1. Introduction
The
IDH1 gene encodes NADP
+-dependent IDH localized in the cytoplasm and peroxisomes [
1,
2,
3,
4,
5]. This enzyme not only catalyzes the oxidative decarboxylation of isocitrate to 2-oxoglutarate but also is critical to reductive carboxylation, which is required for lipogenesis in hypoxia and redox homeostasis during anchorage-independent growth [
6,
7]. In agreement with its physiological function of regulating the intracellular NADP
+/NADPH ratio [
8], IDH1 plays an important role in metabolic adaption in physiology and cancer biology.
Earlier studies revealed widespread
IDH1 mutations at Arg132, an active site of the enzyme, in WHO grade II and III (referred to as lower-grade) glioma and in WHO grade IV secondary glioblastoma [
9,
10,
11]. Among these hotspot mutations, the
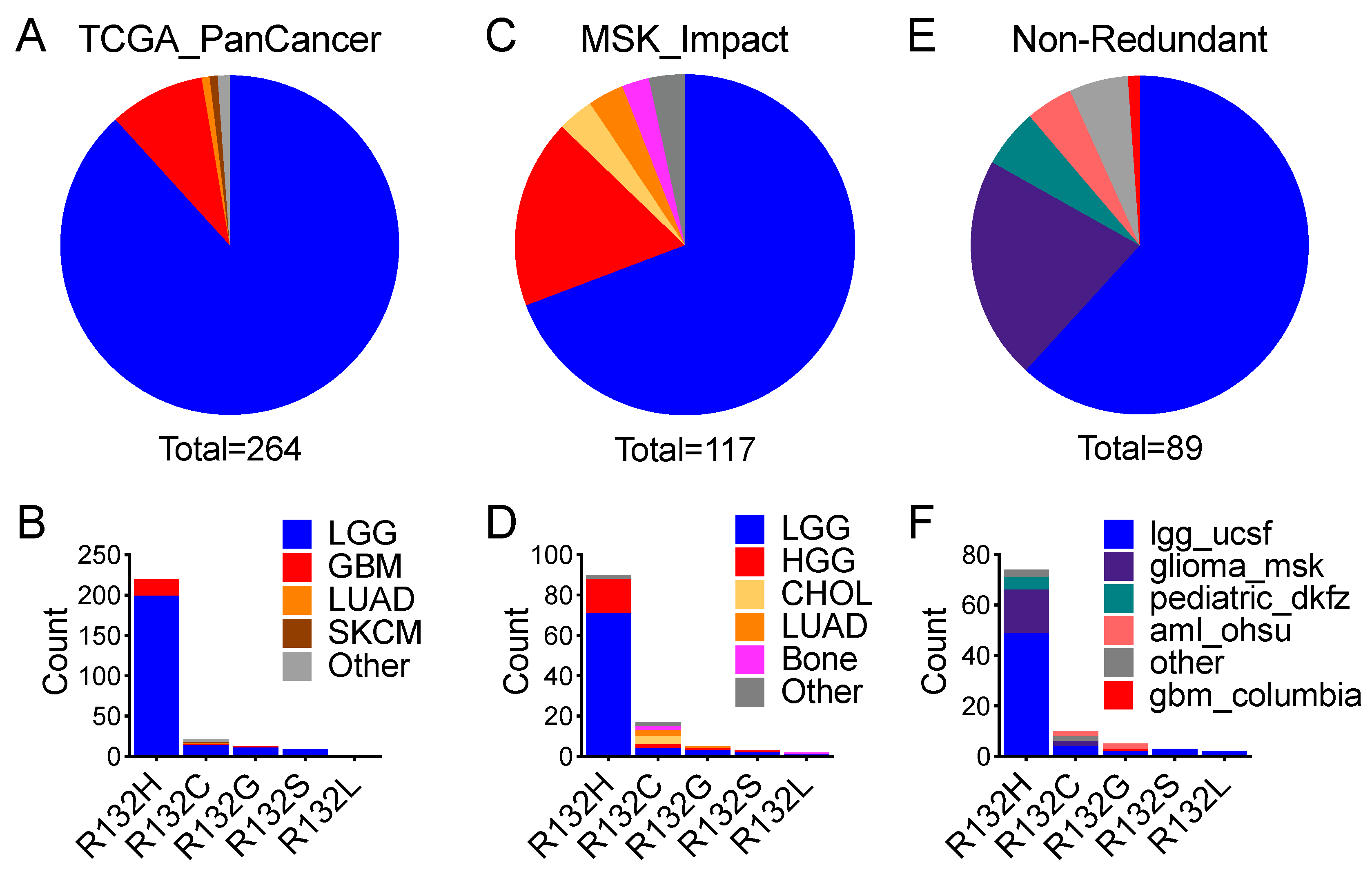
IDH1R132H frequency was >90%, whereas non-canonical
IDH1 mutations, including
IDH1R132C,
IDH1R132G,
IDH1R132S, and
IDH1R132L (referred to collectively as
IDH1R132X), were at very low frequencies [
12]. Furthermore, mutations in the
IDH2 gene (encoding a mitochondrial NADP
+-dependent enzyme) at the analogous Arg172 were relatively uncommon in lower-grade glioma and non-existent in glioblastoma [
13]. Although they were thought to be virtually exclusive in glioma [
11,
14], further mutational analyses revealed
IDH1 and
IDH2 mutations, as well as
IDH2 Arg140 mutations, in various cancer types such as myeloid neoplasia, chondrosarcoma, cholangiocarcinoma, and prostate cancer [
2,
15].
IDH1 and
IDH2 mutations acquire a neomorphic function that produces (D)-2-hydroxyglutarate (D-2HG) from the NADPH-dependent reduction of 2-oxoglutarate [
16]. High levels of D-2HG induce histone and DNA hypermethylation through competitive inhibition of histone and DNA demethylases, thereby blocking cell differentiation and promoting neural stem-like phenotype [
17,
18,
19]; however, neural stem-cell marker genes such as
NES and
PROM1 are generally downregulated in IDH-mutant glioma [
20]. The finding that
IDH1 and
IDH2 mutations occur in various other cancer types has spurred further interest in cancer metabolism, epigenetic regulation, and therapeutic targeting, also promoting the idea that these mutations drive tumorigenesis in general [
1,
2,
3,
4,
5,
21], despite how
IDH1 and
IDH2 mutations promote gliomagenesis remains unclear [
13,
22].
Genetically, lower-grade gliomas with
IDH1 and
IDH2 mutations are associated with either
TP53 and/or
ATRX alteration or 1p/19q codeletion [
23,
24]. Although
TP53 is among the most-mutated tumor-suppressor genes in human cancer [
25,
26,
27], the biological significance of
TP53 alteration in lower-grade glioma requires further investigation. In this study, we analyzed three independent pan-cancer datasets and revealed that the association of
IDH1 mutation with
TP53 alteration is specific to glioma, which indicates a tissue-specific role for
TP53 alteration in gliomagenesis.
2. Materials and Methods
2.1. Pan-Cancer Datasets
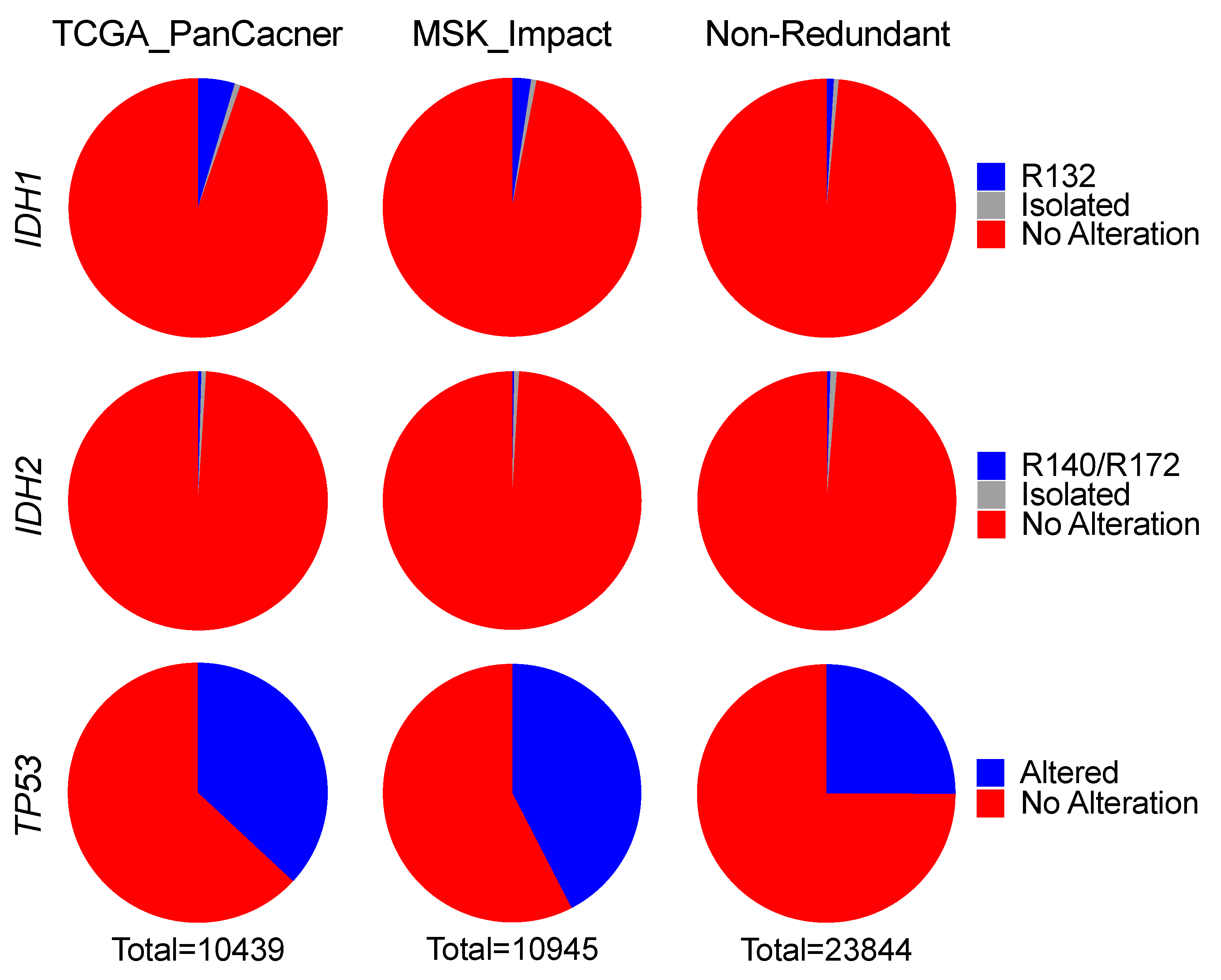
Three independent pan-cancer datasets: TCGA PanCancer dataset (TCGA_PanCancer); MSK-Impact pan-cancer dataset (MSK_Impact); and a combined, non-redundant pan-cancer dataset (Non-Redundant), were downloaded from cBioPortal [
28,
29]. Downloaded data included study ID, sample ID, patient ID, and patient status and survival, with matched genetic alteration data of
IDH1,
IDH2,
TP53,
CDKN2A,
CDKN2B,
CIC,
FUBP1, and 1p/19q codeletion.
TCGA_PanCancer consists of 32 studies comprising 10,953 patients/10,967 samples from cancer types including bladder urothelial carcinoma (BLCA, n = 411), cholangiocarcinoma (CHOL, n = 36), colorectal adenocarcinoma (COADREAD, n = 594), breast invasive carcinoma (BRCA, n = 1084), brain lower-grade glioma (LGG, n = 514), glioblastoma (GBM, n = 592), esophageal adenocarcinoma (ESCA, n = 182), stomach adenocarcinoma (STAD, n = 440), head and neck squamous cell carcinoma (HNSC, n = 523), liver hepatocellular carcinoma (LIHC, n = 372), lung adenocarcinoma (LUAD, n = 566), lung squamous cell carcinoma (LUSC, n = 487), acute myeloid leukemia (LAML, n = 200), ovarian serous cystadenocarcinoma (OV, n = 585), pancreatic adenocarcinoma (PAAD, n = 184), skin cutaneous melanoma (SKCM, n = 448), sarcoma (SARC, n = 255), and uterine corpus endometrial carcinoma (UCEC, n = 529).
MSK_Impact consists of 10,336 patients/10,945 profiled samples, including glioma (n = 553), hepatobiliary cancer (Hepatobiliary, n = 355), bone cancer (Bone, n = 135), skin cancer, non-melanoma (SKNM, n = 148), small cell lung cancer (SCLC, n = 82), melanoma (n = 365), mature T and NK neoplasms (Mature T and NK, n = 134), uterine sarcoma (USARC, n = 93), small bowel cancer (Small Bowel, n = 35), and central nervous system cancer (CNS, n = 48).
Non-Redundant consists of 152 studies published from numerous institutions comprising 25,016 patients/26,922 samples with various cancer types, including acute myeloid leukemia or myelodysplastic syndromes (mnm), skin cutaneous melanoma (skcm), metastatic melanoma (mel), uterine carcinoma (ucs), cutaneous squamous cell carcinoma (cscc), primary central nervous system lymphoma (pcnsl), esophageal squamous cell carcinoma (escc), esophageal adenocarcinoma (esca), ampullary carcinoma (ampca), and basal cell carcinoma (bcc).
2.2. Data Analysis
Relevant data were extracted and processed with GraphPad Prism version 9.0 software (GraphPad Software, San Diego, CA, USA) to present genetic alteration frequency and occurrence. Statistical significance in frequency difference was determined by paired
t-test or Fisher’s exact test, as specified, with two-tailed
p-values. Overall survival was analyzed by the Mantel-Cox log-rank test as previously described [
30,
31]. Multivariate Cox proportional hazards analysis was performed with SPSS Statistics (IBM) software by including
TP53 status,
IDH1 status, age, sex, and histological type, as previously described [
31].
4. Discussion
Through a survey of more than 45,000 pan-cancer samples, we observed that
IDH1 and
IDH2 hotspot mutations are uncommon (2%) and extremely rare (0.4%), respectively, in human cancer, a finding in agreement with an independent pan-cancer analysis [
37]. Therefore, despite being prevalent in glioma, as reported previously [
10,
11,
14,
38], these mutations appear to be selected against in tumorigenesis, which is seemingly at odds with the general thought that these mutations induce oncogenic transformation through epigenetic and metabolic reprogramming resulting from high levels of D-2HG [
13].
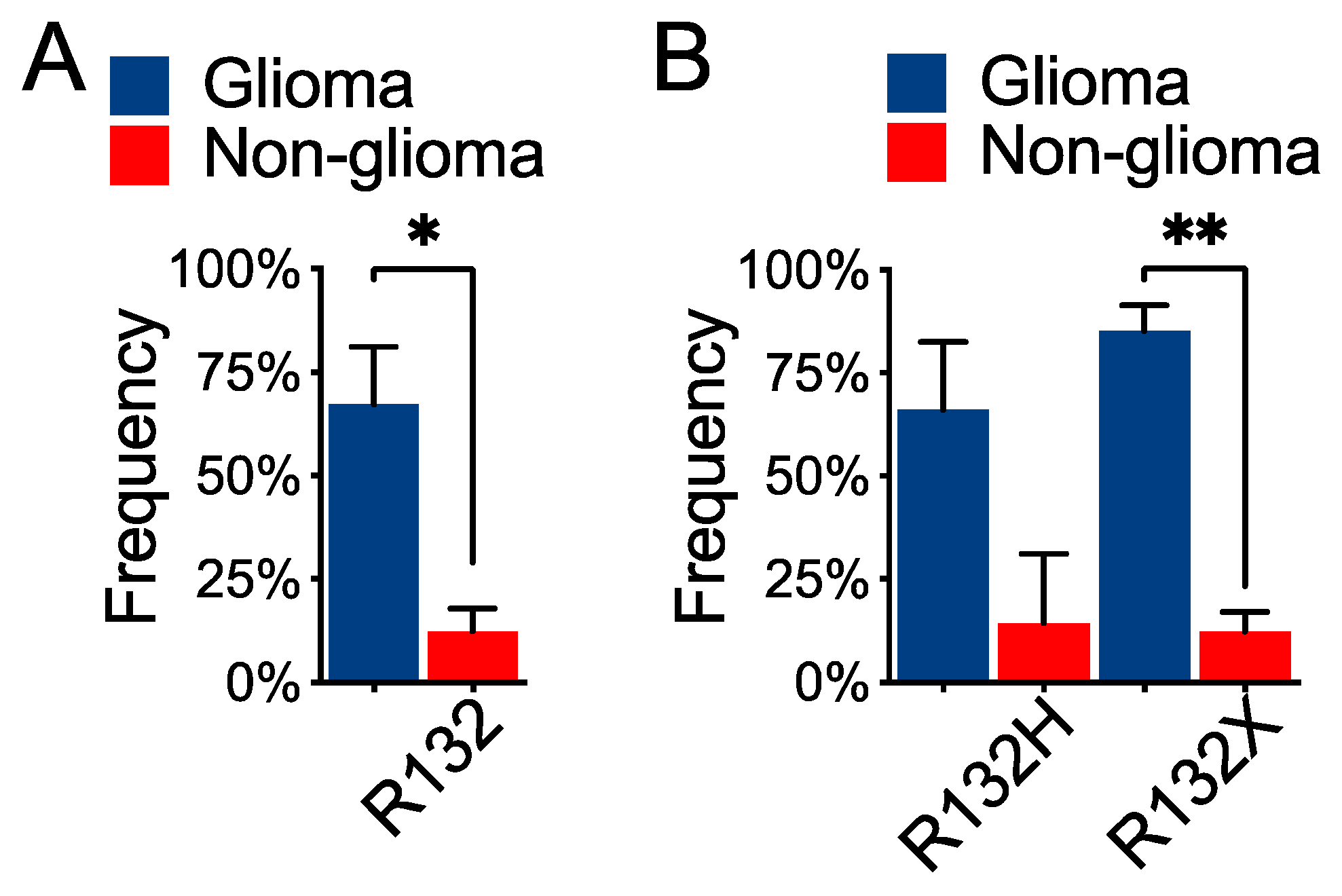
IDH1R132H, the most common form in glioma, produces the least amount of D-2HG and correlates with worse survival compared with the rare
IDH1R132X and
IDH2-R172 mutations, which produce higher levels of D-2HG [
12,
33,
39]. Although the rare occurrence of these mutations in glioma has been attributable to the “cytotoxicity” of high levels of D-2HG [
12,
33], D-2HG sensitizes cells to ferroptosis [
40]—an iron-dependent form of nonapoptotic cell death likely involved in tumor suppression [
41]. D-2HG also exhibits tumor-suppressive activities through the inhibition of aerobic glycolysis in both IDH-mutant and IDH-wildtype leukemia cells [
42]. Together with our previous studies showing that
IDH1 hotspot mutations are intrinsically tumor suppressive [
30,
43,
44], these findings may provide an explanation for the rare occurrence of
IDH1 and
IDH2 hotspot mutations in human cancer.
The prevalence of
IDH1 hotspot mutation in glioma and its co-occurrence with
TP53 alteration indicate a tissue-specific role in gliomagenesis [
13]. Tissue specificity in cancer is best evidenced by hereditary cancer predisposition syndromes in which the underlying gene defects, such as mutations in
APC,
BRCA1, and
VHL, are associated with a high risk of developing tissue-specific cancer types [
45]. In nonhereditary cancers, a subset of recurring genetic alterations can be identified to be associated with a particular type of cancer [
45,
46]. What drives tissue specificity in cancer, however, is complex even though numerous possibilities, including cell of origin, heterogeneity, epigenetic state, and environment, have been proposed [
45,
46]. In keeping with this, studies have shown the requirement of
Trp53 knockout/down to recapitulate a less aggressive phenotype of
IDH1R132H glioma compared with
IDH1-wildtype glioma [
47,
48,
49]; however, the mechanism by which
TP53 alteration contributes to gliomagenesis remains unclear.
Interestingly, p53-mediated ferroptosis, a novel function of p53, has been implicated in tumor suppression independent of its previously recognized tumor-suppressive activities in cell cycle, apoptosis, and senescence [
50,
51,
52,
53,
54]. Given the strongest display of ferroptosis-sensitive gene signature in IDH-mutant lower-grade glioma among all cancer types [
55], we speculate that
TP53 alteration is required to inhibit ferroptosis for gliomagenesis, especially for
IDH1R132X gliomas, including oligodendroglioma, that are supersensitive to ferroptosis owing to the higher levels of D-2HG. For
IDH1R132H gliomas that are relatively less sensitive to ferroptosis, alternative tumor-suppressor pathways, such as 1p/19q codeletion, must be inactivated. Furthermore, our previous studies suggested the importance of the glutamate-rich cerebral environment in IDH-mutant lower-grade gliomagenesis [
44,
56], in agreement with the role of environment for tissue specificity in cancer. Therefore, the requirement of
TP53 alteration and a glutamate-rich environment in gliomagenesis warrants further investigation to account for the prevalence of
IDH1 hotspot mutations in glioma.
The tendency of mutual exclusivity between
IDH2 and
TP53 alteration in human cancer, including glioma, is intriguing, which may suggest alternative mechanisms of tumor-suppressor gene inactivation in tumorigenesis. Given the higher levels of D-2HG and its association with better survival [
33,
39], understanding how
IDH2-R172 glioma cells overcome D-2HG induced sensitization to ferroptosis will shed light on the mechanism of
IDH2-mutant gliomagenesis and the rare occurrence of such a mutation in human cancer.
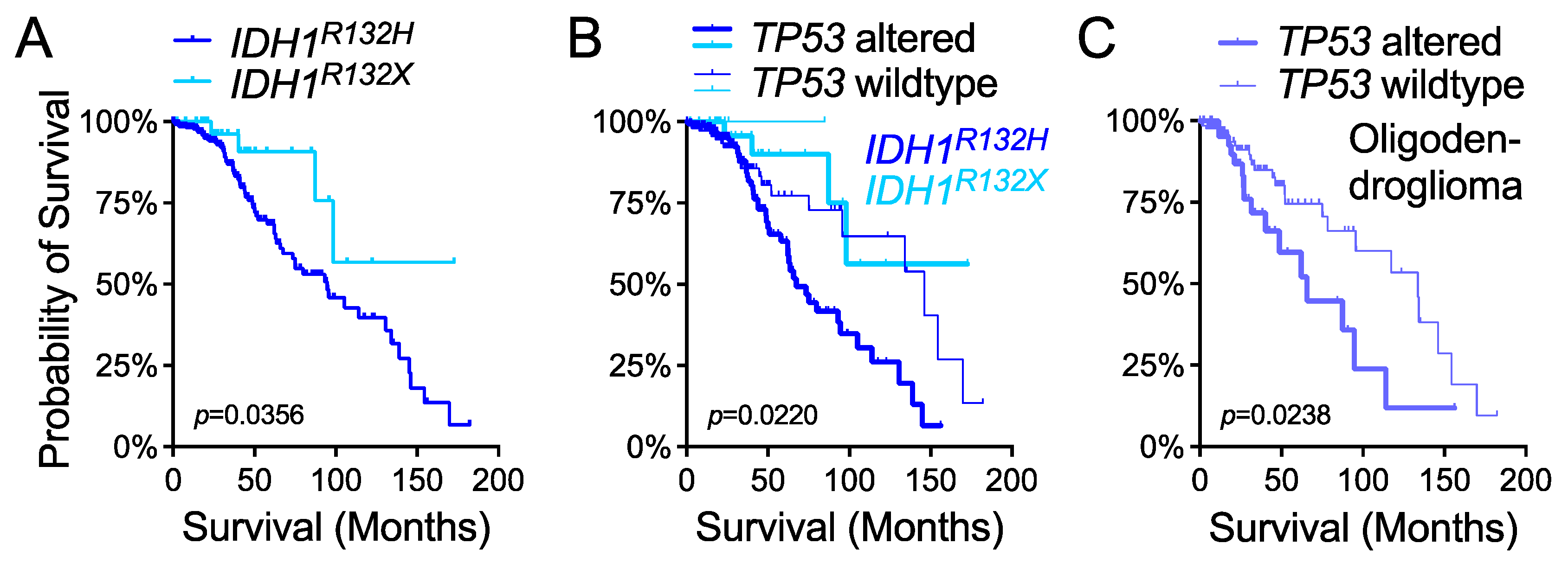
Although the p53 tumor-suppressor pathway is altered at the frequency of 87% in glioblastoma [
57], previously, the
TP53 status has not been associated with patient survival outcomes despite the well-established association of
TP53 alteration with IDH mutations in glioma [
23,
24,
58,
59]. Interestingly,
TP53 alteration has been associated with poor outcomes in pediatric H3 K27M-mutant glioma [
60]. Likewise, in lower-grade glioma we provided evidence that
TP53 status is an independent predictor of overall survival in various molecular and histological subgroups, including the
IDH1R132X subgroup, which was recently reported to have better outcomes than the
IDH1R132H subgroup [
33]. Interestingly, we observed similar overall survival between the
TP53-wildtype
IDH1R132H subgroup and the
TP53-altered
IDH1R132X subgroup based on the available size of samples; however, the paramount importance of
TP53 status in
IDH1R132X glioma requires validation with independent cohorts. Similarly, the importance of
TP53 status in oligodendroglioma patient survival also requires validation. Moreover, additional genetic events, such as
ATRX,
TERT, and
BRAF, which also frequently occur in IDH-mutant glioma [
24,
61,
62], should be considered in future investigations.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}