
Functional IgG Autoantibodies against Autonomic Nervous System Receptors in Symptomatic Women with Silicone Breast Implants

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. IRB Approval/Ethical Clearance
2.3. IgG Purification from Human Subjects
2.4. Human Peripheral Blood Mononuclear Cell Isolation and In Vitro Study
2.5. Quantification of Cytokine Levels
2.6. Mice and Behavioral Study Design
2.7. Open Field Test (OFT)
2.8. Novel-Object Location (NOL)
2.9. Forced Swim Test (FST)
2.10. Quantification of Circulating Auto-Antibody Levels
2.11. Statistical Analysis
3. Results
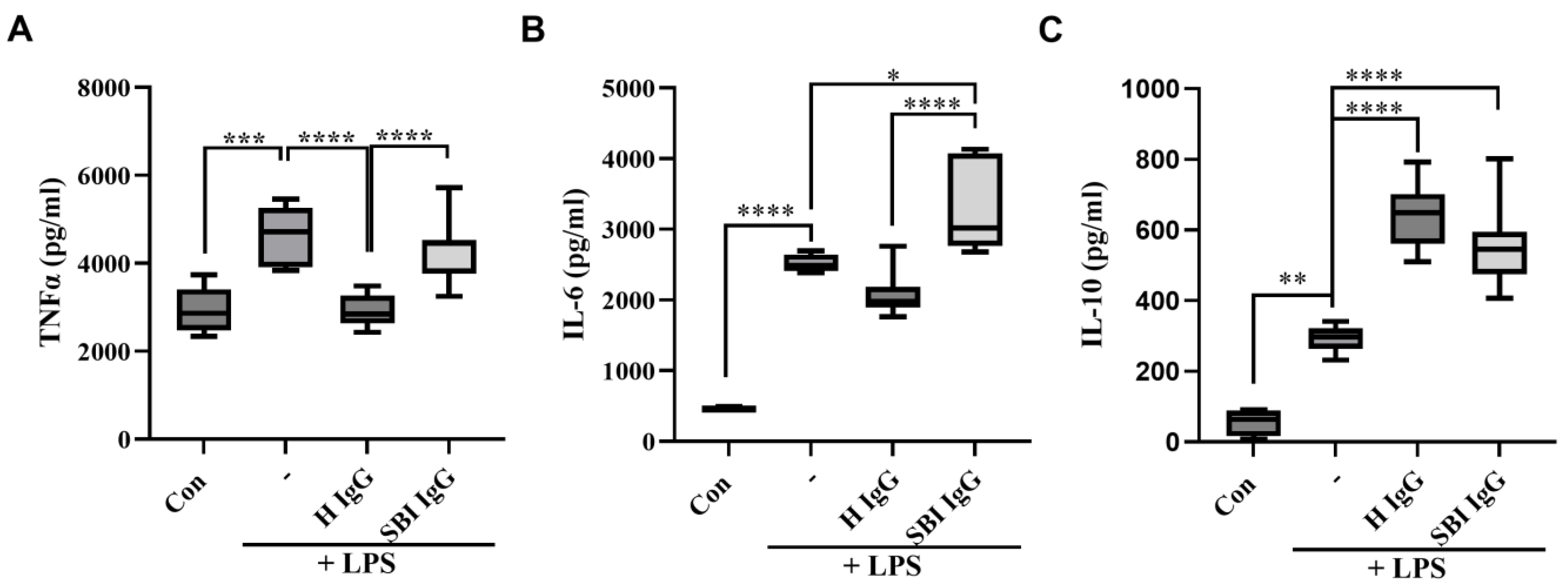
3.1. IgG Derived from Symptomatic Women with Silicone Breast Implants Dysregulates LPS-Induced Cytokine Secretion in Activated Human PBMCs
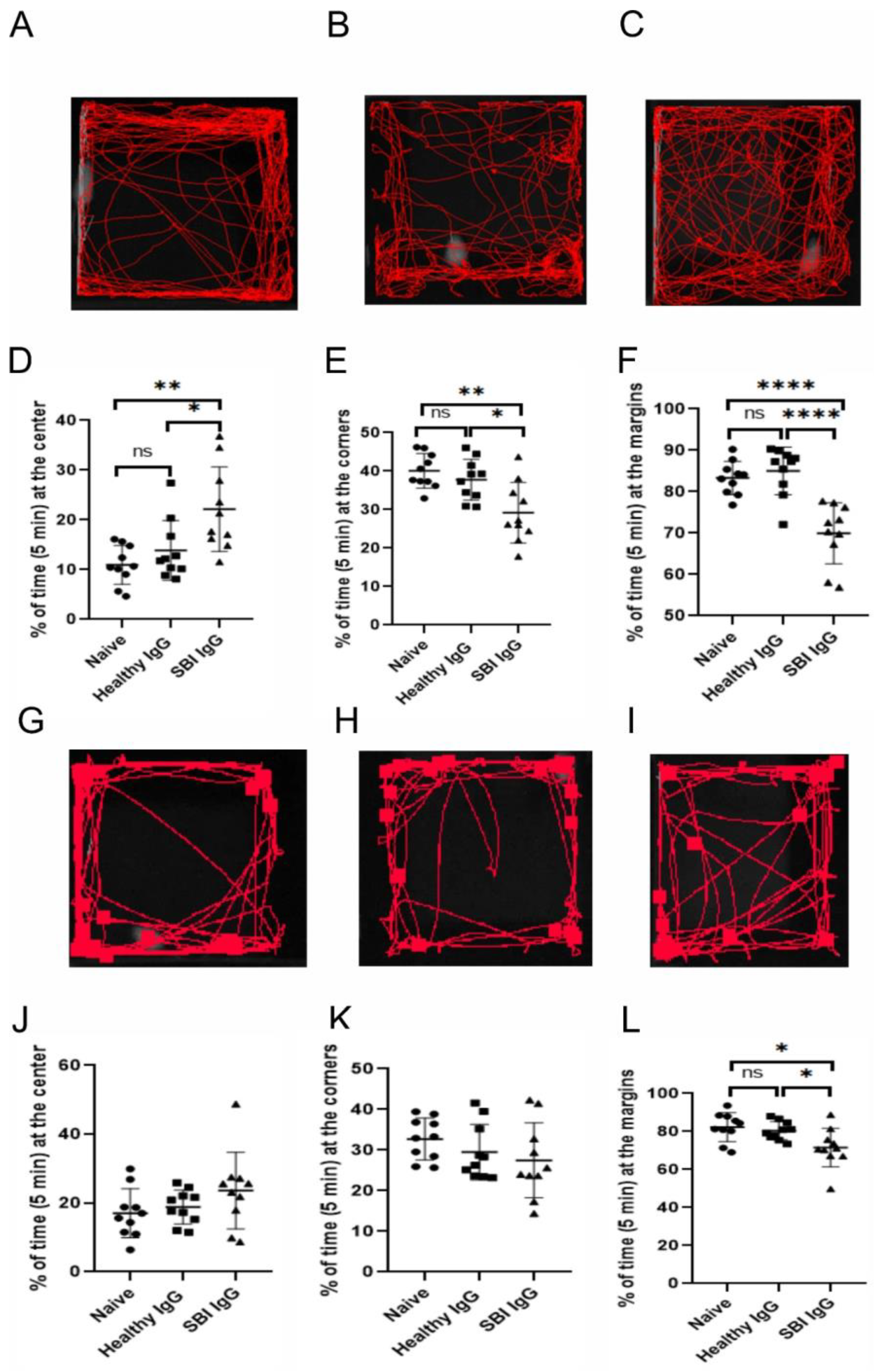
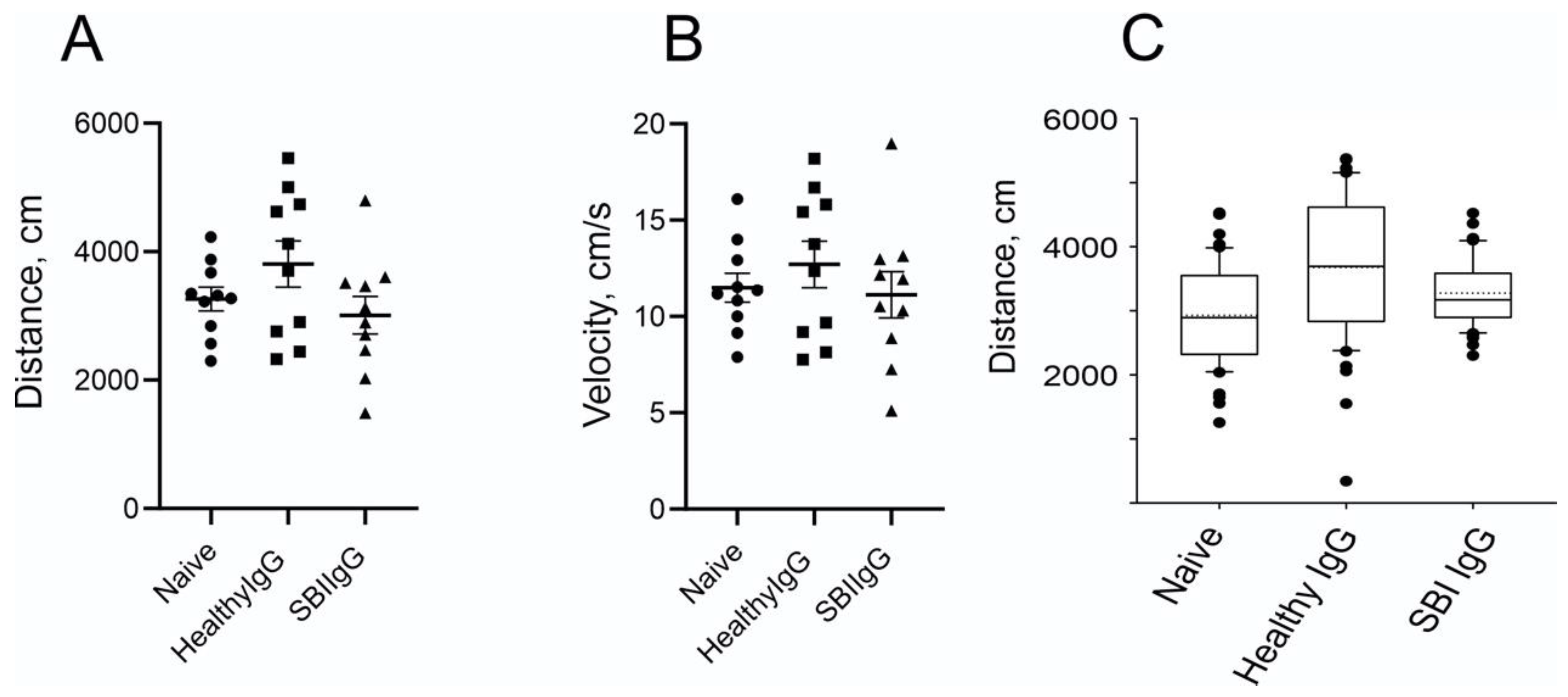
3.2. Passive Transfer of IgGs Derived from Symptomatic Women with SBI, into Mouse Brain, Affects Anxiety-like Behavior and Locomotor Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bridges, A.J.; Vasey, F.B. Silicone Breast Implants. History, Safety, and Potential Complications. Arch. Intern. Med. 1993, 153, 2638–2644. [Google Scholar] [CrossRef]
- Kumagai, Y.; Abe, C.; Shiokawa, Y. Scleroderma after Cosmetic Surgery: Four Cases of Human Adjuvant Disease. Arthritis Rheum. 1979, 22, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, Y.; Shiokawa, Y.; Medsger, T.A.; Rodnan, G.P. Clinical Spectrum of Connective Tissue Disease after Cosmetic Surgery. Observations on Eighteen Patients and a Review of the Japanese Literature. Arthritis Rheum. 1984, 27, 1–12. [Google Scholar] [CrossRef]
- Peters, W.; Smith, D.; Fornasier, V.; Lugowski, S.; Ibanez, D. An Outcome Analysis of 100 Women after Explantation of Silicone Gel Breast Implants. Ann. Plast. Surg. 1997, 39, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Aharon-Maor, A.; Levy, Y.; Schoenfeld, Y. Fibrosarcoma after silicone breast augmentation: Is there a connection? Harefuah 1998, 134, 339–341, 424. [Google Scholar] [PubMed]
- Yoshida, S.H.; Swan, S.; Teuber, S.S.; Gershwin, M.E. Silicone Breast Implants: Immunotoxic and Epidemiologic Issues. Life Sci. 1995, 56, 1299–1310. [Google Scholar] [CrossRef]
- Watad, A.; Bragazzi, N.L.; Amital, H.; Shoenfeld, Y. Hyperstimulation of Adaptive Immunity as the Common Pathway for Silicone Breast Implants, Autoimmunity, and Lymphoma of the Breast. Isr. Med. Assoc. J. 2019, 21, 517–519. [Google Scholar]
- Zandman-Goddard, G.; Blank, M.; Ehrenfeld, M.; Gilburd, B.; Peter, J.; Shoenfeld, Y. A Comparison of Autoantibody Production in Asymptomatic and Symptomatic Women with Silicone Breast Implants. J. Rheumatol. 1999, 26, 73–77. [Google Scholar]
- Watad, A.; Rosenberg, V.; Tiosano, S.; Cohen Tervaert, J.W.; Yavne, Y.; Shoenfeld, Y.; Shalev, V.; Chodick, G.; Amital, H. Silicone Breast Implants and the Risk of Autoimmune/Rheumatic Disorders: A Real-World Analysis. Int. J. Epidemiol. 2018, 47, 1846–1854. [Google Scholar] [CrossRef]
- Tanne, J.H. Breast Implants: US Regulator Issues “Black Box” Warning and Requires More Patient Information. BMJ 2021, n2639. [Google Scholar] [CrossRef]
- Shoenfeld, Y.; Ryabkova, V.A.; Scheibenbogen, C.; Brinth, L.; Martinez-Lavin, M.; Ikeda, S.; Heidecke, H.; Watad, A.; Bragazzi, N.L.; Chapman, J.; et al. Complex Syndromes of Chronic Pain, Fatigue and Cognitive Impairment Linked to Autoimmune Dysautonomia and Small Fiber Neuropathy. Clin. Immunol. 2020, 214, 108384. [Google Scholar] [CrossRef] [PubMed]
- Halpert, G.; Watad, A.; Tsur, A.M.; Dotan, A.; Quiros-Lim, H.E.; Heidecke, H.; Gilburd, B.; Haik, J.; Levy, Y.; Blank, M.; et al. Autoimmune Dysautonomia in Women with Silicone Breast Implants. J. Autoimmun. 2021, 120, 102631. [Google Scholar] [CrossRef] [PubMed]
- Malysheva, O.; Pierer, M.; Wagner, U.; Wahle, M.; Wagner, U.; Baerwald, C.G. Association between Beta2 Adrenergic Receptor Polymorphisms and Rheumatoid Arthritis in Conjunction with Human Leukocyte Antigen (HLA)-DRB1 Shared Epitope. Ann. Rheum. Dis. 2008, 67, 1759–1764. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Haberberger, R.V.; Gordon, T.P.; Jackson, M.W. Antibodies Interfering with the Type 3 Muscarinic Receptor Pathway Inhibit Gastrointestinal Motility and Cholinergic Neurotransmission in Sjögren’s Syndrome. Arthritis Rheum. 2011, 63, 1426–1434. [Google Scholar] [CrossRef]
- Eng, H.; Magnusson, Y.; Matell, G.; Lefvert, A.K.; Saponja, R.; Hoebeke, J. Beta 2-Adrenergic Receptor Antibodies in Myasthenia Gravis. J. Autoimmun. 1992, 5, 213–227. [Google Scholar] [CrossRef]
- Smith, A.J.; Jackson, M.W.; Wang, F.; Cavill, D.; Rischmueller, M.; Gordon, T.P. Neutralization of Muscarinic Receptor Autoantibodies by Intravenous Immunoglobulin in Sjögren Syndrome. Hum. Immunol. 2005, 66, 411–416. [Google Scholar] [CrossRef]
- Cabral-Marques, O.; Marques, A.; Giil, L.M.; De Vito, R.; Rademacher, J.; Günther, J.; Lange, T.; Humrich, J.Y.; Klapa, S.; Schinke, S.; et al. GPCR-Specific Autoantibody Signatures Are Associated with Physiological and Pathological Immune Homeostasis. Nat. Commun. 2018, 9, 5224. [Google Scholar] [CrossRef]
- Hansen, N.; Luedecke, D.; Malchow, B.; Lipp, M.; Vogelgsang, J.; Timäus, C.; Zindler, T.; Gingele, S.; Kühn, S.; Gallinat, J.; et al. + CAP (Cerebrospinal Fluid Analysis in Psychiatry)-Consortium. Autoantibody-Associated Psychiatric Syndromes in Children: Link to Adult Psychiatry. J. Neural. Transm. 2021, 128, 735–747. [Google Scholar] [CrossRef]
- Aldinger, K.A.; Sokoloff, G.; Rosenberg, D.M.; Palmer, A.A.; Millen, K.J. Genetic Variation and Population Substructure in Outbred CD-1 Mice: Implications for Genome-Wide Association Studies. PLoS ONE 2009, 4, e4729. [Google Scholar] [CrossRef]
- Frasier, L.; Kehrer, J.P. Effect of Indomethacin, Aspirin, Nordihydroguairetic Acid, and Piperonyl Butoxide on Cyclophosphamide-Induced Bladder Damage. Drug Chem. Toxicol. 1993, 16, 117–133. [Google Scholar] [CrossRef]
- Salloum, F.; Yin, C.; Xi, L.; Kukreja, R.C. Sildenafil Induces Delayed Preconditioning through Inducible Nitric Oxide Synthase-Dependent Pathway in Mouse Heart. Circ. Res. 2003, 92, 595–597. [Google Scholar] [CrossRef] [PubMed]
- Flammang, A.M.; Erexson, G.L.; Mecchi, M.S.; Murli, H. Genotoxicity Testing of a Salacia Oblonga Extract. Food Chem. Toxicol. 2006, 44, 1868–1874. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.J.; Droll, D.A.; Jiang, X.; Wold, W.S.M.; Nickells, J.A. JE Nakayama/JE SA14-14-2 Virus Structural Region Intertypic Viruses: Biological Properties in the Mouse Model of Neuroinvasive Disease. Virology 2007, 366, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Meini, S.; Cucchi, P.; Bellucci, F.; Catalani, C.; Giuliani, S.; Santicioli, P.; Maggi, C.A. Comparative Antagonist Pharmacology at the Native Mouse Bradykinin B2 Receptor: Radioligand Binding and Smooth Muscle Contractility Studies. Br. J. Pharmacol. 2007, 150, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Mott, K.R.; Osorio, Y.; Brown, D.J.; Morishige, N.; Wahlert, A.; Jester, J.V.; Ghiasi, H. The Corneas of Naive Mice Contain Both CD4+ and CD8+ T Cells. Mol. Vis. 2007, 13, 1802–1812. [Google Scholar]
- Yankelevitch-Yahav, R.; Franko, M.; Huly, A.; Doron, R. The Forced Swim Test as a Model of Depressive-like Behavior. J. Vis. Exp. 2015, 52587. [Google Scholar] [CrossRef]
- Karthivashan, G.; Park, S.-Y.; Kim, J.-S.; Cho, D.-Y.; Ganesan, P.; Choi, D.-K. Comparative Studies on Behavioral, Cognitive and Biomolecular Profiling of ICR, C57BL/6 and Its Sub-Strains Suitable for Scopolamine-Induced Amnesic Models. Int. J. Mol. Sci. 2017, 18, 1735. [Google Scholar] [CrossRef]
- Cetin, A.; Komai, S.; Eliava, M.; Seeburg, P.H.; Osten, P. Stereotaxic Gene Delivery in the Rodent Brain. Nat. Protoc. 2006, 1, 3166–3173. [Google Scholar] [CrossRef]
- Katzav, A.; Arango, M.T.; Kivity, S.; Tanaka, S.; Givaty, G.; Agmon-Levin, N.; Honda, M.; Anaya, J.-M.; Chapman, J.; Shoenfeld, Y. Passive Transfer of Narcolepsy: Anti-TRIB2 Autoantibody Positive Patient IgG Causes Hypothalamic Orexin Neuron Loss and Sleep Attacks in Mice. J. Autoimmun. 2013, 45, 24–30. [Google Scholar] [CrossRef]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The Tail Suspension Test as a Model for Assessing Antidepressant Activity: Review of Pharmacological and Genetic Studies in Mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef]
- Depino, A.M.; Gross, C. Simultaneous Assessment of Autonomic Function and Anxiety-Related Behavior in BALB/c and C57BL/6 Mice. Behav. Brain Res. 2007, 177, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to Measure Locomotor and Anxiety-like Behavior in Mice. J. Vis. Exp. 2015, e52434. [Google Scholar] [CrossRef]
- Weible, A.P.; Rowland, D.C.; Pang, R.; Kentros, C. Neural Correlates of Novel Object and Novel Location Recognition Behavior in the Mouse Anterior Cingulate Cortex. J. Neurophysiol. 2009, 102, 2055–2068. [Google Scholar] [CrossRef]
- Ennaceur, A.; Delacour, J. A New One-Trial Test for Neurobiological Studies of Memory in Rats. 1: Behavioral Data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Sanchez, A.; Dematagoda, S.; Ahmed, R.; Mohanathaas, S.; Odenwald, N.; Arruda-Carvalho, M. Developmental Onset Distinguishes Three Types of Spontaneous Recognition Memory in Mice. Sci. Rep. 2020, 10, 10612. [Google Scholar] [CrossRef]
- Katzav, A.; Ben-Ziv, T.; Blank, M.; Pick, C.G.; Shoenfeld, Y.; Chapman, J. Antibody-Specific Behavioral Effects: Intracerebroventricular Injection of Antiphospholipid Antibodies Induces Hyperactive Behavior While Anti-Ribosomal-P Antibodies Induces Depression and Smell Deficits in Mice. J. Neuroimmunol. 2014, 272, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D.; Bertin, A.; Jalfre, M. Behavioral Despair in Mice: A Primary Screening Test for Antidepressants. Arch. Int. Pharmacodyn. Ther. 1977, 229, 327–336. [Google Scholar]
- Murlanova, K.; Michaelevski, I.; Kreinin, A.; Terrillion, C.; Pletnikov, M.; Pinhasov, A. Link between Temperament Traits, Brain Neurochemistry and Response to SSRI: Insights from Animal Model of Social Behavior. J. Affect Disord. 2021, 282, 1055–1066. [Google Scholar] [CrossRef]
- Cabral-Marques, O.; Halpert, G.; Schimke, L.F.; Ostrinski, Y.; Vojdani, A.; Baiocchi, G.C.; Freire, P.P.; Filgueiras, I.S.; Zyskind, I.; Lattin, M.T.; et al. Autoantibodies Targeting GPCRs and RAS-Related Molecules Associate with COVID-19 Severity. Nat. Commun. 2022, 13, 1220. [Google Scholar] [CrossRef]
- Tocut, M.; Halpert, G.; Tsur, A.M.; Sharif, K.; Heidecke, H.; Levy, Y.; Watad, A.; Amital, H.; Shoenfeld, Y. Cognitive Impairment, Sleep Disturbance, and Depression in Women with Silicone Breast Implants: Association with Autoantibodies against Autonomic Nervous System Receptors. Biomolecules 2022, 12, 776. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The Meaning and Use of the Area under a Receiver Operating Characteristic (ROC) Curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, J.; Sotzny, F.; Bauer, S.; Heidecke, H.; Riemekasten, G.; Dragun, D.; Meisel, C.; Dames, C.; Grabowski, P.; Scheibenbogen, C. IgG Stimulated Β2 Adrenergic Receptor Activation Is Attenuated in Patients with ME/CFS. Brain Behav. Immun. Health 2020, 3, 100047. [Google Scholar] [CrossRef] [PubMed]
- Kozicky, L.K.; Menzies, S.C.; Zhao, Z.Y.; Vira, T.; Harnden, K.; Safari, K.; Del Bel, K.L.; Turvey, S.E.; Sly, L.M. IVIg and LPS Co-Stimulation Induces IL-10 Production by Human Monocytes, Which Is Compromised by an FcγRIIA Disease-Associated Gene Variant. Front. Immunol. 2018, 9, 2676. [Google Scholar] [CrossRef]
- Murakami, K.; Suzuki, C.; Kobayashi, F.; Nakano, A.; Fujii, A.; Sakai, K.; Imada, T. Intravenous Immunoglobulin Preparation Attenuates LPS-Induced Production of pro-Inflammatory Cytokines in Human Monocytic Cells by Modulating TLR4-Mediated Signaling Pathways. Naunyn. Schmiedebergs Arch. Pharmacol. 2012, 385, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Halpert, G.; Katz, I.; Shovman, O.; Tarasov, S.; Ganina, K.K.; Petrova, N.; Tocut, M.; Volkov, A.; Barshack, I.; Blank, M.; et al. IVIG Ameliorate Inflammation in Collagen-Induced Arthritis: Projection for IVIG Therapy in Rheumatoid Arthritis. Clin. Exp. Immunol. 2021, 203, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Iseme, R.A.; McEvoy, M.; Kelly, B.; Agnew, L.; Attia, J.; Walker, F.R. Autoantibodies and Depression: Evidence for a Causal Link? Neurosci. Biobehav. Rev. 2014, 40, 62–79. [Google Scholar] [CrossRef]
- Postal, M.; Appenzeller, S. The Importance of Cytokines and Autoantibodies in Depression. Autoimmun. Rev. 2015, 14, 30–35. [Google Scholar] [CrossRef]
- Roy, B.F.; Rose, J.W.; McFarland, H.F.; McFarlin, D.E.; Murphy, D.L. Anti-Beta-Endorphin Immunoglobulin G in Humans. Proc. Natl. Acad. Sci. USA 1986, 83, 8739–8743. [Google Scholar] [CrossRef]
- Tanaka, S.; Matsunaga, H.; Kimura, M.; Tatsumi, K.I.; Hidaka, Y.; Takano, T.; Uema, T.; Takeda, M.; Amino, N. Autoantibodies against Four Kinds of Neurotransmitter Receptors in Psychiatric Disorders. J. Neuroimmunol. 2003, 141, 155–164. [Google Scholar] [CrossRef]
- Katzav, A.; Solodeev, I.; Brodsky, O.; Chapman, J.; Pick, C.G.; Blank, M.; Zhang, W.; Reichlin, M.; Shoenfeld, Y. Induction of Autoimmune Depression in Mice by Anti-Ribosomal P Antibodies via the Limbic System. Arthritis Rheum. 2007, 56, 938–948. [Google Scholar] [CrossRef]
- Watson, P.; Storbeck, J.; Mattis, P.; Mackay, M. Cognitive and Emotional Abnormalities in Systemic Lupus Erythematosus: Evidence for Amygdala Dysfunction. Neuropsychol. Rev. 2012, 22, 252–270. [Google Scholar] [CrossRef] [PubMed]
- Lapteva, L.; Nowak, M.; Yarboro, C.H.; Takada, K.; Roebuck-Spencer, T.; Weickert, T.; Bleiberg, J.; Rosenstein, D.; Pao, M.; Patronas, N.; et al. Anti-N-Methyl-D-Aspartate Receptor Antibodies, Cognitive Dysfunction, and Depression in Systemic Lupus Erythematosus. Arthritis Rheum. 2006, 54, 2505–2514. [Google Scholar] [CrossRef] [PubMed]
- Goebel, A.; Krock, E.; Gentry, C.; Israel, M.R.; Jurczak, A.; Urbina, C.M.; Sandor, K.; Vastani, N.; Maurer, M.; Cuhadar, U.; et al. Passive Transfer of Fibromyalgia Symptoms from Patients to Mice. J. Clin. Investig. 2021, 131, 144201. [Google Scholar] [CrossRef] [PubMed]
- Scheibenbogen, C.; Loebel, M.; Freitag, H.; Krueger, A.; Bauer, S.; Antelmann, M.; Doehner, W.; Scherbakov, N.; Heidecke, H.; Reinke, P.; et al. Immunoadsorption to Remove SS2 Adrenergic Receptor Antibodies in Chronic Fatigue Syndrome CFS/ME. PLoS ONE 2018, 13, e0193672. [Google Scholar] [CrossRef]
- Gavrilova, N.; Kamaeva, E.; Ignatova, M.; Ryabkova, V.; Lukashenko, M.; Soprun, L.; Churilov, L.P.; Shoenfeld, Y. Intravenouse Immunoglobuline in Dysautonomia. Clin. Immunol. 2022, 240, 109039. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Characteristics | Women with SBI n = 10 | Healthy Women n = 10 |
|---|---|---|
| Mean age ± SEM (y) | 51.1 ± 3.9 | 39.5 ± 4.9 |
| Mean time from implantation to symptom onset ± SEM (y) | 9.3 ± 2.8 | |
| Cosmetic indication for SBI | 7 (70%) | |
| Subjective/Autonomic-related symptoms | ||
| Fatigue | 8 (80%) | |
| Widespread pain | 7 (70%) | |
| Depression | 4 (40%) | |
| Cognitive impairment | 5 (50%) | |
| Memory impairment | 6 (60%) | |
| Palpitations | 3 (30%) | |
| Dry mouth | 3 (30%) | |
| Sleep disturbances | 7 (70%) | |
| Medical History | ||
| Autoimmune thyroid disease | 4 (40%) | |
| Inflammatory bowel disease | 0 (0%) | |
| Endometriosis | 0 (0%) | |
| Familial history for autoimmune disease | 4 (40%) | |
| Drug Use | ||
| Selective serotonin reuptake inhibitors (Cipralex) | 2 (20%) | |
| Stimulants (Ritalin) | 0 (0%) | |
| TNF blockers (Humira) | 0 (0%) | |
| Immunosuppressants (Azathioprine) | 0 (0%) |
| Patient Characteristics | Women with SBI n = 10 | Healthy Women n = 10 | |
|---|---|---|---|
| Mean age ± SEM (y) | 37.7 ± 1.7 | 37.8 ± 4.1 | |
| Mean time from implantation to symptom onset ± SEM (y) | 10.6 ± 1.2 | ||
| Cosmetic indication for SBI | 10 (100%) | ||
| Subjective/Autonomic-related symptoms | |||
| Fatigue | 10 (100%) | ||
| Widespread pain | 10 (100%) | ||
| Depression | 10 (100%) | ||
| Cognitive impairment | 10 (100%) | ||
| Memory impairment | 10 (100%) | ||
| Palpitations | 9 (90%) | ||
| Dry mouth | 8 (80%) | ||
| Sleep disturbances | 8 (80%) | ||
| Medical History | |||
| Autoimmune thyroid disease | 1 (10%) | ||
| Inflammatory bowel disease | 3 (30%) | ||
| Endometriosis | 1 (10%) | ||
| Familial history for autoimmune disease | 5 (50%) | ||
| Drug Use | |||
| Selective serotonin reuptake inhibitors (Cipralex) | 1 (10%) | ||
| Stimulants (Ritalin) | 1 (10%) | ||
| TNF blockers (Humira) | 1 (10%) | ||
| Immunosuppressants (Azathioprine) | 1 (10%) | ||
| Sera autoantibody median levels (Q25, Q75; units/mL) | Healthy women n = 10 | Women with SBI n = 10 | p-Value |
| Anti-β1 adrenergic receptor | 47.8 (22.8, 54.6) | 8.4 (7.4, 10.2) | 0.000 **** |
| Anti-angiotensin II type 1 receptor | 27.6 (13.3, 43.9) | 10.4 (9.9, 11.9) | 0.007 ** |
| Anti-endothelin receptor type A | 14.7 (10.8, 17.9) | 9.0 (7.8, 10.1) | 0.087 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Talalai, E.; Gorobets, D.; Halpert, G.; Tsur, A.M.; Heidecke, H.; Levy, Y.; Watad, A.; Blank, M.; Michaelevski, I.; Shoenfeld, Y.; et al. Functional IgG Autoantibodies against Autonomic Nervous System Receptors in Symptomatic Women with Silicone Breast Implants. Cells 2023, 12, 1510. https://doi.org/10.3390/cells12111510
Talalai E, Gorobets D, Halpert G, Tsur AM, Heidecke H, Levy Y, Watad A, Blank M, Michaelevski I, Shoenfeld Y, et al. Functional IgG Autoantibodies against Autonomic Nervous System Receptors in Symptomatic Women with Silicone Breast Implants. Cells. 2023; 12(11):1510. https://doi.org/10.3390/cells12111510
Chicago/Turabian StyleTalalai, Efrosiniia, Denis Gorobets, Gilad Halpert, Avishai M. Tsur, Harald Heidecke, Yair Levy, Abdulla Watad, Miri Blank, Izhak Michaelevski, Yehuda Shoenfeld, and et al. 2023. "Functional IgG Autoantibodies against Autonomic Nervous System Receptors in Symptomatic Women with Silicone Breast Implants" Cells 12, no. 11: 1510. https://doi.org/10.3390/cells12111510
APA StyleTalalai, E., Gorobets, D., Halpert, G., Tsur, A. M., Heidecke, H., Levy, Y., Watad, A., Blank, M., Michaelevski, I., Shoenfeld, Y., & Amital, H. (2023). Functional IgG Autoantibodies against Autonomic Nervous System Receptors in Symptomatic Women with Silicone Breast Implants. Cells, 12(11), 1510. https://doi.org/10.3390/cells12111510