

Effects of Oral Cannabinoids on Systemic Inflammation and Viral Reservoir Markers in People with HIV on Antiretroviral Therapy: Results of the CTN PT028 Pilot Clinical Trial

 ,
,  ,
,  ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Design
2.2. Blood and Semen Specimens Processing
2.3. Measurements of Soluble Markers and Cytokines in Plasma
2.4. Ex Vivo Immunophenotyping of T-Cells, Monocytes, and Dendritic Cells
2.5. HIV DNA and Cell-Associated HIV RNA Quantification
2.6. Statistical Analyses
3. Results
3.1. Study Participants
3.2. Effect of Oral Cannabinoids on Plasma Markers of Gut Epithelial Damage, Microbial Translocation, and Systemic Inflammation
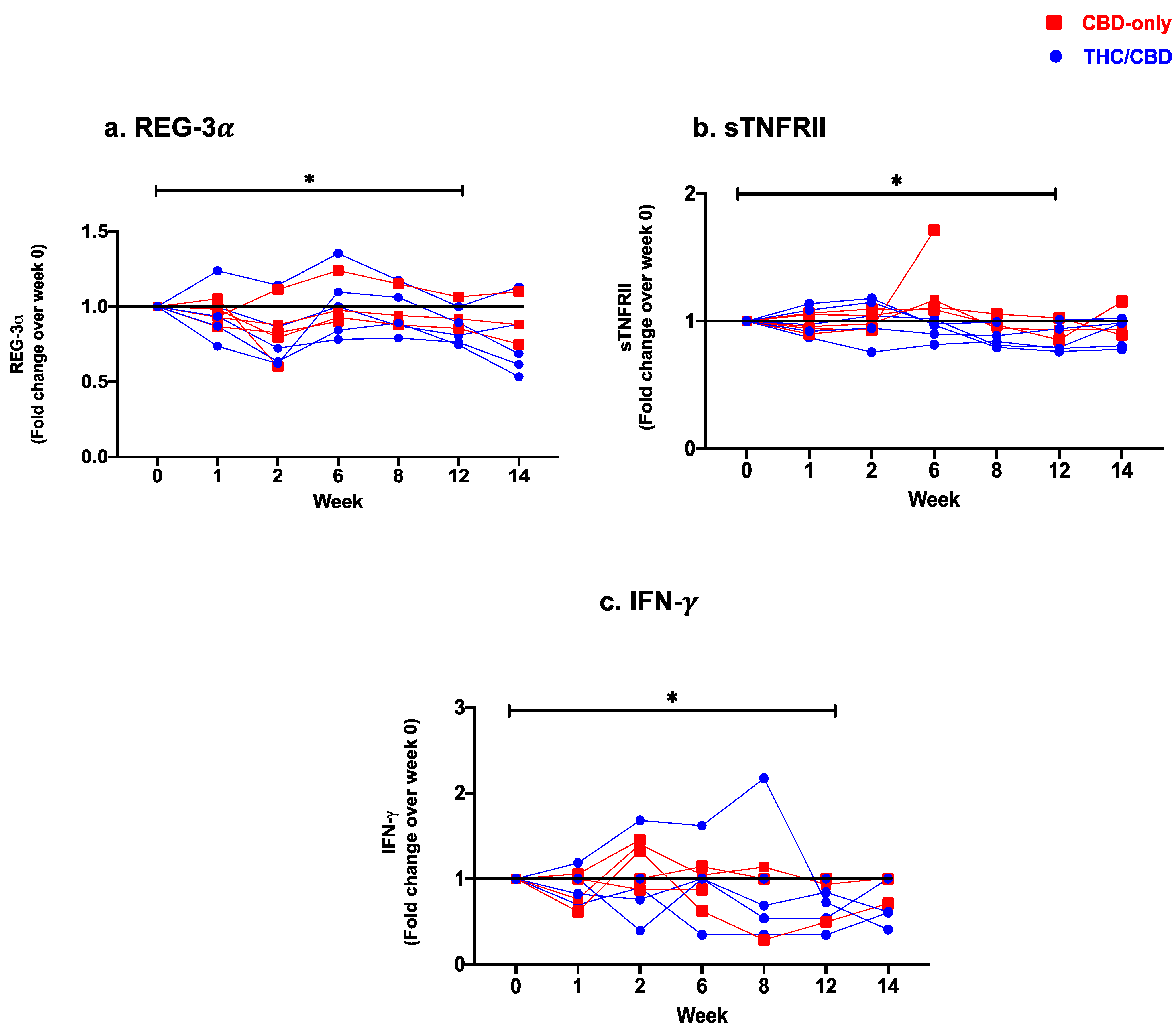
Reduced Levels of Soluble Markers of Gut Epithelial Damage, Microbial Translocation, Immune Activation, and Pro-Inflammatory Cytokines
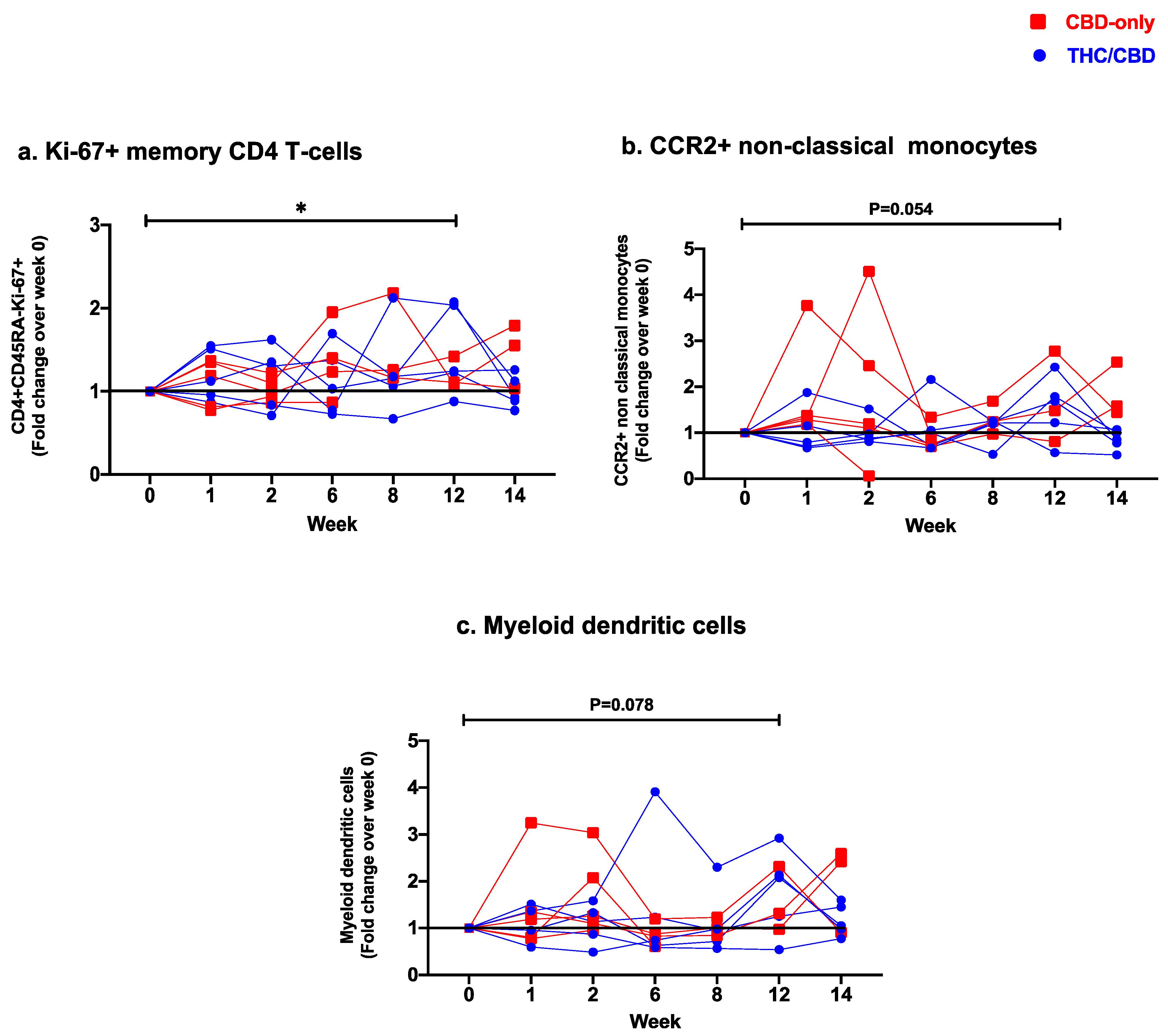
3.3. Effect of Oral Cannabinoids on Blood T-Cell, Monocyte, and Dendritic Cell Subsets
3.3.1. Changes in Circulating CD4 T-Cell Subsets
3.3.2. Changes in Circulating CD8 T-Cell Subsets
3.3.3. Changes in Monocyte Subsets and Dendritic Cell Frequencies
3.4. Effect of Oral Cannabinoids on Total HIV DNA and Cell-Associated HIV RNA in CD4 T-Cells from Blood and Semen
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deeks, S.G. Immune dysfunction, inflammation, and accelerated aging in patients on antiretroviral therapy. Top. HIV Med. 2009, 17, 118–123. [Google Scholar] [PubMed]
- Deeks, S.G. HIV infection, inflammation, immunosenescence, and aging. Annu. Rev. Med. 2011, 62, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Zicari, S.; Sessa, L.; Cotugno, N.; Ruggiero, A.; Morrocchi, E.; Concato, C.; Rocca, S.; Zangari, P.; Manno, E.C.; Palma, P. Immune Activation, Inflammation, and Non-AIDS Co-Morbidities in HIV-Infected Patients under Long-Term ART. Viruses 2019, 11, 200. [Google Scholar] [CrossRef]
- Yero, A.; Bouassa, R.M.; Ancuta, P.; Estaquier, J.; Jenabian, M.A. Immuno-metabolic control of the balance between Th17-polarized and regulatory T-cells during HIV infection. Cytokine Growth Factor. Rev. 2023, 69, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G.; Tracy, R.; Douek, D.C. Systemic effects of inflammation on health during chronic HIV infection. Immunity 2013, 39, 633–645. [Google Scholar] [CrossRef]
- Hunt, P.W.; Sinclair, E.; Rodriguez, B.; Shive, C.; Clagett, B.; Funderburg, N.; Robinson, J.; Huang, Y.; Epling, L.; Martin, J.N.; et al. Gut epithelial barrier dysfunction and innate immune activation predict mortality in treated HIV infection. J. Infect. Dis. 2014, 210, 1228–1238. [Google Scholar] [CrossRef]
- Jenabian, M.A.; El-Far, M.; Vyboh, K.; Kema, I.; Costiniuk, C.T.; Thomas, R.; Baril, J.G.; LeBlanc, R.; Kanagaratham, C.; Radzioch, D.; et al. Immunosuppressive Tryptophan Catabolism and Gut Mucosal Dysfunction Following Early HIV Infection. J. Infect. Dis. 2015, 212, 355–366. [Google Scholar] [CrossRef]
- Yero, A.; Farnos, O.; Rabezanahary, H.; Racine, G.; Estaquier, J.; Jenabian, M.A. Differential Dynamics of Regulatory T-Cell and Th17 Cell Balance in Mesenteric Lymph Nodes and Blood following Early Antiretroviral Initiation during Acute Simian Immunodeficiency Virus Infection. J. Virol. 2019, 93, e00371-19. [Google Scholar] [CrossRef]
- Sokoya, T.; Steel, H.C.; Nieuwoudt, M.; Rossouw, T.M. HIV as a Cause of Immune Activation and Immunosenescence. Mediators Inflamm. 2017, 2017, 6825493. [Google Scholar] [CrossRef]
- Martin, G.E.; Pace, M.; Shearer, F.M.; Zilber, E.; Hurst, J.; Meyerowitz, J.; Thornhill, J.P.; Lwanga, J.; Brown, H.; Robinson, N.; et al. Levels of Human Immunodeficiency Virus DNA Are Determined Before ART Initiation and Linked to CD8 T-Cell Activation and Memory Expansion. J. Infect. Dis. 2020, 221, 1135–1145. [Google Scholar] [CrossRef]
- Khoury, G.; Fromentin, R.; Solomon, A.; Hartogensis, W.; Killian, M.; Hoh, R.; Somsouk, M.; Hunt, P.W.; Girling, V.; Sinclair, E.; et al. Human Immunodeficiency Virus Persistence and T-Cell Activation in Blood, Rectal, and Lymph Node Tissue in Human Immunodeficiency Virus-Infected Individuals Receiving Suppressive Antiretroviral Therapy. J. Infect. Dis. 2017, 215, 911–919. [Google Scholar] [CrossRef]
- Anil, S.M.; Shalev, N.; Vinayaka, A.C.; Nadarajan, S.; Namdar, D.; Belausov, E.; Shoval, I.; Mani, K.A.; Mechrez, G.; Koltai, H. Cannabis compounds exhibit anti-inflammatory activity in vitro in COVID-19-related inflammation in lung epithelial cells and pro-inflammatory activity in macrophages. Sci. Rep. 2021, 11, 1462. [Google Scholar] [CrossRef]
- DeMarino, C.; Cowen, M.; Khatkar, P.; Cotto, B.; Branscome, H.; Kim, Y.; Sharif, S.A.; Agbottah, E.T.; Zhou, W.; Costiniuk, C.T.; et al. Cannabinoids Reduce Extracellular Vesicle Release from HIV-1 Infected Myeloid Cells and Inhibit Viral Transcription. Cells 2022, 11, 723. [Google Scholar] [CrossRef] [PubMed]
- Henriquez, J.E.; Rizzo, M.D.; Schulz, M.A.; Crawford, R.B.; Gulick, P.; Kaminski, N.E. Δ9-Tetrahydrocannabinol Suppresses Secretion of IFNα by Plasmacytoid Dendritic Cells From Healthy and HIV-Infected Individuals. J. Acquir. Immune Defic. Syndr. 2017, 75, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Suryavanshi, S.V.; Zaiachuk, M.; Pryimak, N.; Kovalchuk, I.; Kovalchuk, O. Cannabinoids Alleviate the LPS-Induced Cytokine Storm via Attenuating NLRP3 Inflammasome Signaling and TYK2-Mediated STAT3 Signaling Pathways In Vitro. Cells 2022, 11, 1391. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.D.; Crawford, R.B.; Bach, A.; Sermet, S.; Amalfitano, A.; Kaminski, N.E. Δ(9)-Tetrahydrocannabinol Suppresses Monocyte-Mediated Astrocyte Production of Monocyte Chemoattractant Protein 1 and Interleukin-6 in a Toll-Like Receptor 7-Stimulated Human Coculture. J. Pharmacol. Exp. Ther. 2019, 371, 191–201. [Google Scholar] [CrossRef]
- Carmona-Hidalgo, B.; González-Mariscal, I.; García-Martín, A.; Prados, M.E.; Ruiz-Pino, F.; Appendino, G.; Tena-Sempere, M.; Muñoz, E. Δ9-Tetrahydrocannabinolic Acid markedly alleviates liver fibrosis and inflammation in mice. Phytomedicine 2021, 81, 153426. [Google Scholar] [CrossRef]
- Chandra, L.C.; Kumar, V.; Torben, W.; Vande Stouwe, C.; Winsauer, P.; Amedee, A.; Molina, P.E.; Mohan, M. Chronic administration of Δ9-tetrahydrocannabinol induces intestinal anti-inflammatory microRNA expression during acute simian immunodeficiency virus infection of rhesus macaques. J. Virol. 2015, 89, 1168–1181. [Google Scholar] [CrossRef]
- Kumar, V.; Torben, W.; Kenway, C.S.; Schiro, F.R.; Mohan, M. Longitudinal Examination of the Intestinal Lamina Propria Cellular Compartment of Simian Immunodeficiency Virus-Infected Rhesus Macaques Provides Broader and Deeper Insights into the Link between Aberrant MicroRNA Expression and Persistent Immune Activation. J. Virol. 2016, 90, 5003–5019. [Google Scholar] [CrossRef]
- Kumar, V.; Torben, W.; Mansfield, J.; Alvarez, X.; Vande Stouwe, C.; Li, J.; Byrareddy, S.N.; Didier, P.J.; Pahar, B.; Molina, P.E.; et al. Cannabinoid Attenuation of Intestinal Inflammation in Chronic SIV-Infected Rhesus Macaques Involves T Cell Modulation and Differential Expression of Micro-RNAs and Pro-inflammatory Genes. Front. Immunol. 2019, 10, 914. [Google Scholar] [CrossRef]
- McDew-White, M.; Lee, E.; Alvarez, X.; Sestak, K.; Ling, B.J.; Byrareddy, S.N.; Okeoma, C.M.; Mohan, M. Cannabinoid control of gingival immune activation in chronically SIV-infected rhesus macaques involves modulation of the indoleamine-2,3-dioxygenase-1 pathway and salivary microbiome. EBioMedicine 2022, 75, 103769. [Google Scholar] [CrossRef] [PubMed]
- Couch, D.G.; Cook, H.; Ortori, C.; Barrett, D.; Lund, J.N.; O’Sullivan, S.E. Palmitoylethanolamide and Cannabidiol Prevent Inflammation-induced Hyperpermeability of the Human Gut In Vitro and In Vivo-A Randomized, Placebo-controlled, Double-blind Controlled Trial. Inflamm. Bowel Dis. 2019, 25, 1006–1018. [Google Scholar] [CrossRef] [PubMed]
- Cocetta, V.; Governa, P.; Borgonetti, V.; Tinazzi, M.; Peron, G.; Catanzaro, D.; Berretta, M.; Biagi, M.; Manetti, F.; Dall’Acqua, S.; et al. Cannabidiol Isolated From Cannabis sativa L. Protects Intestinal Barrier From In Vitro Inflammation and Oxidative Stress. Front. Pharmacol. 2021, 12, 641210. [Google Scholar] [CrossRef] [PubMed]
- Kozela, E.; Juknat, A.; Kaushansky, N.; Rimmerman, N.; Ben-Nun, A.; Vogel, Z. Cannabinoids decrease the th17 inflammatory autoimmune phenotype. J. Neuroimmune Pharmacol. 2013, 8, 1265–1276. [Google Scholar] [CrossRef]
- Berg, B.B.; Soares, J.S.; Paiva, I.R.; Rezende, B.M.; Rachid, M.A.; Cau, S.B.A.; Romero, T.R.L.; Pinho, V.; Teixeira, M.M.; Castor, M. Cannabidiol Enhances Intestinal Cannabinoid Receptor Type 2 Receptor Expression and Activation Increasing Regulatory T Cells and Reduces Murine Acute Graft-versus-Host Disease without Interfering with the Graft-versus-Leukemia Response. J. Pharmacol. Exp. Ther. 2021, 377, 273–283. [Google Scholar] [CrossRef]
- McDew-White, M.; Lee, E.; Premadasa, L.S.; Alvarez, X.; Okeoma, C.M.; Mohan, M. Cannabinoids modulate the microbiota-gut-brain axis in HIV/SIV infection by reducing neuroinflammation and dysbiosis while concurrently elevating endocannabinoid and indole-3-propionate levels. J. Neuroinflammation 2023, 20, 62. [Google Scholar] [CrossRef]
- Kaddour, H.; McDew-White, M.; Madeira, M.M.; Tranquille, M.A.; Tsirka, S.E.; Mohan, M.; Okeoma, C.M. Chronic delta-9-tetrahydrocannabinol (THC) treatment counteracts SIV-induced modulation of proinflammatory microRNA cargo in basal ganglia-derived extracellular vesicles. J. Neuroinflammation 2022, 19, 225. [Google Scholar] [CrossRef]
- Kopcho, S.; McDew-White, M.; Naushad, W.; Mohan, M.; Okeoma, C.M. Alterations in Abundance and Compartmentalization of miRNAs in Blood Plasma Extracellular Vesicles and Extracellular Condensates during HIV/SIV Infection and Its Modulation by Antiretroviral Therapy (ART) and Delta-9-Tetrahydrocannabinol (Delta(9)-THC). Viruses 2023, 15, 623. [Google Scholar] [CrossRef]
- Pak, K.; Kantonen, T.; Pekkarinen, L.; Nuutila, P.; Nummenmaa, L. Association of CNR1 gene and cannabinoid 1 receptor protein in the human brain. J. Neurosci. Res. 2023, 101, 327–337. [Google Scholar] [CrossRef]
- Grabon, W.; Rheims, S.; Smith, J.; Bodennec, J.; Belmeguenai, A.; Bezin, L. CB2 receptor in the CNS: From immune and neuronal modulation to behavior. Neurosci. Biobehav. Rev. 2023, 150, 105226. [Google Scholar] [CrossRef]
- Stanzani, A.; Galiazzo, G.; Giancola, F.; Tagliavia, C.; De Silva, M.; Pietra, M.; Fracassi, F.; Chiocchetti, R. Localization of cannabinoid and cannabinoid related receptors in the cat gastrointestinal tract. Histochem. Cell Biol. 2020, 153, 339–356. [Google Scholar] [CrossRef] [PubMed]
- Mboumba Bouassa, R.S.; Sebastiani, G.; Di Marzo, V.; Jenabian, M.A.; Costiniuk, C.T. Cannabinoids and Chronic Liver Diseases. Int. J. Mol. Sci. 2022, 23, 9423. [Google Scholar] [CrossRef]
- Rizzo, M.D.; Henriquez, J.E.; Blevins, L.K.; Bach, A.; Crawford, R.B.; Kaminski, N.E. Targeting Cannabinoid Receptor 2 on Peripheral Leukocytes to Attenuate Inflammatory Mechanisms Implicated in HIV-Associated Neurocognitive Disorder. J. Neuroimmune Pharmacol. 2020, 15, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Costiniuk, C.T.; Jenabian, M.A. Cannabinoids and inflammation: Implications for people living with HIV. Aids 2019, 33, 2273–2288. [Google Scholar] [CrossRef]
- Hsu, D.C.; Sereti, I. Serious Non-AIDS Events: Therapeutic Targets of Immune Activation and Chronic Inflammation in HIV Infection. Drugs 2016, 76, 533–549. [Google Scholar] [CrossRef] [PubMed]
- Zgair, A.; Lee, J.B.; Wong, J.C.M.; Taha, D.A.; Aram, J.; Di Virgilio, D.; McArthur, J.W.; Cheng, Y.K.; Hennig, I.M.; Barrett, D.A.; et al. Oral administration of cannabis with lipids leads to high levels of cannabinoids in the intestinal lymphatic system and prominent immunomodulation. Sci. Rep. 2017, 7, 14542. [Google Scholar] [CrossRef] [PubMed]
- Canada, H. Understanding the New Access to Cannabis for Medical Purposes Regulations. 2016. Available online: https://www.canada.ca/en/health-canada/services/publications/drugs-health-products/understanding-new-access-to-cannabis-for-medical-purposes-regulations.html (accessed on 7 July 2023).
- Furler, M.D.; Einarson, T.R.; Millson, M.; Walmsley, S.; Bendayan, R. Medicinal and recreational marijuana use by patients infected with HIV. AIDS Patient Care STDS 2004, 18, 215–228. [Google Scholar] [CrossRef]
- Harris, G.E.; Dupuis, L.; Mugford, G.J.; Johnston, L.; Haase, D.; Page, G.; Haldane, H.; Harris, N.; Midodzi, W.K.; Dow, G. Patterns and correlates of cannabis use among individuals with HIV/AIDS in Maritime Canada. Can. J. Infect. Dis. Med. Microbiol. 2014, 25, e1–e7. [Google Scholar] [CrossRef]
- Hawke, L.D.; Henderson, J. Legalization of cannabis use in Canada: Impacts on the cannabis use profiles of youth seeking services for substance use. J. Subst. Abuse Treat. 2021, 126, 108340. [Google Scholar] [CrossRef]
- Ellis, R.J.; Peterson, S.N.; Li, Y.; Schrier, R.; Iudicello, J.; Letendre, S.; Morgan, E.; Tang, B.; Grant, I.; Cherner, M. Recent cannabis use in HIV is associated with reduced inflammatory markers in CSF and blood. Neurol. Neuroimmunol. Neuroinflamm 2020, 7. [Google Scholar] [CrossRef]
- Manuzak, J.A.; Gott, T.M.; Kirkwood, J.S.; Coronado, E.; Hensley-McBain, T.; Miller, C.; Cheu, R.K.; Collier, A.C.; Funderburg, N.T.; Martin, J.N.; et al. Heavy Cannabis Use Associated With Reduction in Activated and Inflammatory Immune Cell Frequencies in Antiretroviral Therapy-Treated Human Immunodeficiency Virus-Infected Individuals. Clin. Infect. Dis. 2018, 66, 1872–1882. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.D.; Crawford, R.B.; Henriquez, J.E.; Aldhamen, Y.A.; Gulick, P.; Amalfitano, A.; Kaminski, N.E. HIV-infected cannabis users have lower circulating CD16+ monocytes and IFN-γ-inducible protein 10 levels compared with nonusing HIV patients. Aids 2018, 32, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.W.; Campbell, L.M.; Sun-Suslow, N.; Hong, S.; Umlauf, A.; Ellis, R.J.; Iudicello, J.E.; Letendre, S.; Marcotte, T.D.; Heaton, R.K.; et al. Daily Cannabis Use is Associated With Lower CNS Inflammation in People With HIV. J. Int. Neuropsychol. Soc. 2021, 27, 661–672. [Google Scholar] [CrossRef]
- Yin, L.; Dinasarapu, A.R.; Borkar, S.A.; Chang, K.F.; De Paris, K.; Kim-Chang, J.J.; Sleasman, J.W.; Goodenow, M.M. Anti-inflammatory effects of recreational marijuana in virally suppressed youth with HIV-1 are reversed by use of tobacco products in combination with marijuana. Retrovirology 2022, 19, 10. [Google Scholar] [CrossRef]
- Turner, S.E.; Williams, C.M.; Iversen, L.; Whalley, B.J. Molecular Pharmacology of Phytocannabinoids. Prog. Chem. Org. Nat. Prod. 2017, 103, 61–101. [Google Scholar] [CrossRef]
- Manuzak, J.A.; Granche, J.; Tassiopoulos, K.; Rower, J.E.; Knox, J.R.; Williams, D.W.; Ellis, R.J.; Goodkin, K.; Sharma, A.; Erlandson, K.M. Cannabis Use Is Associated With Decreased Antiretroviral Therapy Adherence Among Older Adults With HIV. Open Forum Infect. Dis. 2023, 10, ofac699. [Google Scholar] [CrossRef] [PubMed]
- Bahji, A.; Li, Y.; Vickers-Smith, R.; Crystal, S.; Kerns, R.D.; Gordon, K.S.; Macmadu, A.; Skanderson, M.; So-Armah, K.; Sung, M.L.; et al. Self-Reported Cannabis Use and HIV Viral Control among Patients with HIV Engaged in Care: Results from a National Cohort Study. Int. J. Environ. Res. Public Health 2022, 19, 5649. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, B.; Miller, M.L.; Hurd, Y.L. Cannabis Use during Adolescent Development: Susceptibility to Psychiatric Illness. Front. Psychiatry 2013, 4, 129. [Google Scholar] [CrossRef]
- Mboumba Bouassa, R.S.; Needham, J.; Nohynek, D.; Singer, J.; Lee, T.; Bobeuf, F.; Samarani, S.; Del Balso, L.; Paisible, N.; Vertzagias, C.; et al. Safety and Tolerability of Oral Cannabinoids in People Living with HIV on Long-Term ART: A Randomized, Open-Label, Interventional Pilot Clinical Trial (CTNPT 028). Biomedicines 2022, 10, 3168. [Google Scholar] [CrossRef]
- Bergamaschi, M.M.; Queiroz, R.H.; Zuardi, A.W.; Crippa, J.A. Safety and side effects of cannabidiol, a Cannabis sativa constituent. Curr. Drug Saf. 2011, 6, 237–249. [Google Scholar] [CrossRef]
- Burstein, S. Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorganic Med. Chem. 2015, 23, 1377–1385. [Google Scholar] [CrossRef]
- Burstein, S.H.; Zurier, R.B. Cannabinoids, endocannabinoids, and related analogs in inflammation. AAPS J. 2009, 11, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Iuvone, T.; Esposito, G.; De Filippis, D.; Scuderi, C.; Steardo, L. Cannabidiol: A promising drug for neurodegenerative disorders? CNS Neurosci. Ther. 2009, 15, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Leweke, F.M.; Rohleder, C.; Gerth, C.W.; Hellmich, M.; Pukrop, R.; Koethe, D. Cannabidiol and Amisulpride Improve Cognition in Acute Schizophrenia in an Explorative, Double-Blind, Active-Controlled, Randomized Clinical Trial. Front. Pharmacol. 2021, 12, 614811. [Google Scholar] [CrossRef]
- Leweke, F.M.; Piomelli, D.; Pahlisch, F.; Muhl, D.; Gerth, C.W.; Hoyer, C.; Klosterkotter, J.; Hellmich, M.; Koethe, D. Cannabidiol enhances anandamide signaling and alleviates psychotic symptoms of schizophrenia. Transl. Psychiatry 2012, 2, e94. [Google Scholar] [CrossRef] [PubMed]
- Thiele, E.A.; Marsh, E.D.; French, J.A.; Mazurkiewicz-Beldzinska, M.; Benbadis, S.R.; Joshi, C.; Lyons, P.D.; Taylor, A.; Roberts, C.; Sommerville, K. Cannabidiol in patients with seizures associated with Lennox-Gastaut syndrome (GWPCARE4): A randomised, double-blind, placebo-controlled phase 3 trial. Lancet 2018, 391, 1085–1096. [Google Scholar] [CrossRef]
- Wade, D.T.; Makela, P.; Robson, P.; House, H.; Bateman, C. Do cannabis-based medicinal extracts have general or specific effects on symptoms in multiple sclerosis? A double-blind, randomized, placebo-controlled study on 160 patients. Mult. Scler. 2004, 10, 434–441. [Google Scholar] [CrossRef]
- Costiniuk, C.T.; Saneei, Z.; Routy, J.P.; Margolese, S.; Mandarino, E.; Singer, J.; Lebouché, B.; Cox, J.; Szabo, J.; Brouillette, M.J.; et al. Oral cannabinoids in people living with HIV on effective antiretroviral therapy: CTN PT028-study protocol for a pilot randomised trial to assess safety, tolerability and effect on immune activation. BMJ Open 2019, 9, e024793. [Google Scholar] [CrossRef]
- Costiniuk, C.T.; Salahuddin, S.; Farnos, O.; Olivenstein, R.; Pagliuzza, A.; Orlova, M.; Schurr, E.; De Castro, C.; Bourbeau, J.; Routy, J.P.; et al. HIV persistence in mucosal CD4+ T cells within the lungs of adults receiving long-term suppressive antiretroviral therapy. Aids 2018, 32, 2279–2289. [Google Scholar] [CrossRef]
- Vandergeeten, C.; Fromentin, R.; Merlini, E.; Lawani, M.B.; DaFonseca, S.; Bakeman, W.; McNulty, A.; Ramgopal, M.; Michael, N.; Kim, J.H.; et al. Cross-clade ultrasensitive PCR-based assays to measure HIV persistence in large-cohort studies. J. Virol. 2014, 88, 12385–12396. [Google Scholar] [CrossRef]
- Isnard, S.; Ramendra, R.; Dupuy, F.P.; Lin, J.; Fombuena, B.; Kokinov, N.; Kema, I.; Jenabian, M.A.; Lebouché, B.; Costiniuk, C.T.; et al. Plasma Levels of C-Type Lectin REG3α and Gut Damage in People With Human Immunodeficiency Virus. J. Infect. Dis. 2020, 221, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Novelli, S.; Lecuroux, C.; Goujard, C.; Reynes, J.; Villemant, A.; Blum, L.; Essat, A.; Avettand-Fenoel, V.; Launay, O.; Molina, J.M.; et al. Persistence of monocyte activation under treatment in people followed since acute HIV-1 infection relative to participants at high or low risk of HIV infection. EBioMedicine 2020, 62, 103129. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Jochems, C.; Anderson, A.M.; Talaie, T.; Jales, A.; Madan, R.A.; Hodge, J.W.; Tsang, K.Y.; Liewehr, D.J.; Steinberg, S.M.; et al. Soluble CD27-pool in humans may contribute to T cell activation and tumor immunity. J. Immunol. 2013, 190, 6250–6258. [Google Scholar] [CrossRef] [PubMed]
- Marafini, I.; Di Sabatino, A.; Zorzi, F.; Monteleone, I.; Sedda, S.; Cupi, M.L.; Antenucci, C.; Biancheri, P.; Giuffrida, P.; Di Stefano, M.; et al. Serum regenerating islet-derived 3-alpha is a biomarker of mucosal enteropathies. Aliment. Pharmacol. Ther. 2014, 40, 974–981. [Google Scholar] [CrossRef]
- Vaishnava, S.; Yamamoto, M.; Severson, K.M.; Ruhn, K.A.; Yu, X.; Koren, O.; Ley, R.; Wakeland, E.K.; Hooper, L.V. The antibacterial lectin RegIIIgamma promotes the spatial segregation of microbiota and host in the intestine. Science 2011, 334, 255–258. [Google Scholar] [CrossRef]
- Cuddihey, H.; MacNaughton, W.K.; Sharkey, K.A. Role of the Endocannabinoid System in the Regulation of Intestinal Homeostasis. Cell Mol. Gastroenterol. Hepatol. 2022, 14, 947–963. [Google Scholar] [CrossRef]
- Alhamoruni, A.; Lee, A.C.; Wright, K.L.; Larvin, M.; O’Sullivan, S.E. Pharmacological effects of cannabinoids on the Caco-2 cell culture model of intestinal permeability. J. Pharmacol. Exp. Ther. 2010, 335, 92–102. [Google Scholar] [CrossRef]
- Alhamoruni, A.; Wright, K.L.; Larvin, M.; O’Sullivan, S.E. Cannabinoids mediate opposing effects on inflammation-induced intestinal permeability. Br. J. Pharmacol. 2012, 165, 2598–2610. [Google Scholar] [CrossRef]
- Taschler, U.; Hasenoehrl, C.; Storr, M.; Schicho, R. Cannabinoid Receptors in Regulating the GI Tract: Experimental Evidence and Therapeutic Relevance. Handb. Exp. Pharmacol. 2017, 239, 343–362. [Google Scholar] [CrossRef]
- Wiley, M.B.; DiPatrizio, N.V. Diet-Induced Gut Barrier Dysfunction Is Exacerbated in Mice Lacking Cannabinoid 1 Receptors in the Intestinal Epithelium. Int. J. Mol. Sci. 2022, 23, 10549. [Google Scholar] [CrossRef]
- Oláh, A.; Szekanecz, Z.; Bíró, T. Targeting Cannabinoid Signaling in the Immune System: “High”-ly Exciting Questions, Possibilities, and Challenges. Front. Immunol. 2017, 8, 1487. [Google Scholar] [CrossRef] [PubMed]
- Pellati, F.; Borgonetti, V.; Brighenti, V.; Biagi, M.; Benvenuti, S.; Corsi, L. Cannabis sativa L. and Nonpsychoactive Cannabinoids: Their Chemistry and Role against Oxidative Stress, Inflammation, and Cancer. Biomed. Res. Int. 2018, 2018, 1691428. [Google Scholar] [CrossRef] [PubMed]
- Mehrpouya-Bahrami, P.; Chitrala, K.N.; Ganewatta, M.S.; Tang, C.; Murphy, E.A.; Enos, R.T.; Velazquez, K.T.; McCellan, J.; Nagarkatti, M.; Nagarkatti, P. Blockade of CB1 cannabinoid receptor alters gut microbiota and attenuates inflammation and diet-induced obesity. Sci. Rep. 2017, 7, 15645. [Google Scholar] [CrossRef] [PubMed]
- Skinner, C.M.; Nookaew, I.; Ewing, L.E.; Wongsurawat, T.; Jenjaroenpun, P.; Quick, C.M.; Yee, E.U.; Piccolo, B.D.; ElSohly, M.; Walker, L.A.; et al. Potential Probiotic or Trigger of Gut Inflammation—The Janus-Faced Nature of Cannabidiol-Rich Cannabis Extract. J. Diet. Suppl. 2020, 17, 543–560. [Google Scholar] [CrossRef]
- Grider, J.R.; Mahavadi, S.; Li, Y.; Qiao, L.Y.; Kuemmerle, J.F.; Murthy, K.S.; Martin, B.R. Modulation of motor and sensory pathways of the peristaltic reflex by cannabinoids. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G539–G549. [Google Scholar] [CrossRef]
- Henshaw, F.R.; Dewsbury, L.S.; Lim, C.K.; Steiner, G.Z. The Effects of Cannabinoids on Pro- and Anti-Inflammatory Cytokines: A Systematic Review of In Vivo Studies. Cannabis Cannabinoid Res. 2021, 6, 177–195. [Google Scholar] [CrossRef]
- Christensen-Quick, A.; Massanella, M.; Frick, A.; Rawlings, S.A.; Spina, C.; Vargas-Meneses, M.; Schrier, R.; Nakazawa, M.; Anderson, C.; Gianella, S. Subclinical Cytomegalovirus DNA Is Associated with CD4 T Cell Activation and Impaired CD8 T Cell CD107a Expression in People Living with HIV despite Early Antiretroviral Therapy. J. Virol. 2019, 93, 10-1128. [Google Scholar] [CrossRef]
- Maidji, E.; Somsouk, M.; Rivera, J.M.; Hunt, P.W.; Stoddart, C.A. Replication of CMV in the gut of HIV-infected individuals and epithelial barrier dysfunction. PLoS Pathog. 2017, 13, e1006202. [Google Scholar] [CrossRef]
- Fenwick, C.; Joo, V.; Jacquier, P.; Noto, A.; Banga, R.; Perreau, M.; Pantaleo, G. T-cell exhaustion in HIV infection. Immunol. Rev. 2019, 292, 149–163. [Google Scholar] [CrossRef]
- Lim, A.; French, M.A.; Price, P. CD4+ and CD8+ T cells expressing FoxP3 in HIV-infected patients are phenotypically distinct and influenced by disease severity and antiretroviral therapy. J. Acquir. Immune Defic. Syndr. 2009, 51, 248–257. [Google Scholar] [CrossRef]
- Lim, A.; Tan, D.; Price, P.; Kamarulzaman, A.; Tan, H.Y.; James, I.; French, M.A. Proportions of circulating T cells with a regulatory cell phenotype increase with HIV-associated immune activation and remain high on antiretroviral therapy. Aids 2007, 21, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Nigam, P.; Velu, V.; Kannanganat, S.; Chennareddi, L.; Kwa, S.; Siddiqui, M.; Amara, R.R. Expansion of FOXP3+ CD8 T cells with suppressive potential in colorectal mucosa following a pathogenic simian immunodeficiency virus infection correlates with diminished antiviral T cell response and viral control. J. Immunol. 2010, 184, 1690–1701. [Google Scholar] [CrossRef] [PubMed]
- Yero, A.; Shi, T.; Routy, J.P.; Tremblay, C.; Durand, M.; Costiniuk, C.T.; Jenabian, M.A. FoxP3(+) CD8 T-cells in acute HIV infection and following early antiretroviral therapy initiation. Front. Immunol. 2022, 13, 962912. [Google Scholar] [CrossRef] [PubMed]
- Schulze Zur Wiesch, J.; Thomssen, A.; Hartjen, P.; Tóth, I.; Lehmann, C.; Meyer-Olson, D.; Colberg, K.; Frerk, S.; Babikir, D.; Schmiedel, S.; et al. Comprehensive analysis of frequency and phenotype of T regulatory cells in HIV infection: CD39 expression of FoxP3+ T regulatory cells correlates with progressive disease. J. Virol. 2011, 85, 1287–1297. [Google Scholar] [CrossRef]
- Song, J.W.; Huang, H.H.; Zhang, C.; Yang, H.G.; Zhang, J.Y.; Xu, R.N.; Jin, L.; Shi, M.; Wang, F.S.; Jiao, Y.M. Expression of CD39 Is Correlated With HIV DNA Levels in Naïve Tregs in Chronically Infected ART Naïve Patients. Front. Immunol. 2019, 10, 2465. [Google Scholar] [CrossRef]
- Antonioli, L.; Pacher, P.; Vizi, E.S.; Haskó, G. CD39 and CD73 in immunity and inflammation. Trends Mol. Med. 2013, 19, 355–367. [Google Scholar] [CrossRef]
- Jenabian, M.A.; Seddiki, N.; Yatim, A.; Carriere, M.; Hulin, A.; Younas, M.; Ghadimi, E.; Kok, A.; Routy, J.P.; Tremblay, A.; et al. Regulatory T cells negatively affect IL-2 production of effector T cells through CD39/adenosine pathway in HIV infection. PLoS Pathog. 2013, 9, e1003319. [Google Scholar] [CrossRef]
- Nikolova, M.; Carriere, M.; Jenabian, M.A.; Limou, S.; Younas, M.; Kok, A.; Hue, S.; Seddiki, N.; Hulin, A.; Delaneau, O.; et al. CD39/adenosine pathway is involved in AIDS progression. PLoS Pathog. 2011, 7, e1002110. [Google Scholar] [CrossRef]
- Yero, A.; Shi, T.; Farnos, O.; Routy, J.P.; Tremblay, C.; Durand, M.; Tsoukas, C.; Costiniuk, C.T.; Jenabian, M.A. Dynamics and epigenetic signature of regulatory T-cells following antiretroviral therapy initiation in acute HIV infection. EBioMedicine 2021, 71, 103570. [Google Scholar] [CrossRef]
- Dutertre, C.A.; Amraoui, S.; DeRosa, A.; Jourdain, J.P.; Vimeux, L.; Goguet, M.; Degrelle, S.; Feuillet, V.; Liovat, A.S.; Müller-Trutwin, M.; et al. Pivotal role of M-DC8⁺ monocytes from viremic HIV-infected patients in TNFα overproduction in response to microbial products. Blood 2012, 120, 2259–2268. [Google Scholar] [CrossRef]
- Chayasirisobhon, S. Mechanisms of Action and Pharmacokinetics of Cannabis. Perm. J. 2020, 25, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Ndhlovu, L.C.; D’Antoni, M.L.; Ananworanich, J.; Byron, M.M.; Chalermchai, T.; Sithinamsuwan, P.; Tipsuk, S.; Ho, E.; Slike, B.M.; Schuetz, A.; et al. Loss of CCR2 expressing non-classical monocytes are associated with cognitive impairment in antiretroviral therapy-naïve HIV-infected Thais. J. Neuroimmunol. 2015, 288, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Amodio, G.; Gregori, S. Dendritic cells a double-edge sword in autoimmune responses. Front. Immunol. 2012, 3, 233. [Google Scholar] [CrossRef] [PubMed]
- Amodio, G.; Cichy, J.; Conde, P.; Matteoli, G.; Moreau, A.; Ochando, J.; Oral, B.H.; Pekarova, M.; Ryan, E.J.; Roth, J.; et al. Role of myeloid regulatory cells (MRCs) in maintaining tissue homeostasis and promoting tolerance in autoimmunity, inflammatory disease and transplantation. Cancer Immunol. Immunother. 2019, 68, 661–672. [Google Scholar] [CrossRef]
- Molina, P.E.; Winsauer, P.; Zhang, P.; Walker, E.; Birke, L.; Amedee, A.; Stouwe, C.V.; Troxclair, D.; McGoey, R.; Varner, K.; et al. Cannabinoid administration attenuates the progression of simian immunodeficiency virus. AIDS Res. Hum. Retroviruses 2011, 27, 585–592. [Google Scholar] [CrossRef]
- Milloy, M.J.; Marshall, B.; Kerr, T.; Richardson, L.; Hogg, R.; Guillemi, S.; Montaner, J.S.; Wood, E. High-intensity cannabis use associated with lower plasma human immunodeficiency virus-1 RNA viral load among recently infected people who use injection drugs. Drug Alcohol. Rev. 2015, 34, 135–140. [Google Scholar] [CrossRef]
- Okafor, C.N.; Zhou, Z.; Burrell, L.E., 2nd; Kelso, N.E.; Whitehead, N.E.; Harman, J.S.; Cook, C.L.; Cook, R.L. Marijuana use and viral suppression in persons receiving medical care for HIV-infection. Am. J. Drug Alcohol. Abuse 2017, 43, 103–110. [Google Scholar] [CrossRef]
- Slawek, D.E.; Arnsten, J.; Sohler, N.; Zhang, C.; Grossberg, R.; Stein, M.; Cunningham, C.O. Daily and near-daily cannabis use is associated with HIV viral load suppression in people living with HIV who use cocaine. AIDS Care 2021, 33, 1350–1357. [Google Scholar] [CrossRef]
- Ghosn, J.; Leruez-Ville, M.; Blanche, J.; Delobelle, A.; Beaudoux, C.; Mascard, L.; Lecuyer, H.; Canestri, A.; Landman, R.; Zucman, D.; et al. HIV-1 DNA levels in peripheral blood mononuclear cells and cannabis use are associated with intermittent HIV shedding in semen of men who have sex with men on successful antiretroviral regimens. Clin. Infect. Dis. 2014, 58, 1763–1770. [Google Scholar] [CrossRef]
- Trone, J.C.; Ollier, E.; Chapelle, C.; Bertoletti, L.; Cucherat, M.; Mismetti, P.; Magne, N.; Laporte, S. Statistical controversies in clinical research: Limitations of open-label studies assessing antiangiogenic therapies with regard to evaluation of vascular adverse drug events-a meta-analysis. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2018, 29, 803–811. [Google Scholar] [CrossRef]
- Chan, A.W.; Tetzlaff, J.M.; Gotzsche, P.C.; Altman, D.G.; Mann, H.; Berlin, J.A.; Dickersin, K.; Hrobjartsson, A.; Schulz, K.F.; Parulekar, W.R.; et al. SPIRIT 2013 explanation and elaboration: Guidance for protocols of clinical trials. BMJ 2013, 346, e7586. [Google Scholar] [CrossRef] [PubMed]
- Grotenhermen, F. Pharmacokinetics and pharmacodynamics of cannabinoids. Clin. Pharmacokinet. 2003, 42, 327–360. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, A.K.; Karanam, A.; Marino, S.E.; Barkley, C.M.; Remmel, R.P.; Roslawski, M.; Gramling-Aden, M.; Leppik, I.E. Food effect on pharmacokinetics of cannabidiol oral capsules in adult patients with refractory epilepsy. Epilepsia 2019, 60, 1586–1592. [Google Scholar] [CrossRef]
- Franco, V.; Gershkovich, P.; Perucca, E.; Bialer, M. The Interplay Between Liver First-Pass Effect and Lymphatic Absorption of Cannabidiol and Its Implications for Cannabidiol Oral Formulations. Clin. Pharmacokinet. 2020, 59, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Tyler, C.J.; Guzman, M.; Lundborg, L.R.; Yeasmin, S.; Perez-Jeldres, T.; Yarur, A.; Behm, B.; Dulai, P.S.; Patel, D.; Bamias, G.; et al. Inherent Immune Cell Variation Within Colonic Segments Presents Challenges for Clinical Trial Design. J. Crohn’s Colitis 2020, 14, 1364–1377. [Google Scholar] [CrossRef]
- Bowcutt, R.; Malter, L.B.; Chen, L.A.; Wolff, M.J.; Robertson, I.; Rifkin, D.B.; Poles, M.; Cho, I.; Loke, P. Isolation and cytokine analysis of lamina propria lymphocytes from mucosal biopsies of the human colon. J. Immunol. Methods 2015, 421, 27–35. [Google Scholar] [CrossRef]
- Papasavvas, E.; Azzoni, L.; Ross, B.N.; Fair, M.; Yuan, Z.; Gyampoh, K.; Mackiewicz, A.; Sciorillo, A.C.; Pagliuzza, A.; Lada, S.M.; et al. Intact Human Immunodeficiency Virus (HIV) Reservoir Estimated by the Intact Proviral DNA Assay Correlates With Levels of Total and Integrated DNA in the Blood During Suppressive Antiretroviral Therapy. Clin. Infect. Dis. 2021, 72, 495–498. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Total Population (n = 10) | |
|---|---|
| Age (Years), median (±IQR) | 57.5 (54.75–61.75) |
| Sex assigned at birth (n (%)) | |
| Male | 8 (80%) |
| Female | 2 (20%) |
| Ethnicity (n (%)) | |
| White-North American | 6 (60%) |
| Black-African | 1 (10%) |
| Asian | 1 (10%) |
| Mixed ethnicity | 2 (20%) |
| Antiretroviral regimens (n (%)) | |
| Biktarvy® (Bictegravir/Tenofovir alafenamide/Emtricitabine) | 5 (50%) |
| Triumeq® (Dolutegravir/Abacavir/Lamivudine) | 1 (10%) |
| Truvada®/Viramure® (Tenofovir/Emtricitabine/Nevirapine) | 1 (10%) |
| Raltegravir/Kivexa®/Biktarvy® (Raltegravir/Abacavir/Lamivudine/Bictegravir/Tenofovir alafenamide/Emtricitabine) | 1 (10%) |
| Genvoya® (Elvitegravir/Cobicistat/Emtricitabine/Tenofovir alafenamide) | 1 (10%) |
| Delstrigo® (Doravirine/Lamivudine/Tenofovir disoproxil fumarate) | 1 (10%) |
| Oral cannabinoids regimens (n (%)) | |
| TN-TC11M2 formulation (CBD: 2.5/THC: 2.5 mg) | 5 (50%) |
| TN-C200M2 formulation (CBD: 200 mg) | 5 (50%) |
| Cannabis use in the past 6 months before study initiation (n (%)) | |
| No | 3 (30%) |
| Yes | 7 (70%) |
| Monthly | 5 (72.43%) |
| Weekly | 2 (28.57%) |
| Daily | 0 (0%) |
| Alcohol use in the past 6 months (n (%)) | |
| No | 5 (50%) |
| Yes | 5 (50%) |
| Drug use in the past 6 months (n (%)) | |
| No | 3 (30%) |
| Yes | 7 (70%) |
| History of infectious diseases (n (%)) | |
| Syphilis (treated) | 2 (20%) |
| Hepatitis B (Anti-HBc Antibodies) | 4 (40%) |
| Hepatitis C (Anti-HCV Antibodies) | 0 (0%) |
| Plasma Markers | Study Time-Line | ||||||
|---|---|---|---|---|---|---|---|
|
Week 0 Treatment Initiation n = 10 | Week 1 n = 10 | Week 2 n = 10 | $ Week 6 n = 9 | $ Week 8 n = 8 | $ Week 12 End of Treatment n = 8 | $ Week 14 Study Termination n = 8 | |
| Gut mucosal damage | |||||||
| REG-3(pg/mL) [Mean (SD)] | 5621 (3299) b,e | 5390 (3294) g | 4663 (2970) b,g,l,m | 5610 (3403) l,q,r | 5315 (3140) m,s,t | 4950 (3018) e,q,s | 4757 (3366) r,t |
| [Median (IQR)] | 5608 (2617–7039) b,e | 5157 (2533–6751) g | 4893 (2220–5740) b,g,l,m | 5620 (2582- 6762) l,q,r | 5784 (2451–6430) m,s,t | 5612 (2074–6013) e,q,s | 4834 (1705–6048) r,t |
| I-FABP (pg/mL) [Mean (SD)] | 895.6 (774.2) | 1129 (1026) | 1117 (1160) | 794.5 (455.6) | 887.4 (768.6) | 802.3 (581.7) | 709.3 (526.4) |
| [Median (IQR)] | 660.6 (436.6–1275) | 731.1 (422.2–2001) | 717.8 (549.3–1174) | 557.5 (354.6–1202) | 665.8 (316.6–1368) | 658.0 (288.6–1338) | 517.6 (388.1–916.1) |
| Microbial translocation and immune activation | |||||||
| LPS (pg/mL) [Mean (SD)] | 98.7 (47.76) d | 99.75 (54.67) i | 89.3 (45.11) m | 104.7 (43.97) p | 138.3 (63.09) d,i,m,p,s,t | 112.2 (45.7) s | 94.73 (37.82) t |
| [Median (IQR)] | 88.29 (60.6–144.0) d | 93.18 (55.7–139.7) i | 76.56 (56.2–142.3) m | 102.8 (64.8–146.4) p | 131.9 (98.8–197.7) d,i,m,p,s,t | 115.8 (80.7–136.2) s | 98.78 (72.6–125.4) t |
| sCD14 (ng/mL) [Mean (SD)] | 2583 (536.9) | 2446 (447.3) h | 2605 (472.9) | 2855 (676.3) h,p,q | 2407 (399.2) p | 2491 (449.7) q | 2650 (602.7) |
| [Median (IQR)] | 2472 (2166–2825) | 2344 (2110–2877) h | 2493 (2248–2995) | 2766 (2371–3257) h,p,q | 2327 (2034–2797) p | 2371 (2181–2800) q | 2523 (2144–2945) |
| sCD27 (U/mL) [Mean (SD)] | 110.9 (22.73) | 113.3 (31.55) | 111.4 (26.81) | 113.3 (28.88) | 112.8 (35.98) | 109.3 (35.76) | 108.6 (28.85) |
| [Median (IQR)] | 114.6 (87.3–127.2) | 110.5 (93.4–122.3) | 105.7 (88.2–135.2) | 103.4 (88.8–137.8) | 108.6 (79.5–136.2) | 91.20 (83.5–141.9) | 106.3 (80.6–132.8) |
| sTNFRII (pg/mL) [Mean (SD)] | 2297 (665.4) d,e | 2279 (713.7) i,j | 2329 (800.1) n | 2631 (1076) | 2096 (481.1) d,i | 2027 (410.0) e,j,n | 2215 (740.8) |
| [Median (IQR)] | 2202 (1708–2823) d,e | 2037 (1680–3061) i,j | 2115 (1639–3158) n | 2608 (1752–3321) | 1971 (1727–2626) d,i | 1938 (1674–2353) e,j,n | 1957 (1719–3002) |
| Pro-inflammatory cytokines | |||||||
| TNF- (pg/mL) [Mean (SD)] | 2.61 (1.64) f | 2.18 (1.19) | 2.57 (2.17) o | 2.06 (1.10) | 2.27 (0.77) | 2.03(0.71) | 2.13 (1.48) f,o |
| [Median (IQR)] | 2.38 (1.38–3.63) f | 2.16 (0.87–3.21) | 1.82 (0.89–3.58) o | 1.93 (1.27–2.69) | 2.09 (1.54–2.91) | 1.94 (1.39–2.67) | 1.56 (0.95–3.78) f,o |
| INF- (pg/mL) [Mean (SD)] | 10.98 (18.11) e | 8.24 (11.43) | 14.31 (24.62) | 9.9 (13.52) q | 9.26 (11.73) | 8.55 (11.63) e,q | 9.93 (15.35) |
| [Median (IQR)] | 3.38 (0.53–13.63) e | 3.2 (0.53–13.28) | 2.28 (0.48–21.01) | 1.63 (0.52–21.62) q | 2.79 (0.54–18.82) | 3.32 (0.54–20.81) e,q | 2.66 (0.67–21.74) |
| IL-1 (pg/mL) [Mean (SD)] | 0.52 (0.45) | 0.62 (0.64) h,j | 0.58 (0.58) | 0.43 (0.34) h | 0.40 (0.20) | 0.40 (0.24) j | 0.62 (0.72) |
| [Median (IQR)] | 0.44 (0.24–0.59) | 0.47 (0.18–0.68) h,j | 0.46 (0.26–0.59) | 0.38 (0.14–0.65) h | 0.46 (0.17–0.60) | 0.44 (0.15–0.52) j | 0.42 (0.19–0.66) |
| IL-6 (pg/mL) [Mean (SD)] | 1.44 (1.00) | 1.11 (0.75) | 1.36 (0.97) | 1.36 (0.91) | 1.40 (1.15) | 1.22 (0.97) | 1.29 (1.13) |
| [Median (IQR)] | 1.14 (0.51–2.30) | 0.91 (0.41–1.91) | 1.30 (0.55–1.86) | 0.92 (0.63–2.30) | 0.87(0.63–1.92) | 0.88 (0.47–1.69) | 1.07 (0.38–1.72) |
| IL-8 (pg/mL) [Mean (SD)] | 5.50 (3.5) | 5.79 (5.6) | 5.12 (3.18) | 5.53 (2.88) | 6.20 (2.41) t | 5.61 (2.99) | 4.21 (1.42) t |
| [Median (IQR)] | 4.97 (3.05–7.38) | 3.88 (2.08–8.10) | 4.89 (2.80–7.67) | 5.94 (2.74–7.92) | 5.84 (4.00–8.37) t | 5.76 (2.66–7.59) | 4.68 (2.86–5.32) t |
| IP-10 (pg/mL) [Mean (SD)] | 55.99 (36.72) | 55.5 (33.47) h | 51.26 (24.24) | 50.55 (26.04) h,p | 61.80 (35.97) p | 56.68 (25.52) | 52.59 (28.58) |
| [Median (IQR)] | 43.74 (30.11–77.91) | 43.18 (31.62–76.80) h | 42.97 (32.30–74.06) | 41.51 (35.77–69.95) h,p | 50.05 (36.14–95.64) p | 48.65 (42.49–81.62) | 41.80 (33.77–75.17) |
| Anti-inflammatory cytokine | |||||||
| IL-10 (pg/mL) [Mean (SD)] | 1.26 (1.37) | 0.92 (0.54) | 1.0 (0.68) | 0.93 (0.44) | 1.1 (0.85) | 1.1 (0.74) | 0.93 (0.49) |
| [Median (IQR)] | 0.77 (0.49–1.55) | 0.79 (0.51–1.28) | 0.85 (0.47–1.26) | 0.86 (0.53–1.29) | 0.82 (0.50–1.36) | 0.87 (0.60–1.52) | 0.90 (0.52–1.18) |
| Cellular Immune Markers | Study Time-Line | ||||||
|---|---|---|---|---|---|---|---|
|
Week 0 Treatment Initiation n = 10 |
Week 1 n = 10 |
Week 2 n = 10 | $
Week 6 n = 9 | $
Week 8 n = 8 | $
Week 12 End of Treatment n = 8 | $
Week 14 Study Termination n = 8 | |
| CD4 T-Cells | |||||||
| Memory T-Cell Subsets | |||||||
| Naïve (CD45RA+CD28+CCR7+) % [Mean (SD)] | 78.2 (13.1) | 79.6 (11.0) | 79.4 (12.5) | 78.0 (14.3) | 76.7 (13.5) | 76.2 (13.6) | 73.9 (15.6) |
| [Median (IQR)] | 83.4 (66.3–87.4) | 83.8 (74.2–86.6) | 82.0 (74.4–87.4) | 86.2 (65.4–87.7) | 81.9 (63.1–85) | 80.7 (63.9–84.8) | 78.7 (59.1–85.4) |
| Central Memory (CD45RA−CD28+CCR7+) % [Mean (SD)] | 32.9 (11.7) | 33.6 (10.6) | 35.3 (12.7) | 34.4 (12.7) | 32.5 (11.5) | 32.7 (12.8) | 32.7 (12.4) |
| [Median (IQR)] | 36.6 (22.3–39.6) | 33.3 (28.3–40.1) | 34.5 (29.3–42.2) | 34.4 (25.8–44.1) | 34.7 (24.8–38.6) | 34.5 (21.1–40.9) | 33.8 (22.8–39.7) |
| Transitional Memory (CD45RA−CD28+CCR7−) % [Mean (SD)] | 64.1 (11.1) | 63.8 (10.6) | 62.4 (12.5) | 63.0 (12.6) | 64.9 (11.4) | 64.4 (12.7) | 63.6 (13.3) |
| [Median (IQR)] | 62 (57.2–71.7) | 63.4 (57.6–69.6) | 62.6 (52.2–68.3) | 60.9 (52.2–73.2) | 63.6 (56.6–73.0) | 61.5 (55.3–77.4) | 62.3 (50.7–75.7) |
| Effector Memory (CD45RA−CD28−CCR7−) % [Mean (SD)] | 2.6 (3.2) c | 2.3 (2.8) i | 2.1 (2.5) | 2.3 (2.7) c | 2.4 (2.7) i | 2.6 (2.3) | 3.2 (3.9) |
| [Median (IQR)] | 1.0 (0.3–5.9) c | 1.0 (0.4–4.6) i | 1.1 (0.3–3.7) | 0.8 (0.3–4.5) c | 1.2 (0.4–5.1) i | 1.4 (0.8–5.3) | 1.6 (0.5–5.6) |
| Terminally Differentiated (CD45RA+CD28−CCR7−) % [Mean (SD)] | 2.1 (4.4) b | 2.1 (4.6) g,i | 1.6 (3.9) b,g | 2.1 (4.4) | 1.8 (4.2) i | 1.8 (3.6) | 2.4 (5.6) |
| [Median (IQR)] | 0.2 (0.04–1.9) b | 0.2 (0.04–1.6) g,i | 0.1 (0.03–1.1) b,g | 0.2 (0.05–2.2) | 0.1 (0.09–1.3) i | 0.2 (0.07–1.8) | 0.1 (0.05–1.8) |
| T-cell functions | |||||||
| HLADR+CD38+ (%) [Mean (SD)] | 3.1 (1.6) | 4.0 (3.1) | 3.8 (4.1) | 2.8 (1.7) | 2.7 (1.7) | 3.6 (2.7) | 3.2 (3.6) |
| [Median (IQR)] | 2.9 (1.6–4.8) | 3.8 (1.6–4.9) | 2.3 (1.6–4.6) | 1.7 (1.3–4.4) | 2.2 (1.5–4.6) | 2.5 (1.3–6.5) | 1.8 (1.2–4.2) |
| CD45RA−Ki-67+ (%) [Mean (SD)] | 0.8 (0.3) e | 0.9 (0.4) | 0.9 (0.4) | 1.0 (0.5) | 1.0 (0.6) | 1.0 (0.4) e | 0.8 (0.3) |
| [Median (IQR)] | 0.7 (0.5–1.0) e | 0.8 (0.5–1.3) | 0.8 (0.5–1.2) | 1.0 (0.6–1.2) | 0.8 (0.6–1.2) | 0.9 (0.7–1.2) e | 0.7 (0.7–0.9) |
| CD45RA−PD-1+ (%) [Mean (SD)] | 32.3 (9.5) f | 32.4 (9.5) j,k | 31.4 (9.5) n,o | 32.2 (10.1) r | 32.6 (10.3) | 30.6 (11.0) j,n | 30.8 (9.2) f,k,o,r |
| [Median (IQR)] | 30.5 (26.3–39.0) f | 30.7 (24.9–38.3) j,k | 30.6 (23.8–37.5) n,o | 32.2 (23.0–37.8) r | 31.2 (24.5–41.7) | 31.3 (22.6–39.2) j,n | 30.5 (22.6–37.8) f,k,o,r |
| CD45RA−CTLA-4+ (%) [Mean (SD)] | 3.0 (1.4) | 3.2 (1.5) | 3.3 (1.7) | 3.6 (2.4) | 3.9 (2.2) | 3.6 (1.4) | 3.8 (1.6) |
| [Median (IQR)] | 3.1 (1.8–4.2) | 2.5 (2.0–4.9) | 2.5 (2.0–4.9) | 3.2 (1.7–5.0) | 3.2 (2.4–5.4) | 3.8 (2.3–5.1) | 3.9 (2.2–4.7) |
| Senescent (CD28−CD57+) (%) [Mean (SD)] | 1.8 (2.5) | 1.5 (2.1) i | 1.3 (1.7) | 1.5 (2.0) | 1.4 (1.6) i | 1.3 (1.2) | 2.1 (3.2) |
| [Median (IQR)] | 0.5 (0.07–4.0) | 0.7 (0.05–3.0) i | 0.7 (0.04–2.3) | 0.7 (0.1–2.6) | 0.7 (0.2–2.2) i | 0.8 (0.5–2.0) | 0.9 (0.3–2.2) |
| CD4+CD39+ (%) [Mean (SD)] | 6.1 (4.7) | 6.3 (5.1) g | 5.9 (4.8) g | 6.4 (5.1) r | 6.4 (5.3) t | 5.9 (5.7) | 5.5 (4.8) r,t |
| [Median (IQR)] | 5.1 (2.1–9.0) | 5.0 (1.9–9.0) g | 4.9 (1.8–8.1) g | 5.8 (1.5–10.3) r | 5.0 (1.8–11.4) t | 5.1 (1.3–7.9) | 4.8 (1.3–7.8) r,t |
| CD4+CD73+ (%) [Mean (SD)] | 6.9 (3.1) d,f | 6.4 (2.6) | 6.6 (2.8) o | 6.2 (3.0) | 6.1 (2.8) d | 6.3 (3.7) | 5.8 (2.9) f,o |
| [Median (IQR)] | 7.1 (4.6–9.2) d,f | 6.8 (4.5–7.9) | 6.5 (4.9–7.9) o | 6.2 (3.9–7.3) | 6.6 (3.6–7.0) d | 6.1 (3.4–6.8) | 5.6 (3.3–6.9) f,o |
| Chemokine receptors expression | |||||||
| CD45RA−CCR4+ (%)[Mean (SD)] | 29.7 (11.4) | 29.2 (10.2) | 30.1 (10.8) | 31.4 (9.5) | 30.5 (9.8) | 29.4 (10.1) | 29.1 (10.4) |
| [Median (IQR)] | 31 (20.5–36.6) | 31.6 (21.1–36.0) | 31.7 (20.8–38.0) | 34.4 (23.2–36.1) | 31.8 (20.3–36.6) | 30.1 (18.6–38.4) | 30.6 (18.3–38.1) |
| CD45RA−CCR6+ (%) [Mean (SD)] | 9.8 (6.1) | 12.6 (7.1) k | 11.7 (5.0) | 15.0 (10.8) | 12.0 (7.2) | 10.9 (4.3) | 9.7 (6.4) k |
| [Median (IQR)] | 8.8 (5.7–12.4) | 11.8 (7.1–19.0) k | 11.2 (8.3–16.2) | 10.6 (7.4–20.8) | 11.4 (8.3–13.3) | 10.3 (8.5–13.2) | 6.7 (5.7–15.5) k |
| CD45RA−CXCR3+ (%) [Mean (SD)] | 2.6 (2.9) | 3.8 (5.2) | 4.1 (3.8) | 5.7 (3.6) | 2.4 (3.1) | 3.5 (3.2) | 3.1 (4.7) |
| [Median (IQR)] | 1.7 (0.3–4.1) | 2.4 (0.4–4.1) | 3.0 (1.8–6.1) | 4.9 (3.0–9.1) | 0.8 (0.5–5.2) | 2.3 (1.7–5.3) | 1.2 (0.9–3.5) |
| Th subsets | |||||||
| Th17 (CD45RA−CCR4+CCR6+ CXCR3−) (%) [Mean (SD)] | 5.2 (3.0) | 6.8 (4.0) k | 6.4 (2.8) | 8.0 (5.9) | 6.3 (3.6) | 5.8 (2.5) | 5.2 (3.5) k |
| [Median (IQR)] | 5.2 (2.8–7.2) | 5.5 (4.4–10.3) k | 6.0 (4.8–9.2) | 6.9 (3.4–10.7) | 5.7 (4.8–7.1) | 5.4 (5.0–6.7) | 3.8 (2.4–8.2) k |
| Th1-Th17 (CD45RA−CCR4−CCR6+CXCR3+) (%) [Mean (SD)] | 0.36 (0.6) c | 0.62 (0.9) k | 0.6 (0.9) | 1.0 (1.1) c | 0.5 (0.9) | 0.3 (0.3) | 0.2 (0.3) k |
| [Median (IQR)] | 0.2 (0.02–0.4) c | 0.3 (0.05–0.8) k | 0.3 (0.1–0.7) | 0.4 (0.3–1.7) c | 0.06 (0.03–0.9) | 0.2 (0.1–0.4) | 0.1 (0.04–0.2) k |
| Th2 (CD45RA−CCR4+CCR6−CXCR3−) (%) [Mean (SD)] | 23.0 (10.5) c | 20.7 (7.6) | 21.9 (8.9) | 20.9 (6.5) c | 22.9 (10.4) | 22.1 (8.9) | 22.6 (8.5) |
| [Median (IQR)] | 21.9 (13.7–32.6) c | 20.0 (14.6–29.1) | 20.7 (14.7–30.7) | 20.9 (14.7–26.9) c | 21.7 (13.4–31.9) | 20.9 (14.0–30.0) | 23.7 (13.6–29.6) |
| Th1 (CD45RA−CCR4−CCR6−CXCR3+) (%) [Mean (SD)] | 1.7 (1.7) | 2.4 (3.6) | 2.7 (2.5) | 3.2 (1.8) p | 1.4 (1.6) p | 2.5 (2.3) | 2.4 (3.8) |
| [Median (IQR)] | 1.3 (0.3–3.0) | 1.3 (0.3–2.5) | 2.0 (1.0–4.5) | 3.6 (1.9–4.7) p | 0.6 (0.4–2.9) p | 1.8 (1.0–4.4) | 0.9 (0.7–2.5) |
| Regulatory T-cells | |||||||
| Treg (CD25hi CD127lo FoxP3+) (%) [Mean (SD)] | 2.9 (1.6) | 2.7 (1.4) | 2.9 (1.2) | 3.2 (1.6) | 3.4 (1.7) | 2.6 (1.1) | 2.9 (1.1) |
| [Median (IQR)] | 2.0 (1.5–4.5) | 2.6 (1.5–3.6) | 2.5 (1.9–3.9) | 2.7 (1.8–4.4) | 3.7 (1.6–4.8) | 2.0 (1.8–3.8) | 2.6 (2.2–4.1) |
| CD73+ Treg (%) [Mean (SD)] | 5.0 (2.4) f | 4.9 (2.6) j,k | 4.8 (2.5) | 4.2 (2.2) | 4.5 (2.1) | 4.2 (1.7) j | 4.2 (1.7) f,k |
| [Median (IQR)] | 4.9 (3.0–7.4) f | 4.1 (3.3–7.3) j,k | 4.4 (2.9–7.2) | 3.5 (3.0–5.8) | 4.3 (3.0–6.6) | 3.9 (3.5–5.7) j | 3.9 (3.4–5.9) f,k |
| CD39+ Treg (%) [Mean (SD)] | 45.1 (21.1) | 45.4 (23.0) | 44.0 (22.2) n | 44.1 (24.5) | 41.3 (23.3) | 41.3 (21.7) n | 41.0 (20.4) |
| [Median (IQR)] | 41.5 (28.5–62.5) | 46.3 (23.4–65.6) | 43.4 (25.2–65.4) n | 43.2 (24.1–69.2) | 36.5 (21.5–66.6) | 37.4 (23.3–59.8) n | 38.1 (25.1–63.3) |
| CD8 T-cells | |||||||
| Memory T-cell subsets | |||||||
| Naïve (CD45RA+CD28+CCR7+) % [Mean (SD)] | 50.4 (25.3) b,d | 52.3 (24.7) i | 54.6 (25.3) b | 51.8 (24.7) | 52.8 (26.8) d,i | 48.2 (25.9) | 52.8 (25.6) |
| [Median (IQR)] | 48.7 (24.4–77.7) b,d | 53.4 (26.5–77.4) i | 55.3 (29.8–80.1) b | 51.2 (27.4–74.2) | 57.5 (25.9–77.7) d,i | 47.0 (24.0–67.5) | 56.2 (28.9–77.8) |
| Central Memory (CD45RA−CD28+CCR7+) % [Mean (SD)] | 9.7 (6.1) | 10.0 (4.7) k | 10.8 (5.3) | 12.1 (6.3) | 11.8 (4.5) | 11.5 (4.5) | 12.1 (5.3) k |
| [Median (IQR)] | 7.6 (4.9–15.9) | 8.6 (5.7–14.6) k | 8.2 (6.5–17.4) | 9.5 (5.8–18.4) | 12.9 (6.8–16.0) | 10.8 (7.3–15.1) | 12.6 (6.4–17.6) k |
| Transitional Memory (CD45RA−CD28+CCR7−) % [Mean (SD)] | 66.7 (12.0) | 67.3 (11.1) | 67.5 (10.7) | 65.2 (9.7) | 67.4 (9.5) | 66.3 (8.1) | 64.7 (13.1) |
| [Median (IQR)] | 70.5 (58.7–75.2) | 72.0 (58.5–74.7) | 69.4 (58.0–74.7) | 69.2 (59.0–72.8) | 72.1 (61.3–73.6) | 70.7 (59.2–72.0) | 70.6 (57.0–72.5) |
| Effector Memory (CD45RA−CD28−CCR7−)% [Mean (SD)] | 22.8 (15.6) | 21.9 (13.9) | 21.0 (13.6) | 21.7 (14.2) | 19.8 (12.8) | 21.2 (10.6) | 21.8 (15.8) |
| [Median (IQR)] | 17.1 (9.8–35.7) | 15.6 (9.8–33.8) | 14.8 (9.3–34.5) | 19.2 (7.9–32.2) | 14.1 (9.9–30.9) | 16.9 (13.3–33.4) | 14.1 (10.7–35.3) |
| Terminally Differentiated (CD45RA+CD28−CCR7−) % [Mean (SD)] | 31.2 (22.2) | 30.0 (22.7) | 28.5 (22.0) | 31.4 (21.6) | 29.8 (23.4) | 33.0 (23.9) | 29.6 (22.5) |
| [Median (IQR)] | 35.5 (5.7–49.3) | 32.1 (4.4–54.1) | 29.5 (4.2–51.6) | 30.6 (9.7–53.1) | 26.5 (6.4–55.4) | 38.4 (6.5–55.4) | 28.6 (6.9–48.2) |
| T-cell functions | |||||||
| HLA-DR+CD38+ (%) [Mean (SD)] | 4.9 (4.1) | 5.3 (4.3) | 5.2 (5.0) | 5.1 (3.8) | 6.1 (6.7) | 6.1 (5.7) | 5.4 (5.0) |
| [Median (IQR)] | 3.6 (2.0–7.4) | 4.3 (2.6–6.9) | 3.5 (1.6–8.2) | 4.4 (2.2–7.8) | 3.5 (1.8–9.0) | 3.4 (2.7–11.1) | 3.8 (2.1–8.5) |
| CD45RA−Ki-67+ (%) [Mean (SD)] | 0.39 (0.19) a | 0.51 (0.25) a | 0.49 (0.27) | 0.55 (0.40) | 0.58 (0.63) | 0.52 (0.18) | 0.58 (0.46) |
| [Median (IQR)] | 0.41 (0.25–0.56) a | 0.47 (0.33–0.73) a | 0.47 (0.32–0.74) | 0.42 (0.35–0.54) | 0.36 (0.28–0.49) | 0.53 (0.34–0.68) | 0.36 (0.30–0.96) |
| CD45RA−PD-1+ (%) [Mean (SD)] | 37.9 (13.1) | 37.9 (11.9) k | 37.7 (14.9) | 35.2 (10.8) | 37.1 (11.6) t | 34.9 (12.8) | 34.7 (10.2) k,t |
| [Median (IQR)] | 37.0 (31.0–45.3) | 38.1 (29.5–46.5) k | 35.3 (27.5–47.8) | 35.9 (27.1–45.7) | 36.4 (29.4–49.2) t | 37.2 (25.1–47.1) | 36.7 (27.2–42.9) k,t |
| CD45RA−CTLA-4+ (%) [Mean (SD)] | 1.0 (0.7) | 1.3 (1.0) | 1.4 (0.9) | 2.1 (3.1) | 1.1 (1.0) | 1.0 (0.6) | 1.3 (0.9) |
| [Median (IQR)] | 0.9 (0.5–1.2) | 1.0 (0.6–1.7) | 1.2 (0.6–2.2) | 0.8 (0.5–2.7) | 0.9 (0.4–1.4) | 1.0 (0.6–1.1) | 1.0 (0.7–2.0) |
| Senescent (CD28−CD57+) (%) [Mean (SD)] | 18.5 (14.5) c,d | 17.0 (13.5) | 16.4 (12.6) m | 17.9 (13.5) c | 16.2 (12.5) d,m | 17.0 (11.1) | 18.1 (15.4) |
| [Median (IQR)] | 16.1 (4.9–31.8) c,d | 13.6 (4.5–31.4) | 12.5 (4.6–29.6) m | 16.8 (5.7–31.6) c | 10.6 (6.4–27.7) d,m | 14.9 (5.8–28.2) | 11.4 (7.2–29.9) |
| CD8+CD39+ (%) [Mean (SD)] | 2.3 (1.4) | 2.6 (1.9) | 2.5 (1.9) | 2.6 (2.2) r | 2.3 (1.9) | 2.2 (1.6) | 2.0 (1.6) r |
| [Median (IQR)] | 2.8 (0.5–3.4) | 2.6 (0.6–3.9) | 2.7 (0.4–3.8) | 2.8 (0.3–4.5) r | 2.1 (0.4–3.8) | 2.1 (0.6–3.4) | 2.2 (0.3–3.3) r |
| CD8+CD73+ (%) [Mean (SD)] | 36.6 (24.4) b | 36.9 (24.6) | 38.1 (24.7) b | 32.9 (19.8) | 32.9 (20.7) | 30.9 (21.4) | 32.1 (21.3) |
| [Median (IQR)] | 35.3 (10.4–54.4) b | 35.8 (10.5–54.3) | 39.5 (11.7–55.6) b | 36.6 (10.8–50.2) | 36.6 (10.8–51.8) | 30.6 (9.2–52.9) | 34.8 (9.3–50.1) |
| CD8+FoxP3+ (%) [Mean (SD)] | 0.29 (0.23) | 0.28 (0.19) | 0.40 (0.29) n | 0.39 (0.35) q | 0.42 (0.51) | 0.20 (0.19) n,q | 0.28 (0.16) |
| [Median (IQR)] | 0.19 (0.13–0.56) | 0.21 (0.17–0.35) | 0.37 (0.12–0.60) n | 0.29 (0.11–0.67) q | 0.14 (0.08–0.80) | 0.12 (0.09–0.29) n,q | 0.25 (0.13–0.44) |
| Chemokine receptors expression | |||||||
| CD45RA−CCR4+ (%) [Mean (SD)] | 15.8 (10.6) | 15.2 (7.5) | 15.6 (8.4) | 18.2 (10.3) | 17.6 (9.5) | 17.7 (11.4) | 18.2 (9.3) |
| [Median (IQR)] | 13.7 (8.1–21.9) | 13.2 (8.6–21.6) | 13.9 (9.0–20.9) | 15.5 (8.1–28.8) | 17.2 (10.2–26.8) | 14.1 (9.4–31.0) | 17.0 (9.6–26.0) |
| CD45RA−CCR6+ (%) [Mean (SD)] | 4.4 (4.2) | 4.9 (3.8) | 4.5 (3.0) | 5.3 (4.2) | 5.1 (4.4) t | 4.0 (2.4) | 4.0 (3.8) t |
| [Median (IQR)] | 3.8 (1.4–5.5) | 3.5 (1.6–6.7) | 3.0 (2.1–7.5) | 4.5 (2.4–6.8) | 3.9 (2.1–6.2) t | 3.2 (2.5–5.7) | 3.1 (1.9–3.8) t |
| CD45RA−CXCR3+ (%) [Mean (SD)] | 4.1 (3.1) b | 6.0 (4.7) | 7.1 (5.5) b | 9.3 (6.4) | 4.7 (4.1) | 6.4 (5.2) | 5.9 (4.2) |
| [Median (IQR)] | 4.0 (1.5–6.3) b | 5.1 (1.8–9.2) | 5.6 (3.2–11.8) b | 8.7 (3.7–14.7) | 2.7 (2.1–8.6) | 4.6 (2.8–10.3) | 3.9 (2.7–10.0) |
| Monocytes | |||||||
| Classical monocytes | |||||||
| Classical (CD14++CD16−) (%) [Mean (SD)] | 78.0 (13.0) | 78.2 (11.8) | 77.5 (10.8) | 75.1 (16.7) | 74.9 (14.5) | 74.7 (14.6) | 73.0 (16.0) |
| [Median (IQR)] | 78.6 (69.7–90.9) | 80.3 (65.4–86.2) | 80.4 (65.5–87.1) | 83.5 (57.6–89.2) | 77.1 (58.4–89.1) | 69.2 (62.6–91.8) | 80.4 (59.9–86.1) |
| Classical CD163+ (%) [Mean (SD)] | 46.4 (19.2) | 44.7 (23.3) | 54.1 (26.7) | 50.8 (19.3) | 56.0 (20.4) t | 43.5 (19.0) | 46.1 (21.0) t |
| [Median (IQR)] | 40.0 (32.0–62.2) | 39.3 (31.6–64.8) | 56.5 (30.3–79.1) | 48.2 (33.8–62.1) | 57.2 (41.0–73.5) t | 49.2 (24.7–53.3) | 41.1 (32.0–64.0) t |
| Classical CX3CR1+ (%) [Mean (SD)] | 51.2 (14.3) b | 52.7 (14.4) | 60.3 (15.8) b | 51.5 (13.4) | 52.3 (11.9) | 52.1 (11.9) | 55.7 (12.9) |
| [Median (IQR)] | 53.8 (38.3–61.3) b | 56.6 (40.8–62.3) | 59.7 (54.3–69.0) b | 56.5 (38.1–63.1) | 51.9 (44.8–64.4) | 56.4 (41.6–61.8) | 57.4 (47.1–66.7) |
| Classical M-DC8+ (%) [Mean (SD)] | 2.56 (3.72) | 3.24 (6.73) k | 3.53 (6.94) o | 1.61 (2.00) | 3.18 (6.28) t | 2.11 (2.93) | 2.64 (5.78) k,o,t |
| [Median (IQR)] | 0.61 (0.42–4.26) | 0.75 (0.48–2.62) k | 0.64 (0.56–3.90) o | 0.72 (0.41–2.31) | 0.74 (0.46–2.42) t | 0.61 (0.38–4.86) | 0.49 (0.33–1.43) k,o,t |
| Classical CCR2+ (%) [Mean (SD)] | 91.6 (3.1) | 92.6 (2.6) | 92.9 (3.2) | 92.1 (3.6) | 93.5 (2.9) | 92.5 (4.6) | 92.2 (5.7) |
| [Median (IQR)] | 92.3 (89.9–93.3) | 92.3 (90.8–94.8) | 93.4 (91.1–95.7) | 91.1 (89.7–95.0) | 94.0 (91.6–95.1) | 93.4 (91.0–95.2) | 94.4 (89.0–95.3) |
| Intermediate monocytes | |||||||
| Intermediate (CD14+CD16+) (%) [Mean (SD)] | 11.0 (4.9) | 10.4 (4.6) | 10.5 (4.4) | 12.0 (7.2) | 12.0 (5.0) | 12.9 (6.7) | 12.7 (5.7) |
| [Median (IQR)] | 11.0 (6.0–15.3) | 10.5 (6.4–14.9) | 12.0 (6.2–13.7) | 9.1 (6.5–21.0) | 14.3 (6.4–15.2) | 14.4 (5.5–18.9) | 11.5 (8.1–18.0) |
| Intermediate CD163+ (%) [Mean (SD)] | 66.2 (11.7) | 66.7 (15.5) | 74.0 (17.6) | 70.4 (12.5) | 73.5 (12.4) t | 64.5 (14.2) | 65.9 (15.0) t |
| [Median (IQR)] | 65.2 (59.4–72.2) | 69.7 (58.1–76.2) | 74.0 (60.4–88.1) | 69.4 (60.3–77.5) | 71.4 (64.9–84.2) t | 59.6 (54.2–78.9) | 64.7 (54.6–78.6) t |
| Intermediate CX3CR1+ (%) [Mean (SD)] | 86.1 (8.2) | 86.8 (8.4) | 88.3 (8.6) | 88.3 (7.3) p | 86.0 (10.5) p | 85.8 (9.8) | 88.8 (7.2) |
| [Median (IQR)] | 87.0 (84.7–91.2) | 87.2 (83.6–94.1) | 89.3 (83.6–94.9) | 91.3 (85.7–92.8) p | 89.3 (84.5–92.2) p | 89.3 (78.6–91.6) | 90.6 (85.9–93.4) |
| Intermediate M-DC8+ (%) [Mean (SD)] | 15.5 (7.7) | 17.9 (10.6) j | 18.8 (13.0) | 15.2 (6.5) | 17.2 (11.2) | 13.4 (7.7) j | 14.6 (6.6) |
| [Median (IQR)] | 11.9 (9.1–23.6) | 15.7 (7.8–26.6) j | 16.1 (9.3–26.1) | 14.1 (11.1–21.1) | 16.0 (7.1–25.5) | 13.4 (6.7–20.0) j | 14.7 (8.3–19.1) |
| Intermediate CCR2+ (%) [Mean (SD)] | 89.3 (2.9) | 88.6 (2.1) | 89.0 (5.5) | 89.6 (1.6) | 90.1 (3.4) | 89.6 (4.5) | 89.6 (4.3) |
| [Median (IQR)] | 88.8 (87.8–91.2) | 88.8 (87.2–90.6) | 91.3 (86.5–92.4) | 90.0 (88.3–91.1) | 90.6 (87.7–92.9) | 91.0 (87.9–92.2) | 89.4 (88.9–93.2) |
| Non classical monocytes | |||||||
| Non classical (CD16++CD14−) (%) [Mean (SD)] | 11.0 (8.8) | 11.4 (7.5) | 12.1 (7.6) | 12.9 (10.2) | 13.0 (10.9) | 12.3 (9.5) | 14.3 (12.2) |
| [Median (IQR)] | 9.3 (3.7–16.5) | 10.7 (5.2–19.9) | 10.2 (5.0–19.4) | 7.1 (5.4–23.1) | 8.3 (4.7–25.7) | 12.2 (3.3–18.7) | 8.6 (6.7–19.1) |
| Non classical CD163+ (%) [Mean (SD)] | 31.2 (7.2) d | 33.7 (9.1) | 38.3 (15.9) | 35.3 (8.2) r | 35.7 (9.5) d | 32.6 (6.3) | 30.9 (6.8) r |
| [Median (IQR)] | 29.7 (26.3–38.0) d | 35.0 (25.8–41.4) | 31.6 (29.3–42.8) | 35.7 (30.0–40.0) r | 31.7 (30.2–46.4) d | 31.8 (27.6–38.5) | 30.2 (25.6–35.7) r |
| Non classical CX3CR1+ (%) [Mean (SD)] | 75.6 (11.5) | 74.8 (14.0) | 76.3 (14.8) | 76.9 (12.2) | 74.6 (15.5) | 75.5 (12.4) | 77.6 (14.9) |
| [Median (IQR)] | 74.7 (71.6–84.0) | 73.7 (67.2–85.9) | 81.9 (61.1–87.6) | 78.9 (65.5–86.6) | 81.0 (57.4–83.8) | 75.5 (63.6–87.9) | 84.5 (60.3–88.7) |
| Non classical M-DC8+ (%) [Mean (SD)] | 28.4 (10.3) | 30.8 (15.3) | 31.3 (18.6) | 25.6 (7.5) | 28.8 (10.8) | 23.2 (7.0) | 26.8 (10.5) |
| [Median (IQR)] | 24.7 (21.5–39.8) | 28.5 (17.8–37.7) | 25.1 (20.5–34.4) | 27.2 (19.1–32.5) | 31.0 (17.9–37.6) | 23.4 (17.6–26.8) | 24.7 (18.7–37.5) |
| Non classical CCR2+ (%) [Mean (SD)] | 7.2 (3.3) e | 8.1 (2.4) | 8.1 (4.7) | 7.2 (5.9) | 7.9 (3.3) | 12.1 (8.2) e | 7.9 (3.8) |
| [Median (IQR)] | 6.3 (4.5–10.5) e | 7.8 (6.8–9.6) | 8.3 (5.4–9.3) | 5.0 (3.9–8.8) | 6.8 (5.7–11.3) | 9.3 (6.7–18.1) e | 7.9 (5.1–11.2) |
| Dendritic cells (DC) | |||||||
| Plasmacytoid DC (CD123+CD11c−) (%) [Mean (SD)] | 4.4 (2.1) | 3.6 (1.1) | 3.8 (1.3) | 3.6 (1.5) | 4.2 (1.1) | 3.8 (1.7) | 4.1 (1.1) |
| [Median (IQR)] | 3.7 (2.7–5.5) | 3.8 (2.5–4.4) | 3.7 (3.1–5.1) | 3.7 (2.4–4.4) | 4.1 (3.2–5.0) | 3.8 (2.5–4.7) | 4.2 (3.3–4.9) |
| Myeloid DC (CD123−CD11c+) (%) [Mean (SD)] | 8.8 (3.2) b,e | 11.5 (2.7) | 12.3 (4.0) b | 10.3 (7.9) | 8.8 (3.9) s | 12.5 (4.8) e,s | 12.6 (6.6) |
| [Median (IQR)] | 8.1 (7.0–10.4) b,e | 11.2 (9.4–14.1) | 11.8 (9.9–13.7) b | 7.8 (5.4–12.0) | 8.1 (5.7–10.5) s | 11.5 (9.4–15.6) e,s | 13.0 (6.4–17.7) |
| HIV DNA and RNA | Study Timeline | ||||||
|---|---|---|---|---|---|---|---|
| Week 0 Treatment Initiation n = 10 | Week 1 n = 10 | Week 2 n = 10 | $ Week 6 n = 9 | $ Week 8 n = 8 | $ Week 12 End of Treatment n = 8 | $ Week 14 Study Termination n = 8 | |
| Total HIV DNA (copies/106 CD4) [Mean (SD)] | 1016 (1081) | 908.1 (817.3) | 920.4 (816.5) | 1053 (1121) | 1292 (1310) | 979.1 (1015) | 1143 (1318) |
| [Median (IQR)] | 708.1 (125.1–1679) | 765.3 (239.3–1449) | 819.8 (176.2–1529) | 911.9 (141- 1886) | 790.3 (165.6–2843) | 629.9 (155.5–2156) | 658.5 (152.3–2317) |
| LTR-gag cell-associated RNA (copies/106 CD4) [Mean (SD)] | 493.5 (557.3) | 593.9 (817.3) i | 529.2 (630.3) | 476.3 (479.6) | 1370 (1687) i,s | 572.9 (752.8) s | 751.9 (858.3) |
| [Median (IQR)] | 308.8 (4.9–915.8) | 224.3 (3.5–1088) i | 271.2 (13.4–1081) | 281.1 (21.9–938.8) | 142 (10.1–3151) i,s | 283.9 (2.1–1290) s | 386.9 (26.0–1770) |
| RNA/DNA ratio [Mean (SD)] | 0.47 (0.45) | 0.56 (0.55) | 0.43 (0.41) | 0.43 (0.37) | 0.62 (0.62) s | 0.44 (0.53) s | 1.85 (3.53) |
| [Median (IQR)] | 0.3 (0.2–1.0) | 0.46 (0.08–0.87) | 0.37 (0.08–0.76) | 0.33 (0.12–0.84) | 0.44 (0.05–1.26) s | 0.17 (0.01–1.07) s | 0.50 (0.28–1.54) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mboumba Bouassa, R.-S.; Comeau, E.; Alexandrova, Y.; Pagliuzza, A.; Yero, A.; Samarani, S.; Needham, J.; Singer, J.; Lee, T.; Bobeuf, F.; et al. Effects of Oral Cannabinoids on Systemic Inflammation and Viral Reservoir Markers in People with HIV on Antiretroviral Therapy: Results of the CTN PT028 Pilot Clinical Trial. Cells 2023, 12, 1811. https://doi.org/10.3390/cells12141811
Mboumba Bouassa R-S, Comeau E, Alexandrova Y, Pagliuzza A, Yero A, Samarani S, Needham J, Singer J, Lee T, Bobeuf F, et al. Effects of Oral Cannabinoids on Systemic Inflammation and Viral Reservoir Markers in People with HIV on Antiretroviral Therapy: Results of the CTN PT028 Pilot Clinical Trial. Cells. 2023; 12(14):1811. https://doi.org/10.3390/cells12141811
Chicago/Turabian StyleMboumba Bouassa, Ralph-Sydney, Eve Comeau, Yulia Alexandrova, Amélie Pagliuzza, Alexis Yero, Suzanne Samarani, Judy Needham, Joel Singer, Terry Lee, Florian Bobeuf, and et al. 2023. "Effects of Oral Cannabinoids on Systemic Inflammation and Viral Reservoir Markers in People with HIV on Antiretroviral Therapy: Results of the CTN PT028 Pilot Clinical Trial" Cells 12, no. 14: 1811. https://doi.org/10.3390/cells12141811
APA StyleMboumba Bouassa, R.-S., Comeau, E., Alexandrova, Y., Pagliuzza, A., Yero, A., Samarani, S., Needham, J., Singer, J., Lee, T., Bobeuf, F., Vertzagias, C., Sebastiani, G., Margolese, S., Mandarino, E., Klein, M. B., Lebouché, B., Routy, J.-P., Chomont, N., Costiniuk, C. T., & Jenabian, M.-A. (2023). Effects of Oral Cannabinoids on Systemic Inflammation and Viral Reservoir Markers in People with HIV on Antiretroviral Therapy: Results of the CTN PT028 Pilot Clinical Trial. Cells, 12(14), 1811. https://doi.org/10.3390/cells12141811