


Neural Progenitor Cells and the Hypothalamus


{kind=link}
{kind=link}
Abstract
1. Introduction
2. NPCs in the Embryonic and Postnatal/Adult Brain
2.1. NPCs in the Embryonic Brain
2.2. NPCs in the Postnatal/Adult Brain
2.3. The Secretome of NPCs
EVs and NPCs
3. Regulation of Adult Neurogenesis
3.1. Regulation of Adult Neurogenesis by Local and/or Distal Neural Circuits
3.1.1. Neurotransmitters
3.1.2. Neuropeptides
3.2. The Effects of Glucocorticoids (GCs) on Adult Neurogenesis
4. Markers of Adult Neurogenesis
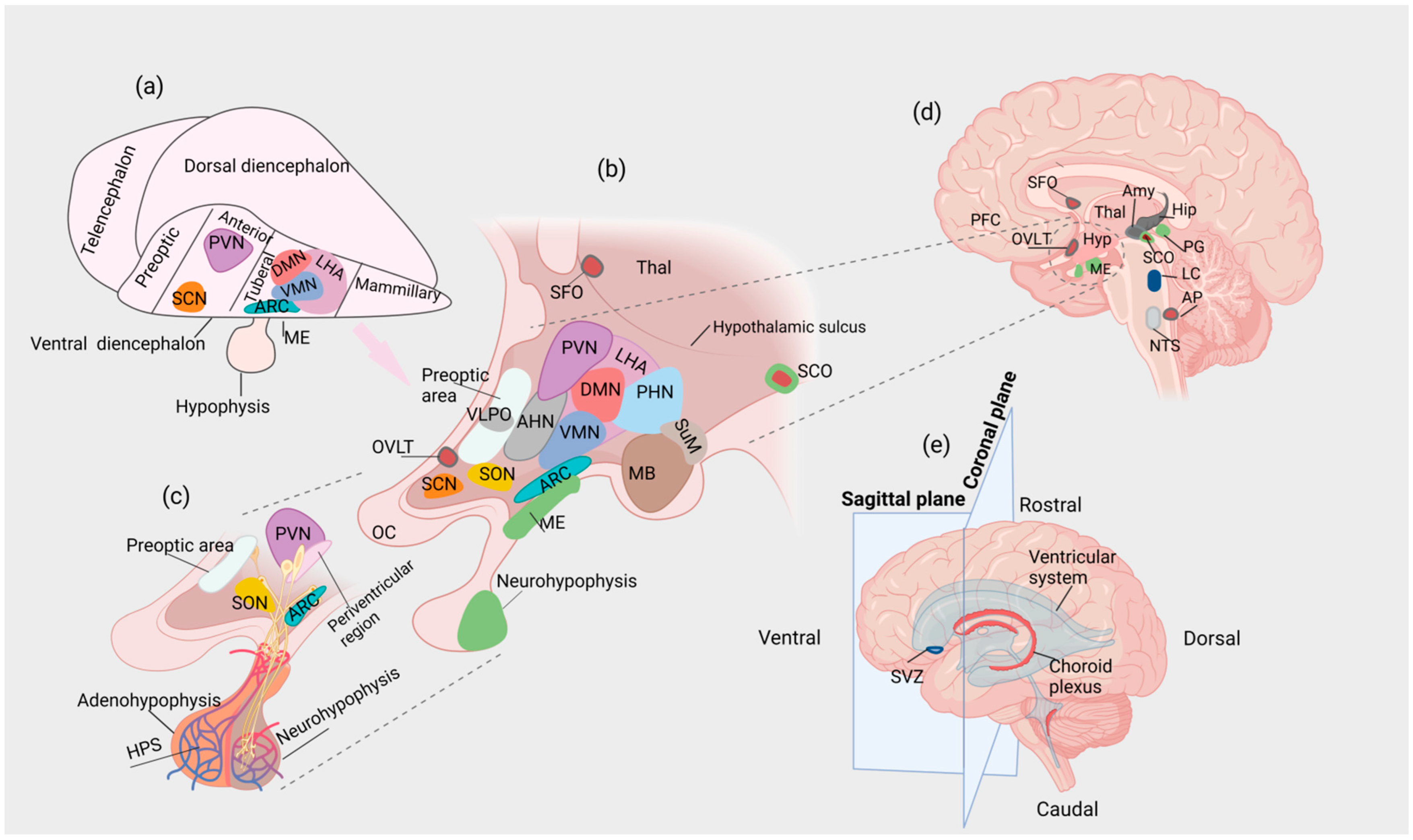
5. The Hypothalamus
5.1. Hypothalamic Nuclei in the MBH
5.1.1. The ARC
5.1.2. The VMN
5.1.3. The DMN
5.2. The SCN
5.2.1. Cellular Time-keeping and Biological Rhythms
5.2.2. Structural and Functional Organization of the SCN
5.3. The PVN
The Entrainment of CRHPVN Neurons by the SCN
5.4. The Hypothalamus as a Regulator of Adult Neurogenesis
6. The CVOs
6.1. The SFO
6.2. The OVLT
6.3. The AP
6.4. CVOs and NPCs
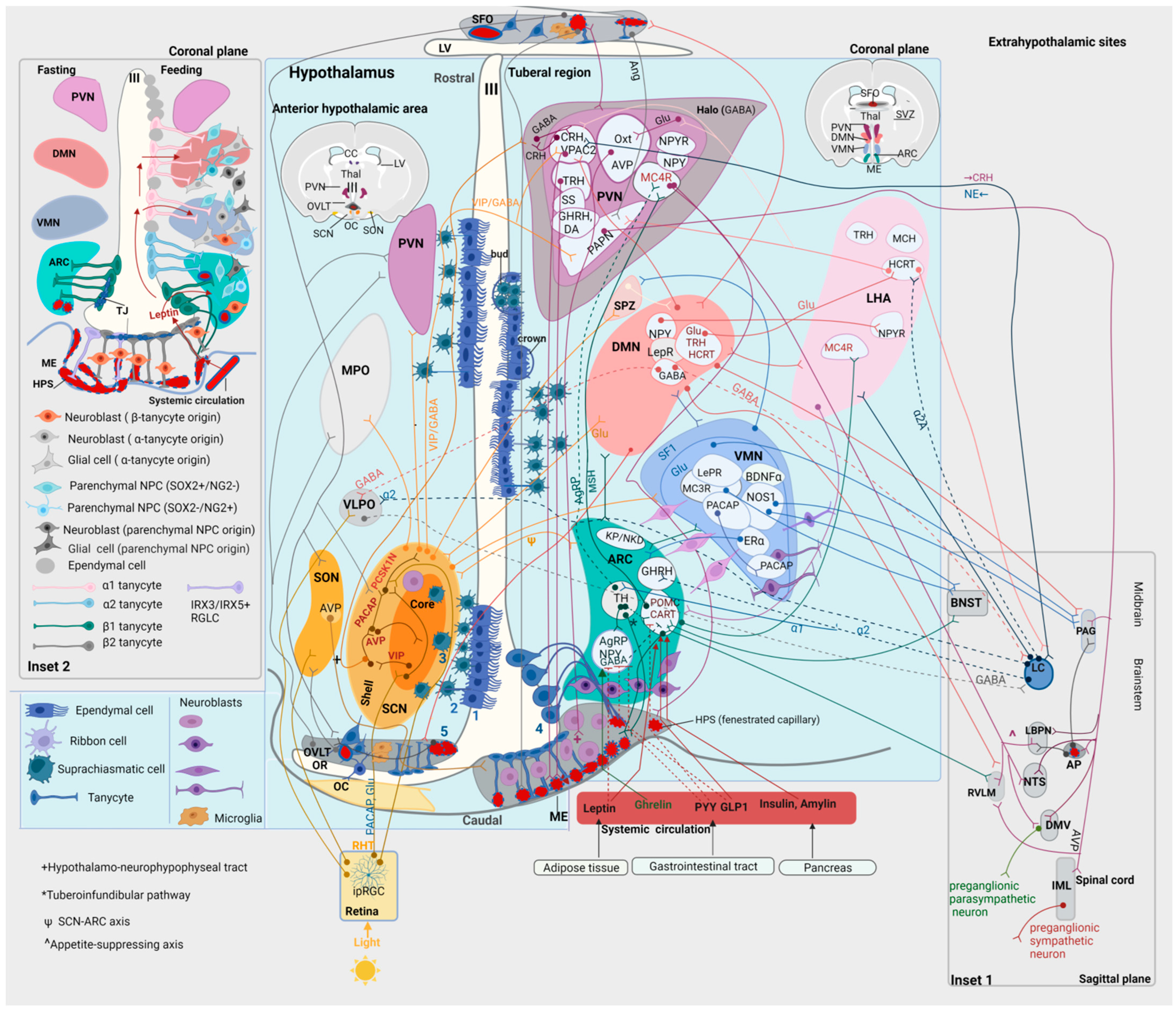
7. The Hypothalamic Neurogenic Niche
7.1. Animal Models
7.1.1. Tanycytes
Tanycytes as NPCs
Τanycytes as Sensors and Integrators of Peripheral Metabolic Signals
Tanycytes as Central Regulators of the Neuroendocrine Secretion
7.2. Humans
7.3. Comparison of the Adult Human and Animal Hypothalamic Neurogenic Niches
7.4. Regulation and Functional Implications of Adult Hypothalamic Neurogenesis
7.4.1. Energy Balance and Metabolism
HFD impairs Neurogenesis in the ARC, similarly to Leptin Deficiency
HFD induces ARC-ME Inflammation, Neuronal Injury and Gliosis
HFD → ↑ NFκB Pathway → MBH Inflammation → Impaired MBH Neurogenesis and NPC Survival → Overeating, Weight Gain, Glucose Intolerance and Hyperinsulinemia
MBH NPC Ablation → Overeating, Weight Gain and Glucose Intolerance
7.4.2. Sleep
↓ Hypothalamic Neurogenesis → Disrupted Sleep Architecture (Aging-reminiscent)
7.4.3. Aging
Aging → MBH Microglial Activation → ↑ NFκB Pathway in Neighboring Cells
Loss of SOX2/BMI1+ Cells in the Wall of the 3rd Ventricle at the Level of MBH drives Aging
Hypothalamic NPCs can form Spheres with Combined Prosurvival and Antidiabetic Effects
7.4.4. Temperature/Heat Acclimation
7.4.5. NE
7.4.6. Transcription Factors of the NFI Family
8. Discussion
9. Outstanding Questions
- What is the secretome of the distinct NPC subpopulations in the adult human hypothalamus? What neuropeptides/receptors do these NPCs express; what is their role?
- What is the role of the PACAP/PAC1 system in hypothalamic neurogenesis?
- What are the content and targets of EVs synthesized and released from NPC subpopulations in the adult human hypothalamus? In the opposite direction, what are the origin and effects of EVs that target hypothalamic NPCs?
- What is the role of ependymal cells, ribbon cells, and buds?
- What is the significance of the morphological similarity of suprachiasmatic cells to ribbon cells? What is the role of suprachiasmatic cells?
- Does neurogenesis occur in the adult human PVN? What are the characteristics and role of this neurogenic subniche?
- What is the role of the OVLT in the hypothalamic neurogenic niche?
- What is the role of glial cells (astrocytes, oligodendrocytes and microglia) in the hypothalamic neurogenic niche?
- What is the molecular mechanism through which HFD induces the activation of the NFκΒ pathway in mouse MBH NPCs?
- What is the molecular mechanism through which activation of the NFκΒ pathway in the mouse hypothalamus induces systemic aging?
- Is the secretome of MBH NPCs responsible for systemic aging in humans as in mice?
- What is the role of biological rhythms and their disruption in adult hypothalamic neurogenesis? What is the role of stress?
- Could brain/hypothalamic organoids/assembloids be utilized to study the human hypothalamic neurogenic niche?
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Martínez-Cerdeño, V.; Noctor, S.C. Neural Progenitor Cell Terminology. Front. Neuroanat. 2018, 12, 104. [Google Scholar] [CrossRef] [PubMed]
- Hassani, S.-N.; Moradi, S.; Taleahmad, S.; Braun, T.; Baharvand, H. Transition of inner cell mass to embryonic stem cells: Mechanisms, facts, and hypotheses. Cell Mol. Life Sci. CMLS 2019, 76, 873–892. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Kimble, J. Asymmetric and symmetric stem-cell divisions in development and cancer. Nature 2006, 441, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Temple, S. The development of neural stem cells. Nature 2001, 414, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Ginhoux, F.; Lim, S.; Hoeffel, G.; Low, D.; Huber, T. Origin and differentiation of microglia. Front. Cell Neurosci. 2013, 7, 45. [Google Scholar] [CrossRef]
- Leathers, T.A.; Rogers, C.D. Time to go: Neural crest cell epithelial-to-mesenchymal transition. Development 2022, 149, dev200712. [Google Scholar] [CrossRef]
- Krispin, S.; Nitzan, E.; Kalcheim, C. The dorsal neural tube: A dynamic setting for cell fate decisions. Dev. Neurobiol. 2010, 70, 796–812. [Google Scholar] [CrossRef]
- Yoo, S.; Blackshaw, S. Regulation and function of neurogenesis in the adult mammalian hypothalamus. Prog. Neurobiol. 2018, 170, 53–66. [Google Scholar] [CrossRef]
- Lim, D.A.; Alvarez-Buylla, A. The Adult Ventricular–Subventricular Zone (V-SVZ) and Olfactory Bulb (OB) Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8, a018820. [Google Scholar] [CrossRef]
- Song, J.; Christian, K.M.; Ming, G.-L.; Song, H. Modification of hippocampal circuitry by adult neurogenesis. Dev. Neurobiol. 2012, 72, 1032–1043. [Google Scholar] [CrossRef]
- Bjornsson, C.S.; Apostolopoulou, M.; Tian, Y.; Temple, S. It takes a village: Constructing the neurogenic niche. Dev. Cell 2015, 32, 435–446. [Google Scholar] [CrossRef]
- Ma, D.K.; A Bonaguidi, M.; Ming, G.-L.; Song, H. Adult neural stem cells in the mammalian central nervous system. Cell Res. 2009, 19, 672–682. [Google Scholar] [CrossRef]
- Daneman, R.; Zhou, L.; Kebede, A.A.; Barres, B.A. Pericytes are required for blood–brain barrier integrity during embryogenesis. Nature 2010, 468, 562–566. [Google Scholar] [CrossRef]
- Dohgu, S.; Takata, F.; Yamauchi, A.; Nakagawa, S.; Egawa, T.; Naito, M.; Tsuruo, T.; Sawada, Y.; Niwa, M.; Kataoka, Y. Brain pericytes contribute to the induction and up-regulation of blood–brain barrier functions through transforming growth factor-beta production. Brain Res. 2005, 1038, 208–215. [Google Scholar] [CrossRef]
- Lehtinen, M.K.; Zappaterra, M.W.; Chen, X.; Yang, Y.J.; Hill, A.D.; Lun, M.; Maynard, T.; Gonzalez, D.; Kim, S.; Ye, P.; et al. The cerebrospinal fluid provides a proliferative niche for neural progenitor cells. Neuron 2011, 69, 893–905. [Google Scholar] [CrossRef]
- Sharif, A.; Fitzsimons, C.P.; Lucassen, P.J. Neurogenesis in the adult hypothalamus: A distinct form of structural plasticity involved in metabolic and circadian regulation, with potential relevance for human pathophysiology. Handb. Clin. Neurol. 2021, 179, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, E.A.B.; Lakshman, N.; Lau, K.S.K.; Morshead, C.M. Regulating Endogenous Neural Stem Cell Activation to Promote Spinal Cord Injury Repair. Cells 2022, 11, 846. [Google Scholar] [CrossRef] [PubMed]
- Lechan, R.M.; Toni, R. Functional Anatomy of the Hypothalamus and Pituitary; Feingold, K.R., Anawalt, B., Blackman, M.R., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Prevot, V.; Dehouck, B.; Sharif, A.; Ciofi, P.; Giacobini, P.; Clasadonte, J. The Versatile Tanycyte: A Hypothalamic Integrator of Reproduction and Energy Metabolism. Endocr. Rev. 2018, 39, 333–368. [Google Scholar] [CrossRef] [PubMed]
- Elshazzly, M.; Lopez, M.J.; Reddy, V.; Caban, O. Embryology, Central Nervous System. In StatPearls; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Noctor, S.C.; Flint, A.C.; Weissman, T.A.; Dammerman, R.S.; Kriegstein, A.R. Neurons derived from radial glial cells establish radial units in neocortex. Nature 2001, 409, 714–720. [Google Scholar] [CrossRef] [PubMed]
- Noctor, S.C.; Flint, A.; Weissman, T.; Wong, W.S.; Clinton, B.K.; Kriegstein, A.R. Dividing precursor cells of the embryonic cortical ventricular zone have morphological and molecular characteristics of radial glia. J. Neurosci. 2002, 22, 3161–3173. [Google Scholar] [CrossRef]
- Redmond, S.A.; Figueres-Oñate, M.; Obernier, K.; Nascimento, M.A.; Parraguez, J.I.; López-Mascaraque, L.; Fuentealba, L.C.; Alvarez-Buylla, A. Development of Ependymal and Postnatal Neural Stem Cells and Their Origin from a Common Embryonic Progenitor. Cell Rep. 2019, 27, 429–441.e3. [Google Scholar] [CrossRef]
- Morest, D.K.; Silver, J. Precursors of neurons, neuroglia, and ependymal cells in the CNS: What are they? Where are they from? How do they get where they are going? Glia 2003, 43, 6–18. [Google Scholar] [CrossRef]
- Götz, M.; Stoykova, A.; Gruss, P. Pax6 controls radial glia differentiation in the cerebral cortex. Neuron 1998, 21, 1031–1044. [Google Scholar] [CrossRef]
- Takahashi, T.; Nowakowski, R.; Caviness, V.S., Jr. The cell cycle of the pseudostratified ventricular epithelium of the embryonic murine cerebral wall. J. Neurosci. 1995, 15, 6046–6057. [Google Scholar] [CrossRef]
- Takahashi, T.; Nowakowski, R.S.; Caviness, V.S., Jr. The leaving or Q fraction of the murine cerebral proliferative epithelium: A general model of neocortical neuronogenesis. J. Neurosci. 1996, 16, 6183–6196. [Google Scholar] [CrossRef] [PubMed]
- Caviness, V.; Goto, T.; Tarui, T.; Takahashi, T.; Bhide, P.; Nowakowski, R. Cell output, cell cycle duration and neuronal specification: A model of integrated mechanisms of the neocortical proliferative process. Cereb. Cortex 2003, 13, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Noctor, S.C.; Martínez-Cerdeño, V.; Ivic, L.; Kriegstein, A.R. Cortical neurons arise in symmetric and asymmetric division zones and migrate through specific phases. Nat. Neurosci. 2004, 7, 136–144. [Google Scholar] [CrossRef]
- Noctor, S.C.; Martínez-Cerdeño, V.; Kriegstein, A.R. Distinct behaviors of neural stem and progenitor cells underlie cortical neurogenesis. J. Comp. Neurol. 2008, 508, 28–44. [Google Scholar] [CrossRef] [PubMed]
- Englund, C.; Fink, A.; Lau, C.; Pham, D.; Daza, R.A.M.; Bulfone, A.; Kowalczyk, T.; Hevner, R.F. Pax6, Tbr2, and Tbr1 are expressed sequentially by radial glia, intermediate progenitor cells, and postmitotic neurons in developing neocortex. J. Neurosci. 2005, 25, 247–251. [Google Scholar] [CrossRef]
- Bello, B.C.; Izergina, N.; Caussinus, E.; Reichert, H. Amplification of neural stem cell proliferation by intermediate progenitor cells in Drosophila brain development. Neural Dev. 2008, 3, 5. [Google Scholar] [CrossRef]
- Lim, D.A.; Alvarez-Buylla, A. Adult neural stem cells stake their ground. Trends Neurosci. 2014, 37, 563–571. [Google Scholar] [CrossRef]
- Seri, B.; García-Verdugo, J.M.; Collado-Morente, L.; McEwen, B.S.; Alvarez-Buylla, A. Cell types, lineage, and architecture of the germinal zone in the adult dentate gyrus. J. Comp. Neurol. 2004, 478, 359–378. [Google Scholar] [CrossRef]
- Rodriguez, M.P.I.; Noctor, S.C.; Muñoz, E.M. Cellular basis of pineal gland development: Emerging role of microglia as phenotype regulator. PLoS ONE 2016, 11, e0167063. [Google Scholar] [CrossRef]
- Deazevedo, L.C.; Moura-Neto, V.; Hedin-Pereira, C.; Lent, R. Cortical radial glial cells in human fetuses: Depth-correlated transformation into astrocytes. J. Neurobiol. 2003, 55, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Rakic, P. Radial glial cells: Scaffolding for cortical development and evolution. In Neuroglia; Oxford University Press: New York, NY, USA, 2004. [Google Scholar] [CrossRef]
- Zecevic, N.; Chen, Y.; Filipovic, R. Contributions of cortical subventricular zone to the development of the human cerebral cortex. J. Comp. Neurol. 2005, 491, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Mathews, K.J.; Allen, K.M.; Boerrigter, D.; Ball, H.; Weickert, C.S.; Double, K.L. Evidence for reduced neurogenesis in the aging human hippocampus despite stable stem cell markers. Aging Cell 2017, 16, 1195–1199. [Google Scholar] [CrossRef] [PubMed]
- Bond, A.M.; Ming, G.; Song, H. Adult mammalian neural stem cells and neurogenesis: Five decades later. Cell Stem. Cell 2015, 17, 385–395. [Google Scholar] [CrossRef]
- Doetsch, F.; Caillé, I.; Lim, D.A.; García-Verdugo, J.M.; Alvarez-Buylla, A. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 1999, 97, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Fuentealba, L.C.; Rompani, S.B.; Parraguez, J.I.; Obernier, K.; Romero, R.; Cepko, C.L.; Alvarez-Buylla, A. Embryonic Origin of Postnatal Neural Stem Cells. Cell 2015, 161, 1644–1655. [Google Scholar] [CrossRef] [PubMed]
- Furutachi, S.; Miya, H.; Watanabe, T.; Kawai, H.; Yamasaki, N.; Harada, Y.; Imayoshi, I.; Nelson, M.; I Nakayama, K.; Hirabayashi, Y.; et al. Slowly dividing neural progenitors are an embryonic origin of adult neural stem cells. Nat. Neurosci. 2015, 18, 657–665. [Google Scholar] [CrossRef]
- Doetsch, F.; García-Verdugo, J.M.; Alvarez-Buylla, A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J. Neurosci. 1997, 17, 5046–5061. [Google Scholar] [CrossRef] [PubMed]
- Doetsch, F.; Wichterle, H.; Lim, D.A.; Alvarez-Buylla, A. Architecture and cell types of the adult subventricular zone: In search of the stem cells. J. Neurobiol. 1998, 36, 234–248. [Google Scholar] [CrossRef]
- Moss, J.; Gebara, E.; Bushong, E.A.; Sánchez-Pascual, I.; O’laoi, R.; El M’ghari, I.; Kocher-Braissant, J.; Ellisman, M.H.; Toni, N. Fine processes of Nestin-GFP–positive radial glia-like stem cells in the adult dentate gyrus ensheathe local synapses and vasculature. Proc. Natl. Acad. Sci. USA 2016, 113, E2536–E2545. [Google Scholar] [CrossRef] [PubMed]
- Seri, B.; García-Verdugo, J.M.; McEwen, B.S.; Alvarez-Buylla, A. Astrocytes give rise to new neurons in the adult mammalian hippocampus. J. Neurosci. 2001, 21, 7153–7160. [Google Scholar] [CrossRef]
- Sun, G.J.; Zhou, Y.; Stadel, R.P.; Moss, J.; Yong, J.H.A.; Ito, S.; Kawasaki, N.K.; Phan, A.T.; Oh, J.H.; Modak, N.; et al. Tangential migration of neuronal precursors of glutamatergic neurons in the adult mammalian brain. Proc. Natl. Acad. Sci. USA 2015, 112, 9484–9489. [Google Scholar] [CrossRef]
- Berg, D.A.; Bond, A.; Ming, G.-L.; Song, H. Radial glial cells in the adult dentate gyrus: What are they and where do they come from? F1000Research 2018, 7, 277. [Google Scholar] [CrossRef]
- Bergmann, O.; Liebl, J.; Bernard, S.; Alkass, K.; Yeung, M.S.; Steier, P.; Kutschera, W.; Johnson, L.; Landén, M.; Druid, H.; et al. The age of olfactory bulb neurons in humans. Neuron 2012, 74, 634–639. [Google Scholar] [CrossRef]
- Ernst, A.; Alkass, K.; Bernard, S.; Salehpour, M.; Perl, S.; Tisdale, J.; Possnert, G.; Druid, H.; Frisén, J. Neurogenesis in the striatum of the adult human brain. Cell 2014, 156, 1072–1083. [Google Scholar] [CrossRef]
- Morales, A.V.; Mira, H. Adult Neural Stem Cells: Born to Last. Front. Cell Dev. Biol. 2019, 7, 96. [Google Scholar] [CrossRef]
- Otsuki, L.; Brand, A.H. Cell cycle heterogeneity directs the timing of neural stem cell activation from quies-cence. Science 2018, 360, 99–102. [Google Scholar] [CrossRef]
- Urbán, N.; Blomfield, I.M.; Guillemot, F. Quiescence of Adult Mammalian Neural Stem Cells: A Highly Regulated Rest. Neuron 2019, 104, 834–848. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, Z.; Zhang, C.; Zhang, L.; Robin, A.; Wang, Y.; Lu, M.; Chopp, M. Stroke transiently increases subventricular zone cell division from asymmetric to symmetric and increases neuronal differentiation in the adult rat. J. Neurosci. 2004, 24, 5810–5815. [Google Scholar] [CrossRef] [PubMed]
- Dause, T.J.; Denninger, J.K.; Smith, B.M.; Kirby, E.D. The neural stem cell secretome across neurodevelopment. Exp. Neurol. 2022, 355, 114142. [Google Scholar] [CrossRef] [PubMed]
- Willis, C.M.; Nicaise, A.M.; Peruzzotti-Jametti, L.; Pluchino, S. The neural stem cell secretome and its role in brain repair. Brain Res. 2020, 1729, 146615. [Google Scholar] [CrossRef]
- Borrett, M.J.; Innes, B.T.; Tahmasian, N.; Bader, G.D.; Kaplan, D.R.; Miller, F.D. A Shared Transcriptional Identity for Forebrain and Dentate Gyrus Neural Stem Cells from Embryogenesis to Adulthood. eNeuro 2022, 9, ENEURO.0271-21.2021. [Google Scholar] [CrossRef]
- Favaro, R.; Valotta, M.; Ferri, A.L.M.; Latorre, E.; Mariani, J.; Giachino, C.; Lancini, C.; Tosetti, V.; Ottolenghi, S.; Taylor, V.; et al. Hippocampal development and neural stem cell maintenance require Sox2-dependent regulation of Shh. Nat. Neurosci. 2009, 12, 1248–1256. [Google Scholar] [CrossRef]
- Arredondo, S.B.; Valenzuela-Bezanilla, D.; Mardones, M.D.; Varela-Nallar, L. Role of Wnt Signaling in Adult Hippocampal Neurogenesis in Health and Disease. Front. Cell Dev. Biol. 2020, 8, 860. [Google Scholar] [CrossRef]
- Wexler, E.M.; Paucer, A.; Kornblum, H.I.; Palmer, T.D.; Geschwind, D.H. Endogenous Wnt signaling maintains neural progenitor cell potency. Stem Cells 2009, 27, 1130–1141. [Google Scholar] [CrossRef]
- Jackson, A.; Panayiotidis, P.; Foroni, L. The human homologue of the Drosophila tailless gene (TLX): Characterization and mapping to a region of common de-letion in human lymphoid leukemia on chromosome 6q21. Genomics 1998, 50, 34–43. [Google Scholar] [CrossRef]
- Shi, Y.; Lie, D.C.; Taupin, P.; Nakashima, K.; Ray, J.; Yu, R.T.; Gage, F.H.; Evans, R.M. Expression and function of orphan nuclear receptor TLX in adult neural stem cells. Nature 2004, 427, 78–83. [Google Scholar] [CrossRef]
- Zhang, C.-L.; Zou, Y.; He, W.; Gage, F.H.; Evans, R.M. A role for adult TLX-positive neural stem cells in learning and behaviour. Nature 2008, 451, 1004–1007. [Google Scholar] [CrossRef] [PubMed]
- Willis, C.M.; Nicaise, A.M.; Hamel, R.; Pappa, V.; Peruzzotti-Jametti, L.; Pluchino, S. Harnessing the Neural Stem Cell Secretome for Regenerative Neuroimmunology. Front. Cell Neurosci. 2020, 14, 590960. [Google Scholar] [CrossRef] [PubMed]
- Makrygianni, E.A.; Chrousos, G.P. Extracellular vesicles and the Stress System. Neuroendocrinology 2022, 113, 120–167. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Adachi, N.; Matsuno, H.; Kawamata, M.; Yoshioka, Y.; Kikuchi, H.; Odaka, H.; Numakawa, T.; Kunugi, H.; Ochiya, T.; et al. The Sox2 promoter-driven CD63-GFP transgenic rat model allows tracking neural stem cell-derived extracellular vesicles. Dis. Model. Mech. 2018, 11, dmm028779. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Ding, L.; Chen, H.; Wang, Y.; Li, C.; Zhao, S.; Yang, X.; Ma, Y.; Zhu, J.; Qi, X.; et al. Neural Stem Cell-Derived Exosomes Regulate Neural Stem Cell Differentiation Through miR-9-Hes1 Axis. Front. Cell Dev. Biol. 2021, 9, 601600. [Google Scholar] [CrossRef]
- Dhanesh, S.B.; Subashini, C.; James, J. Hes1: The maestro in neurogenesis. Cell Mol. Life Sci. CMLS 2016, 73, 4019–4042. [Google Scholar] [CrossRef]
- Zhang, J.; Uchiyama, J.; Imami, K.; Ishihama, Y.; Kageyama, R.; Kobayashi, T. Novel Roles of Small Extracellular Vesicles in Regulating the Quiescence and Proliferation of Neural Stem Cells. Front. Cell Dev. Biol. 2021, 9, 762293. [Google Scholar] [CrossRef]
- Peruzzotti-Jametti, L.; Bernstock, J.D.; Willis, C.M.; Manferrari, G.; Rogall, R.; Fernandez-Vizarra, E.; Williamson, J.C.; Braga, A.; Bosch, A.V.D.; Leonardi, T.; et al. Neural stem cells traffic functional mitochondria via extracellular vesicles. PLoS Biol. 2021, 19, e3001166. [Google Scholar] [CrossRef]
- Morton, M.C.; Neckles, V.N.; Seluzicki, C.M.; Holmberg, J.C.; Feliciano, D.M. Neonatal Subventricular Zone Neural Stem Cells Release Extracellular Vesicles that Act as a Microglial Morphogen. Cell Rep. 2018, 23, 78–89. [Google Scholar] [CrossRef]
- Cossetti, C.; Iraci, N.; Mercer, T.R.; Leonardi, T.; Alpi, E.; Drago, D.; Alfaro-Cervello, C.; Saini, H.K.; Davis, M.P.; Schaeffer, J.; et al. Extracellular vesicles from neural stem cells transfer IFN-γ via Ifngr1 to activate Stat1 signaling in target cells. Mol. Cell 2014, 56, 193–204. [Google Scholar] [CrossRef]
- Liu, Q.; Tan, Y.; Qu, T.; Zhang, J.; Duan, X.; Xu, H.; Mu, Y.; Ma, H.; Wang, F. Therapeutic mechanism of human neural stem cell-derived extracellular vesicles against hypoxia-reperfusion injury in vitro. Life Sci. 2020, 254, 117772. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Giedzinski, E.; Angulo, M.C.; Lui, T.; Lu, C.; Park, A.L.; Tang, S.; Martirosian, V.; Ru, N.; Chmielewski, N.N.; et al. Functional equivalence of stem cell and stem cell-derived extracellular vesicle transplantation to repair the irradiated brain. Stem Cells Transl. Med. 2020, 9, 93–105. [Google Scholar] [CrossRef]
- Upadhya, R.; Madhu, L.N.; Attaluri, S.; Gitaí, D.L.G.; Pinson, M.R.; Kodali, M.; Shetty, G.; Zanirati, G.; Kumar, S.; Shuai, B.; et al. Extracellular vesicles from human iPSC-derived neural stem cells: miRNA and protein signatures, and anti-inflammatory and neurogenic properties. J. Extracell. Vesicles 2020, 9, 1809064. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.K.; Passaro, A.P.; Latchoumane, C.-F.; Spellicy, S.E.; Bowler, M.; Goeden, M.; Martin, W.J.; Holmes, P.V.; Stice, S.L.; Karumbaiah, L. Extracellular Vesicles Mediate Neuroprotection and Functional Recovery after Traumatic Brain Injury. J. Neurotrauma 2020, 37, 1358–1369. [Google Scholar] [CrossRef]
- Lee, E.J.; Choi, Y.; Lee, H.J.; Hwang, D.W.; Lee, D.S. Human neural stem cell-derived extracellular vesicles protect against Parkinson’s disease pathologies. J. Nanobiotechnology 2022, 20, 198. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Olsen, R.H.; Sun, J.; Ming, G.-L.; Song, H. Neuronal Circuitry Mechanisms Regulating Adult Mammalian Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8, a018937. [Google Scholar] [CrossRef]
- Zhao, C.; Deng, W.; Gage, F.H. Mechanisms and Functional Implications of Adult Neurogenesis. Cell 2008, 132, 645–660. [Google Scholar] [CrossRef]
- Jang, M.H.; Song, H.; Ming, G.L. Regulation of adult neurogenesis by neurotransmitters. In Adult Neurogenesis; Gage, F.H., Kempermann, G., Song, H., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2008. [Google Scholar]
- Young, S.Z.; Taylor, M.M.; Bordey, A. Neurotransmitters couple brain activity to subventricular zone neurogenesis. Eur. J. Neurosci. 2011, 33, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Vivar, C.; Potter, M.C.; Choi, J.; Lee, J.-Y.; Stringer, T.P.; Callaway, E.M.; Gage, F.H.; Suh, H.; Van Praag, H. Monosynaptic inputs to new neurons in the dentate gyrus. Nat. Commun. 2012, 3, 1107. [Google Scholar] [CrossRef]
- Deshpande, A.; Bergami, M.; Ghanem, A.; Conzelmann, K.-K.; Lepier, A.; Götz, M.; Berninger, B. Retrograde monosynaptic tracing reveals the temporal evolution of inputs onto new neurons in the adult dentate gyrus and olfactory bulb. Proc. Natl. Acad. Sci. USA 2013, 110, E1152–E1161. [Google Scholar] [CrossRef]
- Song, J.; Zhong, C.; Bonaguidi, M.A.; Sun, G.J.; Hsu, D.; Gu, Y.; Meletis, K.; Huang, Z.J.; Ge, S.; Enikolopov, G.; et al. Neuronal circuitry mechanism regulating adult quiescent neural stem-cell fate decision. Nature 2012, 489, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Sun, J.; Moss, J.; Wen, Z.; Sun, G.J.; Hsu, D.; Zhong, C.; Davoudi, H.; Christian, K.M.; Toni, N.; et al. Parvalbumin interneurons mediate neuronal circuitry–neurogenesis coupling in the adult hippocampus. Nat. Neurosci. 2013, 16, 1728–1730. [Google Scholar] [CrossRef] [PubMed]
- Radley, J.J.; Jacobs, B.L. 5-HT1A receptor antagonist administration decreases cell proliferation in the dentate gyrus. Brain Res. 2002, 955, 264–267. [Google Scholar] [CrossRef]
- Malberg, J.E.; Eisch, A.J.; Nestler, E.J.; Duman, R.S. Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. J. Neurosci. 2000, 20, 9104–9110. [Google Scholar] [CrossRef]
- Bolteus, A.J.; Bordey, A. GABA release and uptake regulate neuronal precursor migration in the postnatal subventricular zone. J. Neurosci. 2004, 24, 7623–7631. [Google Scholar] [CrossRef]
- Bordey, A. Enigmatic GABAergic networks in adult neurogenic zones. Brain Res. Rev. 2007, 53, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Banasr, M.; Hery, M.; Printemps, R.; Daszuta, A. Serotonin-induced increases in adult cell proliferation and neurogenesis are mediated through different and common 5-HT receptor subtypes in the dentate gyrus and the subventricular zone. Neuropsychopharmacology 2003, 29, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Van Kampen, J.; Robertson, H. A possible role for dopamine D3 receptor stimulation in the induction of neurogenesis in the adult rat substantia nigra. Neuroscience 2005, 136, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Zaben, M.; Sheward, W.J.; Shtaya, A.; Abbosh, C.; Harmar, A.J.; Pringle, A.K.; Gray, W.P. The neurotransmitter VIP expands the pool of symmetrically dividing postnatal dentate gyrus precursors via VPAC2 receptors or directs them toward a neuronal fate via VPAC1 receptors. Stem Cells 2009, 27, 2539–2551. [Google Scholar] [CrossRef]
- Decressac, M.; Wright, B.; David, B.; Tyers, P.; Jaber, M.; Barker, R.A.; Gaillard, A. Exogenous neuropeptide Y promotes in vivo hippocampal neurogenesis. Hippocampus 2011, 21, 233–238. [Google Scholar] [CrossRef]
- Geloso, M.C.; Corvino, V.; Di Maria, V.; Marchese, E.; Michetti, F. Cellular targets for neuropeptide Y-mediated control of adult neurogenesis. Front. Cell Neurosci. 2015, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Howell, O.; Doyle, K.; Goodman, J.H.; Scharfman, H.E.; Herzog, H.; Pringle, A.; Beck-Sickinger, A.G.; Gray, W.P. Neuropeptide Y stimulates neuronal precursor proliferation in the post-natal and adult dentate gyrus. J. Neurochem. 2005, 93, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Mercer, A.; Rönnholm, H.; Holmberg, J.; Lundh, H.; Heidrich, J.; Zachrisson, O.; Ossoinak, A.; Frisén, J.; Patrone, C. PACAP promotes neural stem cell proliferation in adult mouse brain. J. Neurosci. Res. 2004, 76, 205–215. [Google Scholar] [CrossRef] [PubMed]
- van den Pol, A.N. Neuropeptide Transmission in Brain Circuits. Neuron 2012, 76, 98–115. [Google Scholar] [CrossRef] [PubMed]
- Anacker, C.; Cattaneo, A.; Luoni, A.; Musaelyan, K.; A Zunszain, P.; Milanesi, E.; Rybka, J.; Berry, A.; Cirulli, F.; Thuret, S.; et al. Glucocorticoid-Related Molecular Signaling Pathways Regulating Hippocampal Neurogenesis. Neuropsychopharmacology 2013, 38, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, R.; Ohtsuka, T. The Notch-Hes pathway in mammalian neural development. Cell Res. 1999, 9, 179–188. [Google Scholar] [CrossRef]
- Graciarena, M.; Depino, A.M.; Pitossi, F.J. Prenatal inflammation impairs adult neurogenesis and memory related behavior through persistent hippocampal TGFbeta1 downregulation. Brain Behav. Immun. 2010, 24, 1301–1309. [Google Scholar] [CrossRef]
- Seuntjens, E.; Umans, L.; Zwijsen, A.; Sampaolesi, M.; Verfaillie, C.M.; Huylebroeck, D. Transforming Growth Factor type beta and Smad family signaling in stem cell function. Cytokine Growth Factor Rev. 2009, 20, 449–458. [Google Scholar] [CrossRef]
- Ahn, S.; Joyner, A.L. In vivo analysis of quiescent adult neural stem cells responding to Sonic hedgehog. Nature 2005, 437, 894–897. [Google Scholar] [CrossRef]
- Cai, C.; Thorne, J.; Grabel, L. Hedgehog serves as a mitogen and survival factor during embryonic stem cell neuro-genesis. Stem Cells 2008, 26, 1097–1108. [Google Scholar] [CrossRef]
- Shimozaki, K. Sox2 transcription network acts as a molecular switch to regulate properties of neural stem cells. World J. Stem Cells 2014, 6, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Episkopou, V. SOX2 functions in adult neural stem cells. Trends Neurosci. 2005, 28, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cui, W. Sox2, a key factor in the regulation of pluripotency and neural differentiation. World J. Stem Cells 2014, 6, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, T.; Lengerke, C. SOX2 protein biochemistry in stemness, reprogramming, and cancer: The PI3K/AKT/SOX2 axis and beyond. Oncogene 2020, 39, 278–292. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Corbi, N.; Basilico, C.; Dailey, L. Developmental-specific activity of the FGF-4 enhancer requires the synergistic action of Sox2 and Oct-3. Genes Dev. 1995, 9, 2635–2645. [Google Scholar] [CrossRef] [PubMed]
- Bernal, A.; Arranz, L. Nestin-expressing progenitor cells: Function, identity and therapeutic implications. Cell Mol. Life Sci. 2018, 75, 2177–2195. [Google Scholar] [CrossRef]
- Liem, R.K.H. Neuronal Intermediate Filaments. In Encyclopedia of Biological Chemistry, 2nd ed.; Lennarz, W.J., Lane, M.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 233–237. ISBN 978-0-12-378631-9. [Google Scholar]
- Suzuki, S.; Namiki, J.; Shibata, S.; Mastuzaki, Y.; Okano, H. The neural stem/progenitor cell marker nestin is expressed in proliferative endothelial cells, but not in mature vasculature. J. Histochem. Cytochem. 2010, 58, 721–730. [Google Scholar] [CrossRef]
- Chou, Y.-H.; Khuon, S.; Herrmann, H.; Goldman, R.D. Nestin promotes the phosphorylation-dependent disassembly of vimentin intermediate filaments during mitosis. Mol. Biol. Cell 2003, 14, 1468–1478. [Google Scholar] [CrossRef]
- Herrmann, H.; Aebi, U. Intermediate filaments and their associates: Multi-talented structural elements specifying cytoarchitecture and cytodynamics. Curr. Opin. Cell Biol. 2000, 12, 79–90. [Google Scholar] [CrossRef]
- Barry, D.; McDermott, K. Differentiation of radial glia from radial precursor cells and transformation into astrocytes in the developing rat spinal cord. Glia 2005, 50, 187–197. [Google Scholar] [CrossRef]
- Johnston, J.A.; Ward, C.L.; Kopito, R.R. Aggresomes: A Cellular Response to Misfolded Proteins. J. Cell Biol. 1998, 143, 1883–1898. [Google Scholar] [CrossRef] [PubMed]
- Morrow, C.S.; Porter, T.J.; Xu, N.; Arndt, Z.P.; Ako-Asare, K.; Heo, H.J.; Thompson, E.A.; Moore, D.L. Vimentin Coordinates Protein Turnover at the Aggresome during Neural Stem Cell Quiescence Exit. Cell Stem Cell 2020, 26, 558–568.e9. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Barragan, J.; Bashiruddin, S.; Smith, C.J.; Tyrrell, C.; Parsons, M.J.; Doris, R.; Kucenas, S.; Downes, G.B.; Velez, C.M.; et al. Gfap-positive radial glial cells are an essential progenitor population for later-born neurons and glia in the zebrafish spinal cord. Glia 2016, 64, 1170–1189. [Google Scholar] [CrossRef]
- Docampo-Seara, A.; Santos-Durán, G.N.; Candal, E.; Díaz, M.R. Expression of radial glial markers (GFAP, BLBP and GS) during telencephalic development in the catshark (Scyliorhinus canicula). Brain Struct. Funct. 2019, 224, 33–56. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, K.K. Glial fibrillary acidic protein: From intermediate filament assembly and gliosis to neurobiomarker. Trends Neurosci. 2015, 38, 364–374. [Google Scholar] [CrossRef]
- Shibata, T.; Yamada, K.; Watanabe, M.; Ikenaka, K.; Wada, K.; Tanaka, K.; Inoue, Y. Glutamate Transporter GLAST Is Expressed in the Radial Glia–Astrocyte Lineage of Developing Mouse Spinal Cord. J. Neurosci. 1997, 17, 9212–9219. [Google Scholar] [CrossRef]
- Imura, T.; Nakano, I.; Kornblum, H.I.; Sofroniew, M.V. Phenotypic and functional heterogeneity of GFAP-expressing cells in vitro: Differential expression of LeX/CD15 by GFAP-expressing multipotent neural stem cells and non-neurogenic astrocytes. Glia 2006, 53, 277–293. [Google Scholar] [CrossRef]
- DeCarolis, N.A.; Mechanic, M.; Petrik, D.; Carlton, A.; Ables, J.L.; Malhotra, S.; Bachoo, R.; Götz, M.; Lagace, D.C.; Eisch, A.J. In vivo contribution of nestin- and GLAST-lineage cells to adult hippocampal neurogenesis. Hippocampus 2013, 23, 708–719. [Google Scholar] [CrossRef]
- Feng, L.; Hatten, M.E.; Heintz, N. Brain lipid-binding protein (BLBP): A novel signaling system in the developing mammalian CNS. Neuron 1994, 12, 895–908. [Google Scholar] [CrossRef]
- Kurtz, A.; Zimmer, A.; Schnütgen, F.; Brüning, G.; Spener, F.; Müller, T. The expression pattern of a novel gene encoding brain-fatty acid binding protein correlates with neuronal and glial cell development. Development 1994, 120, 2637–2649. [Google Scholar] [CrossRef]
- Hartfuss, E.; Galli, R.; Heins, N.; Götz, M. Characterization of CNS Precursor Subtypes and Radial Glia. Dev. Biol. 2001, 229, 15–30. [Google Scholar] [CrossRef]
- E Anthony, T.; Klein, C.; Fishell, G.; Heintz, N. Radial Glia Serve as Neuronal Progenitors in All Regions of the Central Nervous System. Neuron 2004, 41, 881–890. [Google Scholar] [CrossRef]
- Doetsch, F. The glial identity of neural stem cells. Nat. Neurosci. 2003, 6, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Kriegstein, A.R.; Götz, M. Radial glia diversity: A matter of cell fate. Glia 2003, 43, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Thakurela, S.; Tiwari, N.; Schick, S.; Garding, A.; Ivanek, R.; Berninger, B.; Tiwari, V.K. Mapping gene regulatory circuitry of Pax6 during neurogenesis. Cell Discov. 2016, 2, 15045. [Google Scholar] [CrossRef]
- Kanemura, Y.; Yamasaki, M.; Mori, K.; Fujikawa, H.; Hayashi, H.; Nakano, A.; Matsumoto, T.; Tamura, K.; Arita, N.; Sakakibara, S.-I.; et al. Musashi1, an evolutionarily conserved neural RNA-binding protein, is a versatile marker of human glioma cells in determining their cellular origin, malignancy, and proliferative activity. Differ. Res. Biol. Divers. 2001, 68, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Essers, J.; Theil, A.F.; Baldeyron, C.; van Cappellen, W.A.; Houtsmuller, A.B.; Kanaar, R.; Vermeulen, W. Nuclear Dynamics of PCNA in DNA Replication and Repair. Mol. Cell Biol. 2005, 25, 9350–9359. [Google Scholar] [CrossRef]
- Mansilla, S.F.; De La Vega, M.B.; Calzetta, N.L.; Siri, S.O.; Gottifredi, V. CDK-Independent and PCNA-Dependent Functions of p21 in DNA Replication. Genes 2020, 11, 593. [Google Scholar] [CrossRef]
- Limke, T.L.; Cai, J.; Miura, T.; Rao, M.S.; Mattson, M.P. Distinguishing Features of Progenitor Cells in the Late Embryonic and Adult Hippocampus. Dev. Neurosci. 2003, 25, 257–272. [Google Scholar] [CrossRef]
- Kayaselçuk, F.; Zorludemir, S.; Gümürdülü, D.; Zeren, H.; Erman, T. PCNA and Ki-67 in Central Nervous System Tumors: Correlation with the Histological Type and Grade. J. Neuro-Oncol. 2002, 57, 115–121. [Google Scholar] [CrossRef]
- Maslov, A.Y.; Barone, T.A.; Plunkett, R.J.; Pruitt, S.C. Neural Stem Cell Detection, Characterization, and Age-Related Changes in the Subventricular Zone of Mice. J. Neurosci. 2004, 24, 1726–1733. [Google Scholar] [CrossRef]
- Bonfanti, L.; Seki, T. The PSA-NCAM-Positive “Immature” Neurons: An Old Discovery Providing New Vistas on Brain Structural Plasticity. Cells 2021, 10, 2542. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, J.G.; Lin, P.T.; A Flanagan, L.; A Walsh, C. Doublecortin Is a Microtubule-Associated Protein and Is Expressed Widely by Migrating Neurons. Neuron 1999, 23, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Couillard-Despres, S.; Winner, B.; Schaubeck, S.; Aigner, R.; Vroemen, M.; Weidner, N.; Bogdahn, U.; Winkler, J.; Kuhn, H.-G.; Aigner, L. Doublecortin expression levels in adult brain reflect neurogenesis. Eur. J. Neurosci. 2005, 21, 1. [Google Scholar] [CrossRef] [PubMed]
- Batailler, M.; Droguerre, M.; Baroncini, M.; Fontaine, C.; Prevot, V.; Migaud, M. DCX-expressing cells in the vicinity of the hypothalamic neurogenic niche: A comparative study between mouse, sheep, and human tissues. J. Comp. Neurol. 2014, 522, 1966–1985. [Google Scholar] [CrossRef] [PubMed]
- Wojtowicz, J.M.; Kee, N. BrdU assay for neurogenesis in rodents. Nat. Protoc. 2006, 1, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Gratzner, H.G. Monoclonal Antibody to 5-Bromo- and 5-Iododeoxyuridine: A New Reagent for Detection of DNA Replication. Science 1982, 218, 474–475. [Google Scholar] [CrossRef]
- Taupin, P. BrdU immunohistochemistry for studying adult neurogenesis: Paradigms, pitfalls, limitations, and validation. Brain Res. Rev. 2007, 53, 198–214. [Google Scholar] [CrossRef]
- Duan, W.; Zhang, Y.-P.; Hou, Z.; Huang, C.; Zhu, H.; Zhang, C.-Q.; Yin, Q. Novel Insights into NeuN: From Neuronal Marker to Splicing Regulator. Mol. Neurobiol. 2016, 53, 1637–1647. [Google Scholar] [CrossRef]
- Kim, K.K.; Adelstein, R.S.; Kawamoto, S. Identification of Neuronal Nuclei (NeuN) as Fox-3, a New Member of the Fox-1 Gene Family of Splicing Factors. J. Biol. Chem. 2009, 284, 31052–31061. [Google Scholar] [CrossRef]
- Kim, K.K.; Nam, J.; Mukouyama, Y.-S.; Kawamoto, S. Rbfox3-regulated alternative splicing of Numb promotes neuronal differentiation during development. J. Cell Biol. 2013, 200, 443–458. [Google Scholar] [CrossRef]
- Mullen, R.J.; Buck, C.R.; Smith, A.M. NeuN, a neuronal specific nuclear protein in vertebrates. Development 1992, 116, 201–211. [Google Scholar] [CrossRef]
- Wolf, H.K.; Buslei, R.; Schmidt-Kastner, R.; Schmidt-Kastner, P.K.; Pietsch, T.; Wiestler, O.D.; Blumcke, I. NeuN: A useful neuronal marker for diagnostic histopathology. J. Histochem. Cytochem. 1996, 44, 1167–1171. [Google Scholar] [CrossRef]
- Sarnat, H.B.; Nochlin, D.; Born, D.E. Neuronal nuclear antigen (NeuN): A marker of neuronal maturation in early human fetal nervous system. Brain Dev. 1998, 20, 88–94. [Google Scholar] [CrossRef]
- Portiansky, E.L.; Barbeito, C.G.; Gimeno, E.J.; Zuccolilli, G.O.; Goya, R.G. Loss of NeuN immunoreactivity in rat spinal cord neurons during aging. Exp. Neurol. 2006, 202, 519–521. [Google Scholar] [CrossRef]
- Ünal-Çevik, I.; Kılınç, M.; Gürsoy-Özdemir, Y.; Gurer, G.; Dalkara, T. Loss of NeuN immunoreactivity after cerebral ischemia does not indicate neuronal cell loss: A cautionary note. Brain Res. 2004, 1015, 169–174. [Google Scholar] [CrossRef]
- Cannon, J.R.; Greenamyre, J.T. NeuN is not a reliable marker of dopamine neurons in rat substantia nigra. Neurosci. Lett. 2009, 464, 14–17. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Shu, H.F.; Yin, Q.; An, N.; Xu, S.L.; Yin, J.B.; Song, Y.C.; Liu, S.Y.; Yang, H. Expression and cellular distribution of vascular endothelialgrowth factor-C system in cortical tubers of the tuberous sclerosis complex. Brain Pathol. 2012, 22, 205–218. [Google Scholar] [CrossRef]
- Wu, K.-L.; Li, Y.-Q.; Tabassum, A.; Lu, W.-Y.; Aubert, I.; Wong, C.S. Loss of Neuronal Protein Expression in Mouse Hippocampus After Irradiation. J. Neuropathol. Exp. Neurol. 2010, 69, 272–280. [Google Scholar] [CrossRef]
- Rizzoti, K.; Lovell-Badge, R. Pivotal role of median eminence tanycytes for hypothalamic function and neurogenesis. Mol. Cell Endocrinol. 2017, 445, 7–13. [Google Scholar] [CrossRef]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Makrygianni, E.A.; Chrousos, G.P. From Brain Organoids to Networking Assembloids: Implications for Neuroendocrinology and Stress Medicine. Front. Physiol. 2021, 12, 621970. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, M.B. Core Text of Neuroanatomy, 4th ed.; Williams and Wilkins: Baltimore, MD, USA, 1991. [Google Scholar]
- Miyata, S. New aspects in fenestrated capillary and tissue dynamics in the sensory circumventricular organs of adult brains. Front. Neurosci. 2015, 9, 390. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.A.; Morrison, E.H. Neuroanatomy, Nucleus Suprachiasmatic. In StatPearls; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Burbach, J.P.; Luckman, S.M.; Murphy, D.; Gainer, H. Gene Regulation in the Magnocellular Hypothalamo-Neurohypophysial System. Physiol. Rev. 2001, 81, 1197–1267. [Google Scholar] [CrossRef]
- Morris, J.F. Neurosecretion (Regulated Exocytosis in Neuroendocrine Cells). In Encyclopedia of Neuroscience; Larry, R.S., Ed.; Academic Press: Cambridge, MA, USA, 2009; pp. 1007–1014. ISBN 9780080450469. [Google Scholar] [CrossRef]
- Wagner, E. Monoaminergic and Cholinergic Control of the Anterior Pituitary. In Encyclopedia of Neuroscience; Helen, L.H., Anthony, W.N., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 704–712. ISBN 9780123411037. [Google Scholar] [CrossRef]
- Murphy, D.; Waller, S.; Fairhall, K.; Carter, D.A.; Robinson, I.C. Regulation of the synthesis and secretion of vasopressin. Prog. Brain Res. 1999, 119, 137–143. [Google Scholar] [CrossRef]
- Koshimizu, T.-A.; Nakamura, K.; Egashira, N.; Hiroyama, M.; Nonoguchi, H.; Tanoue, A. Vasopressin V1a and V1b Receptors: From Molecules to Physiological Systems. Physiol. Rev. 2012, 92, 1813–1864. [Google Scholar] [CrossRef]
- Golombek, D.A.; Rosenstein, R.E. Physiology of Circadian Entrainment. Physiol. Rev. 2010, 90, 1063–1102. [Google Scholar] [CrossRef]
- Ramkisoensing, A.; Meijer, J.H. Synchronization of Biological Clock Neurons by Light and Peripheral Feedback Systems Promotes Circadian Rhythms and Health. Front. Neurol. 2015, 6, 128. [Google Scholar] [CrossRef]
- Kawakami, N.; Otubo, A.; Maejima, S.; Talukder, A.H.; Satoh, K.; Oti, T.; Takanami, K.; Ueda, Y.; Itoi, K.; Morris, J.F.; et al. Variation of pro-vasopressin processing in parvocellular and magnocellular neurons in the paraventricular nucleus of the hypothalamus: Evidence from the vasopressin-related glycopeptide copeptin. J. Comp. Neurol. 2021, 529, 1372–1390. [Google Scholar] [CrossRef]
- Plotsky, P.M. Regulation of Hypophysiotropic Factors Mediating ACTH Secretion. Ann. N. Y. Acad. Sci. 1987, 512, 205–217. [Google Scholar] [CrossRef]
- Plotsky, P.M.; Sawchenko, P.E. Hypophysial-Portal Plasma Levels, Median Eminence Content, and Immunohistochemical Staining of Corticotropin-Releasing Factor, Arginine Vasopressin, and Oxytocin after Pharmacological Adrenalectomy. Endocrinology 1987, 120, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Whitnall, M.H.; Mezey, Ė.; Gainer, H. Co-localization of corticotropin-releasing factor and vasopressin in median eminence neurosecretory vesicles. Nature 1985, 317, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Sawchenko, P.E. Evidence for differential regulation of corticotropin-releasing factor and vasopressin immunoreactivities in parvocellular neurosecretory and autonomic-related projections of the paraventricular nucleus. Brain Res. 1987, 437, 253–263. [Google Scholar] [CrossRef]
- Xie, Y.; Dorsky, R.I. Development of the hypothalamus: Conservation, modification and innovation. Development 2017, 144, 1588–1599. [Google Scholar] [CrossRef]
- Grandel, H.; Kaslin, J.; Ganz, J.; Wenzel, I.; Brand, M. Neural stem cells and neurogenesis in the adult zebrafish brain: Origin, proliferation dynamics, migration and cell fate. Dev. Biol. 2006, 295, 263–277. [Google Scholar] [CrossRef]
- Wang, X.; Kopinke, D.; Lin, J.; McPherson, A.D.; Duncan, R.N.; Otsuna, H.; Moro, E.; Hoshijima, K.; Grunwald, D.J.; Argenton, F.; et al. Wnt Signaling Regulates Postembryonic Hypothalamic Progenitor Differentiation. Dev. Cell 2012, 23, 624–636. [Google Scholar] [CrossRef]
- Steinbusch, H.W.; Sauren, Y.; Groenewegen, H.; Watanabe, T.; Mulder, A.H. Histaminergic projections from the premammillary and posterior hypothalamic region to the caudate-putamen complex in the rat. Brain Res. 1986, 368, 389–393. [Google Scholar] [CrossRef] [PubMed]
- McPherson, A.D.; Barrios, J.P.; Luks-Morgan, S.J.; Manfredi, J.P.; Bonkowsky, J.L.; Douglass, A.D.; Dorsky, R.I. Motor Behavior Mediated by Continuously Generated Dopaminergic Neurons in the Zebrafish Hypothalamus Recovers after Cell Ablation. Curr. Biol. 2016, 26, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Ruuskanen, J.O.; Wullimann, M.F.; Vernier, P. Two tyrosine hydroxylase genes in vertebrates: New dopaminergic territories revealed in the zebrafish brain. Mol. Cell Neurosci. 2010, 43, 394–402. [Google Scholar] [CrossRef]
- Burbridge, S.; Stewart, I.; Placzek, M. Development of the Neuroendocrine Hypothalamus. Compr. Physiol. 2016, 6, 623–643. [Google Scholar]
- Ferran, J.L.; Puelles, L.; Rubenstein, J.L. Molecular codes defining rostrocaudal domains in the embryonic mouse hypothalamus. Front. Neuroanat. 2015, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Pearson, C.A.; Ohyama, K.; Manning, L.; Aghamohammadzadeh, S.; Sang, H.; Placzek, M. FGF-dependent midline-derived progenitor cells in hypothalamic infundibular development. Development 2011, 138, 2613–2624. [Google Scholar] [CrossRef]
- Pearson, C.A.; Placzek, M. Development of the medial hypothalamus: Forming a functional hypothalamic-neurohypophyseal interface. Curr. Top. Dev. Biol. 2013, 106, 49–88. [Google Scholar] [CrossRef]
- Goto, M.; Hojo, M.; Ando, M.; Kita, A.; Kitagawa, M.; Ohtsuka, T.; Kageyama, R.; Miyamoto, S. Hes1 and Hes5 are required for differentiation of pituicytes and formation of the neurohypophysis in pituitary development. Brain Res. 2015, 1625, 206–217. [Google Scholar] [CrossRef]
- Manning, L.; Ohyama, K.; Saeger, B.; Hatano, O.; Wilson, S.A.; Logan, M.; Placzek, M. Regional Morphogenesis in the Hypothalamus: A BMP-Tbx2 Pathway Coordinates Fate and Proliferation through Shh Downregulation. Dev. Cell 2006, 11, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Szabó, N.-E.; Zhao, T.; Çankaya, M.; Theil, T.; Zhou, X.; Alvarez-Bolado, G. Role of Neuroepithelial Sonic hedgehogin Hypothalamic Patterning. J. Neurosci. 2009, 29, 6989–7002. [Google Scholar] [CrossRef] [PubMed]
- Trowe, M.-O.; Zhao, L.; Weiss, A.-C.; Christoffels, V.; Epstein, D.J.; Kispert, A. Inhibition of Sox2-dependent activation of Shh in the ventral diencephalon by Tbx3 is required for formation of the neurohypophysis. Development 2013, 140, 2299–2309. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zevallos, S.E.; Rizzoti, K.; Jeong, Y.; Lovell-Badge, R.; Epstein, D.J. Disruption of SoxB1-Dependent Sonic hedgehog Expression in the Hypothalamus Causes Septo-optic Dysplasia. Dev. Cell 2012, 22, 585–596. [Google Scholar] [CrossRef]
- Zhao, Y.; Mailloux, C.M.; Hermesz, E.; Palkóvits, M.; Westphal, H. A role of the LIM-homeobox gene Lhx2 in the regulation of pituitary development. Dev. Biol. 2010, 337, 313–323. [Google Scholar] [CrossRef]
- Takuma, N.; Sheng, H.Z.; Furuta, Y.; Ward, J.M.; Sharma, K.; Hogan, B.L.M.; Pfaff, S.L.; Westphal, H.; Kimura, S.; Mahon, K.A. Formation of Rathke’s pouch requires dual induction from the diencephalon. Development 1998, 125, 4835–4840. [Google Scholar] [CrossRef]
- Rodríguez, E.M.; Blázquez, J.L.; Pastor, F.E.; Peláez, B.; Peña, P.; Peruzzo, B.; Amat, P. Hypothalamic Tanycytes: A Key Component of Brain–Endocrine Interaction. Int. Rev. Cytol. 2005, 247, 89–164. [Google Scholar] [CrossRef] [PubMed]
- Salvatierra, J.; Lee, D.A.; Zibetti, C.; Duran-Moreno, M.; Yoo, S.; Newman, E.A.; Wang, H.; Bedont, J.L.; de Melo, J.; Miranda-Angulo, A.L.; et al. The LIM Homeodomain Factor Lhx2 Is Required for Hypothalamic Tanycyte Specification and Differentiation. J. Neurosci. 2014, 34, 16809–16820. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Kar, D.; Gruenig, N.; Zhang, Z.W.; Cousins, N.; Rodgers, H.M.; Swindell, E.C.; Jamrich, M.; Schuurmans, C.; Mathers, P.H.; et al. Rax Is a Selector Gene for Mediobasal Hypothalamic Cell Types. J. Neurosci. 2013, 33, 259–272. [Google Scholar] [CrossRef]
- Shimogori, T.; A Lee, D.; Miranda-Angulo, A.; Yang, Y.; Wang, H.; Jiang, L.; Yoshida, A.C.; Kataoka, A.; Mashiko, H.; Avetisyan, M.; et al. A genomic atlas of mouse hypothalamic development. Nat. Neurosci. 2010, 13, 767–775. [Google Scholar] [CrossRef]
- Guilding, C.; Hughes, A.T.; Brown, T.M.; Namvar, S.; Piggins, H.D. A riot of rhythms: Neuronal and glial circadian oscillators in the mediobasal hypothalamus. Mol. Brain 2009, 2, 28. [Google Scholar] [CrossRef]
- Korf, H.-W.; Møller, M. Arcuate nucleus, median eminence, and hypophysial pars tuberalis. Handb. Clin. Neurol. 2021, 180, 227–251. [Google Scholar] [CrossRef]
- Balthasar, N.; Dalgaard, L.T.; Lee, C.E.; Yu, J.; Funahashi, H.; Williams, T.; Ferreira, M.; Tang, V.; McGovern, R.A.; Kenny, C.D.; et al. Divergence of Melanocortin Pathways in the Control of Food Intake and Energy Expenditure. Cell 2005, 123, 493–505. [Google Scholar] [CrossRef]
- Gropp, E.; Shanabrough, M.; Borok, E.; Xu, A.W.; Janoschek, R.; Buch, T.; Plum, L.; Balthasar, N.; Hampel, B.; Waisman, A.; et al. Agouti-related peptide–expressing neurons are mandatory for feeding. Nat. Neurosci. 2005, 8, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.N.; Macosko, E.Z.; Fenselau, H.; Pers, T.H.; Lyubetskaya, A.; Tenen, D.; Goldman, M.; Verstegen, A.M.; Resch, J.M.; McCarroll, S.A.; et al. A molecular census of arcuate hypothalamus and median eminence cell types. Nat. Neurosci. 2017, 20, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Hrabovszky, E.; Takács, S.; Rumpler, É.; Skrapits, K. The human hypothalamic kisspeptin system: Functional neuroanatomy and clinical perspectives. Handb. Clin. Neurol. 2021, 180, 275–296. [Google Scholar] [CrossRef]
- Rodríguez, E.M.; Blázquez, J.L.; Guerra, M. The design of barriers in the hypothalamus allows the median eminence and the arcuate nucleus to enjoy private milieus: The former opens to the portal blood and the latter to the cerebrospinal fluid. Peptides 2010, 31, 757–776. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, S.; Hori, M.; Ono, H.; Tabuchi, T.; Aoki, S.; Yasumoto, Y. The Infundibular Recess Passes through the Entire Pituitary Stalk. Clin. Neuroradiol. 2016, 26, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, E.; Guerra, M.; Peruzzo, B.; Blázquez, J.L. Tanycytes: A rich morphological history to underpin future molecular and physiological investigations. J. Neuroendocr. 2019, 31, e12690. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, P.; Dinan, T.G. Prolactin and dopamine: What is the connection? A Review Article. J. Psychopharmacol. 2008, 22, 12–19. [Google Scholar] [CrossRef]
- Al-Chalabi, M.; Bass, A.N.; Alsalman, I. Physiology, Prolactin. In StatPearls; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Rui, H.; Nevalainen, M.T. Nevalainen. Chapter 6—Prolactin. In The Cytokine Handbook, 4th ed.; Angus, W.T., Michael, T.L., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 115–147. ISBN 9780126896633. [Google Scholar] [CrossRef]
- Zhang, X.; van den Pol, A.N. Hypothalamic arcuate nucleus tyrosine hydroxylase neurons play orexigenic role in energy homeostasis. Nat. Neurosci 2016, 19, 1341–1347. [Google Scholar] [CrossRef]
- Koch, M.; Varela, L.; Kim, J.G.; Kim, J.D.; Hernández-Nuño, F.; Simonds, S.E.; Castorena, C.M.; Vianna, C.R.; Elmquist, J.K.; Morozov, Y.M.; et al. Hypothalamic POMC neurons promote cannabinoid-induced feeding. Nature 2015, 519, 45–50. [Google Scholar] [CrossRef]
- Cornejo, M.P.; Hentges, S.T.; Maliqueo, M.; Coirini, H.; Becu-Villalobos, D.; Elias, C.F. Neuroendocrine Regulation of Metabolism. J. Neuroendocr. 2016, 28, 1–24. [Google Scholar] [CrossRef]
- E Mayo, K.; Miller, T.; DeAlmeida, V.; Godfrey, P.; Zheng, J.; Cunha, S.R. Regulation of the pituitary somatotroph cell by GHRH and its receptor. Recent Prog. Horm. Res. 2000, 55, 237–267. [Google Scholar]
- Varela, L.; Horvath, T.L. Leptin and insulin pathways in POMC and AgRP neurons that modulate energy balance and glucose homeostasis. EMBO Rep. 2012, 13, 1079–1086. [Google Scholar] [CrossRef]
- Cowley, M.A.; Smart, J.L.; Rubinstein, M.; Cerdan, M.G.; Diano, S.; Horvath, T.L.; Cone, R.D.; Low, M.J. Leptin activates ano-rexigenic POMC neurons through a neural network in the arcuate nucleus. Nature 2001, 411, 480–484. [Google Scholar] [CrossRef]
- Lutz, T.A.; Coester, B.; Whiting, L.; Dunn-Meynell, A.A.; Boyle, C.N.; Bouret, S.G.; Levin, B.E.; Le Foll, C. Amylin Selectively Signals onto POMC Neurons in the Arcuate Nucleus of the Hypothalamus. Diabetes 2018, 67, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.; Le Wang, L.; Xia, B.; Liu, J.; Tahiri, A.; El Ouaamari, A.; Wheeler, M.B.; Pang, Z.P. Activation of arcuate nucleus glucagon-like peptide-1 receptor-expressing neurons suppresses food intake. Cell Biosci. 2022, 12, 178. [Google Scholar] [CrossRef] [PubMed]
- Garfield, A.S.; Li, C.; Madara, J.C.; Shah, B.P.; Webber, E.; Steger, J.S.; Campbell, J.N.; Gavrilova, O.; E Lee, C.; Olson, D.P.; et al. A neural basis for melanocortin-4 receptor–regulated appetite. Nat. Neurosci. 2015, 18, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Carmo, J.M.D.; da Silva, A.A.; Wang, Z.; Fang, T.; Aberdein, N.; de Lara, C.E.P.; Hall, J.E. Role of the brain melanocortins in blood pressure regulation. Biochim. Biophys. Acta. Mol. Basis Dis. 2017, 1863, 2508–2514. [Google Scholar] [CrossRef]
- Liu, Z.; Hruby, V.J. MC4R biased signalling and the conformational basis of biological function selections. J. Cell Mol. Med. 2022, 26, 4125–4136. [Google Scholar] [CrossRef]
- A Takahashi, K.; Cone, R.D. Fasting induces a large, leptin-dependent increase in the intrinsic action potential frequency of orexigenic arcuate nucleus neuropeptide Y/Agouti-related protein neurons. Endocrinology 2005, 146, 1043–1047. [Google Scholar] [CrossRef]
- Zhang, L.; Bijker, M.S.; Herzog, H. The neuropeptide Y system: Pathophysiological and therapeutic implications in obesity and cancer. Pharmacol. Ther. 2011, 131, 91–113. [Google Scholar] [CrossRef]
- Secher, A.; Jelsing, J.; Baquero, A.F.; Hecksher-Sørensen, J.; Cowley, M.A.; Dalbøge, L.S.; Hansen, G.; Grove, K.L.; Pyke, C.; Raun, K.; et al. The arcuate nucleus mediates GLP-1 receptor agonist liraglutide-dependent weight loss. J. Clin. Investig. 2014, 124, 4473–4488. [Google Scholar] [CrossRef]
- Le Roux, C.W.; Bloom, S.R. Peptide YY, appetite and food intake. Proc. Nutr. Soc. 2005, 64, 213–216. [Google Scholar] [CrossRef]
- Acuna-Goycolea, C.; van den Pol, A.N. Peptide YY(3-36) inhibits both anorexigenic proopiomelanocortin and orexigenic neuropeptide Y neurons: Implications for hypothalamic regulation of energy homeostasis. J. Neurosci. 2005, 25, 10510–10519. [Google Scholar] [CrossRef]
- Sato, I.; Arima, H.; Ozaki, N.; Watanabe, M.; Goto, M.; Hayashi, M.; Banno, R.; Nagasaki, H.; Oiso, Y. Insulin Inhibits Neuropeptide Y Gene Expression in the Arcuate Nucleus through GABAergic Systems. J. Neurosci. 2005, 25, 8657–8664. [Google Scholar] [CrossRef]
- Deem, J.D.; Faber, C.L.; Morton, G.J. AgRP neurons: Regulators of feeding, energy expenditure, and behavior. FEBS J. 2022, 289, 2362–2381. [Google Scholar] [CrossRef]
- Ruud, L.E.; Pereira, M.M.A.; de Solis, A.J.; Fenselau, H.; Brüning, J.C. NPY mediates the rapid feeding and glucose metabolism regulatory functions of AgRP neurons. Nat. Commun. 2020, 11, 442. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; A Essner, R.; Kosar, S.; Miller, O.H.; Lin, Y.-C.; Mesgarzadeh, S.; Knight, Z.A. Sustained NPY signaling enables AgRP neurons to drive feeding. eLife 2019, 8, e46348. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Woods, S.C.; Porte, D., Jr.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef]
- Sohn, J.-W.; Elmquist, J.K.; Williams, K.W. Neuronal circuits that regulate feeding behavior and metabolism. Trends Neurosci. 2013, 36, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Jobst, E.E.; Enriori, P.; Cowley, M.A. The electrophysiology of feeding circuits. Trends Endocrinol. Metab. TEM 2004, 15, 488–499. [Google Scholar] [CrossRef]
- Grayson, B.E.; Seeley, R.J.; Sandoval, D.A. Wired on sugar: The role of the CNS in the regulation of glucose homeostasis. Nat. Rev. Neurosci. 2013, 14, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Fioramonti, X.; Contié, S.; Song, Z.; Routh, V.H.; Lorsignol, A.; Pénicaud, L. Characterization of glucosensing neuron subpopulations in the arcuate nucleus: Integration in neuropeptide Y and pro-opio melanocortin networks? Diabetes 2007, 56, 1219–1227. [Google Scholar] [CrossRef]
- Parton, L.E.; Ye, C.P.; Coppari, R.; Enriori, P.J.; Choi, B.; Zhang, C.-Y.; Xu, C.; Vianna, C.R.; Balthasar, N.; Lee, C.E.; et al. Glucose sensing by POMC neurons regulates glucose homeostasis and is impaired in obesity. Nature 2007, 449, 228–232. [Google Scholar] [CrossRef]
- Ewang, D.; Ehe, X.; Ezhao, Z.; Efeng, Q.; Elin, R.; Esun, Y.; Eding, T.; Exu, F.; Eluo, M.; Ezhan, C. Whole-brain mapping of the direct inputs and axonal projections of POMC and AgRP neurons. Front. Neuroanat. 2015, 9, 40. [Google Scholar] [CrossRef]
- Buijs, F.N.; Guzmán-Ruiz, M.; León-Mercado, L.; Basualdo, M.C.; Escobar, C.; Kalsbeek, A.; Buijs, R.M. Suprachiasmatic Nucleus Interaction with the Arcuate Nucleus; Essential for Organizing Physiological Rhythms. eNeuro 2017, 4, ENEURO.0028-17.2017. [Google Scholar] [CrossRef]
- Méndez-Hernández, R.; Escobar, C.; Buijs, R.M. Suprachiasmatic Nucleus-Arcuate Nucleus Axis: Interaction Between Time and Metabolism Essential for Health. Obesity 2020, 28, S10–S17. [Google Scholar] [CrossRef]
- Bouret, S.G.; Draper, S.J.; Simerly, R.B. Formation of Projection Pathways from the Arcuate Nucleus of the Hypothalamus to Hypothalamic Regions Implicated in the Neural Control of Feeding Behavior in Mice. J. Neurosci. 2004, 24, 2797–2805. [Google Scholar] [CrossRef]
- Szabadi, E. Functional neuroanatomy of the central noradrenergic system. J. Psychopharmacol. 2013, 27, 659–693. [Google Scholar] [CrossRef]
- Morgan, P.J. The pars tuberalis of the pituitary: A gateway for neuroendocrine output. Rev. Reprod. 1996, 1, 153–161. [Google Scholar] [CrossRef]
- Khodai, T.; Luckman, S.M. Ventromedial Nucleus of the Hypothalamus Neurons Under the Magnifying Glass. Endocrinology 2021, 162, bqab141. [Google Scholar] [CrossRef]
- Ikeda, Y.; Luo, X.; Abbud, R.; Nilson, J.H.; Parker, K.L. The nuclear receptor steroidogenic factor 1 is essential for the formation of the ventromedial hypothalamic nucleus. Mol. Endocrinol. 1995, 9, 478–486. [Google Scholar] [CrossRef]
- Kim, D.-W.; Yao, Z.; Graybuck, L.T.; Kim, T.K.; Nguyen, T.N.; Smith, K.A.; Fong, O.; Yi, L.; Koulena, N.; Pierson, N.; et al. Multimodal Analysis of Cell Types in a Hypothalamic Node Controlling Social Behavior. Cell 2019, 179, 713–728.e17. [Google Scholar] [CrossRef]
- Cheung, C.C.; Kurrasch, D.M.; Liang, J.K.; Ingraham, H.A. Genetic labeling of steroidogenic factor-1 (SF-1) neurons in mice reveals ventromedial nucleus of the hypothalamus (VMH) circuitry beginning at neurogenesis and development of a separate non-SF-1 neuronal cluster in the ventrolateral VMH. J. Comp. Neurol. 2013, 521, 1268–1288. [Google Scholar] [CrossRef]
- Hetherington, A.W.; Ranson, S.W. The relation of various hypothalamic lesions to adiposity in the rat. J. Comp. Neurol. 1942, 76, 475–499. [Google Scholar] [CrossRef]
- Dhillon, H.; Zigman, J.M.; Ye, C.; Lee, C.E.; McGovern, R.A.; Tang, V.; Kenny, C.D.; Christiansen, L.M.; White, R.D.; Edelstein, E.A.; et al. Leptin Directly Activates SF1 Neurons in the VMH, and This Action by Leptin Is Required for Normal Body-Weight Homeostasis. Neuron 2006, 49, 191–203. [Google Scholar] [CrossRef]
- Majdic, G.; Young, M.; Gomez-Sanchez, E.; Anderson, P.; Szczepaniak, L.S.; Dobbins, R.L.; McGarry, J.D.; Parker, K.L. Knockout Mice Lacking Steroidogenic Factor 1 Are a Novel Genetic Model of Hypothalamic Obesity. Endocrinology 2002, 143, 607–614. [Google Scholar] [CrossRef]
- Viskaitis, P.; Irvine, E.E.; Smith, M.A.; Choudhury, A.I.; Alvarez-Curto, E.; Glegola, J.A.; Hardy, D.G.; Pedroni, S.M.; Pessoa, M.R.P.; Fernando, A.B.; et al. Modulation of SF1 Neuron Activity Coordinately Regulates Both Feeding Behavior and Associated Emotional States. Cell Rep. 2017, 21, 3559–3572. [Google Scholar] [CrossRef]
- Tong, Q.; Ye, C.; McCrimmon, R.J.; Dhillon, H.; Choi, B.; Kramer, M.D.; Yu, J.; Yang, Z.; Christiansen, L.M.; Lee, C.E.; et al. Synaptic glutamate release by ventromedial hypothalamic neurons is part of the neurocircuitry that prevents hypoglycemia. Cell Metab. 2007, 5, 383–393. [Google Scholar] [CrossRef]
- Song, Z.; Levin, B.E.; McArdle, J.J.; Bakhos, N.; Routh, V.H. Convergence of pre- and postsynaptic influences on glucosensing neurons in the ventromedial hypothalamic nucleus. Diabetes 2001, 50, 2673–2681. [Google Scholar] [CrossRef] [PubMed]
- Khodai, T.; Nunn, N.; Worth, A.A.; Feetham, C.H.; Belle, M.; Piggins, H.D.; Luckman, S.M. PACAP Neurons in the Ventromedial Hypothalamic Nucleus Are Glucose Inhibited and Their Selective Activation Induces Hyperglycaemia. Front. Endocrinol. 2018, 9, 632. [Google Scholar] [CrossRef] [PubMed]
- Fioramonti, X.; Marsollier, N.; Song, Z.; Fakira, K.A.; Patel, R.M.; Brown, S.; Duparc, T.; Pica-Mendez, A.; Sanders, N.M.; Knauf, C.; et al. Ventromedial Hypothalamic Nitric Oxide Production Is Necessary for Hypoglycemia Detection and Counterregulation. Diabetes 2010, 59, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Faber, C.L.; Matsen, M.E.; Velasco, K.R.; Damian, V.; Phan, B.A.; Adam, D.; Therattil, A.; Schwartz, M.W.; Morton, G.J. Distinct Neuronal Projections from the Hypothalamic Ventromedial Nucleus Mediate Glycemic and Behavioral Effects. Diabetes 2018, 67, 2518–2529. [Google Scholar] [CrossRef]
- Bentsen, M.A.; Mirzadeh, Z.; Schwartz, M.W. Revisiting How the Brain Senses Glucose—And Why. Cell Metab. 2019, 29, 11–17. [Google Scholar] [CrossRef]
- Sutton, A.K.; Goforth, P.B.; Gonzalez, I.E.; Dell’orco, J.; Pei, H.; Myers, M.G., Jr.; Olson, D.P. Melanocortin 3 receptor-expressing neurons in the ventromedial hypothalamus promote glucose disposal. Proc. Natl. Acad. Sci. USA 2021, 118, e2103090118. [Google Scholar] [CrossRef]
- Mathews, D.; Edwards, D.A. Involvement of the ventromedial and anterior hypothalamic nuclei in the hormonal induction of receptivity in the female rat. Physiol. Behav. 1977, 19, 319–326. [Google Scholar] [CrossRef]
- Lee, H.; Kim, D.-W.; Remedios, R.; Anthony, T.E.; Chang, A.; Madisen, L.; Zeng, H.; Anderson, D.J. Scalable control of mounting and attack by Esr1+ neurons in the ventromedial hypothalamus. Nature 2014, 509, 627–632. [Google Scholar] [CrossRef]
- Hashikawa, K.; Hashikawa, Y.; Falkner, A.; Lin, D. The neural circuits of mating and fighting in male mice. Curr. Opin. Neurobiol. 2016, 38, 27–37. [Google Scholar] [CrossRef]
- Kunwar, P.S.; Zelikowsky, M.; Remedios, R.; Cai, H.; Yilmaz, M.; Meister, M.; Anderson, D.J. Ventromedial hypothalamic neurons control a defensive emotion state. eLife 2015, 4, e06633. [Google Scholar] [CrossRef]
- Todd, W.D.; Fenselau, H.; Wang, J.L.; Zhang, R.; Machado, N.L.; Venner, A.; Broadhurst, R.Y.; Kaur, S.; Lynagh, T.; Olson, D.P.; et al. A hypothalamic circuit for the circadian control of aggression. Nat. Neurosci. 2018, 21, 717–724. [Google Scholar] [CrossRef]
- Falkner, A.L.; Wei, D.; Song, A.; Watsek, L.W.; Chen, I.; Chen, P.; Feng, J.E.; Lin, D. Hierarchical Representations of Aggression in a Hypothalamic-Midbrain Circuit. Neuron 2020, 106, 637–648.e6. [Google Scholar] [CrossRef]
- Pfaff, D.W.; Sakuma, Y. Deficit in the lordosis reflex of female rats caused by lesions in the ventromedial nucleus of the hypothalamus. J. Physiol. 1979, 288, 203–210. [Google Scholar] [CrossRef]
- Goy, R.W.; Phoenix, C.H. Hypothalamic regulation of female sexual behaviour; establishment of behavioural oestrus in spayed guinea-pigs following hypothalamic lesions. Reproduction 1963, 5, 23. [Google Scholar] [CrossRef]
- Pfaff, D.W.; Sakuma, Y. Facilitation of the lordosis reflex of female rats from the ventromedial nucleus of the hypothalamus. J. Physiol. 1979, 288, 189–202. [Google Scholar] [CrossRef]
- Powers, J. Hormonal control of sexual receptivity during the estrous cycle of the rat. Physiol. Behav. 1970, 5, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.I.; Pfaff, D.W. Characterization of estrogen-concentrating hypothalamic neurons by their axonal projections. Science 1982, 217, 1273–1276. [Google Scholar] [CrossRef] [PubMed]
- Arletti, R.; Bertolini, A. Oxytocin stimulates lordosis behavior in female rats. Neuropeptides 1985, 6, 247–253. [Google Scholar] [CrossRef]
- Rachman, I.M.; Unnerstall, J.R.; Pfaff, D.W.; Cohen, R.S. Regulation of neuronal nitric oxide synthase mRNA in lordosis-relevant neurons of the ventromedial hypothalamus following short-term estrogen treatment. Brain Research. Mol. Brain Res. 1998, 59, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Hashikawa, K.; Hashikawa, Y.; Tremblay, R.; Zhang, J.; Feng, J.E.; Sabol, A.; Piper, W.T.; Lee, H.; Rudy, B.; Lin, D. Esr1+ cells in the ventromedial hypothalamus control female aggression. Nat. Neurosci. 2017, 20, 1580–1590. [Google Scholar] [CrossRef]
- Bellinger, L.L.; Bernardis, L.L. The dorsomedial hypothalamic nucleus and its role in ingestive behavior and body weight regulation: Lessons learned from lesioning studies. Physiol. Behav. 2002, 76, 431–442. [Google Scholar] [CrossRef]
- Bernardis, L.L.; Bellinger, L.L. The dorsomedial hypothalamic nucleus revisited: 1998 update. Proceedings of the Society for Experimental Biology and Medicine. Soc. Exp. Biol. Med. 1998, 218, 284–306. [Google Scholar] [CrossRef]
- Bernardis, L.L.; Bellinger, L.L. The dorsomedial hypothalamic nucleus revisited: 1986 update. Brain Res. Rev. 1987, 434, 321–381. [Google Scholar] [CrossRef]
- Thompson, R.; Swanson, L.W. Organization of inputs to the dorsomedial nucleus of the hypothalamus: A reexamination with Fluorogold and PHAL in the rat. Brain Res. Rev. 1998, 27, 89–118. [Google Scholar] [CrossRef]
- Thompson, R.; Canteras, N.; Swanson, L. Organization of projections from the dorsomedial nucleus of the hypothalamus: A PHA-L study in the rat. J. Comp. Neurol. 1996, 376, 143–173. [Google Scholar] [CrossRef]
- Thompson, R.H.; Swanson, L.W. Structural characterization of a hypothalamic visceromotor pattern generator network. Brain Res. Rev. 2003, 41, 153–202. [Google Scholar] [CrossRef]
- Chou, T.C.; Scammell, T.E.; Gooley, J.J.; Gaus, S.E.; Saper, C.B.; Lu, J. Critical Role of Dorsomedial Hypothalamic Nucleus in a Wide Range of Behavioral Circadian Rhythms. J. Neurosci. 2003, 23, 10691–10702. [Google Scholar] [CrossRef]
- Binder, M.D.; Hirokawa, N.; Windhorst, U. (Eds.) Subparaventricular Zone. In Encyclopedia of Neuroscience; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, Y.-H.; Chou, T.C.; Gaus, S.E.; Elmquist, J.K.; Shiromani, P.; Saper, C.B. Contrasting Effects of Ibotenate Lesions of the Paraventricular Nucleus and Subparaventricular Zone on Sleep–Wake Cycle and Temperature Regulation. J. Neurosci. 2001, 21, 4864–4874. [Google Scholar] [CrossRef]
- Elmquist, J.K.; Ahima, R.S.; Elias, C.F.; Flier, J.S.; Saper, C.B. Leptin activates distinct projections from the dorsomedial and ventromedial hypothalamic nuclei. Proc. Natl. Acad. Sci. USA 1998, 95, 741–746. [Google Scholar] [CrossRef]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef]
- Cho, H.; Zhao, X.; Hatori, M.; Yu, R.T.; Barish, G.D.; Lam, M.T.; Chong, L.-W.; DiTacchio, L.; Atkins, A.R.; Glass, C.K.; et al. Regulation of circadian behaviour and metabolism by REV-ERB-α and REV-ERB-β. Nature 2012, 485, 123–127. [Google Scholar] [CrossRef]
- Noguchi, T.; Leise, T.L.; Kingsbury, N.J.; Diemer, T.; Wang, L.L.; Henson, M.A.; Welsh, D.K. Calcium Circadian Rhythmicity in the Suprachiasmatic Nucleus: Cell Autonomy and Network Modulation. Eneuro 2017, 4, ENEURO.0160-17.2017. [Google Scholar] [CrossRef]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef]
- Herzog, E.D.; Aton, S.; Numano, R.; Sakaki, Y.; Tei, H. Temporal Precision in the Mammalian Circadian System: A Reliable Clock from Less Reliable Neurons. J. Biol. Rhythm. 2004, 19, 35–46. [Google Scholar] [CrossRef]
- Patton, A.P.; Chesham, J.E.; Hastings, M.H. Combined Pharmacological and Genetic Manipulations Unlock Unprecedented Temporal Elasticity and Reveal Phase-Specific Modulation of the Molecular Circadian Clock of the Mouse Suprachiasmatic Nucleus. J. Neurosci. 2016, 36, 9326–9341. [Google Scholar] [CrossRef] [PubMed]
- Colwell, C.S. Linking neural activity and molecular oscillations in the SCN. Nat. Rev. Neurosci. 2011, 12, 553–569. [Google Scholar] [CrossRef] [PubMed]
- Meijer, J.H.; Michel, S. Neurophysiological analysis of the suprachiasmatic nucleus: A challenge at multiple levels. Methods Enzymol. 2015, 552, 75–102. [Google Scholar] [CrossRef]
- Hofman, M.; Zhou, J.-N.; Swaab, D. Suprachiasmatic nucleus of the human brain: An immunocytochemical and morphometric analysis. Anat. Rec. 1996, 244, 552–562. [Google Scholar] [CrossRef]
- Abrahamson, E.E.; Moore, R.Y. Suprachiasmatic nucleus in the mouse: Retinal innervation, intrinsic organization and efferent projections. Brain Res. 2001, 916, 172–191. [Google Scholar] [CrossRef]
- Moore, R.Y.; Speh, J.C.; Leak, R.K. Suprachiasmatic nucleus organization. Cell Tissue Res. 2002, 309, 89–98. [Google Scholar] [CrossRef]
- Ebling, F.J. The role of glutamate in the photic regulation of the suprachiasmatic nucleus. Prog. Neurobiol. 1996, 50, 109–132. [Google Scholar] [CrossRef]
- Park, J.; Zhu, H.; O’Sullivan, S.; Ogunnaike, B.A.; Weaver, D.R.; Schwaber, J.S.; Vadigepalli, R. Single-Cell Transcriptional Analysis Reveals Novel Neuronal Phenotypes and Interaction Networks Involved in the Central Circadian Clock. Front. Neurosci. 2016, 10, 481. [Google Scholar] [CrossRef]
- Hastings, M.H.; Brancaccio, M.; Maywood, E.S. Circadian pacemaking in cells and circuits of the suprachiasmatic nucleus. J. Neuroendocr. 2014, 26, 2–10. [Google Scholar] [CrossRef]
- Brancaccio, M.; Maywood, E.S.; Chesham, J.E.; Loudon, A.S.; Hastings, M.H. A Gq-Ca2+ Axis Controls Circuit-Level Encoding of Circadian Time in the Suprachiasmatic Nucleus. Neuron 2013, 78, 714–728. [Google Scholar] [CrossRef]
- Montminy, M. Transcriptional regulation by cyclic AMP. Annu. Rev. Biochem. 1997, 66, 807–822. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.A.; Leise, T.L.; Castanon-Cervantes, O.; Davidson, A.J. Dynamic interactions mediated by nonredundant signaling mechanisms couple circadian clock neurons. Neuron 2013, 80, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Hanna, L.; Harding, C.; Hayter, E.A.; Walmsley, L.; Bechtold, D.A.; Brown, T.M. Output from VIP cells of the mammalian central clock regulates daily physiological rhythms. Nat. Commun. 2020, 11, 1453. [Google Scholar] [CrossRef] [PubMed]
- Biag, J.; Huang, Y.; Gou, L.; Hintiryan, H.; Askarinam, A.; Hahn, J.D.; Toga, A.W.; Dong, H.-W. Cyto- and chemoarchitecture of the hypothalamic paraventricular nucleus in the C57BL/6J male mouse: A study of immunostaining and multiple fluorescent tract tracing. J. Comp. Neurol. 2012, 520, 6–33. [Google Scholar] [CrossRef]
- Swanson, L.W.; Kuypers, H.G.J.M. The paraventricular nucleus of the hypothalamus: Cytoarchitectonic subdivisions and organization of projections to the pituitary, dorsal vagal complex, and spinal cord as demonstrated by retrograde fluorescence double-labeling methods. J. Comp. Neurol. 1980, 194, 555–570. [Google Scholar] [CrossRef]
- Swanson, L.; Sawchenko, P. Paraventricular nucleus: A site for the integration of neuroendocrine and autonomic mechanisms. Neuroendocrinology 1980, 31, 410–417. [Google Scholar] [CrossRef]
- Swanson, L.; Sawchenko, P.; Wiegand, S.; Price, J. Separate neurons in the paraventricular nucleus project to the median eminence and to the medulla or spinal cord. Brain Res. 1980, 198, 190–195. [Google Scholar] [CrossRef]
- Swanson, L.W.; E Sawchenko, P. Hypothalamic Integration: Organization of the Paraventricular and Supraoptic Nuclei. Annu. Rev. Neurosci. 1983, 6, 269–324. [Google Scholar] [CrossRef]
- Franklin, K.B.J.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Simmons, D.M.; Swanson, L.W. Comparison of the spatial distribution of seven types of neuroendocrine neurons in the rat paraventricular nucleus: Toward a global 3D model. J. Comp. Neurol. 2009, 516, 423–441. [Google Scholar] [CrossRef]
- Sawchenko, P.E.; Swanson, L.W. Immunohistochemical identification of neurons in the paraventricular nucleus of the hypothalamus that project to the medulla or to the spinal cord in the rat. J. Comp. Neurol. 1982, 205, 260–272. [Google Scholar] [CrossRef]
- Sawchenko, P.E.; Swanson, L.W. Localization, colocalization, and plasticity of corticotropin-releasing factor immunoreactivity in rat brain. Fed. Proc. 1985, 44, 221–227. [Google Scholar]
- Sawchenko, P.E.; Swanson, L.W.; Vale, W.W. Co-expression of corticotropin-releasing factor and vasopressin immunoreactivity in parvocellular neurosecretory neurons of the adrenalectomized rat. Proc. Natl. Acad. Sci. USA 1984, 81, 1883–1887. [Google Scholar] [CrossRef] [PubMed]
- Sawchenko, P.; Swanson, L.; Vale, W. Corticotropin-releasing factor: Co-expression within distinct subsets of oxytocin-, vasopressin-, and neurotensin-immunoreactive neurons in the hypothalamus of the male rat. J. Neurosci. 1984, 4, 1118–1129. [Google Scholar] [CrossRef] [PubMed]
- Mouri, T.; Itoi, K.; Takahashi, K.; Suda, T.; Murakami, O.; Yoshinaga, K.; Andoh, N.; Ohtani, H.; Masuda, T.; Sasano, N. Colocalization of Corticotropin-Releasing Factor and Vasopressin in the Paraventricular Nucleus of the Human Hypothalamus. Neuroendocrinology 1993, 57, 34–39. [Google Scholar] [CrossRef]
- Pyner, S.; Coote, J.H. Identification of branching paraventricular neurons of the hypothalamus that project to the rostroventrolateral medulla and spinal cord. Neuroscience 2000, 100, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Stocker, S.; Simmons, J.R.; Stornetta, R.; Toney, G.M.; Guyenet, P.G. Water deprivation activates a glutamatergic projection from the hypothalamic paraventricular nucleus to the rostral ventrolateral medulla. J. Comp. Neurol. 2006, 494, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Biancardi, V.C.; Campos, R.R.; Stern, J.E. Altered balance of gamma-aminobutyric acidergic and glutamatergic afferent inputs in rostral ventrolateral medulla-projecting neurons in the paraventricular nucleus of the hypothalamus of renovascular hypertensive rats. J. Comp. Neurol. 2010, 518, 567–585. [Google Scholar] [CrossRef]
- Geerling, J.C.; Shin, J.-W.; Chimenti, P.C.; Loewy, A.D. Paraventricular hypothalamic nucleus: Axonal projections to the brainstem. J. Comp. Neurol. 2010, 518, 1460–1499. [Google Scholar] [CrossRef]
- Loewy, A.D. Central autonomic pathways. In Central Regulation of Autonomic Functions; Loewy, A.D., Spyer, K.M., Eds.; Oxford University Press: Oxford, England, 1990; pp. 88–103. [Google Scholar]
- Mosqueda-Garcia, R. Central autonomic regulation. In Primer on the Autonomic Nervous System; Robertson, D., Low, P.A., Polinsky, R.J., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 3–12. [Google Scholar]
- Ferguson, A.V.; Latchford, K.J.; Samson, W.K. The paraventricular nucleus of the hypothalamus—A potential target for integrative treatment of autonomic dysfunction. Expert Opin. Ther. Targets 2008, 12, 717–727. [Google Scholar] [CrossRef]
- Jiang, Z.; Rajamanickam, S.; Justice, N.J. Local Corticotropin-Releasing Factor Signaling in the Hypothalamic Paraventricular Nucleus. J. Neurosci. 2018, 38, 1874–1890. [Google Scholar] [CrossRef]
- Jiang, Z.; Rajamanickam, S.; Justice, N.J. CRF signaling between neurons in the paraventricular nucleus of the hypothalamus (PVN) coordinates stress responses. Neurobiol. Stress 2019, 11, 100192. [Google Scholar] [CrossRef]
- Jones, J.R.; Chaturvedi, S.; Granados-Fuentes, D.; Herzog, E.D. Circadian neurons in the paraventricular nucleus entrain and sustain daily rhythms in glucocorticoids. Nat. Commun. 2021, 12, 5763. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.Y.; Eichler, V.B. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res. 1972, 42, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Husse, J.; Leliavski, A.; Tsang, A.H.; Oster, H.; Eichele, G. The light-dark cycle controls peripheral rhythmicity in mice with a genetically ablated suprachiasmatic nucleus clock. FASEB J. 2014, 28, 4950–4960. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Chaker, Z.; Doetsch, F. Hypothalamic regulation of regionally distinct adult neural stem cells and neurogenesis. Science 2017, 356, 1383–1386. [Google Scholar] [CrossRef]
- Minocha, S.; Valloton, D.; Arsenijevic, Y.; Cardinaux, J.-R.; Guidi, R.; Hornung, J.-P.; Lebrand, C. Nkx2.1 regulates the generation of telencephalic astrocytes during embryonic development. Sci. Rep. 2017, 7, 43093. [Google Scholar] [CrossRef]
- Orquera, D.P.; Tavella, M.B.; de Souza, F.S.; Nasif, S.; Low, M.J.; Rubinstein, M. The Homeodomain Transcription Factor NKX2.1 Is Essential for the Early Specification of Melanocortin Neuron Identity and ActivatesPomcExpression in the Developing Hypothalamus. J. Neurosci. 2019, 39, 4023–4035. [Google Scholar] [CrossRef]
- Li, Y.; Bao, H.; Luo, Y.; Yoan, C.; Sullivan, H.A.; Quintanilla, L.; Wickersham, I.; Lazarus, M.; Shih, Y.-Y.I.; Song, J. Supramammillary nucleus synchronizes with dentate gyrus to regulate spatial memory retrieval through glutamate release. eLife 2020, 9, e53129. [Google Scholar] [CrossRef]
- Pedersen, N.P.; Ferrari, L.; Venner, A.; Wang, J.L.; Abbott, S.B.G.; Vujovic, N.; Arrigoni, E.; Saper, C.B.; Fuller, P.M. Supramammillary glutamate neurons are a key node of the arousal system. Nat. Commun. 2017, 8, 1405. [Google Scholar] [CrossRef]
- Hashimotodani, Y.; Karube, F.; Yanagawa, Y.; Fujiyama, F.; Kano, M. Supramammillary Nucleus Afferents to the Dentate Gyrus Co-release Glutamate and GABA and Potentiate Granule Cell Output. Cell Rep. 2018, 25, 2704–2715.e4. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Chen, C.-C.; Huang, C.-C.; Nishimori, K.; Hsu, K.-S. Oxytocin stimulates hippocampal neurogenesis via oxytocin receptor expressed in CA3 pyramidal neurons. Nat. Commun. 2017, 8, 537. [Google Scholar] [CrossRef] [PubMed]
- Koutmani, Y.; Gampierakis, I.A.; Polissidis, A.; Ximerakis, M.; Koutsoudaki, P.N.; Polyzos, A.; Agrogiannis, G.; Karaliota, S.; Thomaidou, D.; Rubin, L.; et al. CRH Promotes the Neurogenic Activity of Neural Stem Cells in the Adult Hippocampus. Cell Rep. 2019, 29, 932–945.e7. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, D.; Neri, L.; Canalini, F.; Calevro, A.; Ottani, A.; Vandini, E.; Sena, P.; Zaffe, D.; Guarini, S. NDP-α-MSH induces intense neurogenesis and cognitive recovery in Alzheimer transgenic mice through activation of melanocortin MC4 receptors. Mol. Cell Neurosci. 2015, 67, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Spaccapelo, L.; Galantucci, M.; Neri, L.; Contri, M.; Pizzala, R.; D’Amico, R.; Ottani, A.; Sandrini, M.; Zaffe, D.; Giuliani, D.; et al. Up-regulation of the canonical Wnt-3A and Sonic hedgehog signaling underlies melanocortin-induced neurogenesis after cerebral ischemia. Eur. J. Pharmacol. 2013, 707, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Yabe, T.; Gamo, Y.; Nagai, T.; Oikawa, T.; Yamada, H.; Hanawa, T.I.C.V. administration of orexin-A induces an antidepressive-like effect through hippocampal cell proliferation. Neuroscience 2008, 157, 720–732. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, R.X.; Tang, S.; Ren, Y.Y.; Yang, W.X.; Liu, X.M.; Tang, J.Y. Orexin-A-induced ERK1/2 activation reverses impaired spatial learning and memory in pentylenetetrazol-kindled rats via OX1R-mediated hippocampal neurogenesis. Peptides 2014, 54, 140–147. [Google Scholar] [CrossRef]
- Fry, M.; Hoyda, T.D.; Ferguson, A.V. Making sense of it: Roles of the sensory circumventricular organs in feeding and regulation of energy homeostasis. Exp. Biol. Med. 2007, 232, 14–26. [Google Scholar]
- Benarroch, E.E. Circumventricular organs: Receptive and homeostatic functions and clinical implications. Neurology 2011, 77, 1198–1204. [Google Scholar] [CrossRef]
- Morita, S.; Miyata, S. Different vascular permeability between the sensory and secretory circumventricular organs of adult mouse brain. Cell Tissue Res. 2012, 349, 589–603. [Google Scholar] [CrossRef]
- Hicks, A.-I.; Kobrinsky, S.; Zhou, S.; Yang, J.; Prager-Khoutorsky, M. Anatomical Organization of the Rat Subfornical Organ. Front. Cell Neurosci. 2021, 15, 691711. [Google Scholar] [CrossRef]
- Okamoto, A.; Fujii, R.; Yoshimura, R.; Miyata, S. Transcytosis of tanycytes in the circumventricular organs of adult mouse brain. Neurosci. Lett. 2022, 779, 136633. [Google Scholar] [CrossRef]
- Langlet, F.; Mullier, A.; Bouret, S.G.; Prevot, V.; Dehouck, B. Tanycyte-like cells form a blood-cerebrospinal fluid barrier in the circumventricular organs of the mouse brain. J. Comp. Neurol. 2013, 521, 3389–3405. [Google Scholar] [CrossRef] [PubMed]
- Ayloo, S.; Gu, C. Transcytosis at the blood–brain barrier. Curr. Opin. Neurobiol. 2019, 57, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Balland, E.; Dam, J.; Langlet, F.; Caron, E.; Steculorum, S.; Messina, A.; Rasika, S.; Falluel-Morel, A.; Anouar, Y.; Dehouck, B.; et al. Hypothalamic tanycytes are an ERK-gated conduit for leptin into the brain. Cell Metab. 2014, 19, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Miyata, S. Glial functions in the blood-brain communication at the circumventricular organs. Front. Neurosci. 2022, 16, 991779. [Google Scholar] [CrossRef]
- Vidal-Itriago, A.; Radford, R.A.W.; Aramideh, J.A.; Maurel, C.; Scherer, N.M.; Don, E.K.; Lee, A.; Chung, R.S.; Graeber, M.B.; Morsch, M. Microglia morphophysiological diversity and its implications for the CNS. Front. Immunol. 2022, 13, 997786. [Google Scholar] [CrossRef] [PubMed]
- McKinley, M.J.; McAllen, R.; Davern, P.; Giles, M.E.; Penschow, J.; Sunn, N.; Uschakov, A.; Oldfield, B. The sensory circumventricular organs of the mammalian brain. Adv. Anat. Embryol. Cell Biol. 2003, 172. [Google Scholar] [CrossRef]
- Bennett, L.; Yang, M.; Enikolopov, G.; Iacovitti, L. Circumventricular organs: A novel site of neural stem cells in the adult brain. Mol. Cell Neurosci. 2009, 41, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Furube, E.; Morita, M.; Miyata, S. Characterization of neural stem cells and their progeny in the sensory circumventricular organs of adult mouse. Cell Tissue Res. 2015, 362, 347–365. [Google Scholar] [CrossRef]
- Furube, E.; Ishii, H.; Nambu, Y.; Kurganov, E.; Nagaoka, S.; Morita, M.; Miyata, S. Neural stem cell phenotype of tanycyte-like ependymal cells in the circumventricular organs and central canal of adult mouse brain. Sci. Rep. 2020, 10, 2826. [Google Scholar] [CrossRef]
- Sanin, V.; Heeß, C.; Kretzschmar, H.A.; Schüller, U. Recruitment of neural precursor cells from circumventricular organs of patients with cerebral ischaemia. Neuropathol. Appl. Neurobiol. 2013, 39, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Haan, N.; Goodman, T.; Najdi-Samiei, A.; Stratford, C.M.; Rice, R.; El Agha, E.; Bellusci, S.; Hajihosseini, M.K. Fgf10-expressing tanycytes add new neurons to the appetite/energy-balance regulating centers of the postnatal and adult hypothalamus. J. Neurosci. 2013, 33, 6170–6180. [Google Scholar] [CrossRef] [PubMed]
- Robins, S.; Stewart, I.; McNay, D.; Taylor, V.; Giachino, C.; Goetz, M.; Ninkovic, J.; Briancon, N.; Maratos-Flier, E.; Flier, J.; et al. α-Tanycytes of the adult hypothalamic third ventricle include distinct populations of FGF-responsive neural progenitors. Nat. Commun. 2013, 4, 2049. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tang, Y.; Cai, D. IKKb/NF-kB disrupts adult hypothalamic neural stem cells to mediate a neurodegenerative-mechanism of dietary obesity and pre-diabetes. Nat. Cell Biol. 2012, 14, 999–1012. [Google Scholar] [CrossRef]
- Lee, D.; Bedont, J.L.; Pak, T.; Wang, H.; Song, J.; Miranda-Angulo, A.; Takiar, V.; Charubhumi, V.; Balordi, F.; Takebayashi, H.; et al. Tanycytes of the hypothalamic median eminence form a diet-responsive neurogenic niche. Nat. Neurosci. 2012, 15, 700–702. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Guo, Q.; Scheller, A. Progenies of NG2 glia: What do we learn from transgenic mouse models? Neural Regen. Res. 2021, 16, 43–48. [Google Scholar] [CrossRef]
- Pérez-Martín, M.; Cifuentes, M.; Grondona, J.M.; López-Ávalos, M.D.; Gómez-Pinedo, U.; García-Verdugo, J.M.; Fernández-Llebrez, P. IGF-I stimulates neurogenesis in the hypothalamus of adult rats. Eur. J. Neurosci. 2010, 31, 1533–1548. [Google Scholar] [CrossRef]
- Son, J.E.; Dou, Z.; Kim, K.-H.; Wanggou, S.; Bin Cha, V.S.; Mo, R.; Zhang, X.; Chen, X.; Ketela, T.; Li, X.; et al. Irx3 and Irx5 in Ins2-Cre+ cells regulate hypothalamic postnatal neurogenesis and leptin response. Nat. Metab. 2021, 3, 701–713. [Google Scholar] [CrossRef]
- Mullier, A.; Bouret, S.G.; Prevot, V.; Dehouck, B. Differential distribution of tight junction proteins suggests a role for tanycytes in blood-hypothalamus barrier regulation in the adult mouse brain. J. Comp. Neurol. 2010, 518, 943–962. [Google Scholar] [CrossRef]
- Zabini, A.; Zimmer, Y.; Medová, M. Beyond keratinocyte differentiation: Emerging new biology of small proline-rich proteins. Trends Cell Biol. 2023, 33, 5–8. [Google Scholar] [CrossRef]
- Langlet, F.; Levin, B.E.; Luquet, S.; Mazzone, M.; Messina, A.; Dunn-Meynell, A.A.; Balland, E.; Lacombe, A.; Mazur, D.; Carmeliet, P.; et al. Tanycytic VEGF-A Boosts Blood-Hypothalamus Barrier Plasticity and Access of Metabolic Signals to the Arcuate Nucleus in Response to Fasting. Cell Metab. 2013, 17, 607–617. [Google Scholar] [CrossRef]
- Mirzadeh, Z.; Kusne, Y.; Duran-Moreno, M.; Cabrales, E.; Gil-Perotin, S.; Ortiz, C.; Chen, B.; Garcia-Verdugo, J.M.; Sanai, N.; Alvarez-Buylla, A. Bi- and uniciliated ependymal cells define continuous floor-plate-derived tanycytic territories. Nat. Commun. 2017, 8, 13759. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, O.; Tanahashi, M.; Yokoi, S.; Kaneko, M.; Yanaka, K.; Nakagawa, S.; Maita, H. The cell type-specific ER membrane protein UGS148 is not essential in mice. Genes Cells 2022, 27, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Chaker, Z.; George, C.; Petrovska, M.; Caron, J.-B.; Lacube, P.; Caillé, I.; Holzenberger, M. Hypothalamic neurogenesis persists in the aging brain and is controlled by energy-sensing IGF-I pathway. Neurobiol. Aging 2016, 41, 64–72. [Google Scholar] [CrossRef]
- Traiffort, E.; Charytoniuk, D.; Watroba, L.; Faure, H.; Sales, N.; Ruat, M. Discrete localizations of hedgehog signalling components in the developing and adult rat nervous system. Eur. J. Neurosci. 1999, 11, 3199–3214. [Google Scholar] [CrossRef]
- Shimizu, T.; Hibi, M. Formation and patterning of the forebrain and olfactory system by zinc-finger genes Fezf1 and Fezf2. Dev. Growth Differ. 2009, 51, 221–231. [Google Scholar] [CrossRef]
- Berberoglu, M.A.; Dong, Z.; Li, G.; Zheng, J.; Martinez, L.d.C.G.T.; Peng, J.; Wagle, M.; Reichholf, B.; Petritsch, C.; Li, H.; et al. Heterogeneously expressed fezf2 patterns gradient Notch activity in balancing the quiescence, proliferation, and dif-ferentiation of adult neural stem cells. J. Neurosci. 2014, 34, 13911–13923. [Google Scholar] [CrossRef]
- Bolborea, M.; Langlet, F. What is the physiological role of hypothalamic tanycytes in metabolism? Am. J. Physiol. Integr. Comp. Physiol. 2021, 320, R994–R1003. [Google Scholar] [CrossRef] [PubMed]
- Benford, H.; Bolborea, M.; Pollatzek, E.; Lossow, K.; Hermans-Borgmeyer, I.; Liu, B.; Meyerhof, W.; Kasparov, S.; Dale, N. A sweet taste receptor-dependent mechanism of glucosensing in hypothalamic tanycytes. Glia 2017, 65, 773–789. [Google Scholar] [CrossRef]
- Elizondo-Vega, R.; Cortes-Campos, C.; Barahona, M.J.; Oyarce, K.A.; Carril, C.A.; García-Robles, M.A. The role of tanycytes in hypothalamic glucosensing. J. Cell Mol. Med. 2015, 19, 1471–1482. [Google Scholar] [CrossRef]
- Palma-Chavez, A.; Konar-Nié, M.; Órdenes, P.; Maurelia, F.; Elizondo-Vega, R.; Oyarce, K.; López, S.; Rojas, J.; Steinberg, X.; García-Robles, M.A.; et al. Glucose Increase DAGLα Levels in Tanycytes and Its Inhibition Alters Orexigenic and Anorexigenic Neuropeptides Expression in Response to Glucose. Front. Endocrinol. 2019, 10, 647. [Google Scholar] [CrossRef] [PubMed]
- Schuit, F.C.; Huypens, P.; Heimberg, H.; Pipeleers, D.G. Glucose sensing in pancreatic beta-cells: A model for the study of other glucose-regulated cells in gut, pancreas, and hypothalamus. Diabetes 2001, 50, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sanders, N.M.; Dunn-Meynell, A.A.; Levin, B.E. Third ventricular alloxan reversibly impairs glucose counterregulatory re-sponses. Diabetes 2004, 53, 1230–1236. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fekete, C.; Lechan, R.M. Central regulation of hypothalamic-pituitary-thyroid axis under physiological and pathophysiological conditions. Endocr. Rev. 2014, 35, 159–194. [Google Scholar] [CrossRef]
- Diano, S.; Leonard, J.L.; Meli, R.; Esposito, E.; Schiavo, L. Hypothalamic type II iodothyronine deiodinase: A light and electron microscopic study. Brain Res. 2003, 976, 130–134. [Google Scholar] [CrossRef]
- Guadaño-Ferraz, A.; Obregon, M.J.; Germain, D.L.S.; Bernal, J. The type 2 iodothyronine deiodinase is expressed primarily in glial cells in the neonatal rat brain. Proc. Natl. Acad. Sci. USA 1997, 94, 10391–10396. [Google Scholar] [CrossRef]
- Tu, H.M.; Kim, S.-W.; Salvatore, D.; Bartha, T.; Legradi, G.; Larsen, P.R.; Lechan, R.M. Regional distribution of type 2 thyroxine deiodinase messenger ribonucleic acid in rat hypothalamus and pituitary and its regulation by thyroid hormone. Endocrinology 1997, 138, 3359–3368. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, T.L.; Correa-Medina, M.; Campos, M.P.; Wittmann, G.; Werneck-De-Castro, J.P.; Arrojo e Drigo, R.; Mora-Garzon, M.; Ueta, C.B.; Caicedo, A.; Fekete, C.; et al. Coordination of hypothalamic and pituitary T3 production regulates TSH expression. J. Clin. Investig. 2013, 123, 1492–1500. [Google Scholar] [CrossRef]
- Sánchez, E.; Vargas, M.A.; Singru, P.S.; Pascual, I.; Romero, F.; Fekete, C.; Charli, J.-L.; Lechan, R.M. Tanycyte Pyroglutamyl Peptidase II Contributes to Regulation of the Hypothalamic-Pituitary-Thyroid Axis through Glial-Axonal Associations in the Median Eminence. Endocrinology 2009, 150, 2283–2291. [Google Scholar] [CrossRef]
- Lazcano, I.; Cabral, A.; Uribe, R.M.; Jaimes-Hoy, L.; Perello, M.; Joseph-Bravo, P.; Sánchez-Jaramillo, E.; Charli, J.-L. Fasting Enhances Pyroglutamyl Peptidase II Activity in Tanycytes of the Mediobasal Hypothalamus of Male Adult Rats. Endocrinology 2015, 156, 2713–2723. [Google Scholar] [CrossRef]
- Roberts, L.M.; Woodford, K.; Zhou, M.; Black, D.S.; Haggerty, J.E.; Tate, E.H.; Grindstaff, K.K.; Mengesha, W.; Raman, C.; Zerangue, N. Expression of the Thyroid Hormone Transporters Monocarboxylate Transporter-8 (SLC16A2) and Organic Ion Transporter-14 (SLCO1C1) at the Blood-Brain Barrier. Endocrinology 2008, 149, 6251–6261. [Google Scholar] [CrossRef] [PubMed]
- Segerson, T.P.; Kauer, J.; Wolfe, H.C.; Mobtaker, H.; Wu, P.; Jackson, I.M.D.; Lechan, R.M. Thyroid Hormone Regulates TRH Biosynthesis in the Paraventricular Nucleus of the Rat Hypothalamus. Science 1987, 238, 78–80. [Google Scholar] [CrossRef]
- Dratman, M.B.; Crutchfield, F.L.; Futaesaku, Y.; Goldberger, M.E.; Murray, M. [125I] triiodothyronine in the rat brain: Evidence for neural localization and axonal transport derived from thaw-mount film autoradiography. J. Comp. Neurol. 1987, 260, 392–408. [Google Scholar] [CrossRef]
- Diano, S.; Naftolin, F.; Goglia, F.; Horvath, T.L. Fasting-induced increase in type II iodothyronine deiodinase activity and messenger ribonucleic acid levels is not reversed by thyroxine in the rat hypothalamus. Endocrinology 1998, 139, 2879–2884. [Google Scholar] [CrossRef]
- Coppola, A.; Meli, R.; Diano, S. Inverse shift in circulating corticosterone and leptin levels elevates hypothalamic deiodinase type 2 in fasted rats. Endocrinology 2005, 146, 2827–2833. [Google Scholar] [CrossRef] [PubMed]
- Coppola, A.; Liu, Z.-W.; Andrews, Z.B.; Paradis, E.; Roy, M.-C.; Friedman, J.M.; Ricquier, D.; Richard, D.; Horvath, T.L.; Gao, X.-B.; et al. A central thermogenic-like mechanism in feeding regulation: An interplay between arcuate nucleus T3 and UCP2. Cell Metab. 2007, 5, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Coppola, A.; Hughes, J.; Esposito, E.; Schiavo, L.; Meli, R.; Diano, S. Suppression of hypothalamic deiodinase type II activity blunts TRH mRNA decline during fasting. FEBS Lett. 2005, 579, 4654–4658. [Google Scholar] [CrossRef]
- Müller-Fielitz, H.; Stahr, M.; Bernau, M.; Richter, M.; Abele, S.; Krajka, V.; Benzin, A.; Wenzel, J.; Kalies, K.; Mittag, J.; et al. Tanycytes control the hormonal output of the hypothalamic-pituitary-thyroid axis. Nat. Commun. 2017, 8, 484. [Google Scholar] [CrossRef]
- Prevot, V.; Hanchate, N.K.; Bellefontaine, N.; Sharif, A.; Parkash, J.; Estrella, C.; Allet, C.; de Seranno, S.; Campagne, C.; de Tassigny, X.D.; et al. Function-related structural plasticity of the GnRH system: A role for neuronal–glial–endothelial interactions. Front. Neuroendocr. 2010, 31, 241–258. [Google Scholar] [CrossRef]
- Prevot, V.; Dutoit, S.; Croix, D.; Tramu, G.; Beauvillain, J. Semi-quantitative ultrastructural analysis of the localization and neuropeptide content of gonadotropin releasing hormone nerve terminals in the median eminence throughout the estrous cycle of the rat. Neuroscience 1998, 84, 177–191. [Google Scholar] [CrossRef]
- Prevot, V.; Croix, D.; Bouret, S.; Dutoit, S.; Tramu, G.; Stefano, G.; Beauvillain, J. Definitive evidence for the existence of morphological plasticity in the external zone of the median eminence during the rat estrous cycle: Implication of neuro-glio-endothelial interactions in gonadotropin-releasing hormone release. Neuroscience 1999, 94, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, G.; Trubert, C.; Terrien, J.; Pifferi, F.; Leroy, D.; Loyens, A.; Migaud, M.; Baroncini, M.; Maurage, C.-A.; Fontaine, C.; et al. A comparative study of the neural stem cell niche in the adult hypothalamus of human, mouse, rat and gray mouse lemur (Microcebus murinus). J. Comp. Neurol. 2018, 526, 1419–1443. [Google Scholar] [CrossRef] [PubMed]
- Sanai, N.; Tramontin, A.D.; Quinones-Hinojosa, A.; Barbaro, N.M.; Gupta, N.; Kunwar, S.; Lawton, M.T.; McDermott, M.W.; Parsa, A.T. Unique astrocyte ribbon in adult human brain contains neural stem cells but lacks chain migration. Nature 2004, 427, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.H.; Bouchard-Cannon, P.; Hegazi, S.; Lowden, C.; Fung, S.W.; Chiang, C.-K.; Ness, R.W.; Cheng, H.-Y.M. SOX2-Dependent Transcription in Clock Neurons Promotes the Robustness of the Central Circadian Pacemaker. Cell Rep. 2019, 26, 3191–3202.e8. [Google Scholar] [CrossRef]
- Cheng, A.H.; Fung, S.W.; Hegazi, S.; Abdalla, O.H.M.H.; Cheng, H.-Y.M. SOX2 Regulates Neuronal Differentiation of the Suprachiasmatic Nucleus. Int. J. Mol. Sci. 2021, 23, 229. [Google Scholar] [CrossRef]
- McNay, D.; Briançon, N.; Kokoeva, M.V.; Maratos-Flier, E.; Flier, J.S. Remodeling of the arcuate nucleus energy-balance circuit is inhibited in obese mice. J. Clin. Investig. 2012, 122, 142–152. [Google Scholar] [CrossRef]
- Thaler, J.P.; Yi, C.-X.; Schur, E.A.; Guyenet, S.J.; Hwang, B.H.; Dietrich, M.O.; Zhao, X.; Sarruf, D.A.; Izgur, V.; Maravilla, K.R.; et al. Obesity is associated with hypothalamic injury in rodents and humans. J. Clin. Investig. 2012, 122, 153–162. [Google Scholar] [CrossRef]
- Li, J.; Tang, Y.; Purkayastha, S.; Yan, J.; Cai, D. Control of obesity and glucose intolerance via building neural stem cells in the hypothalamus. Mol. Metab. 2014, 3, 313–324. [Google Scholar] [CrossRef]
- Chattopadhyaya, B.; Di Cristo, G.; Wu, C.Z.; Knott, G.; Kuhlman, S.; Fu, Y.; Palmiter, R.D.; Huang, Z.J. GAD67-Mediated GABA Synthesis and Signaling Regulate Inhibitory Synaptic Innervation in the Visual Cortex. Neuron 2007, 54, 889–903. [Google Scholar] [CrossRef]
- Kong, D.; Tong, Q.; Ye, C.; Koda, S.; Fuller, P.M.; Krashes, M.J.; Vong, L.; Ray, R.S.; Olson, D.; Lowell, B.B. GABAergic RIP-Cre Neurons in the Arcuate Nucleus Selectively Regulate Energy Expenditure. Cell 2012, 151, 645–657. [Google Scholar] [CrossRef]
- Morrison, S.J.; E Perez, S.; Qiao, Z.; Verdi, J.M.; Hicks, C.; Weinmaster, G.; Anderson, D.J. Transient Notch Activation Initiates an Irreversible Switch from Neurogenesis to Gliogenesis by Neural Crest Stem Cells. Cell 2000, 101, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Kostin, A.; Alam, A.; McGinty, D.; Szymusiak, R.; Alam, N. Chronic Suppression of Hypothalamic Cell Proliferation and Neurogenesis Induces Aging-Like Changes in Sleep–Wake Organization in Young Mice. Neuroscience 2019, 404, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, J.; Purkayastha, S.; Tang, Y.; Zhang, H.; Yin, Y.; Li, B.; Liu, G.; Cai, D. Hypothalamic programming of systemic ageing involving IKK-β, NF-κB and GnRH. Nature 2013, 497, 211–216. [Google Scholar] [CrossRef]
- Zhang, Y.; Kim, M.S.; Jia, B.; Yan, J.; Zuniga-Hertz, J.P.; Han, C.; Cai, D. Hypothalamic stem cells control ageing speed partly through exosomal miRNAs. Nature 2017, 548, 52–57. [Google Scholar] [CrossRef]
- Molofsky, A.V.; He, S.; Bydon, M.; Morrison, S.J.; Pardal, R. Bmi-1 promotes neural stem cell self-renewal and neural development but not mouse growth and survival by repressing the p16Ink4a and p19Arf senescence pathways. Genes Dev. 2005, 19, 1432–1437. [Google Scholar] [CrossRef]
- Lowe, S.W.; Sherr, C.J. Tumor suppression by Ink4a–Arf: Progress and puzzles. Curr. Opin. Genet. Dev. 2003, 13, 77–83. [Google Scholar] [CrossRef]
- Tang, Y.; Zuniga-Hertz, J.P.; Han, C.; Yu, B.; Cai, D. Multifaceted secretion of htNSC-derived hypothalamic islets induces survival and antidiabetic effect via peripheral implantation in mice. eLlife 2020, 9, e52580. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Katakura, M.; Hara, T.; Li, G.; Hashimoto, M.; Shido, O. Proliferation of neuronal progenitor cells and neuronal differentiation in the hypothalamus are enhanced in heat-acclimated rats. Pflug. Arch. Eur. J. Physiol. 2009, 458, 661–673. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Katakura, M.; Sugimoto, N.; Hara, T.; Hashimoto, M.; Shido, O. Neural progenitor cell proliferation in the hypothalamus is involved in acquired heat tolerance in long-term heat-acclimated rats. PLoS ONE 2017, 12, e0178787. [Google Scholar] [CrossRef] [PubMed]
- Bullitt, E. Expression ofC-fos-like protein as a marker for neuronal activity following noxious stimulation in the rat. J. Comp. Neurol. 1990, 296, 517–530. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yahiro, T.; Fukushima, A.; Kataoka, N.; Hioki, H.; Nakamura, K. Prostaglandin EP3 receptor–expressing preoptic neurons bidirectionally control body temperature via tonic GABAergic signaling. Sci. Adv. 2022, 8, eadd5463. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, M. From molecular and cellular to integrative heat defense during exposure to chronic heat. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 131, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Mukuda, T.; Hamasaki, S.; Nakane, H.; Kaidoh, T. Short-term Heat Exposure Promotes Hippocampal Neurogenesis via Activation of Angiotensin II Type 1 Receptor in Adult Rats. Neuroscience 2018, 385, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Moon, M.; Kim, H.G.; Lee, T.H.; Oh, M.S. Heat stress-induced memory impairment is associated with neuroinflammation in mice. J. Neuroinflammation 2015, 12, 102. [Google Scholar] [CrossRef] [PubMed]
- Yerkes, R.M.; Dodson, J.D. The relation of strength of stimulus to rapidity of habit-formation. J. Comp. Neurol. Psychol. 1908, 18, 459–482. [Google Scholar] [CrossRef]
- Weselek, G.; Keiner, S.; Fauser, M.; Wagenführ, L.; Müller, J.; Kaltschmidt, B.; Brandt, M.D.; Gerlach, M.; Redecker, C.; Hermann, A.; et al. Norepinephrine is a negative regulator of the adult periventricular neural stem cell niche. Stem Cells 2020, 38, 1188–1201. [Google Scholar] [CrossRef]
- Loughlin, S.; Foote, S.; Grzanna, R. Efferent projections of nucleus locus coeruleus: Morphologic subpopulations have different efferent targets. Neuroscience 1986, 18, 307–319. [Google Scholar] [CrossRef]
- Loy, R.; Koziell, D.A.; Lindsey, J.D.; Moore, R.Y. Noradrenergic innervation of the adult rat hippocampal formation. J. Comp. Neurol. 1980, 189, 699–710. [Google Scholar] [CrossRef]
- Jhaveri, D.J.; Mackay, E.W.; Hamlin, A.S.; Marathe, S.V.; Nandam, L.S.; Vaidya, V.A.; Bartlett, P.F. Norepinephrine Directly Activates Adult Hippocampal Precursors via beta3-Adrenergic Receptors. J. Neurosci. 2010, 30, 2795–2806. [Google Scholar] [CrossRef]
- Masuda, T.; Nakagawa, S.; Boku, S.; Nishikawa, H.; Takamura, N.; Kato, A.; Inoue, T.; Koyama, T. Noradrenaline increases neural precursor cells derived from adult rat dentate gyrus through beta2 receptor. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 36, 44–51. [Google Scholar] [CrossRef]
- Yoo, S.; Kim, J.; Lyu, P.; Hoang, T.V.; Ma, A.; Trinh, V.; Dai, W.; Jiang, L.; Leavey, P.; Duncan, L.; et al. Control of neurogenic competence in mammalian hypothalamic tanycytes. Sci. Adv. 2021, 7, eabg3777. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Hong, Y.; Zhao, T.; Song, H.; Ming, G.-L. What Makes Organoids Good Models of Human Neurogenesis? Front. Neurosci. 2022, 16, 872794. [Google Scholar] [CrossRef]
- Ghatei, M.A.; Takahashi, K.; Suzuki, Y.; Gardiner, J.; Jones, P.M.; Bloom, S.R. Distribution, molecular characterization of pituitary adenylate cyclase-activating polypeptide and its precursor encoding messenger RNA in human and rat tissues. J. Endocrinol. 1993, 136, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Arimura, A.; Somogyvári-Vigh, A.; Miyata, A.; Mizuno, K.; Coy, D.H.; Kitada, C. Tissue Distribution of PACAP as Determined by RIA: Highly Abundant in the Rat Brain and Testes. Endocrinology 1991, 129, 2787–2789. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, T.; Nakamachi, T.; Shioda, S. Discovery of PACAP and its receptors in the brain. J. Headache Pain 2018, 19, 2787–2789. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.C.; Hashimoto, H.; Galas, L.; et al. Pituitary Adenylate Cyclase-Activating Polypeptide and Its Receptors: 20 Years after the Discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Suyama, S.; Yada, T. New insight into GABAergic neurons in the hypothalamic feeding regulation. J. Physiol. Sci. 2018, 68, 717–722. [Google Scholar] [CrossRef]
- A Romanov, R.; Zeisel, A.; Bakker, J.; Girach, F.; Hellysaz, A.; Tomer, R.; Alpár, A.; Mulder, J.; Clotman, F.; Keimpema, E.; et al. Molecular interrogation of hypothalamic organization reveals distinct dopamine neuronal subtypes. Nat. Neurosci. 2016, 20, 176–188. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makrygianni, E.A.; Chrousos, G.P. Neural Progenitor Cells and the Hypothalamus. Cells 2023, 12, 1822. https://doi.org/10.3390/cells12141822
Makrygianni EA, Chrousos GP. Neural Progenitor Cells and the Hypothalamus. Cells. 2023; 12(14):1822. https://doi.org/10.3390/cells12141822
Chicago/Turabian StyleMakrygianni, Evanthia A., and George P. Chrousos. 2023. "Neural Progenitor Cells and the Hypothalamus" Cells 12, no. 14: 1822. https://doi.org/10.3390/cells12141822
APA StyleMakrygianni, E. A., & Chrousos, G. P. (2023). Neural Progenitor Cells and the Hypothalamus. Cells, 12(14), 1822. https://doi.org/10.3390/cells12141822