

Alterations of Lipid Metabolism in the Heart in Spontaneously Hypertensive Rats Precedes Left Ventricular Hypertrophy and Cardiac Dysfunction


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Blood and Tissue Sampling
2.4. Echocardiography
2.5. Gene Expression Analysis
2.6. Western Blot Analysis
2.7. Measurement of Lipids
2.8. Desaturation and Elongation Indices
2.9. Statistical Analysis
3. Results
3.1. Plasma Parameters
3.2. Myocardial Structure and Function
3.3. Molecular Pathways of Myocardial Remodeling in SHRs
3.3.1. Molecular Indicators of Myocardial Remodeling
3.3.2. AKT Pathway
3.3.3. Other Signaling Pathways Involved in the Development of Cardiac Hypertrophy
3.4. Lipid Content in the Heart
3.5. Lipogenesis Pathway in Cardiomyocytes
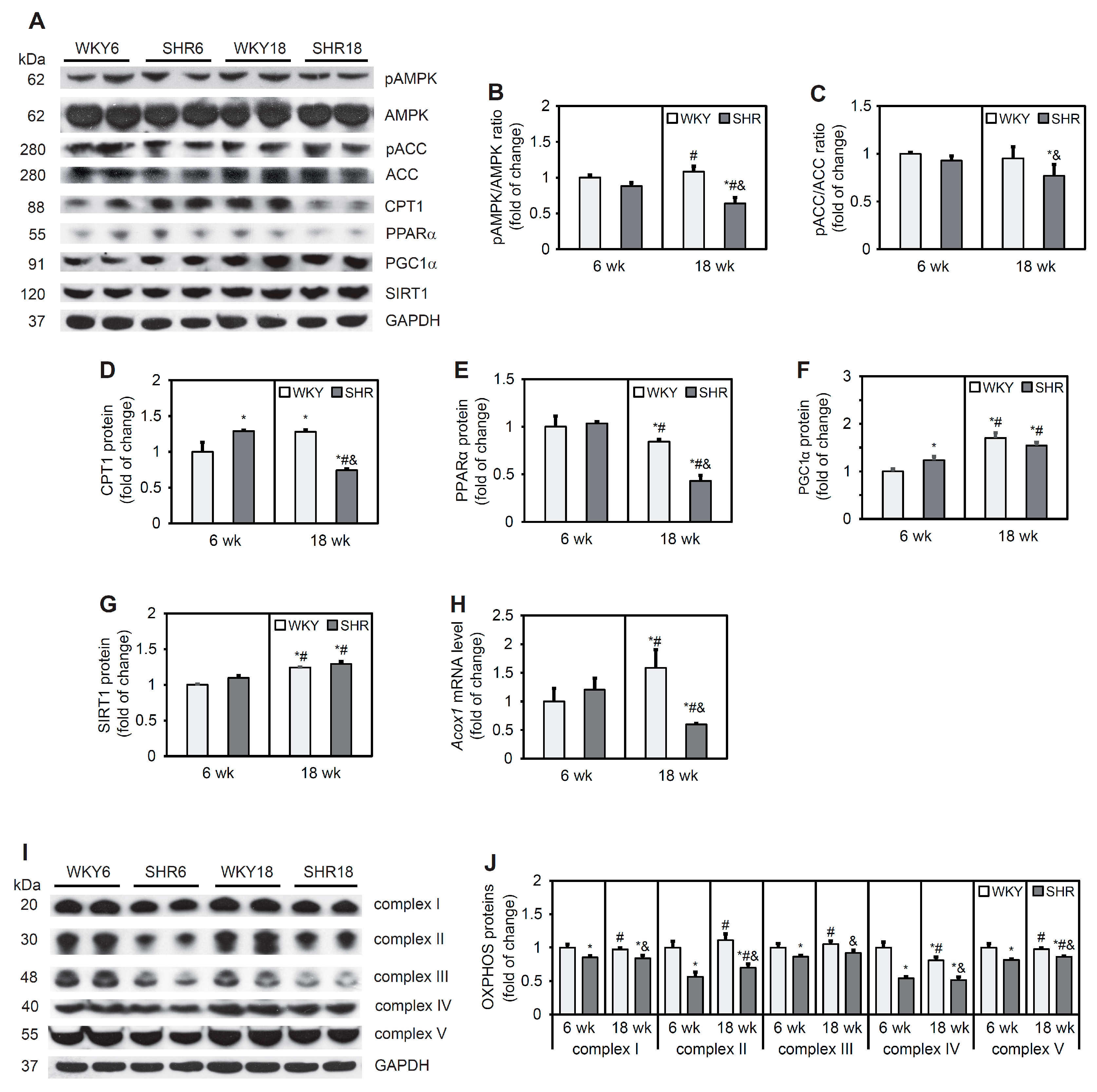
3.6. Fatty Acid β-Oxidation
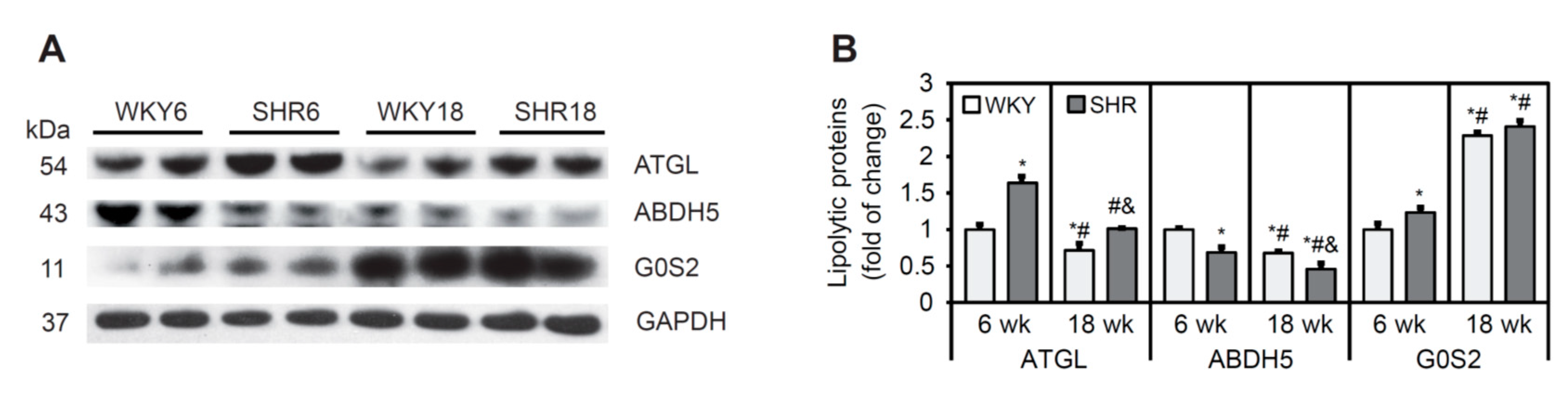
3.7. Lipolysis Process in Cardiomyocytes in SHRs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fazio, S.; Major, A.S.; Swift, L.L.; Gleaves, L.A.; Accad, M.; Linton, M.F.; Farese, R.V., Jr. Increased atherosclerosis in LDL receptor-null mice lacking ACAT1 in macrophages. J. Clin. Investig. 2001, 107, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Lopaschuk, G.D.; Karwi, Q.G.; Tian, R.; Wende, A.R.; Abel, E.D. Cardiac energy metabolism in heart failure. Circ. Res. 2021, 128, 1487–1513. [Google Scholar] [CrossRef] [PubMed]
- Pietri, P.; Georgiopoulos, G.; Tsiachris, D.; Kordalis, A.; Vlachopoulos, C.; Vyssoulis, G.; Stefanadis, C. Triglycerides are related to left ventricular mass in hypertensive patients independently of other cardiometabolic risk factors: The effect of gender. Sci. Rep. 2020, 10, 13253. [Google Scholar] [CrossRef] [PubMed]
- Sai, E.; Shimada, K.; Yokoyama, T.; Hiki, M.; Sato, S.; Hamasaki, N.; Maruyama, M.; Morimoto, R.; Miyazaki, T.; Fujimoto, S.; et al. Myocardial triglyceride content in patients with left ventricular hypertrophy: Comparison between hypertensive heart disease and hypertrophic cardiomyopathy. Heart Vessels 2017, 32, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Sanlialp, S.C.; Sanlialp, M.; Nar, G.; Malcok, A. Triglyceride glucose index reflects the unfavorable changes of left ventricular diastolic functions and structure in uncomplicated newly diagnosed hypertensive patients. Clin. Exp. Hypertens. 2022, 44, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Finck, B.N.; Han, X.; Courtois, M.; Aimond, F.; Nerbonne, J.M.; Kovacs, A.; Gross, R.W.; Kelly, D.P. A critical role for PPARalpha-mediated lipotoxicity in the pathogenesis of diabetic cardiomyopathy: Modulation by dietary fat content. Proc. Natl. Acad. Sci. USA 2003, 100, 1226–1231. [Google Scholar] [CrossRef]
- Chokshi, A.; Drosatos, K.; Cheema, F.H.; Ji, R.; Khawaja, T.; Yu, S.; Kato, T.; Khan, R.; Takayama, H.; Knöll, R.; et al. Ventricular assist device implantation corrects myocardial lipotoxicity, reverses insulin resistance, and normalizes cardiac metabolism in patients with advanced heart failure. Circulation 2012, 125, 2844–2853. [Google Scholar] [CrossRef]
- Yan, A.; Xie, G.; Ding, X.; Wang, Y.; Guo, L. Effects of lipid overload on heart in metabolic diseases. Horm. Metab. Res. 2021, 53, 771–778. [Google Scholar] [CrossRef]
- Lahey, R.; Wang, X.; Carley, A.N.; Lewandowski, E.D. Dietary fat supply to failing hearts determines dynamic lipid signaling for nuclear receptor activation and oxidation of stored triglyceride. Circulation 2014, 130, 1790–1799. [Google Scholar] [CrossRef]
- Goldberg, I.J.; Reue, K.; Abumrad, N.A.; Bickel, P.E.; Cohen, S.; Fisher, E.A.; Galis, Z.S.; Granneman, J.G.; Lewandowski, E.D.; Murphy, R.; et al. Deciphering the Role of Lipid Droplets in Cardiovascular Disease: A Report From the 2017 National Heart, Lung, and Blood Institute Workshop. Circulation 2018, 138, 305–315. [Google Scholar] [CrossRef]
- Marrocco, V.; Bogomolovas, J.; Ehler, E.; dos Remedios, C.G.; Yu, J.; Gao, C.; Lange, S. PKC and PKN in heart disease. J. Mol. Cell. Cardiol. 2019, 128, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Saponaro, C.; Gaggini, M.; Carli, F.; Gastaldelli, A. The subtle balance between lipolysis and lipogenesis: A critical point in metabolic homeostasis. Nutrients 2015, 7, 9453–9474. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.H.Y.; Lemaitre, R.N.; Imamura, F.; King, I.B.; Song, X.; Spiegelman, D.; Siscovick, D.S.; Mozaffarian, D. Fatty acids in the de novo lipogenesis pathway and risk of coronary heart disease: The Cardiovascular Health Study. Am. J. Clin. Nutr. 2011, 94, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Marfella, R.; Di Filippo, C.; Portoghese, M.; Barbieri, M.; Ferraraccio, F.; Siniscalchi, M.; Cacciapuoti, F.; Rossi, F.; D’Amico, M.; Paolisso, G. Myocardial lipid accumulation in patients with pressure-overloaded heart and metabolic syndrome. J. Lipid Res. 2009, 50, 2314–2323. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bell, M.; Sreenivasan, U.; Sreenevasan, U.; Hu, H.; Liu, J.; Dalen, K.; Londos, C.; Yamaguchi, T.; Rizzo, M.A.; et al. Unique regulation of adipose triglyceride lipase (ATGL) by perilipin 5, a lipid droplet-associated protein. J. Biol. Chem. 2011, 286, 15707–15715. [Google Scholar] [CrossRef] [PubMed]
- Sahu-Osen, A.; Montero-Moran, G.; Schittmayer, M.; Fritz, K.; Dinh, A.; Chang, Y.F.; McMahon, D.; Boeszoermenyi, A.; Cornaciu, I.; Russell, D.; et al. CGI-58/ABHD5 is phosphorylated on Ser239 by protein kinase A: Control of subcellular localization. J. Lipid Res. 2015, 56, 109–121. [Google Scholar] [CrossRef]
- Lu, X.; Yang, X.; Liu, J. Differential control of ATGL-mediated lipid droplet degradation by CGI-58 and G0S2. Cell Cycle 2010, 9, 2791–2797. [Google Scholar] [CrossRef]
- Ma, T.; Lopez-Aguiar, A.G.N.; Li, A.; Lu, Y.; Sekula, D.; Nattie, E.E.; Freemantle, S.; Dmitrovsky, E. Mice lacking G0S2 are lean and cold-tolerant. Cancer Biol. Ther. 2014, 15, 643–650. [Google Scholar] [CrossRef]
- Hajri, T.; Ibrahimi, A.; Coburn, C.T.; Knapp, F.F.; Kurtz, T.; Pravenec, M.; Abumrad, N.A. Defective fatty acid uptake in the spontaneously hypertensive rat is a primary determinant of altered glucose metabolism, hyperinsulinemia, and myocardial hypertrophy. J. Biol. Chem. 2001, 276, 23661–23666. [Google Scholar] [CrossRef]
- Dodd, M.S.; Ball, D.R.; Schroeder, M.A.; Le Page, L.M.; Atherton, H.J.; Heather, L.C.; Seymour, A.M.; Ashrafian, H.; Watkins, H.; Clarke, K.; et al. In Vivo alterations in cardiac metabolism and function in the Spontaneously Hypertensive Rat heart. Cardiovasc. Res. 2012, 95, 69–76. [Google Scholar] [CrossRef]
- LaPier, T.L.; Rodnick, K.J. Changes in cardiac energy metabolism during early development of female SHR. Am. J. Hypertens. 2000, 13, 1074–1081. [Google Scholar] [CrossRef]
- Purushothaman, S.; Nair, R.R.; Harikrishnan, V.S.; Fernandez, A.C. Temporal relation of cardiac hypertrophy, oxidative stress, and fatty acid metabolism in spontaneously hypertensive rat. Mol. Cell. Biochem. 2011, 351, 59–64. [Google Scholar] [CrossRef]
- Sambandam, N.; Chen, X.; Cam, C.; Rodrigues, B. Cardiac lipoprotein lipase in the spontaneously hypertensive rat. Cardiovasc. Res. 1997, 33, 460–468. [Google Scholar] [CrossRef]
- Saifudeen, I.; Subhadra, L.; Konnottil, R.; Nair, R.R. Metabolic modulation by edium-chain triglycerides reduces oxidative stress and ameliorates CD36-mediated cardiac remodeling in Spontaneously Hypertensive Rat in the initial and established stages of hypertrophy. J. Card. Fail. 2017, 23, 240–251. [Google Scholar] [CrossRef]
- Kondo, J.; Yamada, Y.; Okumura, K.; Hashimoto, H.; Ito, T.; Satake, T. 1,2-diacylglycerol content in myocardium from spontaneously hypertensive rats during the development of hypertension. Basic Res. Cardiol. 1990, 85, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Liška, F.; Landa, V.; Zídek, V.; Mlejnek, P.; Šilhavý, J.; Šimáková, M.; Strnad, H.; Trnovská, J.; Škop, V.; Kazdová, L.; et al. Downregulation of Plzf gene ameliorates metabolic and cardiac traits in the spontaneously hypertensive rat. Hypertension 2017, 69, 1084–1091. [Google Scholar] [CrossRef]
- Khairallah, R.J.; O’Shea, K.M.; Brown, B.H.; Khanna, N.; Des Rosiers, C.; Stanley, W.C. Treatment with docosahexaenoic acid, but not eicosapentaenoic acid, delays Ca2+-induced mitochondria permeability transition in normal and hypertrophied myocardium. J. Pharmacol. Exp. Ther. 2010, 335, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, A.; Sanchez-Rodriguez, E.; Vargas, F.; Montoro-Molina, S.; Romero, M.; Espejo-Calvo, J.A.; Vilchez, P.; Jaramillo, S.; Olmo-García, L.; Carrasco-Pancorbo, A.; et al. Cardioprotective effect of a virgin olive oil enriched with bioactive compounds in spontaneously hypertensive rats. Nutrients 2019, 11, 1728. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biol. Chem. 1959, 37, 911–917. [Google Scholar]
- Dobrzyn, A.; Dobrzyn, P.; Lee, S.H.; Miyazaki, M.; Cohen, P.; Asilmaz, E.; Hardie, D.G.; Friedman, J.M.; Ntambi, J.M. Stearoyl-CoA desaturase-1deficiency reduces ceramide synthesis by downregulating serine palmitoyltransferase and increasing beta-oxidation in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E599–E607. [Google Scholar] [CrossRef]
- Yecies, J.L.; Zhang, H.H.; Menon, S.; Liu, S.; Yecies, D.; Lipovsky, A.I.; Gorgun, C.; Kwiatkowski, D.J.; Hotamisligil, G.S.; Lee, C.H.; et al. Akt stimulates hepatic SREBP1c and lipogenesis through parallel mTORC1-dependent and independent pathways. Cell Metab. 2011, 14, 21–32. [Google Scholar] [CrossRef]
- Dillmann, W. Cardiac hypertrophy and thyroid hormone signaling. Heart Fail. Rev. 2010, 15, 125–132. [Google Scholar] [CrossRef]
- Kazantzis, M.; Stahl, A. Fatty acid transport proteins, implications in physiology and disease. Biochim. Biophys. Acta 2012, 1821, 852–857. [Google Scholar] [CrossRef]
- Shao, W.; Espenshade, P.J. Expanding roles for SREBP in metabolism. Cell Metab. 2012, 16, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.M.; Dutta, R.; Seeds, M.C.; Lake, K.N.; Hallmark, B.; Mathias, R.A.; Howard, T.D.; Chilton, F.H. FADS genetic and metabolomic analyses identify the ∆5 desaturase (FADS1) step as a critical control point in the formation of biologically important lipids. Sci. Rep. 2020, 10, 15873. [Google Scholar] [CrossRef]
- Nie, L.; Pike, A.C.W.; Pascoa, T.C.; Bushell, S.R.; Quigley, A.; Ruda, G.F.; Chu, A.; Cole, V.; Speedman, D.; Moreira, T.; et al. The structural basis of fatty acid elongation by the ELOVL elongases. Nat. Struct. Mol. Biol. 2021, 28, 512–520. [Google Scholar] [CrossRef]
- Lehman, J.J.; Barger, P.M.; Kovacs, A.; Saffitz, J.E.; Medeiros, D.M.; Kelly, D.P. Peroxisome proliferator-activated receptor gamma coactivator-1 promotes cardiac mitochondrial biogenesis. J. Clin. Investig. 2000, 106, 847–856. [Google Scholar] [CrossRef]
- Heier, C.; Radner, F.P.; Moustafa, T.; Schreiber, R.; Grond, S.; Eichmann, T.O.; Schweiger, M.; Schmidt, A.; Cerk, I.K.; Oberer, M.; et al. G0/G1 switch Gene 2 regulates cardiac lipolysis. J. Biol. Chem. 2015, 290, 26141–26150. [Google Scholar] [CrossRef]
- Zhou, Y.T.; Grayburn, P.; Karim, A.; Shimabukuro, M.; Higa, M.; Baetens, D.; Orci, L.; Unger, R.H. Lipotoxic heart disease in obese rats: Implications for human obesity. Proc. Natl. Acad. Sci. USA 2000, 97, 1784–1789. [Google Scholar] [CrossRef] [PubMed]
- Koehler, U.; Medugorac, I. Left ventricular enzyme activities of the energy-supplying metabolism in Goldblatt-II rats. Res. Exp. Med. 1985, 185, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Raizada, V.; Pathak, D.; Avery, G.; Woodfin, B. Accelerated glycolysis in early hypertensive left ventricular hypertrophy. Cardiology 1993, 83, 160–164. [Google Scholar] [CrossRef]
- Park, H.J.; Georgescu, S.P.; Du, C.; Madias, C.; Aronovitz, M.J.; Welzig, C.M.; Wang, B.; Begley, U.; Zhang, Y.; Blaustein, R.O.; et al. Parasympathetic response in chick myocytes and mouse heart is controlled by SREBP. J. Clin. Investig. 2008, 118, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.; Kotzka, J.; Kremer, L.; Lehr, S.; Lohaus, C.; Meyer, H.E.; Krone, W.; Müller-Wieland, D. MAP kinases Erk1/2 phosphorylate sterol regulatory element-binding protein (SREBP)-1a at serine 117 in vitro. J. Biol. Chem. 2000, 275, 33302–33307. [Google Scholar] [CrossRef]
- Soesanto, W.; Lin, H.Y.; Hu, E.; Lefler, S.; Litwin, S.E.; Sena, S.; Abel, E.D.; Symons, J.D.; Jalili, T. Mammalian target of rapamycin is a critical regulator of cardiac hypertrophy in spontaneously hypertensive rats. Hypertension 2009, 54, 1321–1327. [Google Scholar] [CrossRef] [PubMed]
- Aceros, H.; Farah, G.; Cobos-Puc, L.; Stabile, A.M.; Noiseux, N.; Mukaddam-Daher, S. Moxonidine improves cardiac structure and performance in SHR through inhibition of cytokines, p38 MAPK and Akt. Br. J. Pharmacol. 2011, 164, 946–957. [Google Scholar] [CrossRef]
- Wang, Y.; Viscarra, J.; Kim, S.J.; Sul, H.S. Transcriptional regulation of hepatic lipogenesis. Nat. Rev. Mol. Cell. Biol. 2015, 16, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Raso, G.M.; Bianco, G.; Iacono, A.; Esposito, E.; Autore, G.; Ferrante, M.C.; Calignano, A.; Meli, R. Maternal adaptations to pregnancy in spontaneously hypertensive rats: Leptin and ghrelin evaluation. J. Endocrinol. 2007, 194, 611–619. [Google Scholar] [CrossRef]
- Hall, M.E.; Maready, M.W.; Hall, J.E.; Stec, D.E. Rescue of cardiac leptin receptors in db/db mice prevents myocardial triglyceride accumulation. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E316–E325. [Google Scholar] [CrossRef]
- Wang, D.; Feng, D.; Wang, Y.; Dong, P.; Wang, Y.; Zhong, L.; Li, B.; Fu, J.; Xiao, X.; Speakman, J.R.; et al. Angiopoietin-like protein 8/leptin crosstalk influences cardiac mass in youths with cardiometabolic risk: The BCAMS study. Front. Endocrinol. 2022, 12, 788549. [Google Scholar] [CrossRef]
- Morelli, M.B.; Chavez, C.; Santulli, G. Angiopoietin-like proteins as therapeutic targets for cardiovascular disease: Focus on lipid disorders. Expert Opin. Ther. Targets 2020, 24, 79–88. [Google Scholar] [CrossRef]
- Aitman, T.J.; Glazier, A.M.; Wallace, C.A.; Cooper, L.D.; Norsworthy, P.J.; Wahid, F.N.; Al-Majali, K.M.; Trembling, P.M.; Mann, C.J.; Shoulders, C.C.; et al. Identification of Cd36 (Fat) as an insulin-resistance gene causing defective fatty acid and glucose metabolism in hypertensive rats. Nat. Genet. 1999, 21, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Jump, D.B.; Tripathy, S.; Depner, C.M. Fatty acid-regulated transcription factors in the liver. Annu. Rev. Nutr. 2013, 33, 249–269. [Google Scholar] [CrossRef] [PubMed]
- Lovic, D.; Erdine, S.; Catakoğlu, A.B. How to estimate left ventricular hypertrophy in hypertensive patients. Anatol. J. Cardiol. 2014, 14, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Shi, X.; Bharadwaj, K.G.; Ikeda, S.; Yamashita, H.; Yagyu, H.; Schaffer, J.E.; Yu, Y.H.; Goldberg, I.J. DGAT1 expression increases heart triglyceride content but ameliorates lipotoxicity. J. Biol. Chem. 2009, 284, 36312–36323. [Google Scholar] [CrossRef] [PubMed]
- Roe, N.D.; Handzlik, M.K.; Li, T.; Tian, R. The Role of Diacylglycerol Acyltransferase (DGAT) 1 and 2 in Cardiac Metabolism and Function. Sci. Rep. 2018, 8, 4983. [Google Scholar]
- Ardiansyah; Ohsaki, Y.; Shirakawa, H.; Koseki, T.; Komai, M. Novel effects of a single administration of ferulic acid on the regulation of blood pressure and the hepatic lipid metabolic profile in stroke-prone spontaneously hypertensive rats. J. Agric. Food Chem. 2008, 56, 2825–2830. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Minćzuk, K.; Massey, J.C.; Howell, N.L.; Roy, R.J.; Paul, S.; Patrie, J.T.; Kramer, C.M.; Epstein, F.H.; Carey, R.M.; et al. Metformin improves cardiac metabolism and function, and prevents left ventricular hypertrophy in spontaneously hypertensive rats. J. Am. Heart Assoc. 2020, 9, e015154. [Google Scholar] [CrossRef]
- Rubattu, S.; Stanzione, R.; Volpe, M. Mitochondrial dysfunction contributes to hypertensive target organ damage: Lessons from an animal model of human disease. Oxid. Med. Cell Longev. 2016, 2016, 1067801. [Google Scholar] [CrossRef]
- Haemmerle, G.; Lass, A.; Zimmermann, R.; Gorkiewicz, G.; Meyer, C.; Rozman, J.; Heldmaier, G.; Maier, R.; Theussl, C.; Eder, S.; et al. Defective lipolysis and altered energy metabolism in mice lacking adipose triglyceride lipase. Science 2006, 312, 734–737. [Google Scholar] [CrossRef]
- Gao, H.; Feng, X.; Li, Z.; Li, M.; Gao, S.; He, Y.; Wang, J.; Zeng, S.; Liu, X.; Huang, X.; et al. Downregulation of adipose triglyceride lipase promotes cardiomyocyte hypertrophy by triggering the accumulation of ceramides. Arch. Biochem. Biophys. 2015, 565, 76–88. [Google Scholar] [CrossRef]
- Kienesberger, P.C.; Pulinilkunnil, T.; Sung, M.M.Y.; Nagendran, J.; Haemmerle, G.; Kershaw, E.E.; Young, M.E.; Light, P.E.; Oudit, G.Y.; Zechner, R.; et al. Myocardial ATGL overexpression decreases the reliance on fatty acid oxidation and protects against pressure overload-induced cardiac dysfunction. Mol. Cell. Biol. 2012, 32, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Pulinilkunnil, T.; Kienesberger, P.C.; Nagendran, J.; Sharma, N.; Young, M.E.; Dyck, J.R.B. Cardiac-specific adipose triglyceride lipase overexpression protects from cardiac steatosis and dilated cardiomyopathy following diet-induced obesity. Int. J. Obes. 2014, 38, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Zierler, K.A.; Jaeger, D.; Pollak, N.M.; Eder, S.; Rechberger, G.N.; Radner, F.P.W.; Woelkart, G.; Kolb, D.; Schmidt, A.; Kumari, M.; et al. Functional cardiac lipolysis in mice critically depends on comparative gene identification-58. J. Biol. Chem. 2013, 288, 9892–9904. [Google Scholar] [CrossRef]
- Xie, X.; Tie, Y.F.; Lai, S.; Zhang, Y.L.; Li, H.H.; Liu, Y. Cardiac-specific CGI-58 deficiency activates the ER stress pathway to promote heart failure in mice. Cell Death Dis. 2021, 12, 1003. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WKY6 | SHR6 | WKY18 | SHR18 | |
|---|---|---|---|---|
| Glucose (mg/dL) | 182.9 ± 23.0 | 109.2 ± 19.9 * | 213.9 ± 36.1 | 152.1 ± 24.1 & |
| FFA (mg/dL) | 4.9 ± 0.5 | 16.4 ± 5.5 * | 9.7 ± 1.3 | 10.1 ± 2.0 |
| TG (mg/dL) | 73.7 ± 9.2 | 104.9 ± 23.2 * | 86.3 ± 13.0 | 140.3 ± 24.6 & |
| Cholesterol (mg/dL) | 57.1 ± 5.0 | 66.6 ± 7.8 * | 69.9 ± 11.3 | 37.8 ± 5.1 & |
| WKY6 | SHR6 | WKY18 | SHR18 | |
|---|---|---|---|---|
| HW/BW (g/g × 100) | 0.38 ± 0.06 | 0.47 ± 0.07 * | 0.29 ± 0.05 | 0.41 ± 0.03 & |
| HR (beats/min) | 312 ± 17 | 404 ± 26 * | 351 ± 4 | 381 ± 26 & |
| AWTd (mm) | 1.14 ± 0.13 | 1.19 ± 0.11 | 1.60 ± 0.08 | 1.97 ± 0.14 & |
| PWTd (mm) | 1.24 ± 0.13 | 1.31 ± 0.11 | 1.68 ± 0.05 | 2.15 ± 0.15 & |
| AWTs (mm) | 2.46 ± 0.16 | 2.54 ± 0.21 | 3.18 ± 0.10 | 3.52 ± 0.26 & |
| PWTs (mm) | 2.73 ± 0.21 | 2.72 ± 0.16 | 3.33 ± 0.10 | 3.98 ± 0.20 & |
| EDD (mm) | 5.03 ± 0.60 | 4.79 ± 0.38 | 7.00 ± 0.24 | 6.83 ± 0.44 |
| ESD (mm) | 2.09 ± 0.37 | 2.10 ± 0.26 | 3.30 ± 0.26 | 3.83 ± 0.25 & |
| RWT | 0.50 ± 0.02 | 0.55 ± 0.03 * | 0.48 ± 0.01 | 0.63 ± 0.03 & |
| EDV (mL) | 0.14 ± 0.05 | 0.12 ± 0.03 | 0.36 ± 0.04 | 0.34 ± 0.07 |
| ESV (mL) | 0.010 ± 0.006 | 0.010 ± 0.004 | 0.038 ± 0.010 | 0.06 ± 0.012 & |
| EF (%) | 92.5 ± 2.5 | 91.5 ± 1.8 | 89.5 ± 1.5 | 82.3 ± 1.3 & |
| FS (%) | 058 ± 0.05 | 0.56 ± 0.03 | 0.53 ± 0.02 | 0.44 ± 0.01 & |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bednarski, T.K.; Duda, M.K.; Dobrzyn, P. Alterations of Lipid Metabolism in the Heart in Spontaneously Hypertensive Rats Precedes Left Ventricular Hypertrophy and Cardiac Dysfunction. Cells 2022, 11, 3032. https://doi.org/10.3390/cells11193032
Bednarski TK, Duda MK, Dobrzyn P. Alterations of Lipid Metabolism in the Heart in Spontaneously Hypertensive Rats Precedes Left Ventricular Hypertrophy and Cardiac Dysfunction. Cells. 2022; 11(19):3032. https://doi.org/10.3390/cells11193032
Chicago/Turabian StyleBednarski, Tomasz K., Monika K. Duda, and Pawel Dobrzyn. 2022. "Alterations of Lipid Metabolism in the Heart in Spontaneously Hypertensive Rats Precedes Left Ventricular Hypertrophy and Cardiac Dysfunction" Cells 11, no. 19: 3032. https://doi.org/10.3390/cells11193032
APA StyleBednarski, T. K., Duda, M. K., & Dobrzyn, P. (2022). Alterations of Lipid Metabolism in the Heart in Spontaneously Hypertensive Rats Precedes Left Ventricular Hypertrophy and Cardiac Dysfunction. Cells, 11(19), 3032. https://doi.org/10.3390/cells11193032