

Interplay between Protein Kinase C Epsilon and Reactive Oxygen Species during Myogenic Differentiation
 , ,
, ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Gene Silencing
2.2.1. Short Hairpin RNA (shRNA) Cell Infection
2.2.2. Small Interfering RNA (siRNA) Cell Transfection
2.3. Measurement of ROS
2.3.1. ROS Detection with Flow Cytometry
2.3.2. ROS Detection with Immunofluorescence
2.4. RNA Extraction and Quantitative RT-PCR
2.5. Analysis of Protein Content with Western Blot
2.6. Immunofluorescence
2.7. Immunohistochemistry
2.8. Immunoprecipitation
2.9. Statistical Analysis
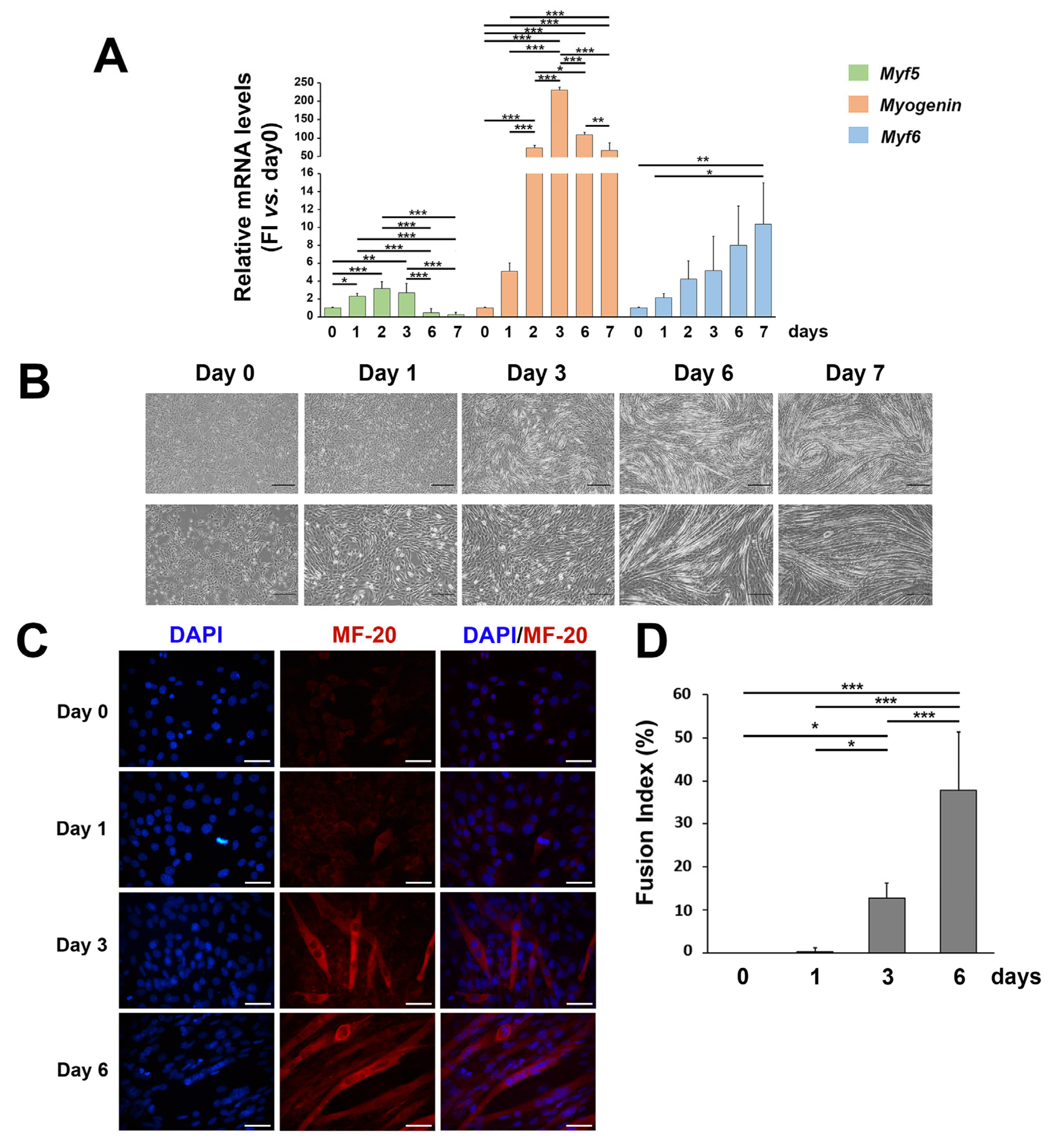
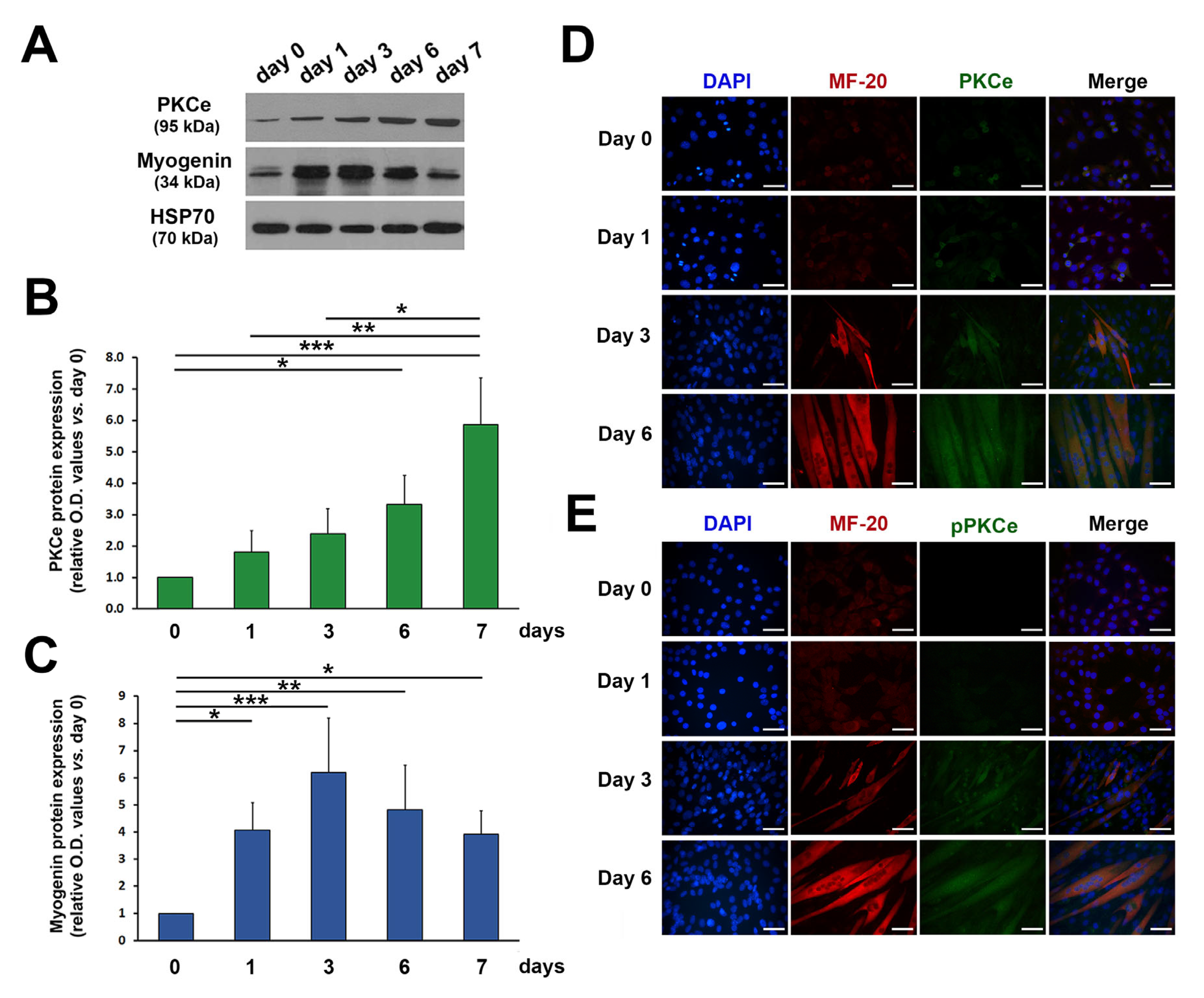
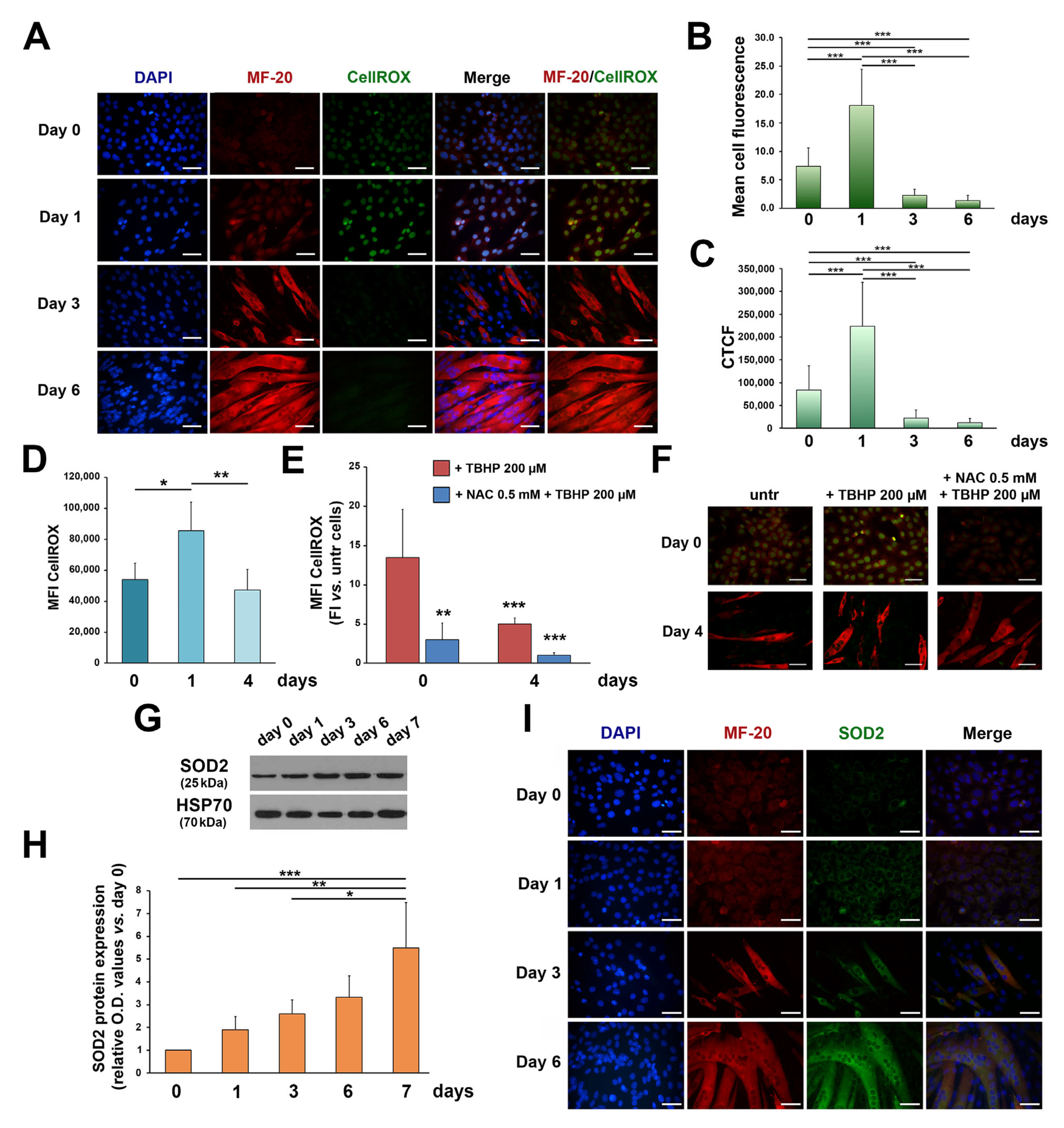
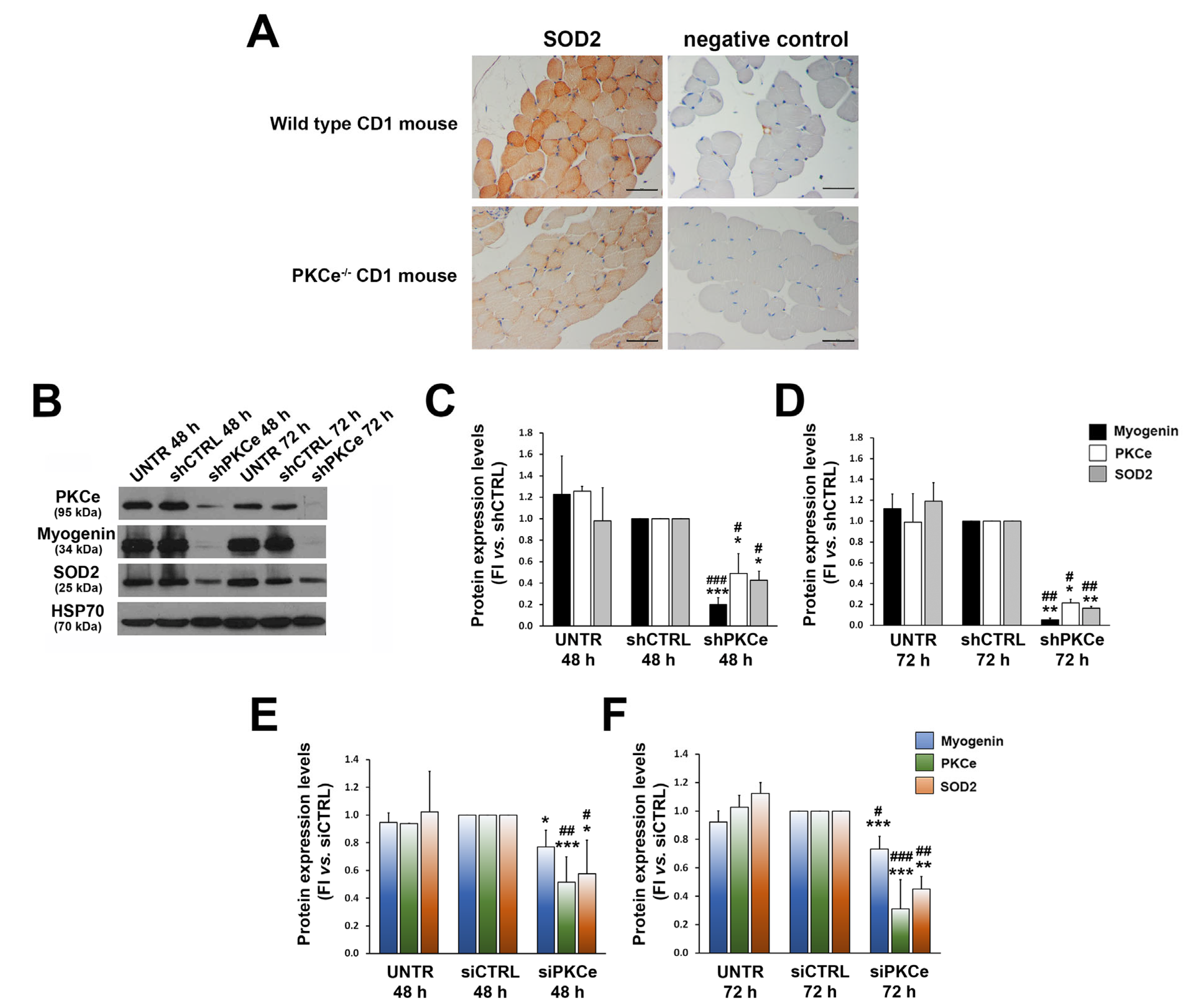
3. Results
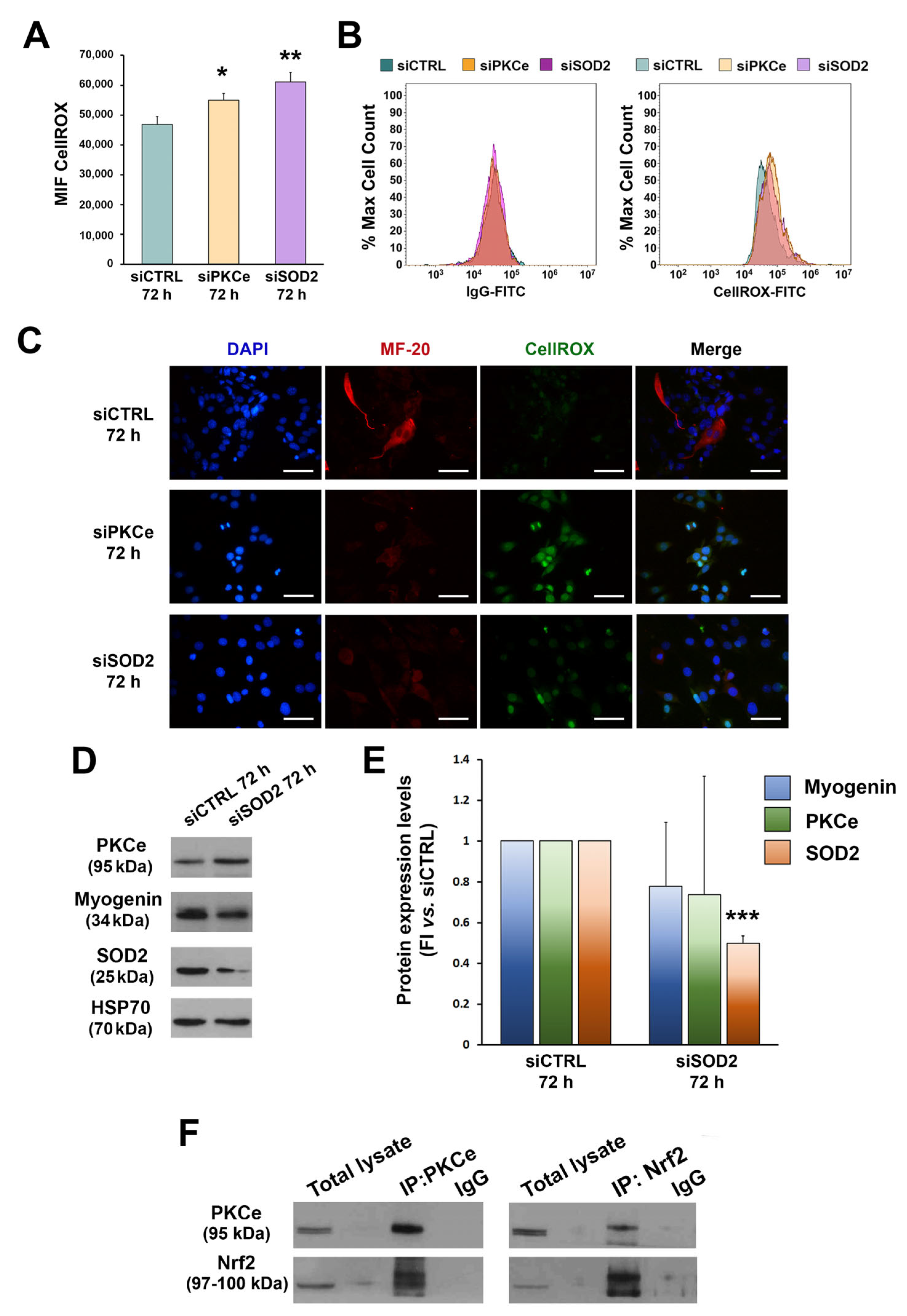
3.1. ROS and SOD2 Protein Content Are Modulated during Myogenic Differentiation
3.2. PKCe Down-Regulation Affects SOD2 Protein Content via Nrf2
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zuo, L.; Pannell, B.K. Redox Characterization of Functioning Skeletal Muscle. Front. Physiol. 2015, 6, 338. [Google Scholar] [CrossRef] [PubMed]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building Muscle: Molecular Regulation of Myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef]
- Ardite, E.; Barbera, J.A.; Roca, J.; Fernández-Checa, J.C. Glutathione Depletion Impairs Myogenic Differentiation of Murine Skeletal Muscle C2C12 Cells through Sustained NF-ΚB Activation. Am. J. Pathol. 2004, 165, 719–728. [Google Scholar] [CrossRef]
- Fulle, S.; Protasi, F.; Di Tano, G.; Pietrangelo, T.; Beltramin, A.; Boncompagni, S.; Vecchiet, L.; Fanò, G. The Contribution of Reactive Oxygen Species to Sarcopenia and Muscle Ageing. Exp. Gerontol. 2004, 39, 17–24. [Google Scholar] [CrossRef]
- Hansen, J.; Klass, M.; Harris, C.; Csete, M. A Reducing Redox Environment Promotes C2C12 Myogenesis: Implications for Regeneration in Aged Muscle. Cell Biol. Int. 2007, 31, 546–553. [Google Scholar] [CrossRef]
- Sestili, P.; Barbieri, E.; Martinelli, C.; Battistelli, M.; Guescini, M.; Vallorani, L.; Casadei, L.; D’Emilio, A.; Falcieri, E.; Piccoli, G.; et al. Creatine Supplementation Prevents the Inhibition of Myogenic Differentiation in Oxidatively Injured C2C12 Murine Myoblasts. Creatine supplementation prevents the inhibition of myogenic differentiation in oxidatively injured C2C12 murine myoblasts. Mol. Nutr. Food Res. 2009, 53, 1187–1204. [Google Scholar] [CrossRef] [PubMed]
- Sandiford, S.D.; Kennedy, K.A.; Xie, X.; Pickering, J.; Li, S.S. Dual Oxidase Maturation Factor 1 (DUOXA1) Overexpression Increases Reactive Oxygen Species Production and Inhibits Murine Muscle Satellite Cell Differentiation. Cell Commun. Signal. 2014, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Kozakowska, M.; Pietraszek-Gremplewicz, K.; Jozkowicz, A.; Dulak, J. The Role of Oxidative Stress in Skeletal Muscle Injury and Regeneration: Focus on Antioxidant Enzymes. J. Muscle Res. Cell Motil. 2015, 36, 377–393. [Google Scholar] [CrossRef] [PubMed]
- Guttridge, D.C.; Mayo, M.W.; Madrid, L.V.; Wang, C.-Y.; Baldwin, A.S., Jr. NF-ΚB-Induced Loss of MyoD Messenger RNA: Possible Role in Muscle Decay and Cachexia. Science 2000, 289, 2363–2366. [Google Scholar] [CrossRef]
- Guttridge, D.C.; Albanese, C.; Reuther, J.Y.; Pestell, R.G.; Baldwin, A.S. NF-ΚB Controls Cell Growth and Differentiation through Transcriptional Regulation of Cyclin D1. Mol. Cell Biol. 1999, 19, 5785–5799. [Google Scholar] [CrossRef]
- Wang, H.; Hertlein, E.; Bakkar, N.; Sun, H.; Acharyya, S.; Wang, J.; Carathers, M.; Davuluri, R.; Guttridge, D.C. NF-ΚB Regulation of YY1 Inhibits Skeletal Myogenesis through Transcriptional Silencing of Myofibrillar Genes. Mol. Cell Biol. 2007, 27, 4374–4387. [Google Scholar] [CrossRef] [PubMed]
- Rochard, P.; Rodier, A.; Casas, F.; Cassar-Malek, I.; Marchal-Victorion, S.; Daury, L.; Wrutniak, C.; Cabello, G. Mitochondrial Activity Is Involved in the Regulation of Myoblast Differentiation through Myogenin Expression and Activity of Myogenic Factors. J. Biol. Chem. 2000, 275, 2733–2744. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, E.; Sestili, P. Reactive Oxygen Species in Skeletal Muscle Signaling. J. Signal. Transduct. 2012, 2012, 982794. [Google Scholar] [CrossRef] [PubMed]
- Langen, R.C.J.; Schols, A.M.W.J.; Kelders, M.C.J.M.; van der Velden, J.L.J.; Wouters, E.F.M.; Janssen-Heininger, Y.M.W. Tumor Necrosis Factor-α Inhibits Myogenesis through Redox-Dependent and -Independent Pathways. Am. J. Physiol. Cell Physiol. 2002, 283, C714–C721. [Google Scholar] [CrossRef]
- Langen, R.C.J.; Velden, J.L.J.; Schols, A.M.W.J.; Kelders, M.C.J.M.; Wouters, E.F.M.; Janssen-Heininger, Y.M.W. Tumor Necrosis Factor-Alpha Inhibits Myogenic Differentiation through MyoD Protein Destabilization. FASEB J. 2004, 18, 227–237. [Google Scholar] [CrossRef]
- Fedorova, M.; Kuleva, N.; Hoffmann, R. Reversible and Irreversible Modifications of Skeletal Muscle Proteins in a Rat Model of Acute Oxidative Stress. Biochim. Biophys. Acta 2009, 1792, 1185–1193. [Google Scholar] [CrossRef]
- Le Moal, E.; Pialoux, V.; Juban, G.; Groussard, C.; Zouhal, H.; Chazaud, B.; Mounier, R. Redox Control of Skeletal Muscle Regeneration. Antioxid. Redox Signal. 2017, 27, 276–310. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, N.S.; Shelar, S.B.; Jones, D.P.; Hoidal, J.R. Reductive Stress Impairs Myogenic Differentiation. Redox Biol. 2020, 34, 101492. [Google Scholar] [CrossRef] [PubMed]
- Lyons, C.N.; Leary, S.C.; Moyes, C.D. Bioenergetic remodeling during cellular differentiation: Changes in Cytochrome c Oxidase Regulation Do Not Affect the Metabolic Phenotype. Biochem. Cell Biol. 2004, 82, 391–399. [Google Scholar] [CrossRef]
- Jahnke, V.E.; Sabido, O.; Freyssenet, D. Control of mitochondrial biogenesis, ROS level, and cytosolic Ca2+ concentration during the cell cycle and the onset of differentiation in L6E9 myoblasts. Am. J. Physiol. Cell Physiol. 2009, 296, C1185–C1194. [Google Scholar] [CrossRef]
- Malinska, D.; Kudin, A.P.; Bejtka, M.; Kunz, W.S. Changes in Mitochondrial Reactive Oxygen Species Synthesis during Differentiation of Skeletal Muscle Cells. Mitochondrion 2012, 12, 144–148. [Google Scholar] [CrossRef]
- Bakkar, N.; Guttridge, D.C. NF-kappaB signaling: A tale of two pathways in skeletal myogenesis. Physiol. Rev. 2010, 90, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Togliatto, G.; Trombetta, A.; Dentelli, P.; Cotogni, P.; Rosso, A.; Tschöp, M.H.; Granata, R.; Ghigo, E.; Brizzi, M.F. Unacylated ghrelin promotes skeletal muscle regeneration following hindlimb ischemia via SOD-2-mediated miR-221/222 expression. J. Am. Heart Assoc. 2013, 2, e000376. [Google Scholar] [CrossRef] [PubMed]
- He, W.A.; Berardi, E.; Cardillo, V.M.; Acharyya, S.; Aulino, P.; Thomas-Ahner, J.; Wang, J.; Bloomston, M.; Muscarella, P.; Nau, P.; et al. NF-κB-mediated Pax7 dysregulation in the muscle microenvironment promotes cancer cachexia. J. Clin. Investig. 2013, 123, 4821–4835. [Google Scholar] [CrossRef]
- Brzeszczyńska, J.; Johns, N.; Schilb, A.; Degen, S.; Degen, M.; Langen, R.; Schols, A.; Glass, D.J.; Roubenoff, R.; Greig, C.A.; et al. Loss of oxidative defense and potential blockade of satellite cell maturation in the skeletal muscle of patients with cancer but not in the healthy elderly. Aging 2016, 8, 1690–1702. [Google Scholar] [CrossRef]
- Jang, Y.C.; Lustgarten, M.S.; Liu, Y.; Muller, F.L.; Bhattacharya, A.; Liang, H.; Salmon, A.B.; Brooks, S.V.; Larkin, L.; Hayworth, C.R.; et al. Increased superoxide in vivo accelerates age-associated muscle atrophy through mitochondrial dysfunction and neuromuscular junction degeneration. FASEB J. 2010, 24, 1376–1390. [Google Scholar] [CrossRef]
- Jang, Y.C.; Liu, Y.; Hayworth, C.R.; Bhattacharya, A.; Lustgarten, M.S.; Muller, F.L.; Chaudhuri, A.; Qi, W.; Li, Y.; Huang, J.Y.; et al. Dietary restriction attenuates age-associated muscle atrophy by lowering oxidative stress in mice even in complete absence of CuZnSOD. Aging Cell 2012, 11, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.C.; Rodriguez, K.; Lustgarten, M.S.; Muller, F.L.; Bhattacharya, A.; Pierce, A.; Choi, J.J.; Lee, N.H.; Chaudhuri, A.; Richardson, A.G.; et al. Superoxide-mediated oxidative stress accelerates skeletal muscle atrophy by synchronous activation of proteolytic systems. GeroScience 2020, 42, 1579–1591. [Google Scholar] [CrossRef]
- Zhang, Y.; Davis, C.; Sakellariou, G.K.; Shi, Y.; Kayani, A.C.; Pulliam, D.; Bhattacharya, A.; Richardson, A.; Jackson, M.J.; McArdle, A.; et al. CuZnSOD gene deletion targeted to skeletal muscle leads to loss of contractile force but does not cause muscle atrophy in adult mice. FASEB J. 2013, 27, 3536–3548. [Google Scholar] [CrossRef]
- Sakellariou, G.K.; McDonagh, B.; Porter, H.; Giakoumaki, I.I.; Earl, K.E.; Nye, G.A.; Vasilaki, A.; Brooks, S.V.; Richardson, A.; Van Remmen, H.; et al. Comparison of Whole Body SOD1 Knockout with Muscle-Specific SOD1 Knockout Mice Reveals a Role for Nerve Redox Signaling in Regulation of Degenerative Pathways in Skeletal Muscle. Antioxid. Redox Signal. 2018, 28, 275–295. [Google Scholar] [CrossRef]
- Gordon, S.J.V.; Fenker, D.E.; Vest, K.E.; Padilla-Benavides, T. Manganese influx and expression of ZIP8 is essential in primary myoblasts and contributes to activation of SOD2. Metallomics 2019, 11, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, A.; Muller, F.L.; Liu, Y.; Ng, R.; Faulkner, J.; Hamilton, M.; Richardson, A.; Huang, T.T.; Epstein, C.J.; Van Remmen, H. Alterations in mitochondrial function, hydrogen peroxide release and oxidative damage in mouse hind-limb skeletal muscle during aging. Mech. Ageing Dev. 2006, 127, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Lustgarten, M.S.; Jang, Y.C.; Liu, Y.; Qi, W.; Qin, Y.; Dahia, P.L.; Shi, Y.; Bhattacharya, A.; Muller, F.L.; Shimizu, T.; et al. MnSOD deficiency results in elevated oxidative stress and decreased mitochondrial function but does not lead to muscle atrophy during aging. Aging Cell 2011, 10, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, A.; Yang, C.; Liu, Y.; Tan, Y.; Bond, S.T.; Walker, S.; Sikora, T.; Laskowski, A.; Sharma, A.; de Haan, J.B.; et al. SOD2 in skeletal muscle: New insights from an inducible deletion model. Redox Biol. 2021, 47, 102135. [Google Scholar] [CrossRef] [PubMed]
- Di Marcantonio, D.; Galli, D.; Carubbi, C.; Gobbi, G.; Queirolo, V.; Martini, S.; Merighi, S.; Vaccarezza, M.; Maffulli, N.; Sykes, S.M.; et al. PKCε as a novel promoter of skeletal muscle differentiation and regeneration. Exp. Cell Res. 2015, 339, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, G.; Galli, D.; Carubbi, C.; Neri, L.M.; Masselli, E.; Pozzi, G.; Vitale, M.; Mirandola, P. PKC Proteins and Muscular Dystrophy. J. Funct. Morphol. Kinesiol. 2018, 3, 12. [Google Scholar] [CrossRef]
- Mirandola, P.; Gobbi, G.; Ponti, C.; Sponzilli, I.; Cocco, L.; Vitale, M. PKCε controls protection against TRAIL in erythroid progenitors. Blood 2006, 107, 508–513. [Google Scholar] [CrossRef]
- Gobbi, G.; Mirandola, P.; Sponzilli, I.; Micheloni, C.; Malinverno, C.; Cocco, L.; Vitale, M. Timing and Expression Level of Protein Kinase Cε Regulate the Megakaryocytic Differentiation of Human CD34 Cells. Stem Cells. 2007, 25, 2322–2329. [Google Scholar] [CrossRef]
- Gobbi, G.; Di Marcantonio, D.; Micheloni, C.; Carubbi, C.; Galli, D.; Vaccarezza, M.; Bucci, G.; Vitale, M.; Mirandola, P. TRAIL Up-Regulation Must Be Accompanied by a Reciprocal PKCε Down-Regulation during Differentiation of Colonic Epithelial Cell: Implications for Colorectal Cancer Cell Differentiation. J. Cell Physiol. 2011, 227, 630–638. [Google Scholar] [CrossRef]
- Gobbi, G.; Mirandola, P.; Carubbi, C.; Masselli, E.; Sykes, S.M.; Ferraro, F.; Nouvenne, A.; Thon, J.N.; Italiano, J.E.; Vitale, M. Proplatelet Generation in the Mouse Requires PKCε-Dependent RhoA Inhibition. Blood 2013, 122, 1305–1311. [Google Scholar] [CrossRef]
- Galli, D.; Gobbi, G.; Carrubbi, C.; Marcantonio, D.; Benedetti, L.; De Angelis, M.G.C.; Meschi, T.; Vaccarezza, M.; Sampaolesi, M.; Mirandola, P.; et al. The Role of PKCε-Dependent Signaling for Cardiac Differentiation. Histochem. Cell Biol. 2012, 139, 35–46. [Google Scholar] [CrossRef]
- Masselli, E.; Carubbi, C.; Gobbi, G.; Mirandola, P.; Galli, D.; Martini, S.; Bonomini, S.; Crugnola, M.; Craviotto, L.; Aversa, F.; et al. Protein Kinase Cɛ Inhibition Restores Megakaryocytic Differentiation of Hematopoietic Progenitors from Primary Myelofibrosis Patients. Leukemia 2015, 29, 2192–2201. [Google Scholar] [CrossRef] [PubMed]
- Galli, D.; Carubbi, C.; Masselli, E.; Corradi, D.; Dei Cas, A.; Nouvenne, A.; Bucci, G.; Arcari, M.L.; Mirandola, P.; Vitale, M.; et al. PKCε Is a Negative Regulator of PVAT-Derived Vessel Formation. Exp. Cell Res. 2015, 330, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Queirolo, V.; Galli, D.; Masselli, E.; Borzì, R.M.; Martini, S.; Vitale, F.; Gobbi, G.; Carubbi, C.; Mirandola, P. PKCε Is a Regulator of Hypertrophic Differentiation of Chondrocytes in Osteoarthritis. Osteoarthr. Cartil. 2016, 24, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Carubbi, C.; Masselli, E.; Martini, S.; Galli, D.; Aversa, F.; Mirandola, P.; Italiano, J.E.; Gobbi, G.; Vitale, M. Human Thrombopoiesis Depends on Protein Kinase Cδ /Protein Kinase Cε Functional Couple. Haematologica 2016, 101, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Masselli, E.; Carubbi, C.; Pozzi, G.; Martini, S.; Aversa, F.; Galli, D.; Gobbi, G.; Mirandola, P.; Vitale, M. Platelet Expression of PKCepsilon Oncoprotein in Myelofibrosis Is Associated with Disease Severity and Thrombotic Risk. Ann. Transl. Med. 2017, 5, 273. [Google Scholar] [CrossRef] [PubMed]
- Martini, S.; Pozzi, G.; Carubbi, C.; Masselli, E.; Galli, D.; Di Nuzzo, S.; Banchini, A.; Gobbi, G.; Vitale, M.; Mirandola, P. PKCε Promotes Human Th17 Differentiation: Implications in the Pathophysiology of Psoriasis. Eur. J. Immunol. 2018, 48, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, G.; Mirandola, P.; Carubbi, C.; Galli, D.; Vitale, M. Protein kinase C ε in hematopoiesis: Conductor or selector? Semin. Thromb. Hemost. 2013, 39, 59–65. [Google Scholar] [CrossRef]
- Parker, P.J.; Lockwood, N.; Davis, K.; Kelly, J.R.; Soliman, T.N.; Pardo, A.L.; Marshall, J.J.T.; Redmond, J.M.; Vitale, M.; Martini, S. A cancer-associated, genome protective programme engaging PKCε. Adv. Biol. Regul. 2020, 78, 100759. [Google Scholar] [CrossRef]
- Dobrowolny, G.; Martini, M.; Scicchitano, B.M.; Romanello, V.; Boncompagni, S.; Nicoletti, C.; Pietrangelo, L.; De Panfilis, S.; Catizone, A.; Bouchè, M.; et al. Muscle Expression of SOD1G93A Triggers the Dismantlement of Neuromuscular Junction Via PKC-Theta. Antioxid. Redox Signal. 2018, 28, 1105–1119. [Google Scholar] [CrossRef]
- Di Marcantonio, D.; Martinez, E.; Sidoli, S.; Vadaketh, J.; Nieborowska-Skorska, M.; Gupta, A.; Meadows, J.M.; Ferraro, F.; Masselli, E.; Challen, G.A.; et al. Protein Kinase c Epsilon Is a Key Regulator of Mitochondrial Redox Homeostasis in Acute Myeloid Leukemia. Clin. Cancer Res. 2018, 24, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishna, R.; Jaken, S. Protein Kinase C Signaling and Oxidative Stress. Free. Radic. Biol. Med. 2000, 9, 1349–1361. [Google Scholar] [CrossRef] [PubMed]
- Del Carlo, M.; Loeser, R.F. Chondrocyte cell death mediated by reactive oxygen species-dependent activation of PKC-betaI. Am. J. Physiol. Cell Physiol. 2006, 290, 802–811. [Google Scholar] [CrossRef] [PubMed]
- Cosentino-Gomes, D.; Rocco-Machado, N.; Meyer-Fernandes, J.R. Cell Signaling through Protein Kinase c Oxidation and Activation. Int. J. Mol. Sci. 2012, 13, 10697–10721. [Google Scholar] [CrossRef]
- Giorgi, C.; Agnoletto, C.; Baldini, C.; Bononi, A.; Bonora, M.; Marchi, S.; Missiroli, S.; Patergnani, S.; Poletti, F.; Rimessi, A.; et al. Redox Control of Protein Kinase C: Cell- and Disease-Specific Aspects. Antioxid. Redox Signal. 2010, 13, 1051–1085. [Google Scholar] [CrossRef]
- Korchak, H.M.; Rossi, M.W.; Kilpatrick, L.E. Selective Role for β-Protein Kinase c in Signaling for O-2 Generation but Not Degranulation or Adherence in Differentiated HL60 Cells. J. Biol. Chem. 1998, 16, 27292–27299. [Google Scholar] [CrossRef]
- El-Benna, J.; Dang, P.M.-C.; Gougerot-Pocidalo, M.-A.; Marie, J.-C.; Braut-Boucher, F. P47phox, the Phagocyte NADPH Oxidase/NOX2 Organizer: Structure, Phosphorylation and Implication in Diseases. Exp. Mol. Med. 2009, 30, 217–225. [Google Scholar] [CrossRef]
- Plumb, R.D.; El-Sherbeeny, N.A.; Dixon, L.J.; Hughes, S.M.T.; Devine, A.B.; Leahey, W.J.; McVeigh, G.E. NAD(P)H-Dependent Superoxide Production in Platelets: The Role of Angiotensin II and Protein Kinase C. Clin. Biochem. 2005, 38, 607–613. [Google Scholar] [CrossRef]
- Griendling, K.K.; Ushio-Fukai, M. Reactive Oxygen Species as Mediators of Angiotensin II Signaling. Regul. Pept. 2000, 28, 21–27. [Google Scholar] [CrossRef]
- Chung, Y.W.; Kim, H.K.; Kim, I.Y.; Yim, M.B.; Chock, P.B. Dual function of protein kinase C (PKC) in 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced manganese superoxide dismutase (MnSOD) expression: Activation of CREB and FOXO3a by PKC-alpha phosphorylation and by PKC-mediated inactivation of Akt, respectively. J. Biol. Chem. 2011, 286, 29681–29690. [Google Scholar]
- Sen, A.; Nelson, T.J.; Alkon, D.L.; Hongpaisan, J. Loss in PKC Epsilon Causes Downregulation of MnSOD and BDNF Expression in Neurons of Alzheimer’s Disease Hippocampus. J. Alzheimers Dis. 2018, 63, 1173–1189. [Google Scholar] [CrossRef] [PubMed]
- Mylroie, H.; Dumont, O.; Bauer, A.; Thornton, C.C.; Mackey, J.; Calay, D.; Hamdulay, S.S.; Choo, J.R.; Boyle, J.J.; Samarel, A.M.; et al. PKCε-CREB-Nrf2 signalling induces HO-1 in the vascular endothelium and enhances resistance to inflammation and apoptosis. Cardiovasc. Res. 2015, 106, 509–519. [Google Scholar] [CrossRef]
- Huang, H.C.; Nguyen, T.; Pickett, C.B. Regulation of the antioxidant response element by protein kinase C-mediated phosphorylation of NF-E2-related factor 2. Proc. Natl. Acad. Sci. USA 2000, 97, 12475–12480. [Google Scholar] [CrossRef]
- Ivaska, J.; Vuoriluoto, K.; Huovinen, T.; Izawa, I.; Inagaki, M.; Parker, P.J. PKCɛ-Mediated Phosphorylation of Vimentin Controls Integrin Recycling and Motility. EMBO J. 2005, 24, 3834–3845. [Google Scholar] [CrossRef] [PubMed]
- Saurin, A.T.; Durgan, J.; Cameron, A.J.; Faisal, A.; Marber, M.S.; Parker, P.J. The Regulated Assembly of a PKCɛ Complex Controls the Completion of Cytokinesis. Nat. Cell Biol. 2008, 10, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Ludin, A.; Gur-Cohen, S.; Golan, K.; Kaufmann, K.B.; Itkin, T.; Medaglia, C.; Lu, X.-J.; Ledergor, G.; Kollet, O.; Lapidot, T. Reactive Oxygen Species Regulate Hematopoietic Stem Cell Self-Renewal, Migration and Development, as Well as Their Bone Marrow Microenvironment. Antioxid. Redox Signal. 2014, 21, 1605–1619. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Tak, E.; Lee, J.; Rashid, M.; Murphy, M.P.; Ha, J.; Kim, S.S. Mitochondrial H2O2 Generated from Electron Transport Chain Complex I Stimulates Muscle Differentiation. Cell Res. 2011, 21, 817–834. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Protein Name NCBI Reference Sequence Identifier | Forward Primer | Reverse Primer |
|---|---|---|
| Mrf4 (Myf6) (NM_008657.3) | GAGATTCTGCGGAGTGCCAT | TTCTTGCTTGGGTTTGTAGC |
| Myog (NM_031189.2) | ATCCAGTACATTGAGCGCCT | GCAAATGATCTCCTGGGTTG |
| Myf5 (NM_008656.5) | TGAGGGAACAGGTGGAGAAC | AGCTGGACACGGAGCTTTTA |
| Gusb (NM_001357025.1) | CCGCTGAGAGTAATCGGAAAC | TCTCGCAAAATAAAGGCCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pozzi, G.; Presta, V.; Masselli, E.; Condello, G.; Cortellazzi, S.; Arcari, M.L.; Micheloni, C.; Vitale, M.; Gobbi, G.; Mirandola, P.; et al. Interplay between Protein Kinase C Epsilon and Reactive Oxygen Species during Myogenic Differentiation. Cells 2023, 12, 1792. https://doi.org/10.3390/cells12131792
Pozzi G, Presta V, Masselli E, Condello G, Cortellazzi S, Arcari ML, Micheloni C, Vitale M, Gobbi G, Mirandola P, et al. Interplay between Protein Kinase C Epsilon and Reactive Oxygen Species during Myogenic Differentiation. Cells. 2023; 12(13):1792. https://doi.org/10.3390/cells12131792
Chicago/Turabian StylePozzi, Giulia, Valentina Presta, Elena Masselli, Giancarlo Condello, Samuele Cortellazzi, Maria Luisa Arcari, Cristina Micheloni, Marco Vitale, Giuliana Gobbi, Prisco Mirandola, and et al. 2023. "Interplay between Protein Kinase C Epsilon and Reactive Oxygen Species during Myogenic Differentiation" Cells 12, no. 13: 1792. https://doi.org/10.3390/cells12131792
APA StylePozzi, G., Presta, V., Masselli, E., Condello, G., Cortellazzi, S., Arcari, M. L., Micheloni, C., Vitale, M., Gobbi, G., Mirandola, P., & Carubbi, C. (2023). Interplay between Protein Kinase C Epsilon and Reactive Oxygen Species during Myogenic Differentiation. Cells, 12(13), 1792. https://doi.org/10.3390/cells12131792