
β-Catenin Elicits Drp1-Mediated Mitochondrial Fission Activating the Pro-Apoptotic Caspase-1/IL-1β Signalosome in Aeromonas hydrophila-Infected Zebrafish Macrophages

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Care and Maintenance
2.3. Bacterial Culture
2.4. ZKM Isolation
2.5. Infection Studies
2.6. Reagents and Chemicals
2.7. Apoptosis Studies
2.8. RNA Isolation, cDNA Synthesis and RT-qPCR
2.9. siRNA Transfection
2.10. Intracellular Bacterial Quantification
2.11. β-Catenin Assay
2.12. Superoxide Measurement
2.13. mtROS Production
2.14. Mitochondrial Membrane Potential (ΔΨm) Assay
2.15. Measurement of Cyt c
2.16. Mitochondrial Morphology Assessment
2.17. IL-1β Assay
2.18. Caspase-1 and Caspase-3 Assays
2.19. Statistical Analysis
3. Results
3.1. A. hydrophila Induces ZKM Apoptosis
3.2. A. hydrophila Induces Canonical Wnt Signaling in ZKM
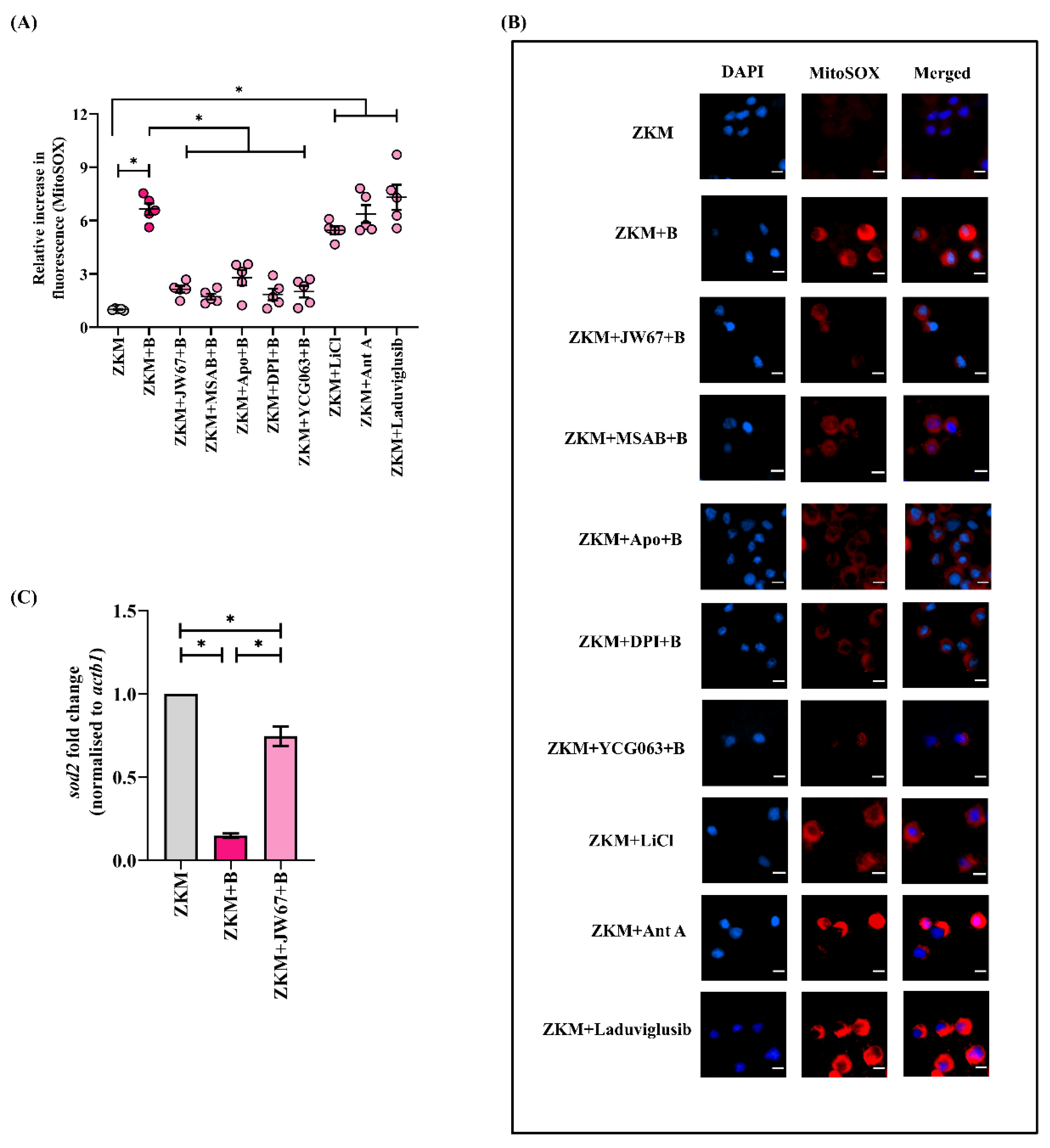
3.3. β-Catenin Mediates NOX-Dependent Oxidative Stress in A. hydrophila-Infected ZKM
3.4. β-Catenin-Mediated Cytosolic ROS Positively Impacts mtROS Generation
3.5. β-Catenin Alters ΔΨm in A. hydrophila-Infected ZKM
3.6. β-Catenin-Induced mtROS Abets Mitochondrial Fission in A. hydrophila-Infected ZKM
3.7. β-Catenin-Induced Mitochondrial Fission Favors ZKM Apoptosis and Pathogen Removal
3.8. Cyt c Induces Pro-Apoptotic Caspase-1/IL-1β/Caspase-3 Axis in A. hydrophila-Infected ZKM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging Aeromonas species infections and their significance in public health. Sci. World J. 2012, 2012, 625023. [Google Scholar] [CrossRef] [PubMed]
- Batra, P.; Mathur, P.; Misra, M.C. Aeromonas spp.: An emerging nosocomial pathogen. J. Lab. Physicians 2016, 8, 001–004. [Google Scholar] [CrossRef] [PubMed]
- Chopra, A.; Xu, X.J.; Ribardo, D.; Gonzalez, M.; Kuhl, K.; Peterson, J.; Houston, C.W. The cytotoxic enterotoxin of Aeromonas hydrophila induces proinflammatory cytokine production and activates arachidonic acid metabolism in macrophages. Infect. Immun. 2000, 68, 2808–2818. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Ding, F.; Cui, P.; Ao, J.; Hu, S.; Chen, X.J. Transcriptome and expression profiling analysis revealed changes of multiple signaling pathways involved in immunity in the large yellow croaker during Aeromonas hydrophila infection. BMC Genom. 2010, 11, 1–14. [Google Scholar] [CrossRef]
- Srivastava, N.; Shelly, A.; Kumar, M.; Pant, A.; Das, B.; Majumdar, T.; Mazumder, S. Aeromonas hydrophila utilizes TLR4 topology for synchronous activation of MyD88 and TRIF to orchestrate anti-inflammatory responses in zebrafish. Cell Death Discov. 2017, 3, 1–9. [Google Scholar] [CrossRef]
- Kumar, M.; Kumar, J.; Sharma, S.; Hussain, M.A.; Shelly, A.; Das, B.; Yadav, A.K.; Mazumder, S. TLR22-mediated activation of TNF-α-caspase-1/IL-1β inflammatory axis leads to apoptosis of Aeromonas hydrophila-infected macrophages. Mol. Immunol. 2021, 137, 114–123. [Google Scholar] [CrossRef]
- Cheesman, S.E.; Neal, J.T.; Mittge, E.; Seredick, B.M.; Guillemin, K. Epithelial cell proliferation in the developing zebrafish intestine is regulated by the Wnt pathway and microbial signaling via Myd88. Proc. Natl. Acad. Sci. USA 2011, 108, 4570–4577. [Google Scholar] [CrossRef]
- Lü, A.J.; Hu, X.C.; Wang, Y.; Zhu, A.H.; Shen, L.L.; Tian, J.; Feng, Z.Z.; Feng, Z.J. Skin immune response in the zebrafish, Danio rerio (Hamilton), to Aeromonas hydrophila infection: A transcriptional profiling approach. J. Fish Dis. 2015, 38, 137–150. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Cao, M.; Wang, X.; Wang, G.; Li, J. Identification of wnt2 in the pearl mussel Hyriopsis cumingii and its role in innate immunity and gonadal development. Fish Shellfish Immunol. 2021, 118, 85–93. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/β-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, J.K.; Kling, J.C.; Tran, T.T.; Blumenthal, A. Functions of the WNT signaling network in shaping host responses to infection. Front. Immunol. 2019, 10, 2521. [Google Scholar] [CrossRef]
- Mukherjee, T.; Balaji, K.N. The WNT framework in shaping immune cell responses during bacterial infections. Front. Immunol. 2019, 10, 1985. [Google Scholar] [CrossRef]
- Rogan, M.R.; Patterson, L.L.; Wang, J.Y.; McBride, J.W. Bacterial manipulation of Wnt signaling: A host-pathogen tug-of-Wnt. Front. Immunol. 2019, 10, 2390. [Google Scholar] [CrossRef] [PubMed]
- Vallee, A.; Lecarpentier, Y.; Vallée, J. Interplay of opposing effects of the WNT/β-Catenin pathway and PPARγ and implications for SARS-CoV2 treatment. Front. Immunol. 2021, 12, 666693. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Zhong, G. The Chlamydia pneumoniae inclusion membrane protein Cpn1027 interacts with host cell Wnt signaling pathway regulator cytoplasmic activation/proliferation-associated protein 2 (Caprin2). PLoS ONE 2015, 10, e0127909. [Google Scholar] [CrossRef]
- Majumdar, T.; Sharma, S.; Kumar, M.; Hussain, M.; Chauhan, N.; Kalia, I.; Sahu, A.K.; Rana, V.S.; Bharti, R.; Haldar, A.K.; et al. Tryptophan-kynurenine pathway attenuates β-catenin-dependent pro-parasitic role of STING-TICAM2-IRF3-IDO1 signalosome in Toxoplasma gondii infection. Cell Death Dis. 2019, 10, 1–19. [Google Scholar] [CrossRef]
- Cott, C.; Thuenauer, R.; Landi, A.; Kühn, K.; Juillot, S.; Imberty, A.; Madl, J.; Eierhoff, T.; Römer, W. Pseudomonas aeruginosa lectin LecB inhibits tissue repair processes by triggering β-catenin degradation. Biochim. Biophys. Acta 2016, 1863, 1106–1118. [Google Scholar] [CrossRef]
- Tao, L.; Zhang, J.; Meraner, P.; Tovaglieri, A.; Wu, X.; Gerhard, R.; Zhang, X.; Stallcup, W.B.; Miao, J.; He, X.; et al. Frizzled proteins are colonic epithelial receptors for C. difficile toxin B. Nature 2016, 538, 350–355. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Pan, Y.; Zhang, C.; Yan, B.X.; Yu, S.S.; Wu, D.L.; Shi, M.M.; Shi, K.; Cai, X.X.; Zhou, S.S. Wnt/β-catenin signaling induces the aging of mesenchymal stem cells through promoting the ROS production. Mol. Cell. Biochem. 2013, 374, 13–20. [Google Scholar] [CrossRef]
- Staehlke, S.; Haack, F.; Waldner, A.-C.; Koczan, D.; Moerke, C.; Mueller, P.; Uhrmacher, A.M.; Nebe, J.B. ROS Dependent Wnt/β-Catenin Pathway and Its Regulation on Defined Micro-Pillars—A Combined In Vitro and In Silico Study. Cells 2020, 9, 1784. [Google Scholar] [CrossRef] [PubMed]
- Goswamy, D.; Irazoqui, J.E. A unifying hypothesis on the central role of reactive oxygen species in bacterial pathogenesis and host defense in C. elegans. Curr. Opin. Immunol. 2021, 68, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid. Med. Cell Longev. 2019, 2019, 1–13. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: Outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Takeshima, Y.; Fujio, K. Basic mechanism of immune system activation by mitochondria. Immunol. Med. 2020, 43, 142–147. [Google Scholar] [CrossRef]
- Silwal, P.; Kim, J.K.; Kim, Y.J.; Jo, E.K. Mitochondrial reactive oxygen species: Double-edged weapon in host defense and pathological inflammation during infection. Front. Immunol. 2020, 11, 1649. [Google Scholar] [CrossRef] [PubMed]
- Shekhova, E. Mitochondrial reactive oxygen species as major effectors of antimicrobial immunity. PLoS Pathog. 2020, 16, e1008470. [Google Scholar] [CrossRef]
- Herb, M.; Schramm, M. Functions of ROS in macrophages and antimicrobial immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef]
- Shafique, E.; Torina, A.; Reichert, K.; Colantuono, B.; Nur, N.; Zeeshan, K.; Ravichandran, V.; Liu, Y.; Feng, J.; Zeeshan, K.; et al. Mitochondrial redox plays a critical role in the paradoxical effects of NAPDH oxidase-derived ROS on coronary endothelium. Cardiovasc. Res. 2017, 113, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.C.; Ng, A.; Kim, B.H.; Bianco, A.; Xavier, R.J.; Elledge, S. J Wnt signaling regulates mitochondrial physiology and insulin sensitivity. Genes Dev. 2010, 24, 1507–1518. [Google Scholar] [CrossRef]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Aparicio-Trejo, O.E.; Pedraza-Chaverri, J. Mitochondrial redox signaling and oxidative stress in kidney diseases. Biomolecules 2021, 11, 1144. [Google Scholar] [CrossRef]
- Poirault-Chassac, S.; Nivet-Antoine, V.; Houvert, A.; Kauskot, A.; Lauret, E.; Lai-Kuen, R.; Dusanter-Fourt, I.; Baruch, D. Mitochondrial dynamics and reactive oxygen species initiate thrombopoiesis from mature megakaryocytes. Blood Adv. 2021, 5, 1706–1718. [Google Scholar] [CrossRef]
- Zhang, L.; Li, S.; Wang, R.; Chen, C.; Ma, W.; Cai, H. Anti-tumor effect of LATS2 on liver cancer death: Role of DRP1-mediated mitochondrial division and the Wnt/β-catenin pathway. Biomed. Pharmacother. 2019, 114, 108825. [Google Scholar] [CrossRef]
- Banerjee, C.; Singh, A.; Das, T.K.; Raman, R.; Shrivastava, A.; Mazumder, S. Ameliorating ER-stress attenuates Aeromonas hydrophila-induced mitochondrial dysfunctioning and caspase mediated HKM apoptosis in Clarias batrachus. Sci. Rep. 2014, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Shelly, A.; Dahiya, P.; Ray, A.; Mazumder, S. Aeromonas hydrophila inhibits autophagy triggering cytosolic translocation of mtDNA which activates the pro-apoptotic caspase-1/IL-1β-nitric oxide axis in headkidney macrophages. Virulence 2022, 13, 60–76. [Google Scholar] [CrossRef]
- Chen, S.; Guttridge, D.C.; You, Z.; Zhang, Z.; Fribley, A.; Mayo, M.W.; Kitajewski, J.; Wang, C.Y. Wnt-1 signaling inhibits apoptosis by activating β-catenin/T cell factor–mediated transcription. J. Cell Biol. 2001, 152, 87–96. [Google Scholar] [CrossRef]
- You, Z.; Saims, D.; Chen, S.; Zhang, Z.; Guttridge, D.C.; Guan, K.L.; MacDougald, O.A.; Brown, A.M.; Evan, G.; Kitajewski, J.; et al. Wnt signaling promotes oncogenic transformation by inhibiting c-Myc–induced apoptosis. J. Cell Biol. 2002, 157, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Rosowski, E.E. Illuminating macrophage contributions to host-pathogen interactions in vivo: The power of Zebrafish. Infect. Immun. 2020, 88, e00906-19. [Google Scholar] [CrossRef]
- Sharma, S.; Kumar, M.; Kumar, J.; Srivastava, N.; Hussain, M.A.; Shelly, A.; Mazumder, S.M. fortuitum-induced CNS-pathology: Deciphering the role of canonical Wnt signaling, blood brain barrier components and cytokines. Dev. Comp. Immunol. 2021, 122, 104111. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Datta, D.; Singh, R.; Kumar, M.; Kumar, J.; Mazumder, S. TLR-2 mediated cytosolic-Ca2+ surge activates ER-stress-superoxide-NO signalosome augmenting TNF-α production leading to apoptosis of Mycobacterium smegmatis-infected fish macrophages. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Barbu, E.M.; Shirazi, F.; McGrath, D.M.; Albert, N.; Sidman, R.L.; Pasqualini, R.; Arap, W.; Kontoyiannis, D.P. An antimicrobial peptidomimetic induces Mucorales cell death through mitochondria-mediated apoptosis. PLoS ONE 2013, 8, e76981. [Google Scholar] [CrossRef]
- Krzymińska, S.; Kaznowski, A.; Chodysz, M. Aeromonas spp. human isolates induce apoptosis of murine macrophages. Curr. Microbiol. 2009, 58, 252–257. [Google Scholar] [CrossRef]
- Doonan, F.; Cotter, T.G. Morphological assessment of apoptosis. Methods 2008, 44, 200–204. [Google Scholar] [CrossRef]
- Schutte, B.; Nuydens, R.; Geerts, H.; Ramaekers, F. Annexin V binding assay as a tool to measure apoptosis in differentiated neuronal cells. J. Neurosci. Methods 1998, 86, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Xu, M. DNA fragmentation in apoptosis. Cell Res. 2000, 10, 205–211. [Google Scholar] [CrossRef]
- Ming, M.; Wang, S.; Wu, W.; Senyuk, V.; Le Beau, M.M.; Nucifora, G.; Qian, Z. Activation of Wnt/β-catenin protein signaling induces mitochondria-mediated apoptosis in hematopoietic progenitor cells. J. Biol. Chem. 2012, 287, 22683–22690. [Google Scholar] [CrossRef]
- Nusse, R. Wnt signaling in disease and in development. Cell Res. 2005, 15, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Galimov, E.R. The role of p66shc in oxidative stress and apoptosis. Acta Nat. 2010, 2, 44–51. [Google Scholar] [CrossRef]
- Kuznetsov, A.V.; Javadov, S.; Saks, V.; Margreiter, R.; Grimm, M. Synchronism in mitochondrial ROS flashes, membrane depolarization and calcium sparks in human carcinoma cells. Biochim. Biophys. Acta. Bioenerg. 2017, 1858, 418–431. [Google Scholar] [CrossRef]
- Ježek, J.; Cooper, K.F.; Strich, R. Reactive oxygen species and mitochondrial dynamics: The yin and yang of mitochondrial dysfunction and cancer progression. Antioxidants 2018, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Perfettini, J.L.; Roumier, T.; Kroemer, G. Mitochondrial fusion and fission in the control of apoptosis. Trends Cell Biol. 2005, 15, 179–183. [Google Scholar] [CrossRef]
- Prouty, M.G.; Correa, N.E.; Barker, L.P.; Jagadeeswaran, P.; Klose, K.E. Zebrafish-Mycobacterium marinum model for mycobacterial pathogenesis. FEMS Microbiol. 2003, 225, 177–182. [Google Scholar] [CrossRef]
- Majumdar, T.; Chattopadhyay, P.; Saha, D.R.; Sau, S.; Mazumder, S. Virulence plasmid of Aeromonas hydrophila induces macrophage apoptosis and helps in developing systemic infection in mice. Microb. Pathog. 2009, 46, 98–107. [Google Scholar] [CrossRef]
- Ashida, H.; Mimuro, H.; Ogawa, M.; Kobayashi, T.; Sanada, T.; Kim, M.; Sasakawa, C. Cell death and infection: A double-edged sword for host and pathogen survival. J. Cell Biol. 2011, 195, 931–942. [Google Scholar] [CrossRef]
- McCoy, A.J.; Koizumi, Y.; Toma, C.; Higa, N.; Dixit, V.; Taniguchi, S.I.; Tschopp, J.; Suzuki, T. Cytotoxins of the human pathogen Aeromonas hydrophila trigger, via the NLRP3 inflammasome, caspase-1 activation in macrophages. Eur. J. Immunol. 2010, 40, 2797–2803. [Google Scholar] [CrossRef]
- Franco, A.T.; Israel, D.A.; Washington, M.K.; Krishna, U.; Fox, J.G.; Rogers, A.B.; Neish, A.S.; Collier-Hyams, L.; Perez-Perez, G.I.; Hatakeyama, M.; et al. Activation of β-catenin by carcinogenic Helicobacter pylori. Proc. Natl. Acad. Sci. USA 2005, 102, 10646–10651. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, L.E.; Piazuelo, M.B.; Chaturvedi, R.; Schumacher, M.; Aihara, E.; Feng, R.; Noto, J.M.; Delgado, A.; Israel, D.A.; Zavros, Y.; et al. Helicobacter pyloritargets cancer-associated apical-junctional constituents in gastroids and gastric epithelial cells. Gut 2015, 64, 720–730. [Google Scholar] [CrossRef]
- Wu, X.; Deng, G.; Hao, X.; Li, Y.; Zeng, J.; Ma, C.; He, Y.; Liu, X.; Wang, Y. A caspase-dependent pathway is involved in Wnt/β-catenin signaling promoted apoptosis in Bacillus Calmette-Guerin infected RAW264. 7 macrophages. Int. J. Mol. Sci. 2014, 15, 5045–5062. [Google Scholar] [CrossRef] [PubMed]
- Canton, M.; Sánchez-Rodríguez, R.; Spera, I.; Venegas, F.C.; Favia, M.; Viola, A.; Castegna, A. Reactive Oxygen Species in Macrophages: Sources and Targets. Front. Immunol. 2021, 12, 734229. [Google Scholar] [CrossRef]
- Boo, J.H.; Song, H.; Kim, J.E.; Kang, D.E.; Mook-Jung, I. Accumulation of phosphorylated β-catenin enhances ROS-induced cell death in presenilin-deficient cells. PLoS ONE 2009, 4, e4172. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A.; Kim, Y.R.; Kumar, S.; Naqvi, A.; Hoffman, T.A.; Kumar, A.; Miller, F.J., Jr.; Kim, C.S.; Irani, K. Canonical Wnt signaling induces vascular endothelial dysfunction via p66Shc-regulated reactive oxygen species. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2301–2309. [Google Scholar] [CrossRef]
- Kumar, M.; Sharma, S.; Haque, M.; Kumar, J.; Hathi, U.P.S.; Mazumder, S. TLR22-Induced Pro-Apoptotic mtROS Abets UPRmt-Mediated Mitochondrial Fission in Aeromonas hydrophila-Infected Headkidney Macrophages of Clarias gariepinus. Front. Immunol. 2022, 13, 931021. [Google Scholar] [CrossRef]
- Roca, F.J.; Whitworth, L.J.; Redmond, S.; Jones, A.A.; Ramakrishnan, L. TNF induces pathogenic programmed macrophage necrosis in tuberculosis through a mitochondrial-lysosomal-endoplasmic reticulum circuit. Cell 2019, 178, 1344–1361. [Google Scholar] [CrossRef] [PubMed]
- Hatinguais, R.; Pradhan, A.; Brown, G.D.; Brown, A.J.; Warris, A.; Shekhova, E. Mitochondrial reactive oxygen species regulate immune responses of macrophages to Aspergillus fumigatus. Front. Immunol. 2021, 12, 929. [Google Scholar] [CrossRef]
- Rharass, T.; Lemcke, H.; Lantow, M.; Kuznetsov, S.A.; Weiss, D.G.; Panáková, D. Ca2+-mediated mitochondrial reactive oxygen species metabolism augments Wnt/β-catenin pathway activation to facilitate cell differentiation. J. Biol. Chem. 2014, 289, 27937–27951. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, S.; Ji, Z.; Xu, H.; Zhao, W.; Xia, Z.; Xu, R. Mechanistic study of mtROS-JNK-SOD2 signaling in bupivacaine-induced neuron oxidative stress. Aging 2020, 12, 13463. [Google Scholar] [CrossRef] [PubMed]
- Peterman, E.; Sullivan, C.; Goody, M.; Rodriguez-Nunez, I.; Yoder, J.; Kim, C.H. Neutralization of mitochondrial superoxide by superoxide dismutase 2 promotes bacterial clearance and regulates phagocyte numbers in zebrafish. Infect. Immun. 2015, 83, 430–440. [Google Scholar] [CrossRef]
- Jain, P.; Luo, Z.Q.; Blanke, S.R. Helicobacter pylori vacuolating cytotoxin A (VacA) engages the mitochondrial fission machinery to induce host cell death. Proc. Natl. Acad. Sci. USA 2011, 108, 16032–16037. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, M.; Pyakurel, A.; Glytsou, C.; Scorrano, L. The cell biology of mitochondrial membrane dynamics. Nat. Rev. Mol. Cell Biol. 2020, 21, 204–224. [Google Scholar] [CrossRef]
- Suen, D.F.; Norris, K.L.; Youle, R.J. Mitochondrial dynamics and apoptosis. Genes Dev. 2008, 22, 1577–1590. [Google Scholar] [CrossRef]
- Estaquier, J.; Arnoult, D. Inhibiting Drp1-mediated mitochondrial fission selectively prevents the release of cytochrome c during apoptosis. Cell Death Differ. 2007, 14, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Donath, S.; Li, Y.; Qin, D.; Prabhakar, B.S.; Li, P. miR-30 regulates mitochondrial fission through targeting p53 and the dynamin-related protein-1 pathway. PLoS Genet. 2010, 6, e1000795. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Chemicals Used | Company | Function |
|---|---|---|---|
| 1. | JW67 | Sigma | β-catenin inhibitor |
| 2. | MSAB | MedChemExpress | β-catenin inhibitor |
| 3. | LiCl | Sigma | Wnt/β-catenin signaling activator |
| 4. | Laduviglusib | MedChemExpress | Wnt/β-catenin signaling activator |
| 5. | Apo | Sigma | NADPH oxidase (NOX) inhibitor |
| 6. | DPI | Sigma | NADPH oxidase (NOX) inhibitor |
| 7. | Mdivi-1 | Sigma | Drp1 inhibitor |
| 8. | CsA | Sigma | Mitochondrial permeability transition pore (MPTP) inhibitor |
| 9. | YCG063 | Calbiochem | Mitochondrial ROS (mtROS) inhibitor |
| 10. | Ant A | Sigma | Mitochondrial ROS (mtROS) inducer |
| 11. | Z-YVAD-FMK | Sigma | Caspase-1 inhibitor |
| 12. | Ac-DEVD-CHO | Biovision | Caspase-3 inhibitor |
| S. No. | Gene Name | Accession Number | Primer Sequence | Amplicon Length (bp) |
|---|---|---|---|---|
| 1. | actb1 | FJ915059.1 | FP:5′-CGAGCAGGAGATGGGAACC-3′ RP: 5′-CAACGGAAACGCTCATTGC-3′ | 104 |
| 2. | wnt2 | NM_130950.1 | FP:5′-CACAATCTGTTCGGGAGGCT-3′ RP: 5′-ACGCCCTGGTCAATGTGTAG-3′ | 100 |
| 3. | wnt3a | NM_001007185.1 | FP:5′-TAAGCAAGCAAAGGCCACCAG-3′ RP: 5′-GACACCATGCTGCCGAACTC-3′ | 198 |
| 4. | lrp6 | NM_001134684.1 | FP:5′-GTCAACACACCGCTCCTACA-3′ RP:5′-CCCGGCGTATAGTCACTGTC-3′ | 105 |
| 5. | fzd5 | NM_131134.2 | FP:5′-CCTTGCCACCAACCCTACTT-3′ RP: 5′-CGCTCCATGTCGATGAGGAA-3′ | 128 |
| 6. | gsk3b | NM_131381.1 | FP:5′-CATCTTTGGAGCCACCGACT-3′ RP: 5′-TGGCCGAAACACCTTAGTCC-3′ | 240 |
| 7. | axin | NM_131503.2 | FP: 5′-TCTGGCCAATCACAGGGTTC-3′ RP: 5′-TCGTGTGCATCCCTTAGCTG-3′ | 102 |
| 8. | ctnnb1 | NM_131059.2 | FP:5′-GCTCCCCACAGATGGTATCG-3′ RP: 5′-GGAGCCGAGCATATTGACGA-3′ | 170 |
| 9. | shc2 | NM_001044973.1 | FP: 5′-TCGGGCTCAAACTTCACATCT-3′ RP: 5′-GGCCGAATCTACTCCCCTCT-3′ | 130 |
| 10. | sod2 | NM_199976.1 | FP: 5′-TAGGTCTGTTGGTTGGTCGC-3′ RP: 5′-GCACCTAACAGGGGGTTGAA-3′ | 109 |
| 11. | dnm1l | NM_200922.1 | FP: 5′-GCAGAGTAGCGGGAAGAGTT-3′ RP: 5′-TCCATCCACTCCGTTCTCCT-3′ | 160 |
| 12. | mfn1 | BC057468.1 | FP: 5′-AACTGATGTGACCACCGAGC-3′ RP: 5′-CGTCCCAGCGATTGTTCAAG-3′ | 179 |
| 13. | mfn2 | NM_001128254.2 | FP: 5′-AGACAGTGTTTCGCCCTCAG-3′ RP: 5′-CCTGGTCGTTGTAGCCCATT-3′ | 163 |
| S. No. | Gene | siRNA Sequence |
|---|---|---|
| 1. | il1b | Sense: 5′-GGAAAGACACCGAGCGCAUUU-3′ Antisense: 5′-PAUGCGCUCGGUGUCUUUCCUU-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, S.; Kumar, M.; Kumar, J.; Mazumder, S. β-Catenin Elicits Drp1-Mediated Mitochondrial Fission Activating the Pro-Apoptotic Caspase-1/IL-1β Signalosome in Aeromonas hydrophila-Infected Zebrafish Macrophages. Cells 2023, 12, 1509. https://doi.org/10.3390/cells12111509
Sharma S, Kumar M, Kumar J, Mazumder S. β-Catenin Elicits Drp1-Mediated Mitochondrial Fission Activating the Pro-Apoptotic Caspase-1/IL-1β Signalosome in Aeromonas hydrophila-Infected Zebrafish Macrophages. Cells. 2023; 12(11):1509. https://doi.org/10.3390/cells12111509
Chicago/Turabian StyleSharma, Shagun, Manmohan Kumar, Jai Kumar, and Shibnath Mazumder. 2023. "β-Catenin Elicits Drp1-Mediated Mitochondrial Fission Activating the Pro-Apoptotic Caspase-1/IL-1β Signalosome in Aeromonas hydrophila-Infected Zebrafish Macrophages" Cells 12, no. 11: 1509. https://doi.org/10.3390/cells12111509
APA StyleSharma, S., Kumar, M., Kumar, J., & Mazumder, S. (2023). β-Catenin Elicits Drp1-Mediated Mitochondrial Fission Activating the Pro-Apoptotic Caspase-1/IL-1β Signalosome in Aeromonas hydrophila-Infected Zebrafish Macrophages. Cells, 12(11), 1509. https://doi.org/10.3390/cells12111509