



Male DAT Val559 Mice Exhibit Compulsive Behavior under Devalued Reward Conditions Accompanied by Cellular and Pharmacological Changes

 , and
, and 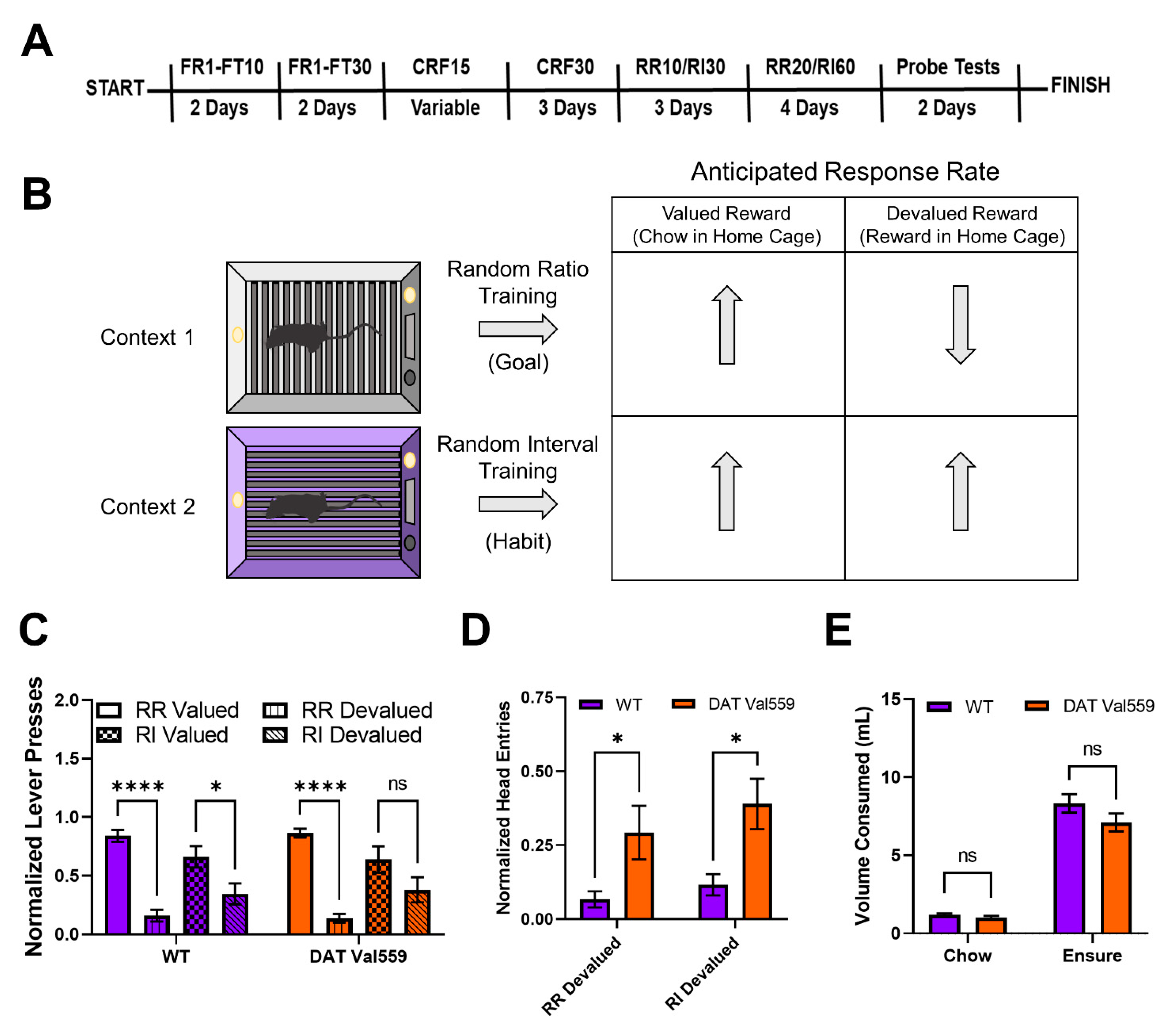{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Goal vs. Habit Nose Poke Paradigm
- The training schedule consisted of two days of FR1-FT10 (fixed ratio 1-fixed time 10) where the trial started with the delivery of a free reward (Ensure®) that the mouse had 5 s to consume upon head entry into the reward delivery zone. The mouse could then earn a reward by making a nose poke into a nose poke hole backlit by LEDs, otherwise a reward was freely delivered every 10 s (FR1-FT10). Mice had to nose poke into an active hole (indicated by a lit LED in the back of the hole) to elicit delivery of a reward. Mice were tested under the FR1-FT10 schedule for 15 min sessions and 15 rewards/session for each training context. Only the holes to the left and right of the reward delivery zone could be active holes. This FR1-FT10 schedule was followed by 2 days of FR1-FT30 (15 min sessions, 15 rewards/session).
- Mice then underwent CRF training (60 min sessions, 15 rewards/context). Under this protocol mice received a free reward at the beginning of the session, but then had to complete a nose poke on the active hole (indicated by LED) to earn additional rewards. All mice had to reach the criterion of earning 15 rewards within 60 min in both training contexts before moving to the next training stage as a group. To prevent overtraining, mice that obtained this criterion sooner were rested, and total training sessions at CRF 15 were kept equivalent across mice +/− 1 training session.
- Mice were then run three days on CRF with 30 rewards possible.
- Mice were then trained on random ratio 10 (RR10; reward was delivered on average every 10 nose pokes with a 0.1 probability that a given nose poke would produce a reward) and random interval 30 s (RI30; reward was delivered upon nose poke on average every 30 s regardless of nose poke vigor) for 3 days. Mice were balanced across training context for which hole was the active hole (i.e., for mouse 1 the active nose poke hole was the left hole in the clear plastic environmental context while the right hole was the active nose poke hole in the striped environmental context, but this condition was reversed for mouse 2).
- After the RR10 and RI30 training the intensity of the schedules were increased to random ratio 20 (RR20) and random interval 60 (RI60) where animals were given 60 min in each context to earn 30 rewards.
- Upon completion of RR20/RI60 training mice underwent two days of five-minute non-reinforced probe tests done under devalued (Ensure® given in home cage for 1 h) and valued states (mouse chow given in home cage for 1 h), counterbalanced across days, context, and genotype. Prior to the start of the non-reinforced probe tests, mice were separated into clean cages and allowed access to either mouse chow (valued probe test days) or Ensure® (devalued probe test days) for an hour. The amount of chow and Ensure® consumed in home cage was measured to confirm that there were no consumption differences across genotypes. The full training schedule is shown in Figure 1A.
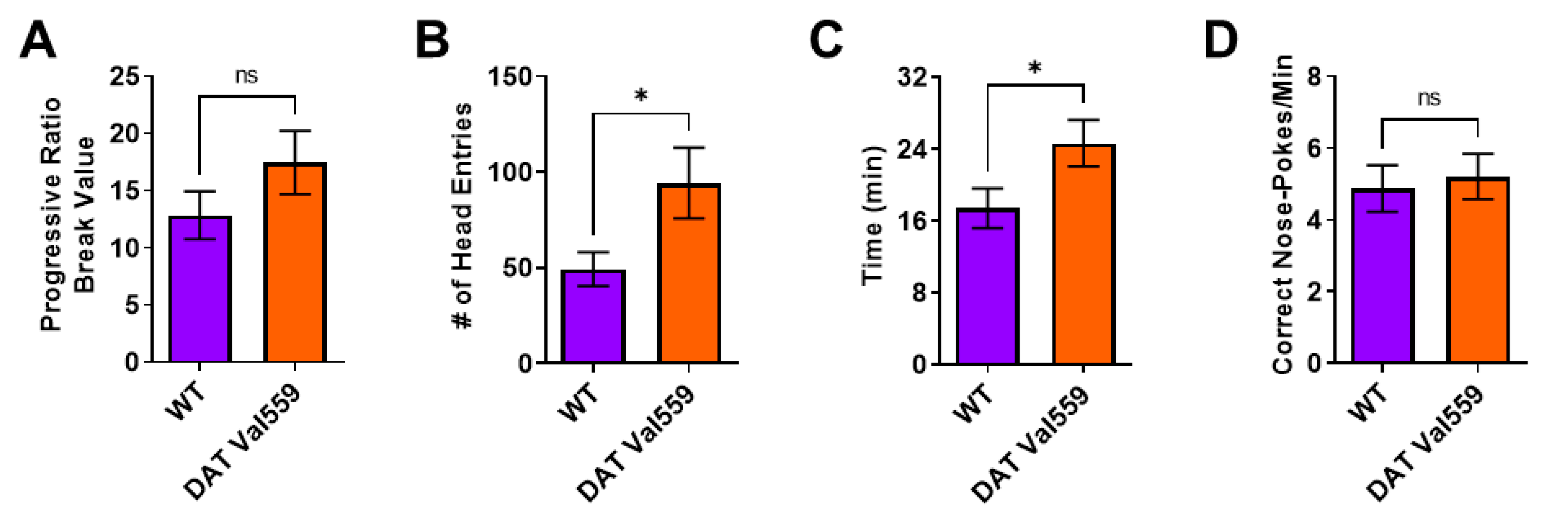
2.3. Devaluation of Progressive Ratio
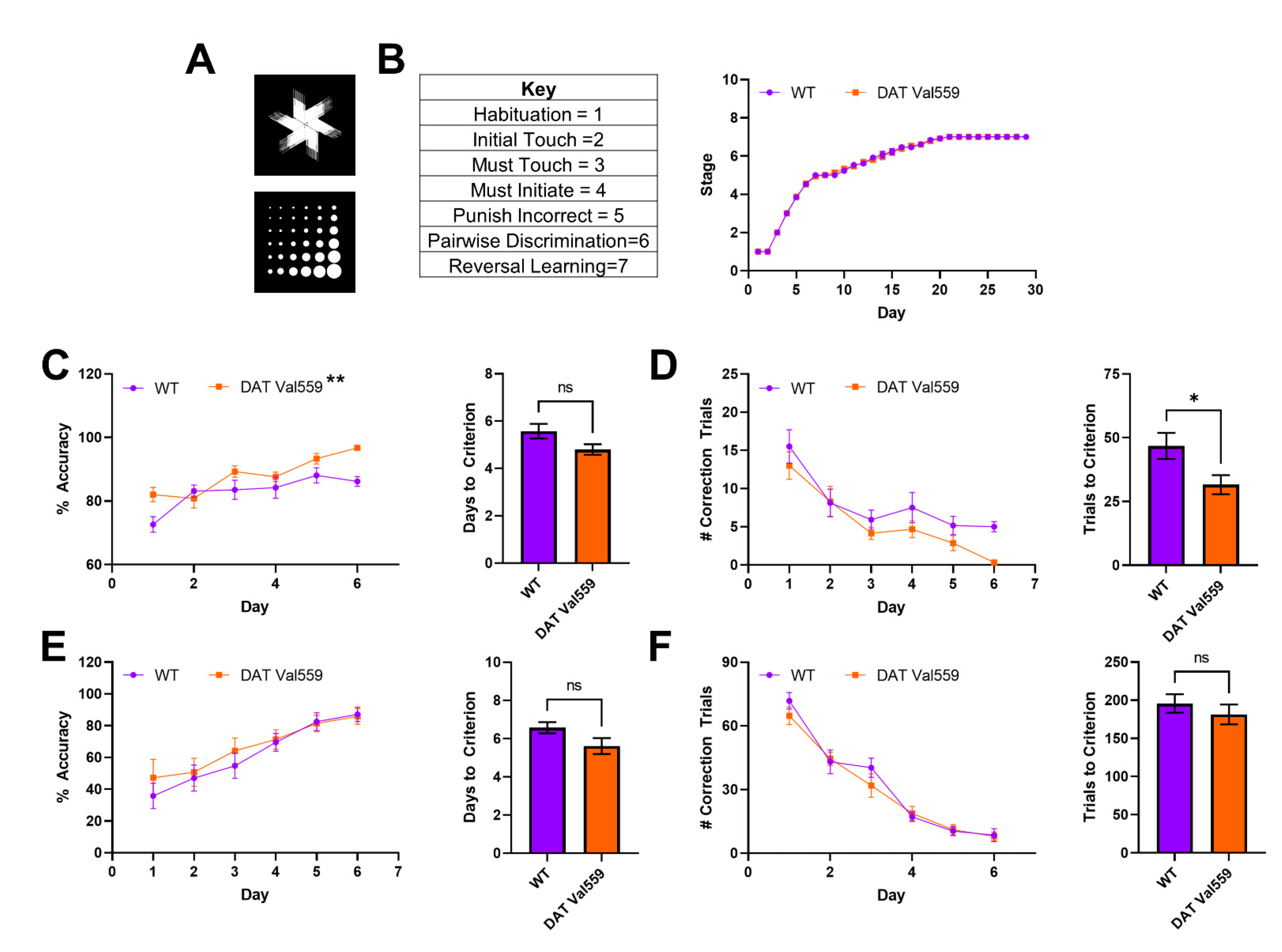
2.4. Pairwise Discrimination and Reversal Learning
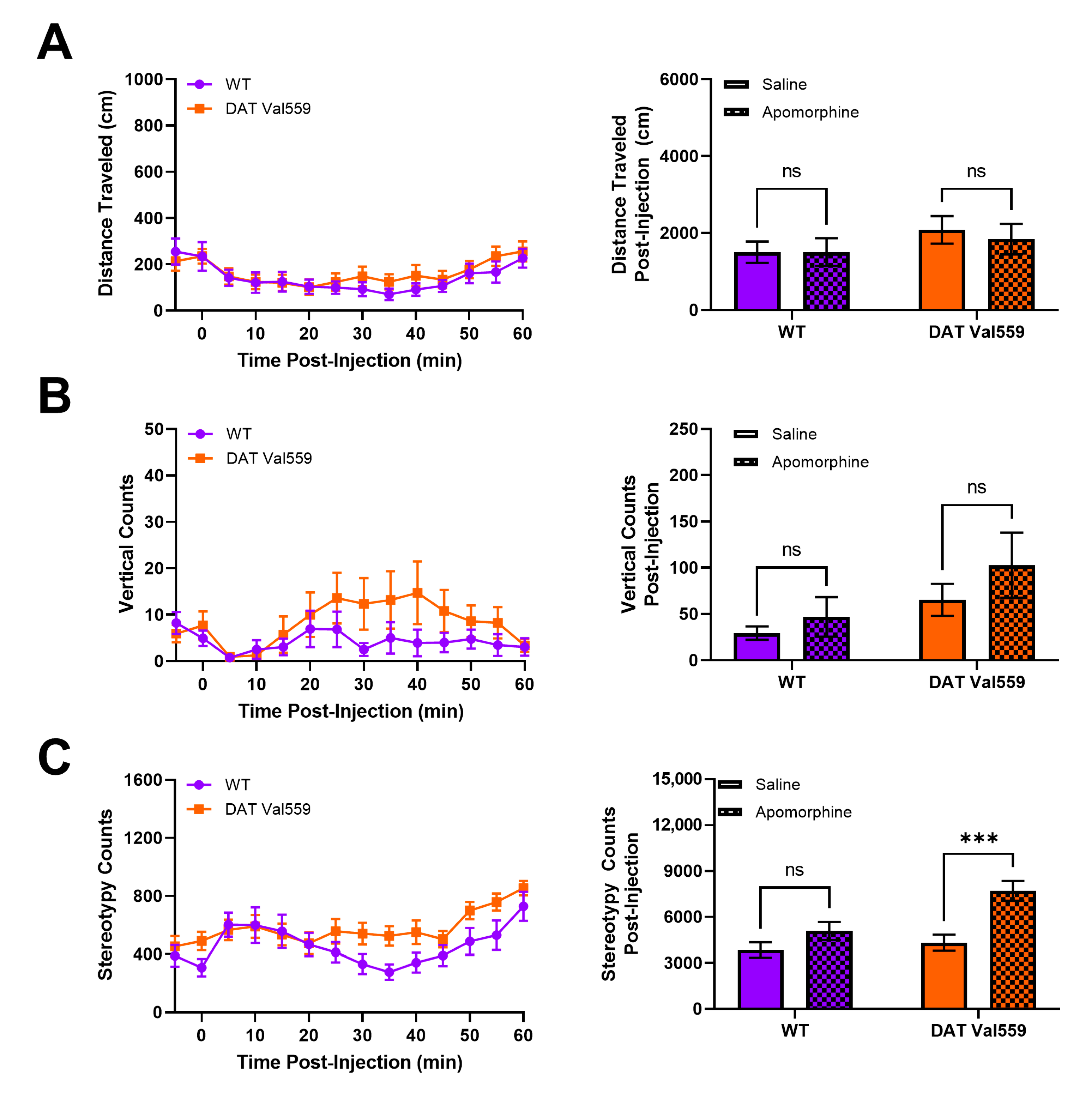
2.5. Apomorphine-Induced Locomotion
2.6. Golgi Staining and Dendritic Morphology
2.7. Statistical Analyses
3. Results
3.1. DAT VAL559 Males Display Compulsive Reward Seeking in Devalued States for Both Goal-Directed and Habit Driven Behavioral Contexts
3.2. DAT Val559 Dysregulated Reward Seeking Is also Present with Progressive Ratio Testing under Devalued Conditions
3.3. DAT Val559 Males Display Enhanced Learning in Pairwise Discrimination Task but No Deficits in Cognitive Flexibility
3.4. DAT Val559 Males Exhibit Increased Sensitivity to DA Agonist-Induced Repetitive Motor Behaviors
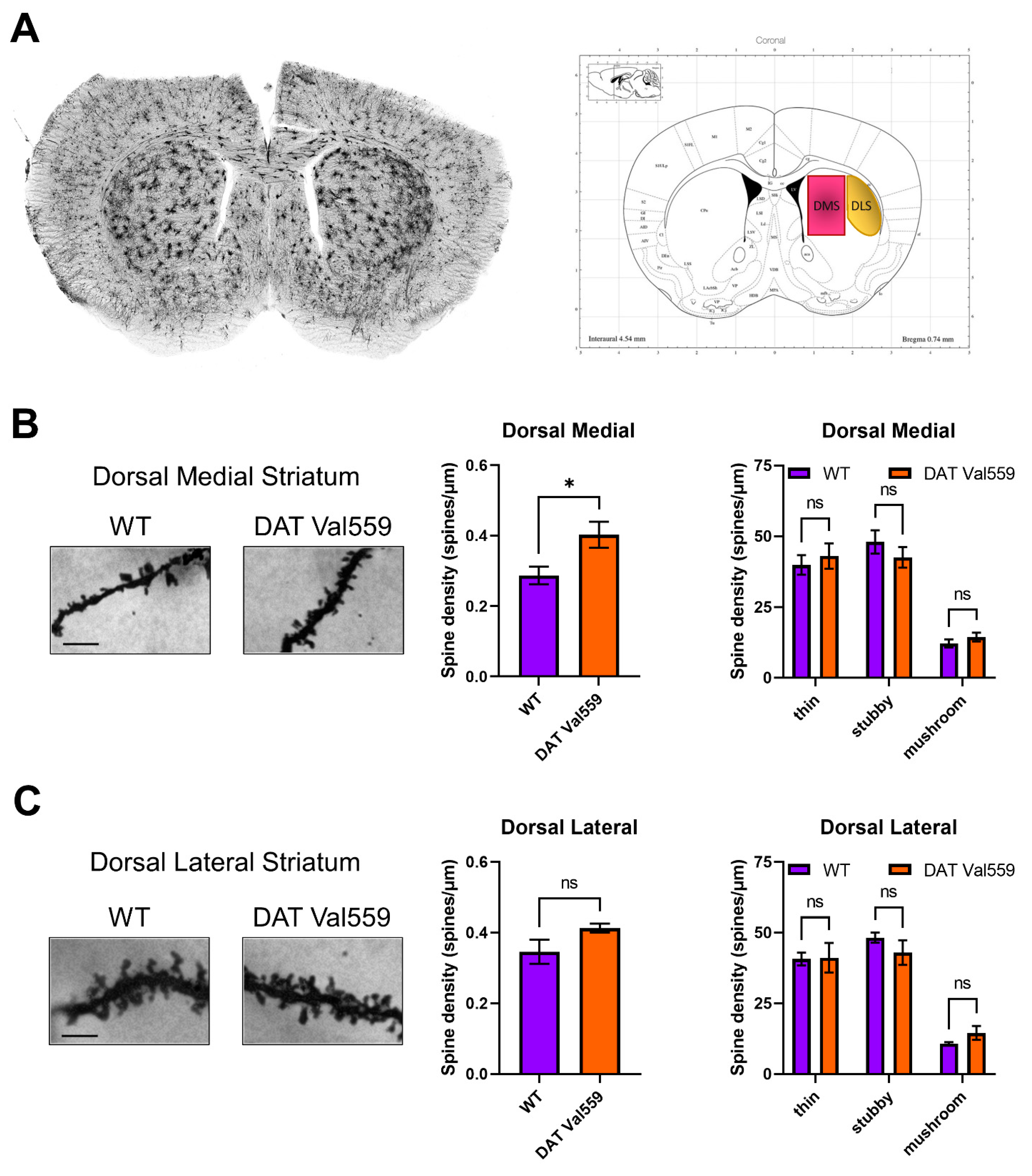
3.5. Dendritic Spine Density Is Increased in the DMS of Male DAT Val559 Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goodman, J.; Marsh, R.; Peterson, B.S.; Packard, M.G. Annual Research Review: The neurobehavioral development of multiple memory systems—Implications for childhood and adolescent psychiatric disorders. J. Child Psychol. Psychiatry 2014, 55, 582–610. [Google Scholar] [CrossRef]
- Lipton, D.M.; Gonzales, B.J.; Citri, A. Dorsal striatal circuits for habits, compulsions and addictions. Front. Syst. Neurosci. 2019, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.J.; Robbins, T.W. From the ventral to the dorsal striatum: Devolving views of their roles in drug addiction. Neurosci. Biobehav. Rev. 2013, 37, 1946–1954. [Google Scholar] [CrossRef]
- Yin, H.H.; Mulcare, S.P.; Hilario, M.R.; Clouse, E.; Holloway, T.; Davis, M.I.; Hansson, A.C.; Lovinger, D.M.; Costa, R.M. Dynamic reorganization of striatal circuits during the acquisition and consolidation of a skill. Nat. Neurosci. 2009, 12, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Gremel, C.M.; Costa, R.M. Orbitofrontal and striatal circuits dynamically encode the shift between goal-directed and habitual actions. Nat. Commun. 2013, 4, 2264. [Google Scholar] [CrossRef]
- Kupferschmidt, D.A.; Juczewski, K.; Cui, G.; Johnson, K.A.; Lovinger, D.M. Parallel, but dissociable, processing in discrete corticostriatal inputs encodes skill learning. Neuron 2017, 96, 476–489.e5. [Google Scholar] [CrossRef]
- Vandaele, Y.; Mahajan, N.R.; Ottenheimer, D.J.; Richard, J.M.; Mysore, S.P.; Janak, P.H. Distinct recruitment of dorsomedial and dorsolateral striatum erodes with extended training. eLife 2019, 8, e49536. [Google Scholar] [CrossRef]
- Yin, H.H.; Knowlton, B.J.; Balleine, B.W. Lesions of dorsolateral striatum preserve outcome expectancy but disrupt habit formation in instrumental learning. Eur. J. Neurosci. 2004, 19, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.H.; Knowlton, B.J.; Balleine, B.W. Blockade of NMDA receptors in the dorsomedial striatum prevents action-outcome learning in instrumental conditioning. Eur. J. Neurosci. 2005, 22, 505–512. [Google Scholar] [CrossRef]
- Kim, H.F.; Ghazizadeh, A.; Hikosaka, O. Dopamine neurons encoding long-term memory of bbject value for habitual behavior. Cell 2015, 163, 1165–1175. [Google Scholar] [CrossRef]
- Faure, A.; Haberland, U.; Conde, F.; El Massioui, N. Lesion to the nigrostriatal dopamine system disrupts stimulus-response habit formation. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 2771–2780. [Google Scholar] [CrossRef]
- Leong, K.C.; Berini, C.R.; Ghee, S.M.; Reichel, C.M. Extended cocaine-seeking produces a shift from goal-directed to habitual responding in rats. Physiol. Behav. 2016, 164, 330–335. [Google Scholar] [CrossRef]
- Nelson, A.; Killcross, S. Amphetamine exposure enhances habit formation. J. Neuro. 2006, 26, 3805–3812. [Google Scholar] [CrossRef]
- DiFeliceantonio, A.G.; Berridge, K.C. Dorsolateral neostriatum contribution to incentive salience: Opioid or dopamine stimulation makes one reward cue more motivationally attractive than another. Eur. J. Neurosci. 2016, 43, 1203–1218. [Google Scholar] [CrossRef] [PubMed]
- Seiler, J.L.; Cosme, C.V.; Sherathiya, V.N.; Schaid, M.D.; Bianco, J.M.; Bridgemohan, A.S.; Lerner, T.N. Dopamine signaling in the dorsomedial striatum promotes compulsive behavior. Curr. Biol. 2022, 32, 1175–1188.e5. [Google Scholar] [CrossRef]
- Van Elzelingen, W.; Warnaar, P.; Matos, J.; Bastet, W.; Jonkman, R.; Smulders, D.; Goedhoop, J.; Denys, D.; Arbab, T.; Willuhn, I. Striatal dopamine signals are region specific and temporally stable across action-sequence habit formation. Curr. Biol. 2022, 32, 1163–1174.e6. [Google Scholar] [CrossRef]
- Giros, B.; Jaber, M.; Jones, S.R.; Wightman, R.M.; Caron, M.G. Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter. Nature 1996, 379, 606–612. [Google Scholar] [CrossRef]
- Jones, S.R.; Gainetdinov, R.R.; Jaber, M.; Giros, B.; Wightman, R.M.; Caron, M.G. Profound neuronal plasticity in response to inactivation of the dopamine transporter. Proc. Natl. Acad. Sci. USA 1998, 95, 4029–4034. [Google Scholar] [CrossRef]
- Benoit-Marand, M.; Jaber, M.; Gonon, F. Release and elimination of dopamine in vivo in mice lacking the dopamine transporter: Functional consequences. Eur. J. Neurosci. 2000, 12, 2985–2992. [Google Scholar] [CrossRef]
- Nordquist, R.E.; Voorn, P.; de Mooij-van Malsen, J.G.; Joosten, R.N.; Pennartz, C.M.; Vanderschuren, L.J. Augmented reinforcer value and accelerated habit formation after repeated amphetamine treatment. Eur. Neuropsychopharmacol. 2007, 17, 532–540. [Google Scholar] [CrossRef]
- Mergy, M.A.; Gowrishankar, R.; Davis, G.L.; Jessen, T.N.; Wright, J.; Stanwood, G.D.; Hahn, M.K.; Blakely, R.D. Genetic targeting of the amphetamine and methylphenidate-sensitive dopamine transporter: On the path to an animal model of attention-deficit hyperactivity disorder. Neurochem. Int. 2014, 73, 56–70. [Google Scholar] [CrossRef]
- Gowrishankar, R.; Hahn, M.K.; Blakely, R.D. Good riddance to dopamine: Roles for the dopamine transporter in synaptic function and dopamine-associated brain disorders. Neurochem. Int. 2014, 73, 42–48. [Google Scholar] [CrossRef]
- Mazei-Robison, M.S.; Couch, R.S.; Shelton, R.C.; Stein, M.A.; Blakely, R.D. Sequence variation in the human dopamine transporter gene in children with attention deficit hyperactivity disorder. Neuropharmacology 2005, 49, 724–736. [Google Scholar] [CrossRef]
- Hamilton, P.J.; Campbell, N.G.; Sharma, S.; Erreger, K.; Herborg Hansen, F.; Saunders, C.; Belovich, A.N.; Consortium, N.A.A.S.; Sahai, M.A.; Cook, E.H.; et al. De novo mutation in the dopamine transporter gene associates dopamine dysfunction with autism spectrum disorder. Mol. Psychiatry 2013, 18, 1315–1323. [Google Scholar] [CrossRef]
- Hansen, F.H.; Skjorringe, T.; Yasmeen, S.; Arends, N.V.; Sahai, M.A.; Erreger, K.; Andreassen, T.F.; Holy, M.; Hamilton, P.J.; Neergheen, V.; et al. Missense dopamine transporter mutations associate with adult parkinsonism and ADHD. J. Clin. Investig. 2014, 124, 3107–3120. [Google Scholar] [CrossRef]
- Sakrikar, D.; Mazei-Robison, M.S.; Mergy, M.A.; Richtand, N.W.; Han, Q.; Hamilton, P.J.; Bowton, E.; Galli, A.; Veenstra-Vanderweele, J.; Gill, M.; et al. Attention deficit/hyperactivity disorder-derived coding variation in the dopamine transporter disrupts microdomain targeting and trafficking regulation. J. Neuro. 2012, 32, 5385–5397. [Google Scholar] [CrossRef]
- Bowton, E.; Saunders, C.; Reddy, I.A.; Campbell, N.G.; Hamilton, P.J.; Henry, L.K.; Coon, H.; Sakrikar, D.; Veenstra-VanderWeele, J.M.; Blakely, R.D.; et al. SLC6A3 coding variant Ala559Val found in two autism probands alters dopamine transporter function and trafficking. Transl. Psychiatry 2014, 4, e464. [Google Scholar] [CrossRef]
- Grunhage, F.; Schulze, T.G.; Muller, D.J.; Lanczik, M.; Franzek, E.; Albus, M.; Borrmann-Hassenbach, M.; Knapp, M.; Cichon, S.; Maier, W.; et al. Systematic screening for DNA sequence variation in the coding region of the human dopamine transporter gene (DAT1). Mol. Psychiatry 2000, 5, 275–282. [Google Scholar] [CrossRef]
- Mazei-Robison, M.S.; Bowton, E.; Holy, M.; Schmudermaier, M.; Freissmuth, M.; Sitte, H.H.; Galli, A.; Blakely, R.D. Anomalous dopamine release associated with a human dopamine transporter coding variant. J. Neuro. 2008, 28, 7040–7046. [Google Scholar] [CrossRef]
- Gowrishankar, R.; Gresch, P.J.; Davis, G.L.; Katamish, R.M.; Riele, J.R.; Stewart, A.M.; Vaughan, R.A.; Hahn, M.K.; Blakely, R.D. Region-specific regulation of presynaptic dopamine homeostasis by D2 autoreceptors shapes the in vivo—Impact of the neuropsychiatric disease-associated DAT variant Val559. J. Neuro. 2018, 38, 5302–5312. [Google Scholar] [CrossRef]
- Mergy, M.A.; Gowrishankar, R.; Gresch, P.J.; Gantz, S.C.; Williams, J.; Davis, G.L.; Wheeler, C.A.; Stanwood, G.D.; Hahn, M.K.; Blakely, R.D. The rare DAT coding variant Val559 perturbs DA neuron function, changes behavior, and alters in vivo responses to psychostimulants. Proc. Natl. Acad. Sci. USA 2014, 111, E4779–E4788. [Google Scholar] [CrossRef]
- Stewart, A.; Davis, G.L.; Gresch, P.J.; Katamish, R.M.; Peart, R.; Rabil, M.J.; Gowrishankar, R.; Carroll, F.I.; Hahn, M.K.; Blakely, R.D. Serotonin transporter inhibition and 5-HT2C receptor activation drive loss of cocaine-induced locomotor activation in DAT Val559 mice. Neuropsychopharmacology 2018, 44, 994–1006. [Google Scholar] [CrossRef]
- Davis, G.L.; Stewart, A.; Stanwood, G.D.; Gowrishankar, R.; Hahn, M.K.; Blakely, R.D. Functional coding variation in the presynaptic dopamine transporter associated with neuropsychiatric disorders drives enhanced motivation and context-dependent impulsivity in mice. Behav. Brain Res. 2018, 337, 61–69. [Google Scholar] [CrossRef]
- Schultz, W. Multiple dopamine functions at different time courses. Annu. Rev. Neurosci. 2007, 30, 259–288. [Google Scholar] [CrossRef]
- Schultz, W. Behavioral dopamine signals. Trends Neurosci. 2007, 30, 203–210. [Google Scholar] [CrossRef]
- Izquierdo, A.; Jentsch, J.D. Reversal learning as a measure of impulsive and compulsive behavior in addictions. Psychopharmacology 2012, 219, 607–620. [Google Scholar] [CrossRef]
- Torregrossa, M.M.; Quinn, J.J.; Taylor, J.R. Impulsivity, compulsivity, and habit: The role of orbitofrontal cortex revisited. Biol. Psychiatry 2008, 63, 253–255. [Google Scholar] [CrossRef]
- Klanker, M.; Feenstra, M.; Denys, D. Dopaminergic control of cognitive flexibility in humans and animals. Front. Neurosci. 2013, 7, 201. [Google Scholar] [CrossRef]
- Miller, H.L.; Ragozzino, M.E.; Cook, E.H.; Sweeney, J.A.; Mosconi, M.W. Cognitive set shifting deficits and their relationship to repetitive behaviors in autism spectrum disorder. J. Autism. Dev. Disord. 2015, 45, 805–815. [Google Scholar] [CrossRef]
- O’Donnell, L.A.; Deldin, P.J.; Pester, B.; McInnis, M.G.; Langenecker, S.A.; Ryan, K.A. Cognitive flexibility: A trait of bipolar disorder that worsens with length of illness. J. Clin. Exp. Neuropsychol. 2017, 39, 979–987. [Google Scholar] [CrossRef]
- Getahun, D.; Jacobsen, S.J.; Fassett, M.J.; Chen, W.; Demissie, K.; Rhoads, G.G. Recent trends in childhood attention-deficit/hyperactivity disorder. JAMA Pediatrics 2013, 167, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.J.; Naughton, B.J.; Thirtamara-Rajamani, K.; Yoon, D.J.; Han, D.D.; Devries, A.C.; Gu, H.H. Dopamine transporter inhibition is necessary for cocaine-induced increases in dendritic spine density in the nucleus accumbens. Synapse 2011, 65, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zeng, C.; Li, J.; Zhou, Z.; Ju, X.; Xia, S.; Li, Y.; Liu, A.; Teng, H.; Zhang, K.; et al. PAK2 haploinsufficiency results in synaptic cytoskeleton impairment and autism-related behavior. Cell Rep. 2018, 24, 2029–2041. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Kim, Y.; Kim, A.M.; Helmin, K.; Nairn, A.C.; Greengard, P. Cocaine-induced dendritic spine formation in D1 and D2 dopamine receptor-containing medium spiny neurons in nucleus accumbens. Proc. Natl. Acad. Sci. USA 2006, 103, 3399–3404. [Google Scholar] [CrossRef]
- Rodriguez, A.; Ehlenberger, D.B.; Dickstein, D.L.; Hof, P.R.; Wearne, S.L. Automated three-dimensional detection and shape classification of dendritic spines from fluorescence microscopy images. PLoS ONE 2008, 3, e1997. [Google Scholar] [CrossRef]
- Stewart, A.; Mayer, F.P.; Gowrishankar, R.; Davis, G.L.; Areal, L.B.; Gresch, P.J.; Katamish, R.M.; Peart, R.; Stilley, S.E.; Spiess, K.; et al. Behaviorally penetrant, anomalous dopamine efflux exposes sex and circuit dependent regulation of dopamine transporters. Mol. Psychiatry 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Allichon, M.C.; Ortiz, V.; Pousinha, P.; Andrianarivelo, A.; Petitbon, A.; Heck, N.; Trifilieff, P.; Barik, J.; Vanhoutte, P. Cell-type-specific adaptions in striatal medium-sized spiny neurons and their roles in Behavioral Responses to Drugs of Abuse. Front. Synaptic. Neurosci. 2021, 13, 799274. [Google Scholar] [CrossRef]
- Dos Santos, M.; Cahill, E.N.; Bo, G.D.; Vanhoutte, P.; Caboche, J.; Giros, B.; Heck, N. Cocaine increases dopaminergic connectivity in the nucleus accumbens. Brain Struct. Funct. 2018, 223, 913–923. [Google Scholar] [CrossRef]
- Alcantara, A.A.; Lim, H.Y.; Floyd, C.E.; Garces, J.; Mendenhall, J.M.; Lyons, C.L.; Berlanga, M.L. Cocaine- and morphine-induced synaptic plasticity in the nucleus accumbens. Synapse 2011, 65, 309–320. [Google Scholar] [CrossRef]
- Marie, N.; Canestrelli, C.; Noble, F. Transfer of neuroplasticity from nucleus accumbens core to shell is required for cocaine reward. PLoS ONE 2012, 7, e30241. [Google Scholar] [CrossRef]
- Everitt, B.J.; Robbins, T.W. Neural systems of reinforcement for drug addiction: From actions to habits to compulsion. Nat. Neurosci. 2005, 8, 1481. [Google Scholar] [CrossRef]
- O’Sullivan, S.S.; Wu, K.; Politis, M.; Lawrence, A.D.; Evans, A.H.; Bose, S.K.; Djamshidian, A.; Lees, A.J.; Piccini, P. Cue-induced striatal dopamine release in Parkinson’s disease-associated impulsive-compulsive behaviours. Brain 2011, 134, 969–978. [Google Scholar] [CrossRef]
- Stuchlik, A.; Radostova, D.; Hatalova, H.; Vales, K.; Nekovarova, T.; Koprivova, J.; Svoboda, J.; Horacek, J. Validity of quinpirole sensitization rat model of OCD: Linking evidence from animal and clinical studies. Front. Behav. Neurosci. 2016, 10, 209. [Google Scholar] [CrossRef]
- Eagle, D.M.; Noschang, C.; d’Angelo, L.-S.C.; Noble, C.A.; Day, J.O.; Dongelmans, M.L.; Theobald, D.E.; Mar, A.C.; Urcelay, G.P.; Morein-Zamir, S.; et al. The dopamine D2/D3 receptor agonist quinpirole increases checking-like behaviour in an operant observing response task with uncertain reinforcement: A novel possible model of OCD? Behav. Brain Res. 2014, 264, 207–229. [Google Scholar] [CrossRef]
- Dorfman, A.; Szechtman, H.; Eilam, D. Social interaction modulates the intensity of compulsive checking in a rat model of obsessive-compulsive disorder (OCD). Behav. Brain Res. 2019, 359, 156–164. [Google Scholar] [CrossRef]
- Argiolas, A.; Hedlund, H. The pharmacology and clinical pharmacokinetics of apomorphine SL. BJU Int. 2001, 88, 18–21. [Google Scholar] [CrossRef]
- De Steno, D.A.; Schmauss, C. A role for dopamine D2 receptors in reversal learning. Neuroscience 2009, 162, 118–127. [Google Scholar] [CrossRef][Green Version]
- Kim, B.; Yoon, S.; Nakajima, R.; Lee, H.J.; Lim, H.J.; Lee, Y.K.; Choi, J.S.; Yoon, B.J.; Augustine, G.J.; Baik, J.H. Dopamine D2 receptor-mediated circuit from the central amygdala to the bed nucleus of the stria terminalis regulates impulsive behavior. Proc. Natl. Acad. Sci. USA 2018, 115, E10730–E10739. [Google Scholar] [CrossRef]
- Linden, J.; James, A.S.; McDaniel, C.; Jentsch, J.D. Dopamine D2 receptors in dopaminergic neurons modulate performance in a reversal learning task in mice. eNeuro 2018, 5, 1. [Google Scholar] [CrossRef]
- Jia, J.M.; Zhao, J.; Hu, Z.; Lindberg, D.; Li, Z. Age-dependent regulation of synaptic connections by dopamine D2 receptors. Nat. Neurosci. 2013, 16, 1627–1636. [Google Scholar] [CrossRef]
- Berlanga, M.L.; Price, D.L.; Phung, B.S.; Giuly, R.; Terada, M.; Yamada, N.; Cyr, M.; Caron, M.G.; Laakso, A.; Martone, M.E.; et al. Multiscale imaging characterization of dopamine transporter knockout mice reveals regional alterations in spine density of medium spiny neurons. Brain Res. 2011, 1390, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Chohan, M.O.; Kopelman, J.M.; Yueh, H.; Fazlali, Z.; Greene, N.; Harris, A.Z.; Balsam, P.D.; Leonardo, E.D.; Kramer, E.R.; Veenstra-VanderWeele, J.; et al. Developmental impact of glutamate transporter overexpression on dopaminergic neuron activity and stereotypic behavior. Mol. Psychiatry 2022, 27, 1515–1526. [Google Scholar] [CrossRef] [PubMed]
- Pascoli, V.; Hiver, A.; Li, Y.; Harada, M.; Esmaeili, V.; Luscher, C. Cell-type specific synaptic plasticity in dorsal striatum is associated with punishment-resistance compulsive-like cocaine self-administration in mice. Neuropsychopharmacology 2022. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, K.C.; Connor, M.; Pierson, J.; Shuffrey, L.C.; Blakely, R.D.; Ahmari, S.E.; Veenstra-VanderWeele, J. Serotonin 5-HT1B receptor-mediated behavior and binding in mice with the overactive and dysregulated serotonin transporter Ala56 variant. Psychopharmacology 2021, 238, 1111–1120. [Google Scholar] [CrossRef]
- Kang, J.; Kim, H.; Hwang, S.H.; Han, M.; Lee, S.H.; Kim, H.F. Primate ventral striatum maintains neural representations of the value of previously rewarded objects for habitual seeking. Nat. Commun. 2021, 12, 2100. [Google Scholar] [CrossRef]
- Tanner, M.K.; Davis, J.K.P.; Jaime, J.; Moya, N.A.; Hohorst, A.A.; Bonar, K.; Abrams, K.A.; Jamil, N.; Han, R.; Hubert, T.J.; et al. Duration- and sex-dependent neural circuit control of voluntary physical activity. Psychopharmacology 2022, 239, 3697–3709. [Google Scholar] [CrossRef]
- Matyjek, M.; Bayer, M.; Dziobek, I. Autistic Traits Affect Reward Anticipation but not Reception. Sci. Rep. 2020, 10, 8396. [Google Scholar] [CrossRef]
- Grimm, O.; van Rooij, D.; Tshagharyan, A.; Yildiz, D.; Leonards, J.; Elgohary, A.; Buitelaar, J.; Reif, A. Effects of comorbid disorders on reward processing and connectivity in adults with ADHD. Transl. Psychiatry 2021, 11, 636. [Google Scholar] [CrossRef]
- Nimarko, A.F.; Gorelik, A.J.; Carta, K.E.; Gorelik, M.G.; Singh, M.K. Neural correlates of reward processing distinguish healthy youth at familial risk for bipolar disorder from youth at familial risk for major depressive disorder. Transl. Psychiatry 2022, 12, 31. [Google Scholar] [CrossRef]
- Rosch, K.S.; Hawk, L.W., Jr. The effects of performance-based rewards on neurophysiological correlates of stimulus, error, and feedback processing in children with ADHD. Psychophysiology 2013, 50, 1157–1173. [Google Scholar] [CrossRef]
- Jones, E.J.; Webb, S.J.; Estes, A.; Dawson, G. Rule learning in autism: The role of reward type and social context. Dev. Neuropsychol. 2013, 38, 58–77. [Google Scholar] [CrossRef] [PubMed]
- Kohls, G.; Thonessen, H.; Bartley, G.K.; Grossheinrich, N.; Fink, G.R.; Herpertz-Dahlmann, B.; Konrad, K. Differentiating neural reward responsiveness in autism versus ADHD. Dev. Cogn. Neurosci. 2014, 10, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Kohls, G.; Antezana, L.; Mosner, M.G.; Schultz, R.T.; Yerys, B.E. Altered reward system reactivity for personalized circumscribed interests in autism. Mol. Autism 2018, 9, 9. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stewart, A.; Davis, G.L.; Areal, L.B.; Rabil, M.J.; Tran, V.; Mayer, F.P.; Blakely, R.D. Male DAT Val559 Mice Exhibit Compulsive Behavior under Devalued Reward Conditions Accompanied by Cellular and Pharmacological Changes. Cells 2022, 11, 4059. https://doi.org/10.3390/cells11244059
Stewart A, Davis GL, Areal LB, Rabil MJ, Tran V, Mayer FP, Blakely RD. Male DAT Val559 Mice Exhibit Compulsive Behavior under Devalued Reward Conditions Accompanied by Cellular and Pharmacological Changes. Cells. 2022; 11(24):4059. https://doi.org/10.3390/cells11244059
Chicago/Turabian StyleStewart, Adele, Gwynne L. Davis, Lorena B. Areal, Maximilian J. Rabil, Vuong Tran, Felix P. Mayer, and Randy D. Blakely. 2022. "Male DAT Val559 Mice Exhibit Compulsive Behavior under Devalued Reward Conditions Accompanied by Cellular and Pharmacological Changes" Cells 11, no. 24: 4059. https://doi.org/10.3390/cells11244059
APA StyleStewart, A., Davis, G. L., Areal, L. B., Rabil, M. J., Tran, V., Mayer, F. P., & Blakely, R. D. (2022). Male DAT Val559 Mice Exhibit Compulsive Behavior under Devalued Reward Conditions Accompanied by Cellular and Pharmacological Changes. Cells, 11(24), 4059. https://doi.org/10.3390/cells11244059