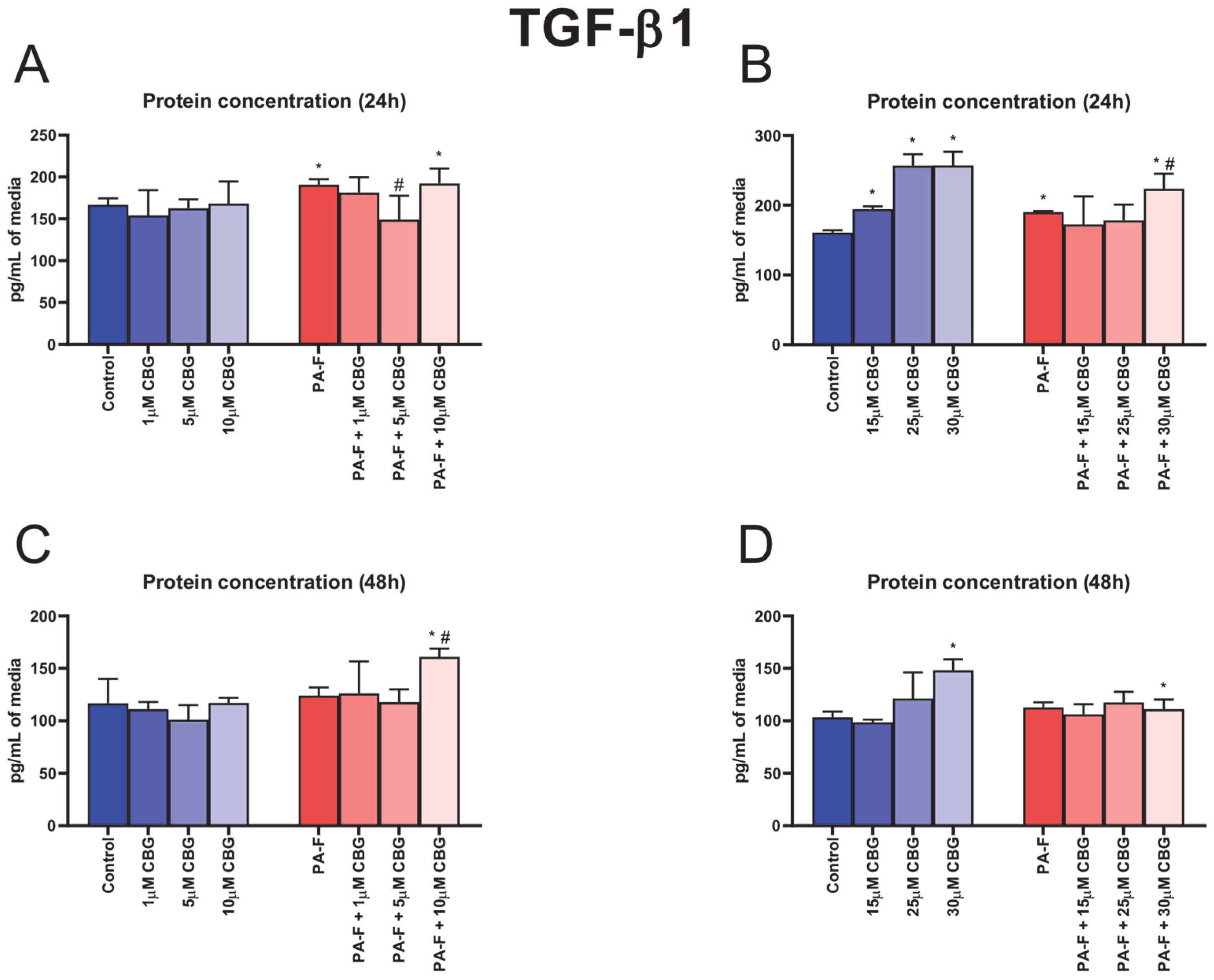
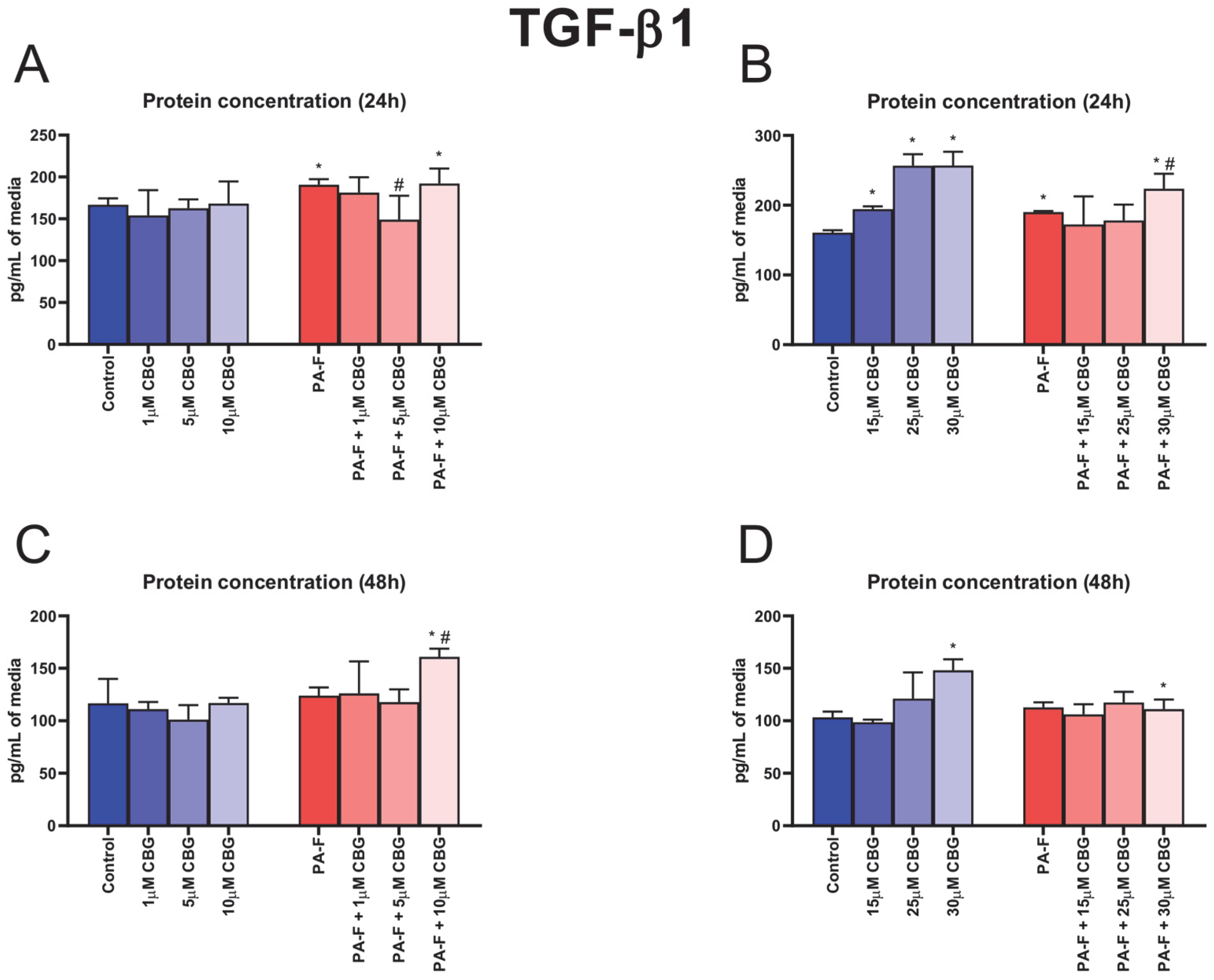
3.2. Concentration-Dependent Influence of Cannabigerol on Transforming Growth Factor Beta 1 Changes in Post-Incubation Media and Primary Rat Hepatocytes Exposed to Palmitate in Combination with Fructose
3.3. Concentration-Dependent Influence of Cannabigerol on the Hydroxyproline Content in Primary Rat Hepatocytes Exposed to Palmitate in Combination with Fructose
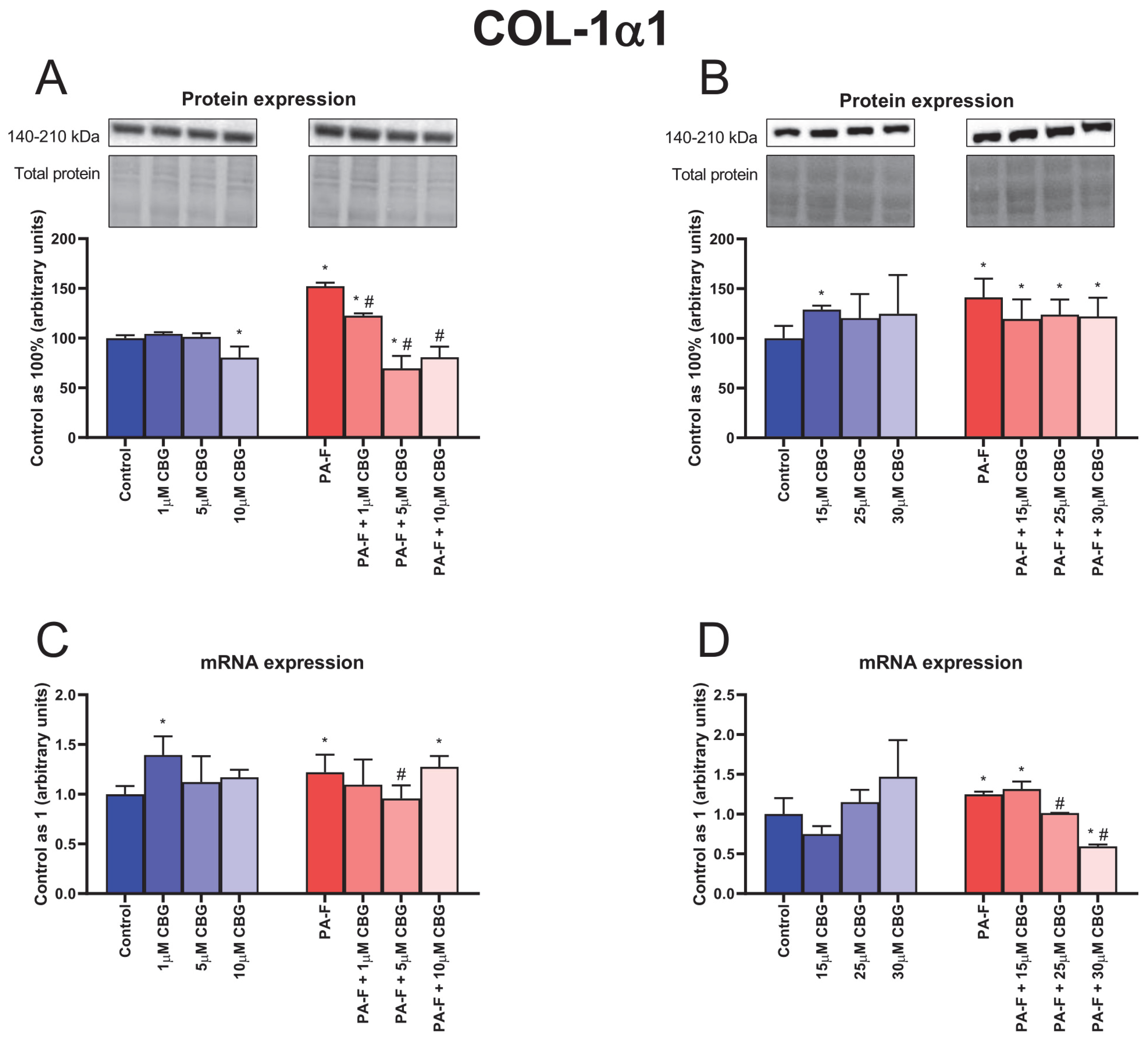
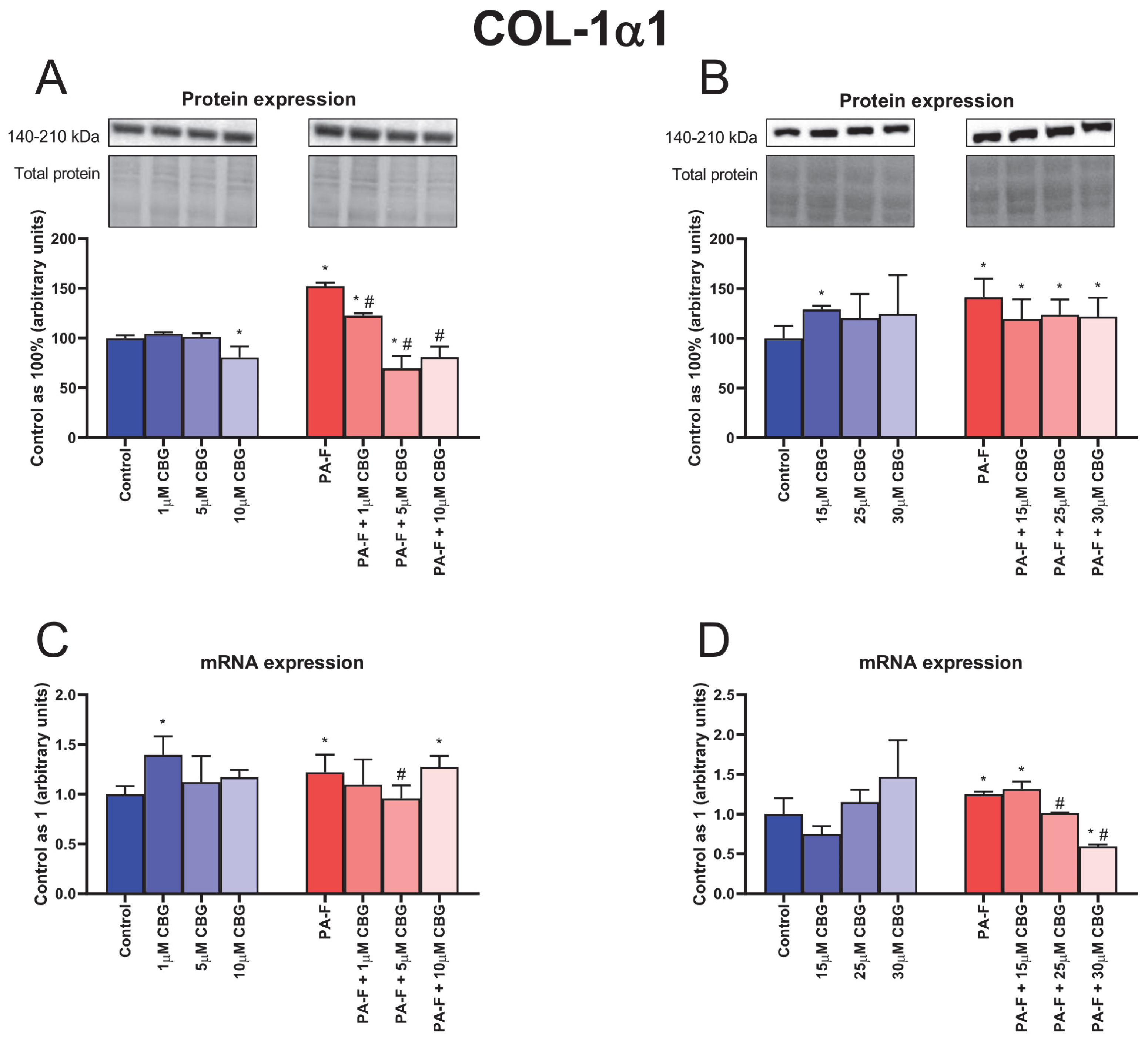
3.4. Concentration-Dependent Influence of Cannabigerol on Collagen Type 1 Alpha 1 Changes in Primary Rat Hepatocytes Exposed to Palmitate in Combination with Fructose
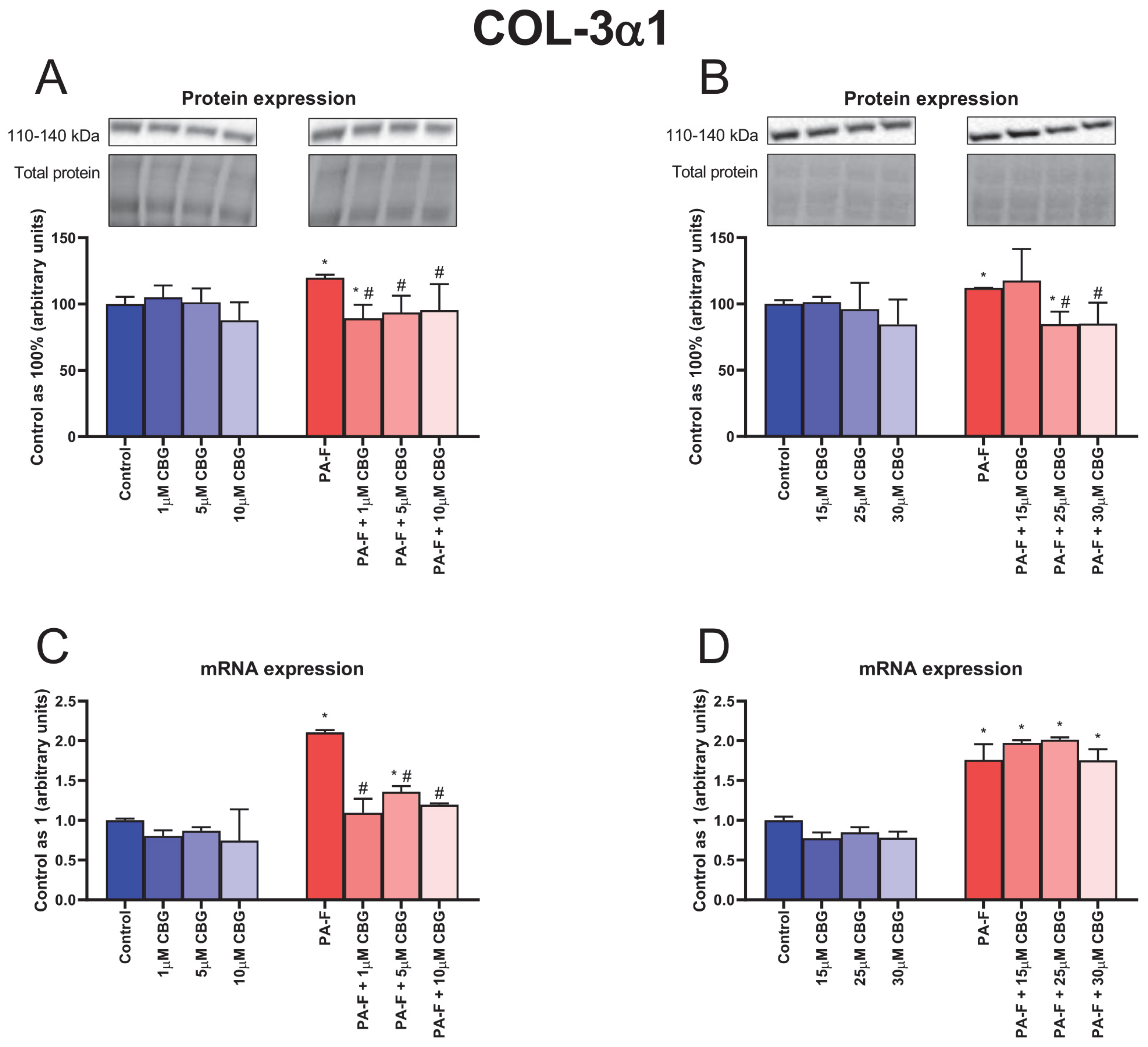
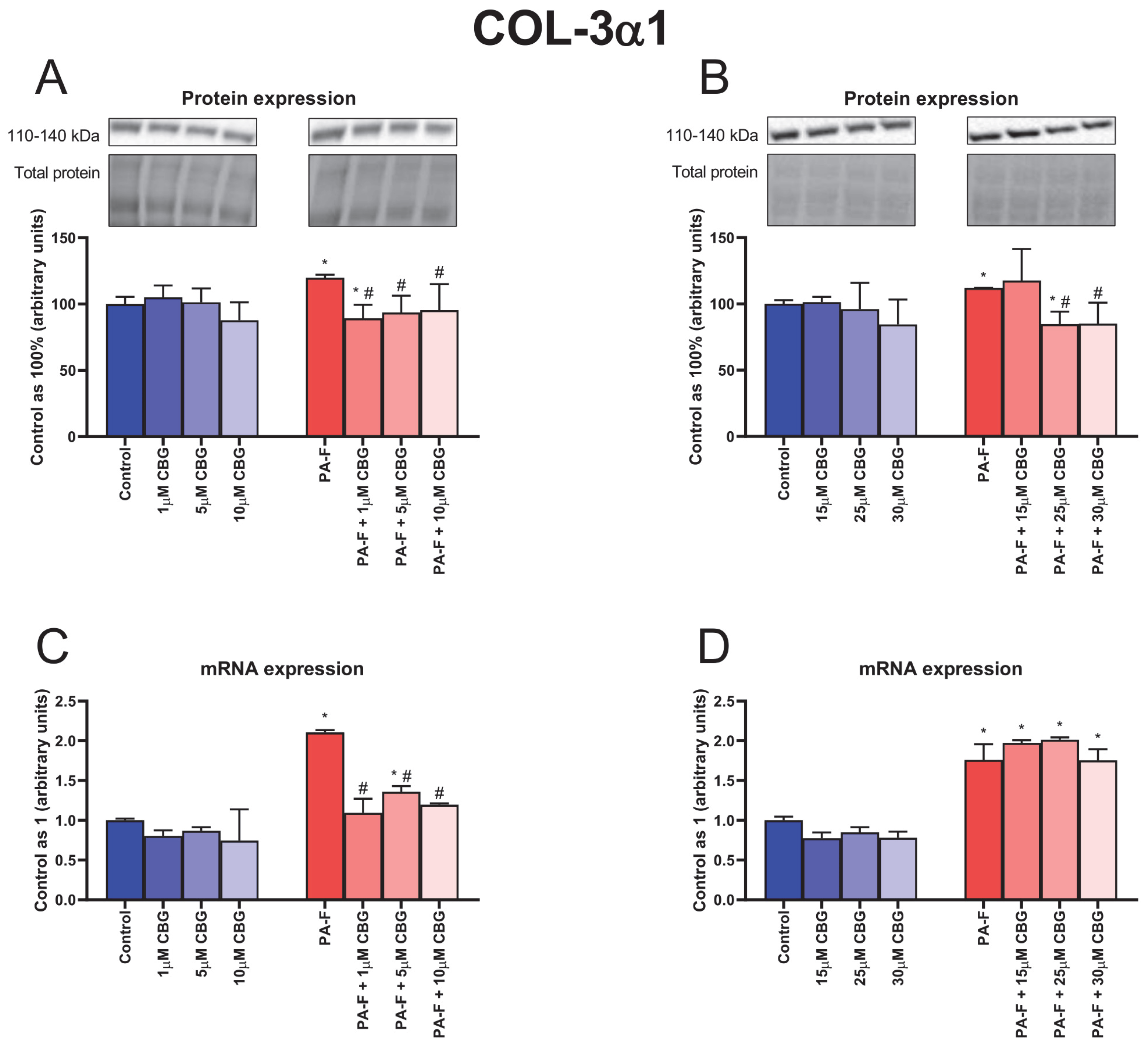
3.5. Concentration-Dependent Influence of Cannabigerol on Collagen Type 3 Alpha 1 Changes in Primary Rat Hepatocytes Exposed to Palmitate in Combination with Fructose
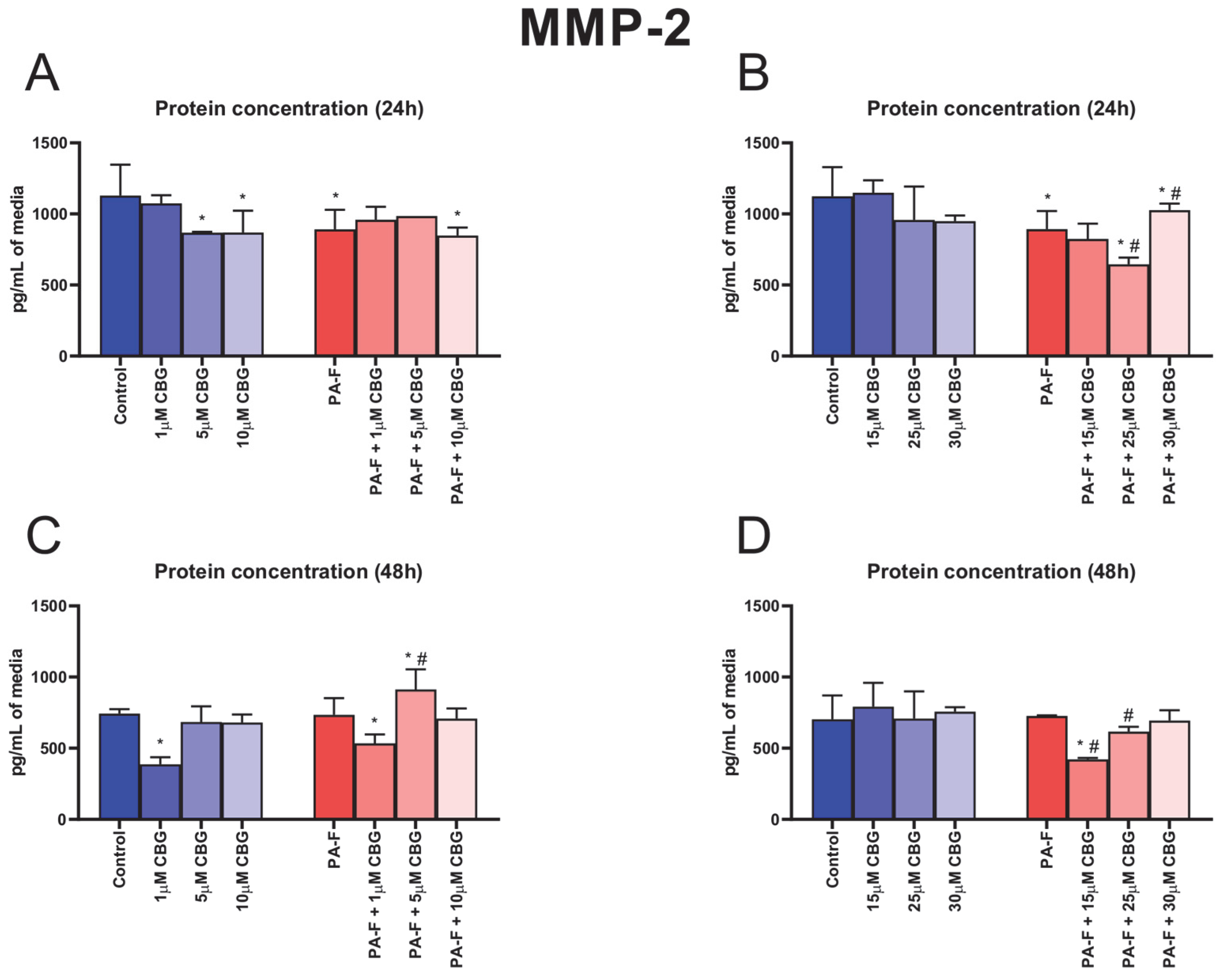
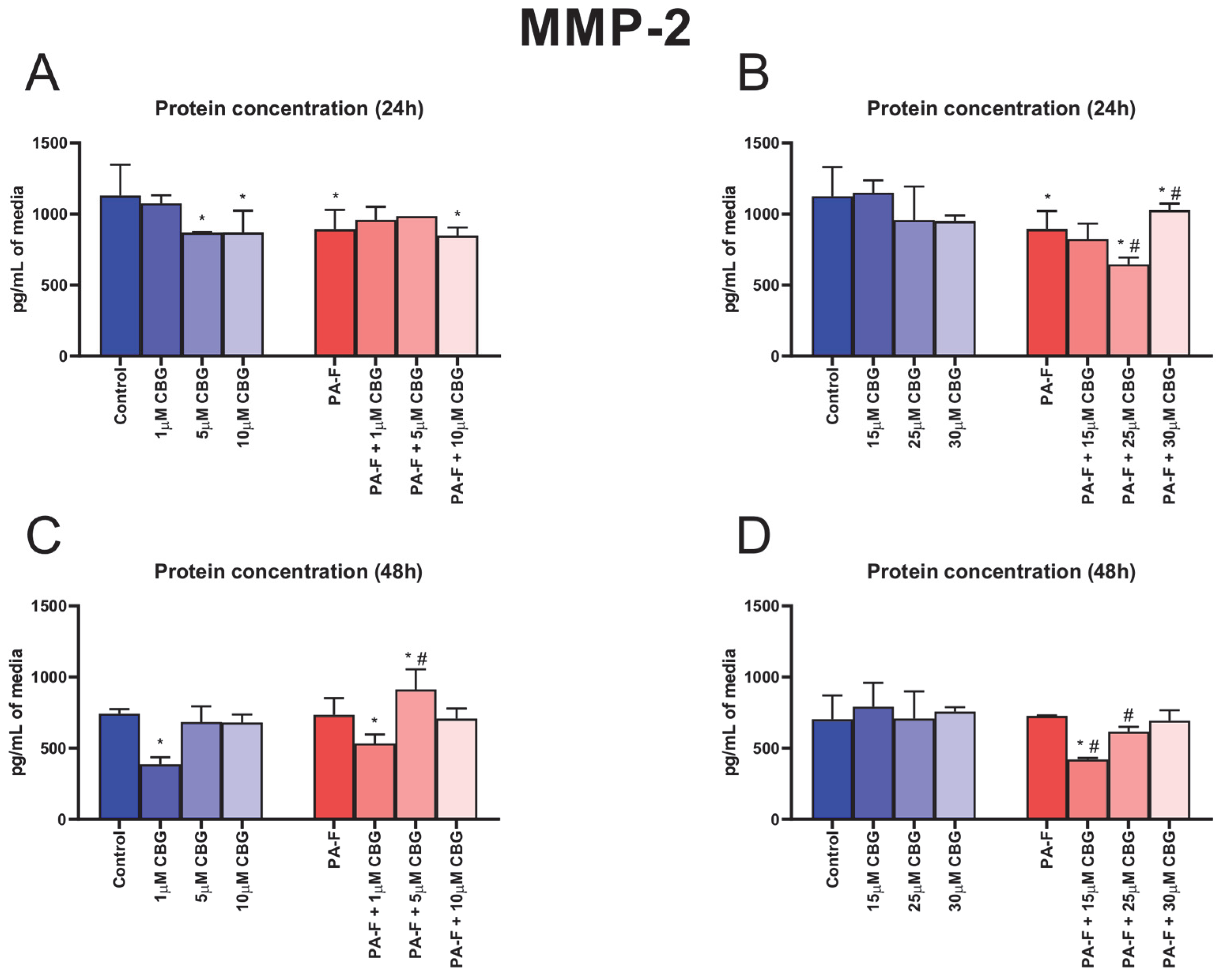
3.6. Concentration-Dependent Influence of Cannabigerol on Matrix Metalloproteinase 2 Changes in Post-Incubation Media and Primary Rat Hepatocytes Exposed to Palmitate in Combination with Fructose
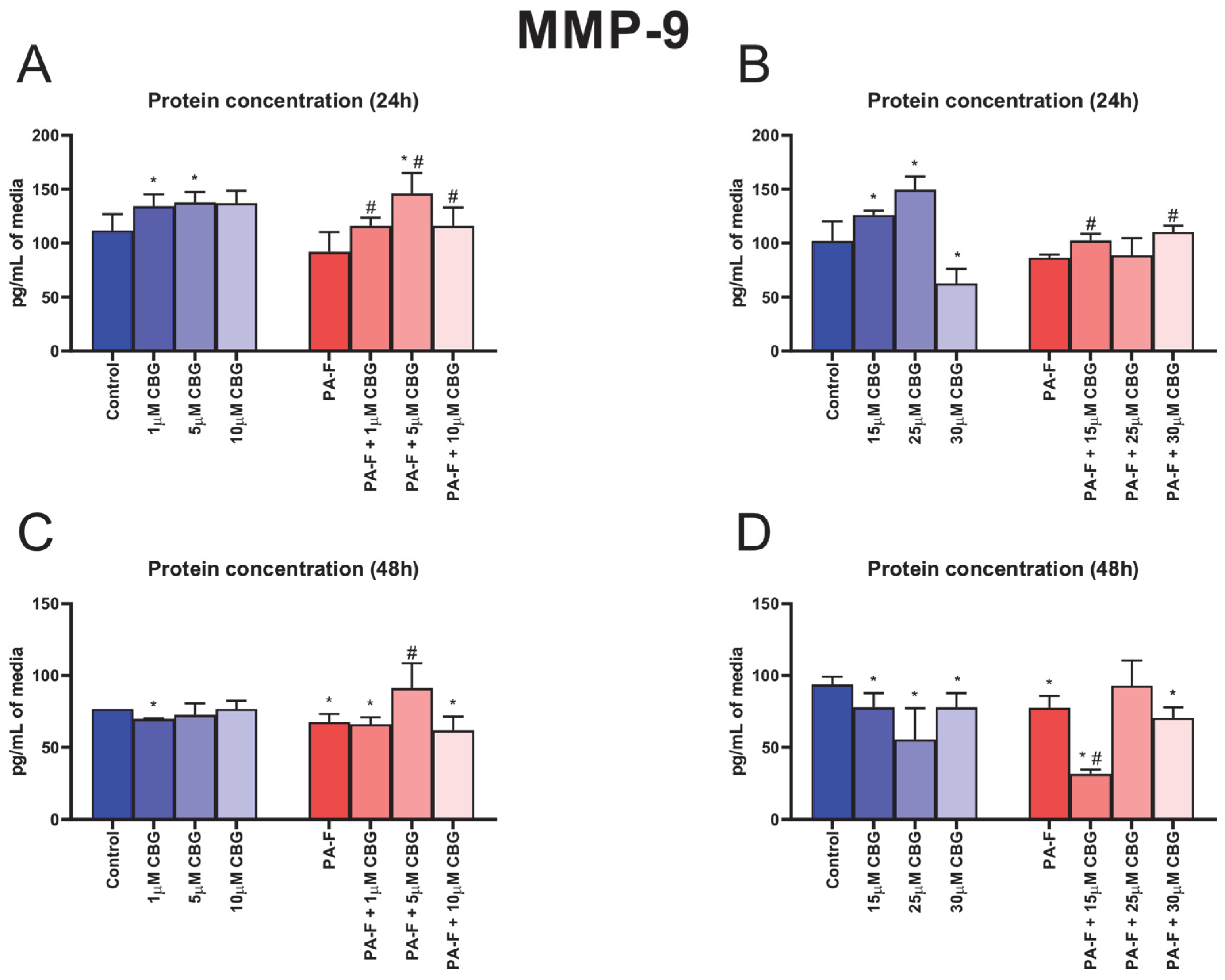
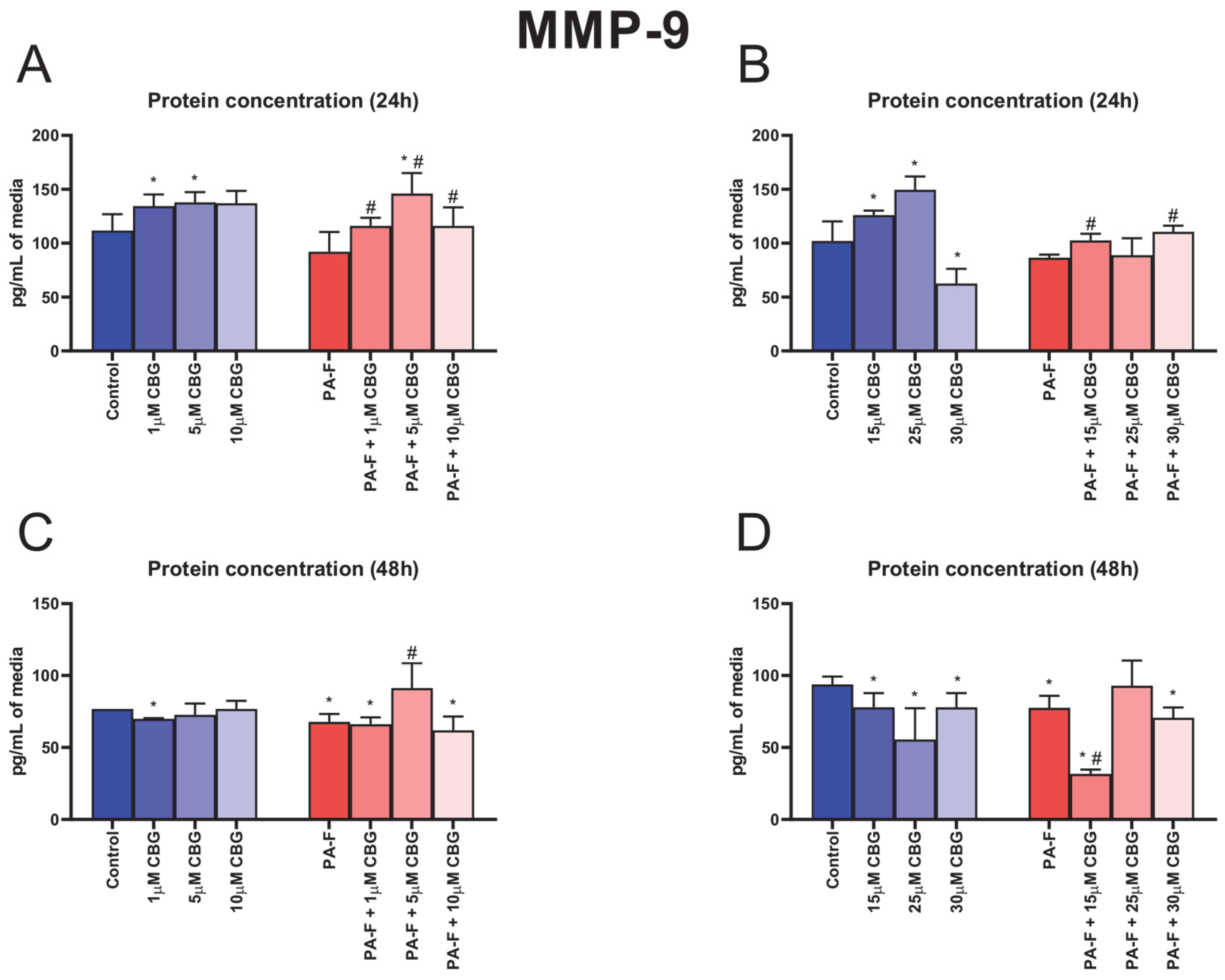
3.7. Concentration-Dependent Influence of Cannabigerol on Matrix Metalloproteinase 9 Changes in Post-Incubation Media and Primary Rat Hepatocytes Exposed to Palmitate in Combination with Fructose
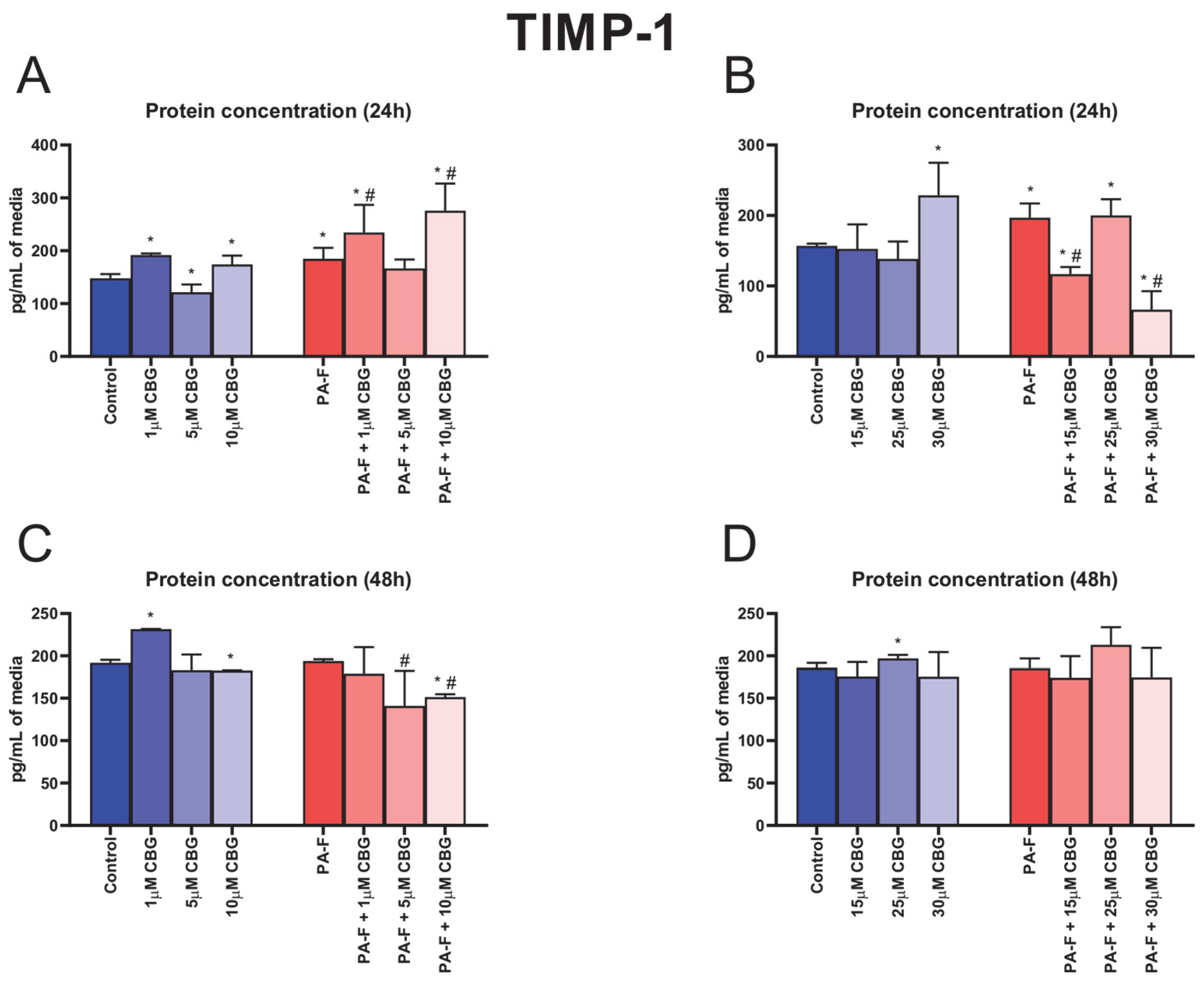
3.8. Concentration-Dependent Influence of Cannabigerol on Tissue Inhibitor of Metalloproteinase 1 Changes in Post-Incubation Media and Primary Rat Hepatocytes Exposed to Palmitate in Combination with Fructose
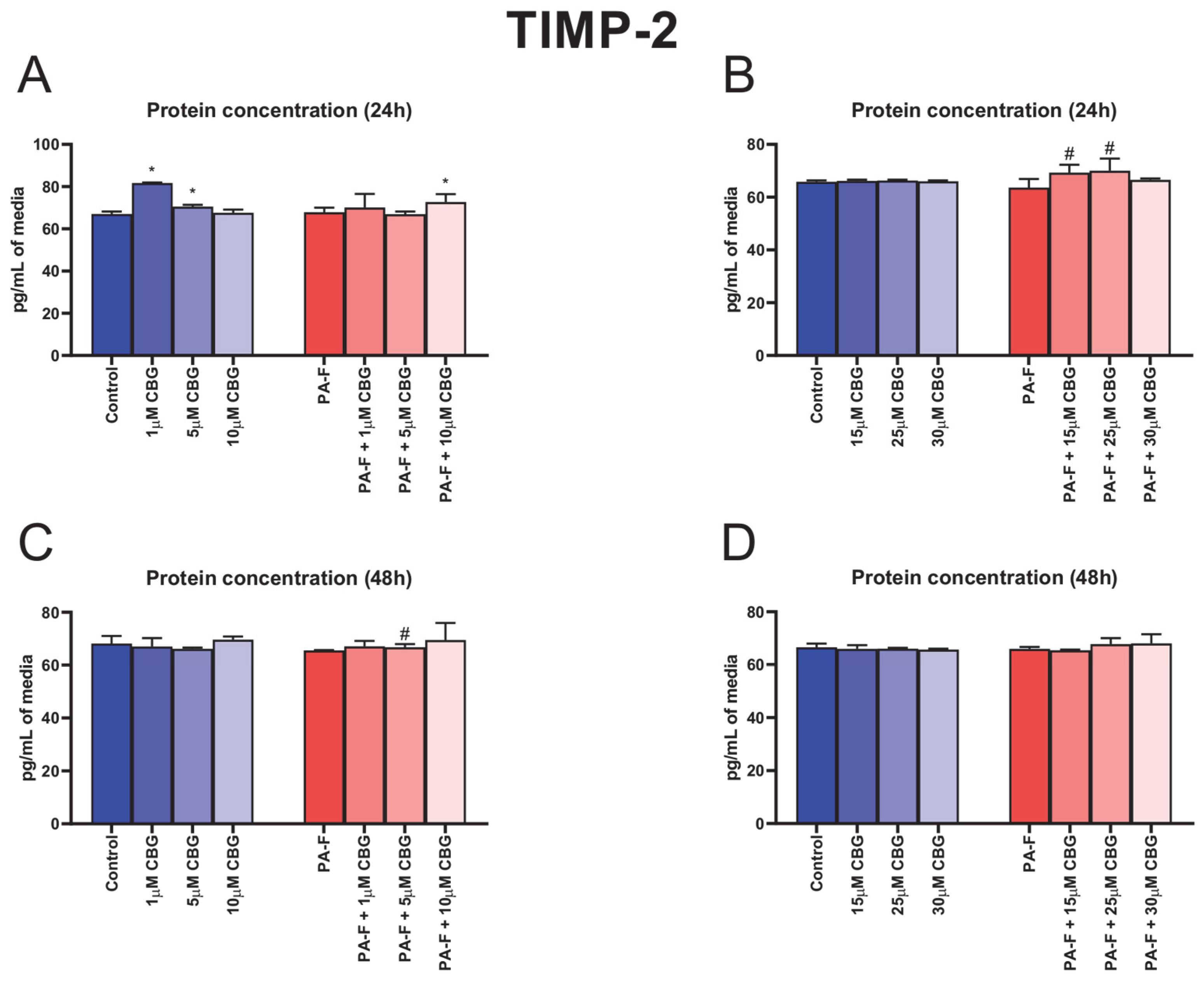
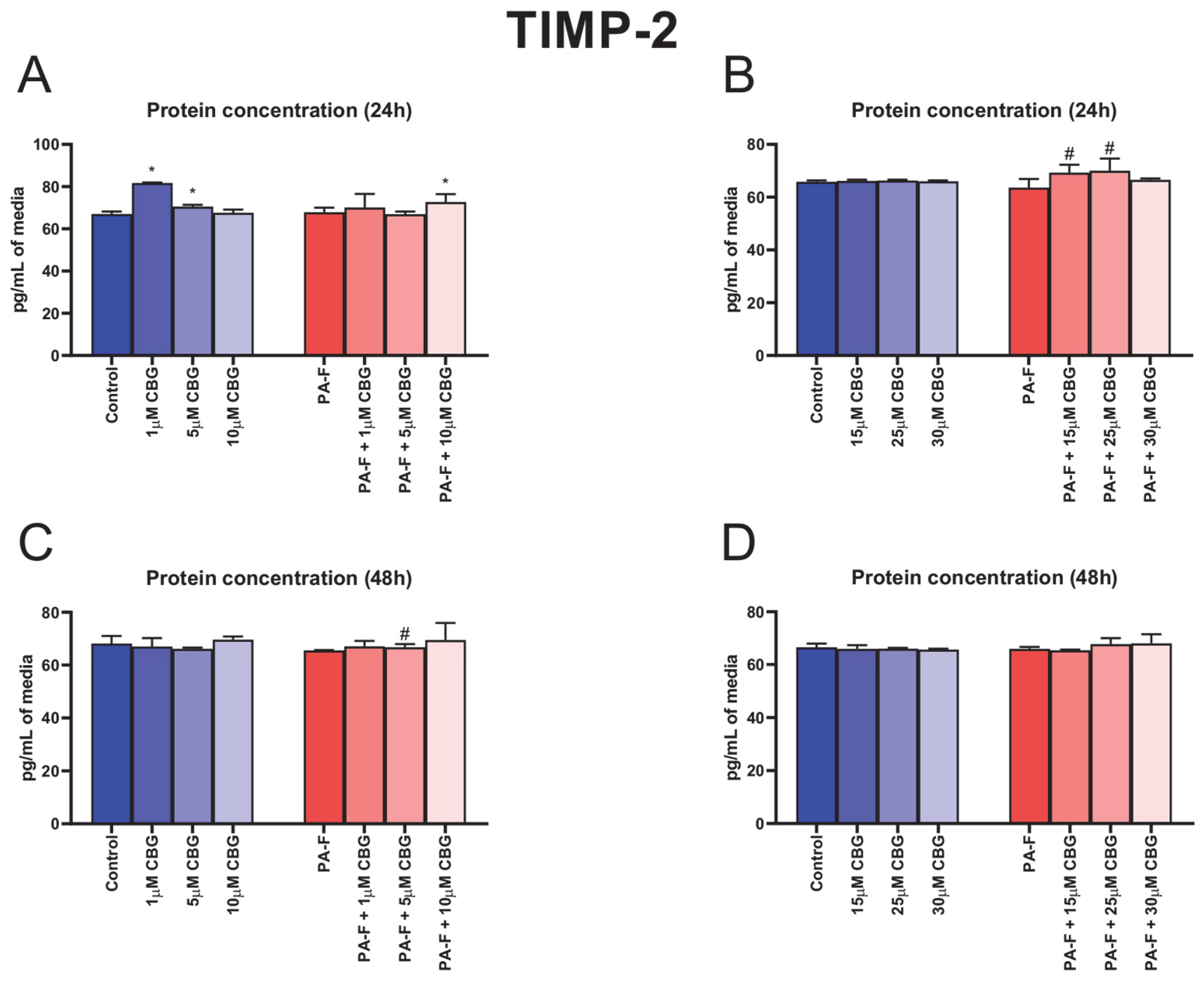
3.9. Concentration-Dependent Influence of Cannabigerol on Tissue Inhibitor of Metalloproteinase 2 Changes in Post-Incubation Media and Primary Rat Hepatocytes Exposed to Palmitate in Combination with Fructose
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karsdal, M.A.; Daniels, S.J.; Nielsen, S.H.; Bager, C.; Rasmussen, D.G.K.; Loomba, R.; Surabattula, R.; Villesen, I.F.; Luo, Y.; Shevell, D.; et al. Collagen biology and non-invasive biomarkers of liver fibrosis. Liver Int. 2020, 40, 736–750. [Google Scholar] [CrossRef] [PubMed]
- Elpek, G.Ö. Cellular and molecular mechanisms in the pathogenesis of liver fibrosis: An update. World J. Gastroenterol. 2014, 20, 7260–7276. [Google Scholar] [CrossRef] [PubMed]
- Acharya, P.; Chouhan, K.; Weiskirchen, S.; Weiskirchen, R. Cellular Mechanisms of Liver Fibrosis. Front. Pharmacol. 2021, 12, 671640. [Google Scholar] [CrossRef]
- Zoubek, M.E.; Trautwein, C.; Strnad, P. Reversal of liver fibrosis: From fiction to reality. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 129–141. [Google Scholar] [CrossRef]
- Zbodakova, O.; Chalupsky, K.; Tureckova, J.; Sedlacek, R. Metalloproteinases in liver fibrosis: Current insights. Met. Med. 2017, ume 4, 25–35. [Google Scholar] [CrossRef]
- Tan, Z.; Sun, H.; Xue, T.; Gan, C.; Liu, H.; Xie, Y.; Yao, Y.; Ye, T. Liver Fibrosis: Therapeutic Targets and Advances in Drug Therapy. Front. Cell Dev. Biol. 2021, 9, 730176. [Google Scholar] [CrossRef] [PubMed]
- Wobser, H.; Dorn, C.; Weiss, T.S.; Amann, T.; Bollheimer, C.; Büttner, R.; Schölmerich, J.; Hellerbrand, C. Lipid accumulation in hepatocytes induces fibrogenic activation of hepatic stellate cells. Cell Res. 2009, 19, 996–1005. [Google Scholar] [CrossRef]
- Piras, I.S.; Gerhard, G.S.; DiStefano, J.K. Palmitate and Fructose Interact to Induce Human Hepatocytes to Produce Pro-Fibrotic Transcriptional Responses in Hepatic Stellate Cells Exposed to Conditioned Media. Cell. Physiol. Biochem. 2020, 54, 1068–1082. [Google Scholar] [CrossRef]
- Borrelli, F.; Fasolino, I.; Romano, B.; Capasso, R.; Maiello, F.; Coppola, D.; Orlando, P.; Battista, G.; Pagano, E.; Di Marzo, V.; et al. Beneficial effect of the non-psychotropic plant cannabinoid cannabigerol on experimental inflammatory bowel disease. Biochem. Pharmacol. 2013, 85, 1306–1316. [Google Scholar] [CrossRef]
- Aljobaily, N.; Krutsinger, K.; Viereckl, M.J.; Joly, R.; Menlove, B.; Cone, B.; Suppes, A.; Han, Y. Low-Dose Administration of Cannabigerol Attenuates Inflammation and Fibrosis Associated with Methionine/Choline Deficient Diet-Induced NASH Model via Modulation of Cannabinoid Receptor. Nutrients 2022, 15, 178. [Google Scholar] [CrossRef]
- Nachnani, R.; Raup-Konsavage, W.M.; Vrana, K.E. The Pharmacological Case for Cannabigerol. Experiment 2020, 376, 204–212. [Google Scholar] [CrossRef]
- Duarte, S.; Baber, J.; Fujii, T.; Coito, A.J. Matrix metalloproteinases in liver injury, repair and fibrosis. Matrix Biol. 2015, 44–46, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Julien, B.; Grenard, P.; Teixeira-Clerc, F.; Van Nhieu, J.T.; Li, L.; Karsak, M.; Zimmer, A.; Mallat, A.; Lotersztajn, S. Antifibrogenic role of the cannabinoid receptor CB2 in the liver. Gastroenterology 2005, 128, 742–755. [Google Scholar] [CrossRef]
- Seglen, P.O. Chapter 4 Preparation of Isolated Rat Liver Cells. Methods Cell Biol. 1976, 13, 29–83. [Google Scholar] [CrossRef] [PubMed]
- Charni-Natan, M.; Goldstein, I. Protocol for Primary Mouse Hepatocyte Isolation. STAR Protoc. 2020, 1, 100086. [Google Scholar] [CrossRef] [PubMed]
- Lah, T.T.; Novak, M.; Pena Almidon, M.A.; Marinelli, O.; Žvar Baškovič, B.; Majc, B.; Mlinar, M.; Bošnjak, R.; Breznik, B.; Zomer, R.; et al. Cannabigerol Is a Potential Therapeutic Agent in a Novel Combined Therapy for Glioblastoma. Cells 2021, 10, 340. [Google Scholar] [CrossRef]
- Díaz-Alonso, J.; Paraíso-Luna, J.; Navarrete, C.; Del Río, C.; Cantarero, I.; Palomares, B.; Aguareles, J.; Fernández-Ruiz, J.; Bellido, M.L.; Pollastro, F.; et al. VCE-003.2, a novel cannabigerol derivative, enhances neuronal progenitor cell survival and alleviates symptomatology in murine models of Huntington’s disease. Sci. Rep. 2016, 6, 29789. [Google Scholar] [CrossRef]
- Gomez-Torres, O.; Amatya, S.; Kamberov, L.; Dhaibar, H.A.; Khanna, P.; Rom, O.; Yurdagul, A.; Orr, A.W.; Nunez, K.; Thevenot, P.; et al. SLAMF1 is expressed and secreted by hepatocytes and the liver in nonalcoholic fatty liver disease. Am. J. Physiol. Liver Physiol. 2022, 323, G177–G187. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the Lactate Dehydrogenase Assay. Cold Spring Harb. Protoc. 2018, 2018. [Google Scholar] [CrossRef]
- Stone, N.L.; England, T.J.; O’Sullivan, S.E. Protective Effects of Cannabidivarin and Cannabigerol on Cells of the Blood–Brain Barrier Under Ischemic Conditions. Cannabis Cannabinoid Res. 2021, 6, 315–326. [Google Scholar] [CrossRef]
- Muriel, P.; López-Sánchez, P.; Ramos-Tovar, E. Fructose and the Liver. Int. J. Mol. Sci. 2021, 22, 6969. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Ahmad, A. Understanding the mechanism of hepatic fibrosis and potential therapeutic approaches. Saudi J. Gastroenterol. 2012, 18, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Seki, K.; Kitade, M.; Nishimura, N.; Kaji, K.; Asada, K.; Namisaki, T.; Moriya, K.; Kawaratani, H.; Okura, Y.; Takaya, H.; et al. Oral administration of fructose exacerbates liver fibrosis and hepatocarcinogenesis via increased intestinal permeability in a rat steatohepatitis model. Oncotarget 2018, 9, 28638–28651. [Google Scholar] [CrossRef] [PubMed]
- Al Fadel, F.; Fayyaz, S.; Japtok, L.; Kleuser, B. Involvement of Sphingosine 1-Phosphate in Palmitate-Induced Non-Alcoholic Fatty Liver Disease. Cell. Physiol. Biochem. 2016, 40, 1637–1645. [Google Scholar] [CrossRef]
- Lee, H.-S.; Shun, C.-T.; Chiou, L.-L.; Chen, C.-H.; Huang, G.-T.; Sheu, J.-C. Hydroxyproline content of needle biopsies as an objective measure of liver fibrosis: Emphasis on sampling variability. J. Gastroenterol. Hepatol. 2005, 20, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liu, C.; Hong, S.; Min, J.; Yang, Q.; Hu, M.; Zhao, Y.; Hong, L. Excess mechanical stress and hydrogen peroxide remodel extracellular matrix of cultured human uterosacral ligament fibroblasts by disturbing the balance of MMPs/TIMPs via the regulation of TGF-β1 signaling pathway. Mol. Med. Rep. 2016, 15, 423–430. [Google Scholar] [CrossRef]
- Hayashi, M.; Nomoto, S.; Hishida, M.; Inokawa, Y.; Kanda, M.; Okamura, Y.; Nishikawa, Y.; Tanaka, C.; Kobayashi, D.; Yamada, S.; et al. Identification of the collagen type 1 alpha 1 gene (COL1A1) as a candidate survival-related factor associated with hepatocellular carcinoma. BMC Cancer 2014, 14, 108. [Google Scholar] [CrossRef]
- Brierley, D.I.; Samuels, J.; Duncan, M.; Whalley, B.J.; Williams, C.M. Cannabigerol is a novel, well-tolerated appetite stimulant in pre-satiated rats. Psychopharmacol. 2016, 233, 3603–3613. [Google Scholar] [CrossRef]
- Hinz, B.; Lagares, D. Evasion of apoptosis by myofibroblasts: A hallmark of fibrotic diseases. Nat. Rev. Rheumatol. 2019, 16, 11–31. [Google Scholar] [CrossRef]
- Roeb, E. Matrix metalloproteinases and liver fibrosis (translational aspects). Matrix Biol. 2018, 68–69, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Han, X.; Yin, L.; Xu, L.; Qi, Y.; Xu, Y.; Sun, H.; Lin, Y.; Liu, K.; Peng, J. Potent effects of dioscin against liver fibrosis. Sci. Rep. 2015, 5, 9713. [Google Scholar] [CrossRef] [PubMed]
- Robert, S.; Gicquel, T.; Bodin, A.; Lagente, V.; Boichot, E. Characterization of the MMP/TIMP Imbalance and Collagen Production Induced by IL-1β or TNF-α Release from Human Hepatic Stellate Cells. PLoS ONE 2016, 11, e0153118. [Google Scholar] [CrossRef] [PubMed]
- Prystupa, A.; Boguszewska-Czubara, A.; Bojarska-Junak, A.; Toruń-Jurkowska, A.; Roliński, J.; Załuska, W. Activity of MMP-2, MMP-8 and MMP-9 in serum as a marker of progression of alcoholic liver disease in people from Lublin Region, eastern Poland. J. Neurol. Sci. 2015, 22, 325–328. [Google Scholar] [CrossRef]
- Onozuka, I.; Kakinuma, S.; Kamiya, A.; Miyoshi, M.; Sakamoto, N.; Kiyohashi, K.; Watanabe, T.; Funaoka, Y.; Ueyama, M.; Nakagawa, M.; et al. Cholestatic liver fibrosis and toxin-induced fibrosis are exacerbated in matrix metalloproteinase-2 deficient mice. Biochem. Biophys. Res. Commun. 2011, 406, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Radbill, B.D.; Gupta, R.; Ramirez, M.C.M.; DiFeo, A.; Martignetti, J.A.; Alvarez, C.E.; Friedman, S.L.; Narla, G.; Vrabie, R.; Bowles, R.; et al. Loss of Matrix Metalloproteinase-2 Amplifies Murine Toxin-Induced Liver Fibrosis by Upregulating Collagen I Expression. Dig. Dis. Sci. 2010, 56, 406–416. [Google Scholar] [CrossRef]
- Takahara, T.; Furui, K.; Yata, Y.; Jin, B.; Zhang, L.P.; Nambu, S.; Sato, H.; Seiki, M.; Watanabe, A. Dual expression of matrix metalloproteinase-2 and membrane-type 1-matrix metalloproteinase in fibrotic human livers. Hepatology 1997, 26, 1521–1529. [Google Scholar] [CrossRef]
- Hori, T.; Uemoto, S.; Walden, L.B.; Chen, F.; Baine, A.-M.T.; Hata, T.; Kogure, T.; Nguyen, J.H. Matrix metalloproteinase-9 as a therapeutic target for the progression of fulminant liver failure with hepatic encephalopathy: A pilot study in mice. Hepatol. Res. 2013, 44, 651–662. [Google Scholar] [CrossRef]
- Nart, D.; Yaman, B.; Yılmaz, F.; Zeytunlu, M.; Karasu, Z.; Kılıç, M. Expression of matrix metalloproteinase-9 in predicting prognosis of hepatocellular carcinoma after liver transplantation. Liver Transplant. 2010, 16, 621–630. [Google Scholar] [CrossRef]
- Roderfeld, M.; Rath, T.; Lammert, F.; Dierkes, C.; Graf, J.; Roeb, E. Innovative immunohistochemistry identifies MMP-9 expressing macrophages at the invasive front of murine HCC. World J. Hepatol. 2010, 2, 175–179. [Google Scholar] [CrossRef]
- Masuzaki, R.; Kanda, T.; Sasaki, R.; Matsumoto, N.; Ogawa, M.; Matsuoka, S.; Karp, S.J.; Moriyama, M. Noninvasive Assessment of Liver Fibrosis: Current and Future Clinical and Molecular Perspectives. Int. J. Mol. Sci. 2020, 21, 4906. [Google Scholar] [CrossRef] [PubMed]
- Kossakowska, A.E.; Edwards, D.R.; Lee, S.S.; Urbanski, L.S.; Stabbler, A.L.; Zhang, C.-L.; Phillips, B.W.; Zhang, Y.; Urbanski, S.J. Altered Balance Between Matrix Metalloproteinases and Their Inhibitors in Experimental Biliary Fibrosis. Am. J. Pathol. 1998, 153, 1895–1902. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Protein | Source (Clone Number) | Dilution | Catalog Number | Manufacturer |
|---|---|---|---|---|
| TGF-β1 | rabbit (monoclonal) | 1:200 | MA5-15065 | Thermo Fisher Scientific, Inc., MA, USA |
| MMP-2 | mouse (monoclonal) | 1:500 | sc-13595 | Santa Cruz Biotechnology, Inc., TX, USA |
| MMP-9 | mouse (monoclonal) | 1:500 | sc-393859 | Santa Cruz Biotechnology, Inc., TX, USA |
| TIMP-1 | rabbit (polyclonal) | 1:200 | PA5-9959 | Thermo Fisher Scientific, Inc., MA, USA |
| TIMP-2 | mouse (monoclonal) | 1:100 | sc-365671 | Santa Cruz Biotechnology, Inc., TX, USA |
| COL-1α1 | mouse (monoclonal) | 1:200 | sc-293182 | Santa Cruz Biotechnology, Inc., TX, USA |
| COL-3α1 | mouse (monoclonal) | 1:100 | sc-514601 | Santa Cruz Biotechnology, Inc., TX, USA |
| Target Gene | Forward Primer Sequences (5′→3′) | Reverse Primer Sequence (5′→3′) | Annealing Temperature |
|---|---|---|---|
| TGF-β1 | CTTCAGCTCCACAGAGAAGAACTGC | CACGATCATGTTGGACAACTGCTCC | 64 °C |
| MMP-2 | ACCATCGCCCATCATCAAGT | CGAGCAAAAGCATCATCCAC | 60 °C |
| MMP-9 | CTTGAAGTCTCAGAAGGTGG | AACAAGAAAGGACAGCGTGC | 60 °C |
| TIMP-1 | GGGCTAAATTCATGGGTTCC | GTTCAGGCTTCAGCTTTTGC | 64 °C |
| TIMP-2 | CCAGGTCCTTTTCATCCTGA | CTGGGACTCCTAGGCAAATG | 64 °C |
| COL-1α1 | TGGCCAAGAAGACATCCCTGAAGT | ACATCAGGTTTCCACGTCTCACCA | 62 °C |
| COL-3α1 | AGGCCAATGGCAATGTAAAG | TGTCTTGCTCCATTCACCAG | 64 °C |
| GAPDH | TGCACCACCAACTGCTTA | GGATGCAGGGATGATGTTC | 62 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sztolsztener, K.; Konstantynowicz-Nowicka, K.; Pędzińska-Betiuk, A.; Chabowski, A. Concentration-Dependent Attenuation of Pro-Fibrotic Responses after Cannabigerol Exposure in Primary Rat Hepatocytes Cultured in Palmitate and Fructose Media. Cells 2023, 12, 2243. https://doi.org/10.3390/cells12182243
Sztolsztener K, Konstantynowicz-Nowicka K, Pędzińska-Betiuk A, Chabowski A. Concentration-Dependent Attenuation of Pro-Fibrotic Responses after Cannabigerol Exposure in Primary Rat Hepatocytes Cultured in Palmitate and Fructose Media. Cells. 2023; 12(18):2243. https://doi.org/10.3390/cells12182243
Chicago/Turabian StyleSztolsztener, Klaudia, Karolina Konstantynowicz-Nowicka, Anna Pędzińska-Betiuk, and Adrian Chabowski. 2023. "Concentration-Dependent Attenuation of Pro-Fibrotic Responses after Cannabigerol Exposure in Primary Rat Hepatocytes Cultured in Palmitate and Fructose Media" Cells 12, no. 18: 2243. https://doi.org/10.3390/cells12182243
APA StyleSztolsztener, K., Konstantynowicz-Nowicka, K., Pędzińska-Betiuk, A., & Chabowski, A. (2023). Concentration-Dependent Attenuation of Pro-Fibrotic Responses after Cannabigerol Exposure in Primary Rat Hepatocytes Cultured in Palmitate and Fructose Media. Cells, 12(18), 2243. https://doi.org/10.3390/cells12182243