
Neuroglobin: A New Possible Marker of Estrogen-Responsive Breast Cancer


 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Human Subjects
2.3. Breast Cancer Tissues
2.4. Breast Tissue Immunohistochemistry
2.5. Protein Extraction and Western Blot Assay
2.6. Mitochondria Isolation
2.7. Statistical Analysis
3. Results
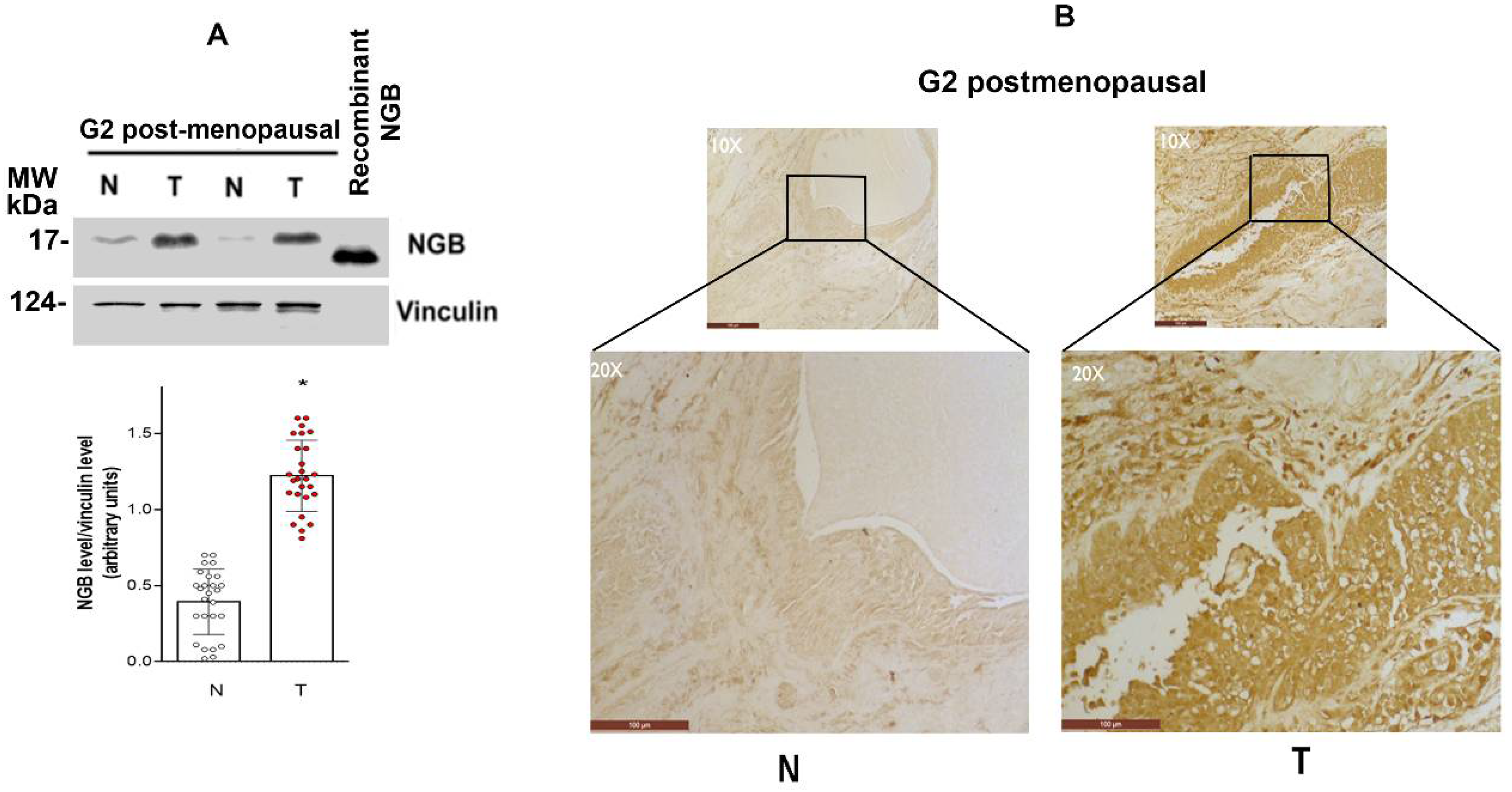
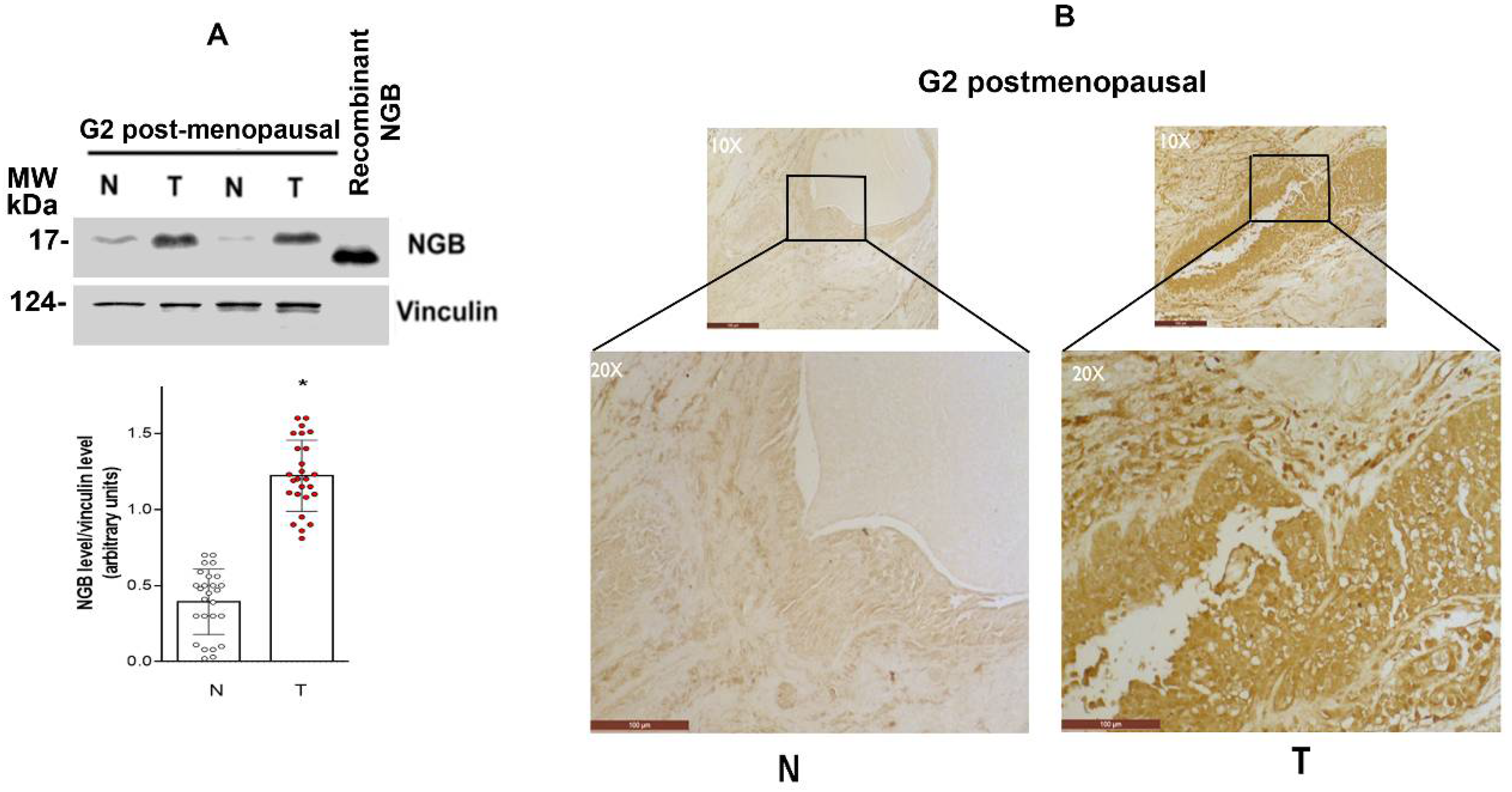
3.1. NGB Expression in Breast Tumors
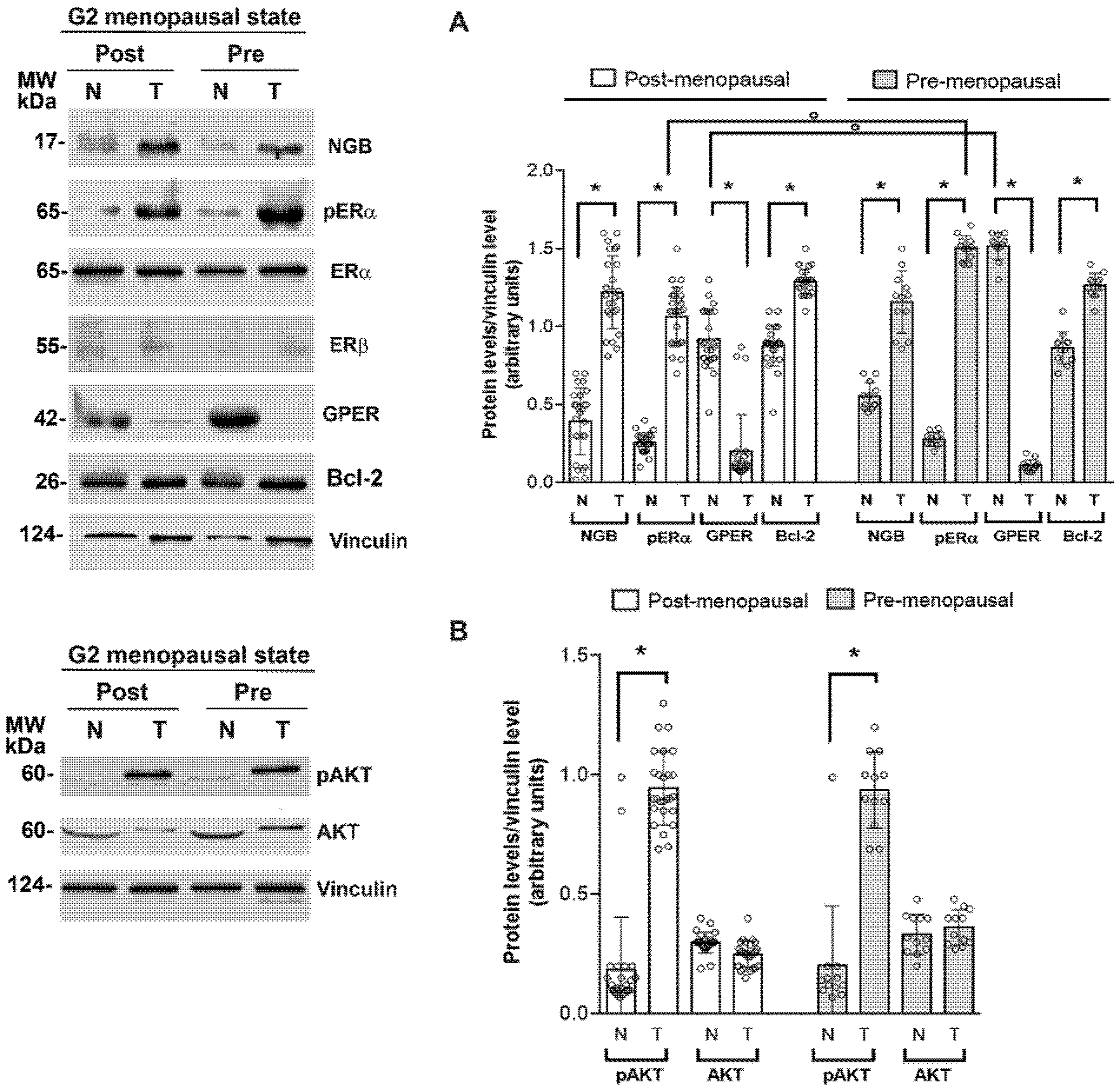
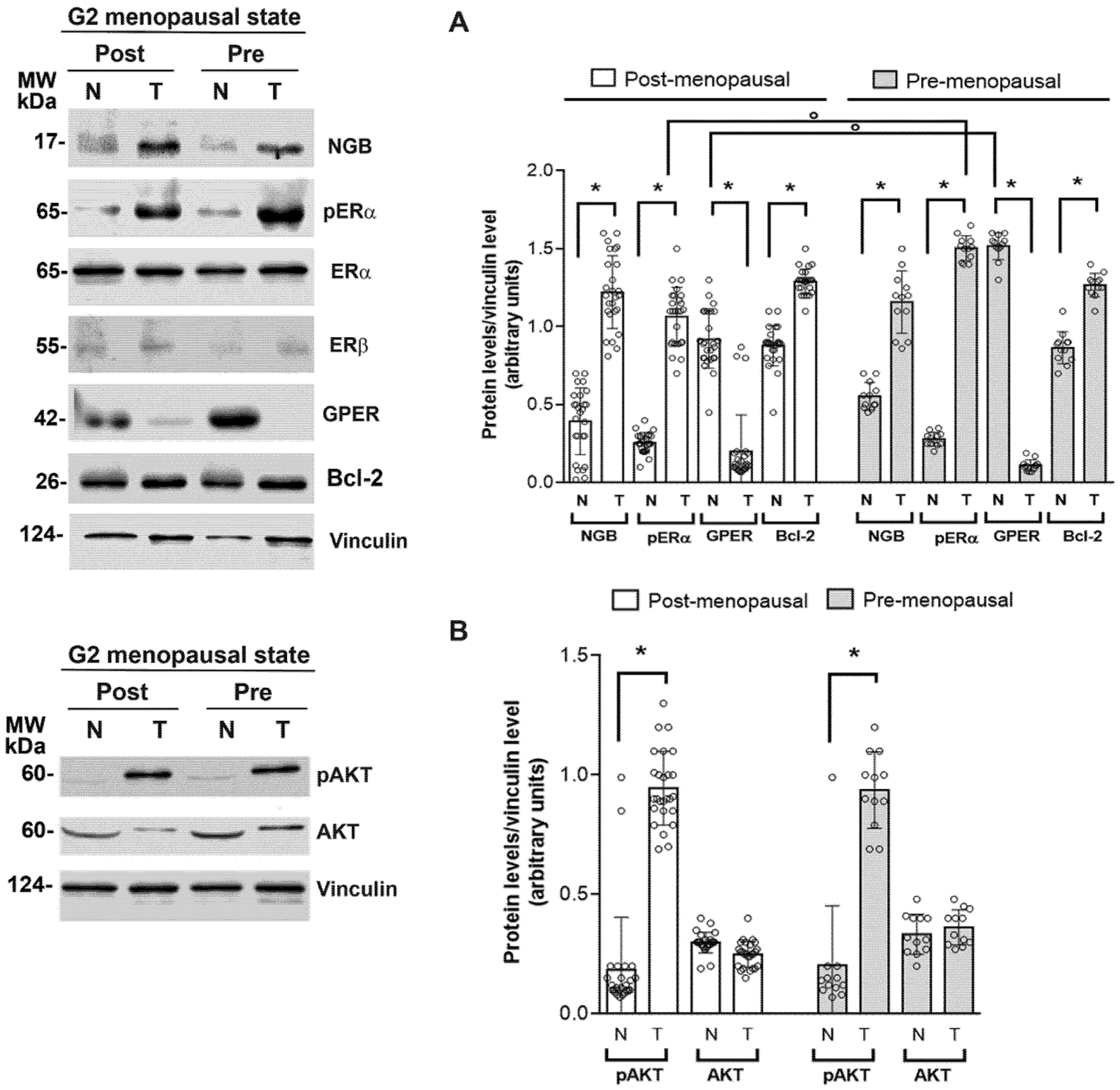
3.2. Association between NGB and Estrogen Receptor Levels and Activities in Breast Tumors
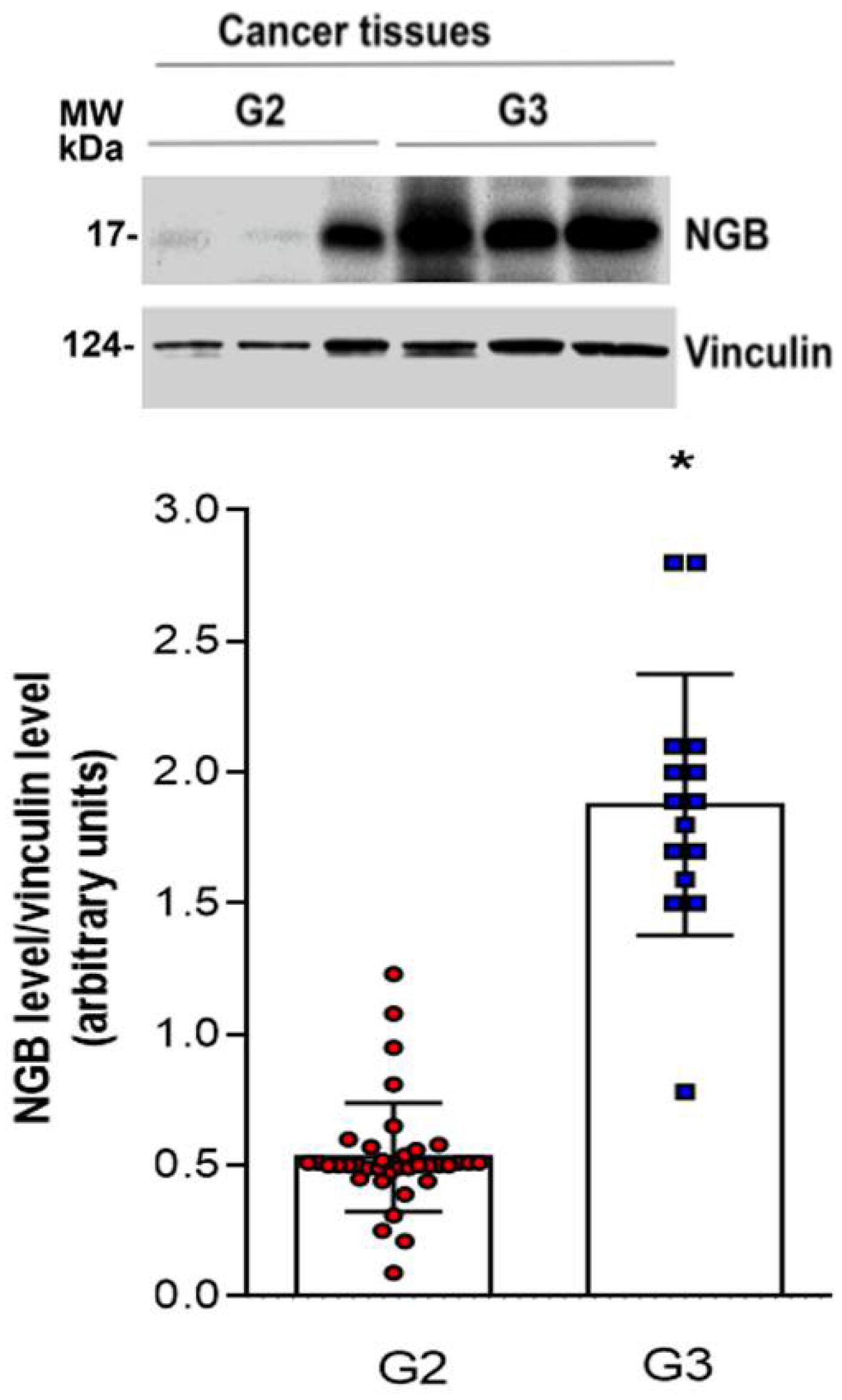
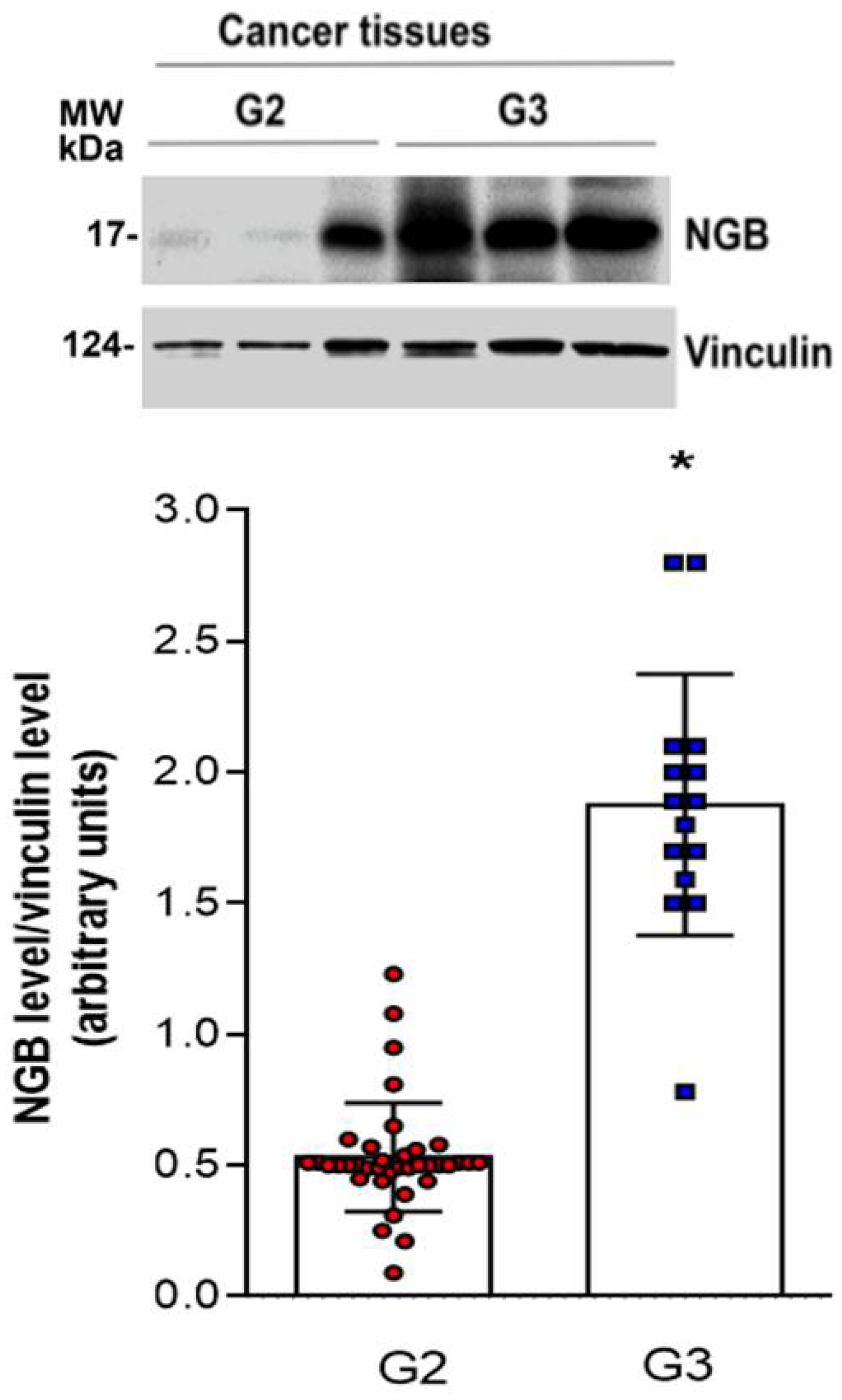
3.3. NGB Association with Tumor Grade
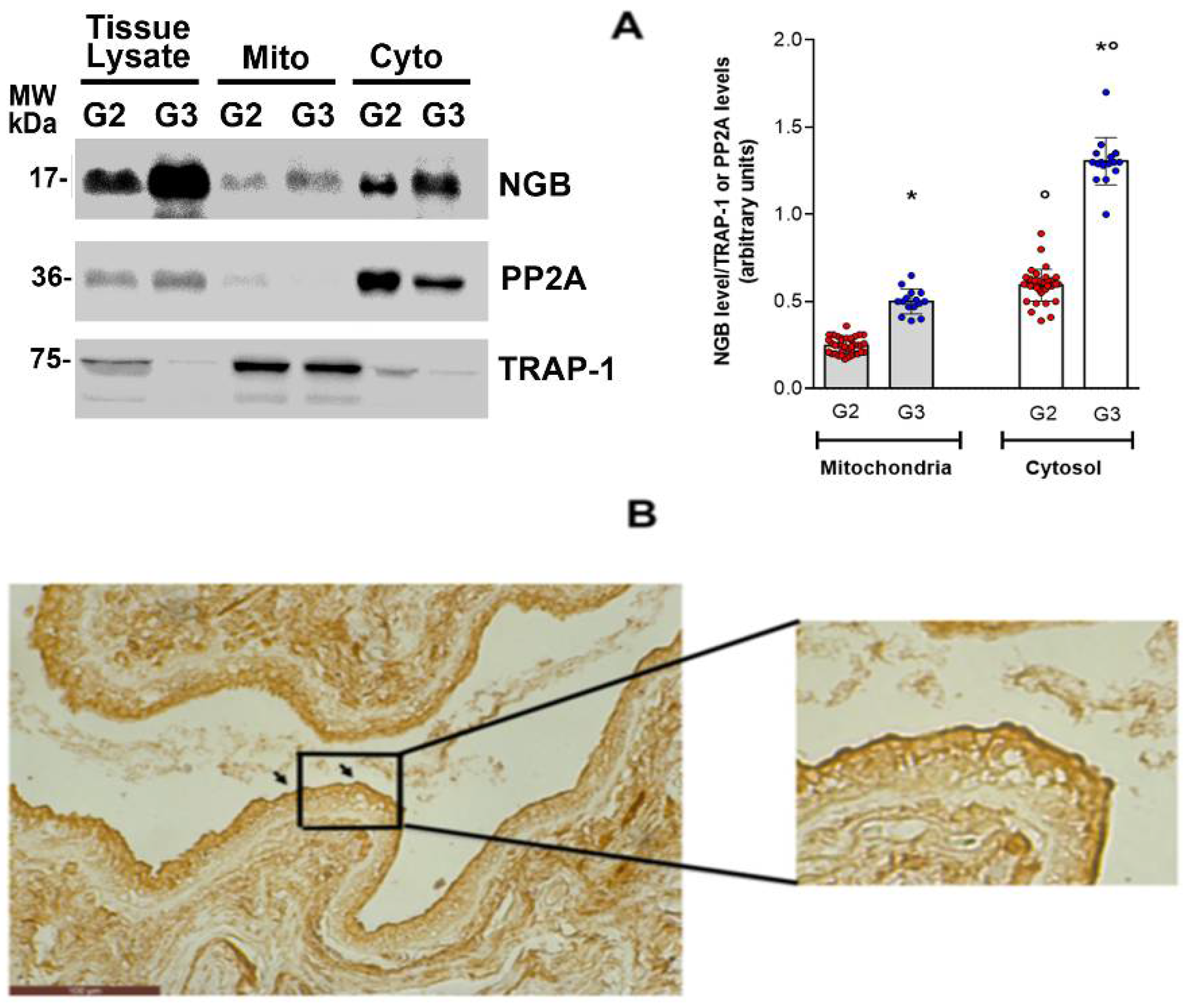
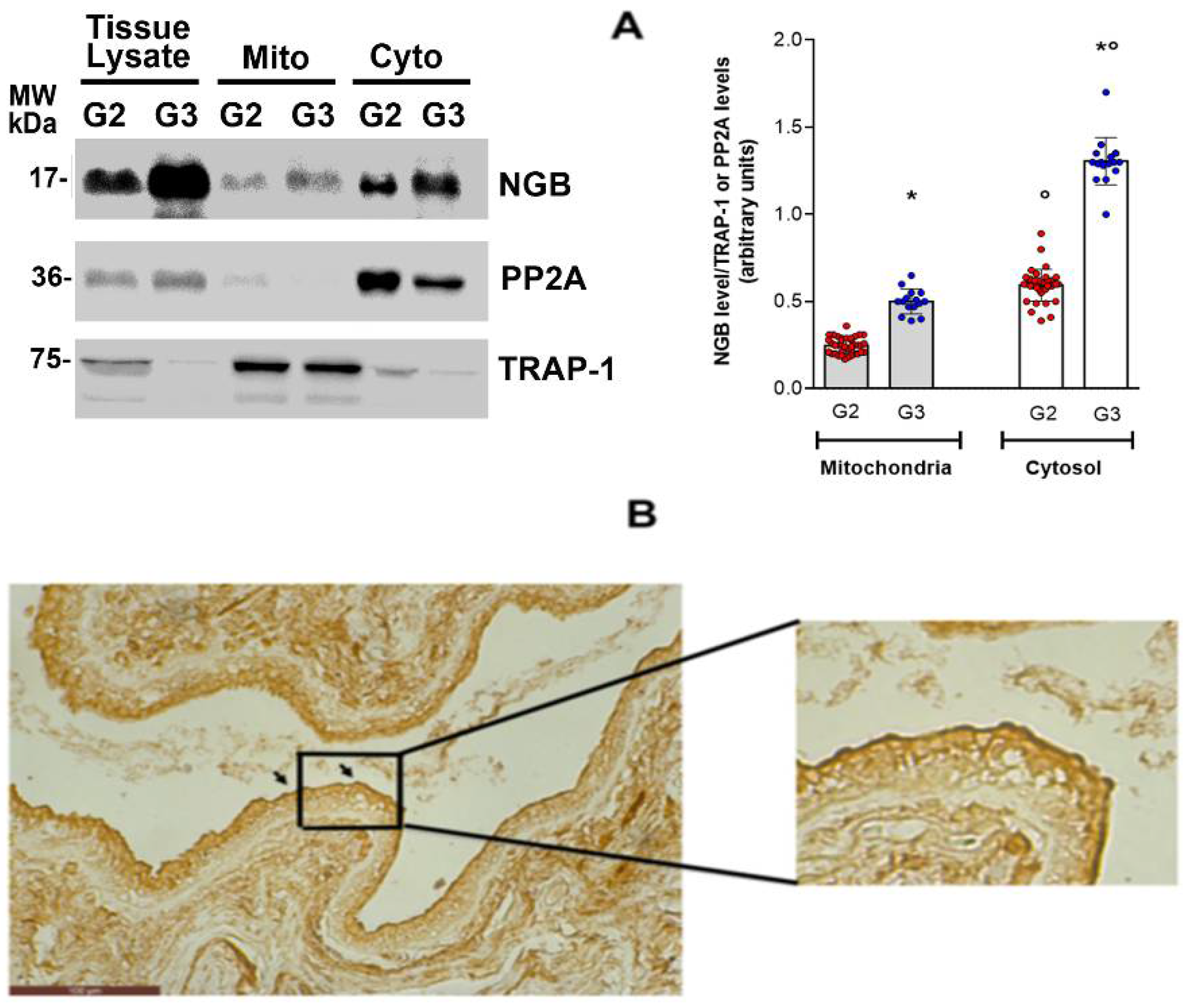
3.4. NGB Localization in Breast Cancer Tissues
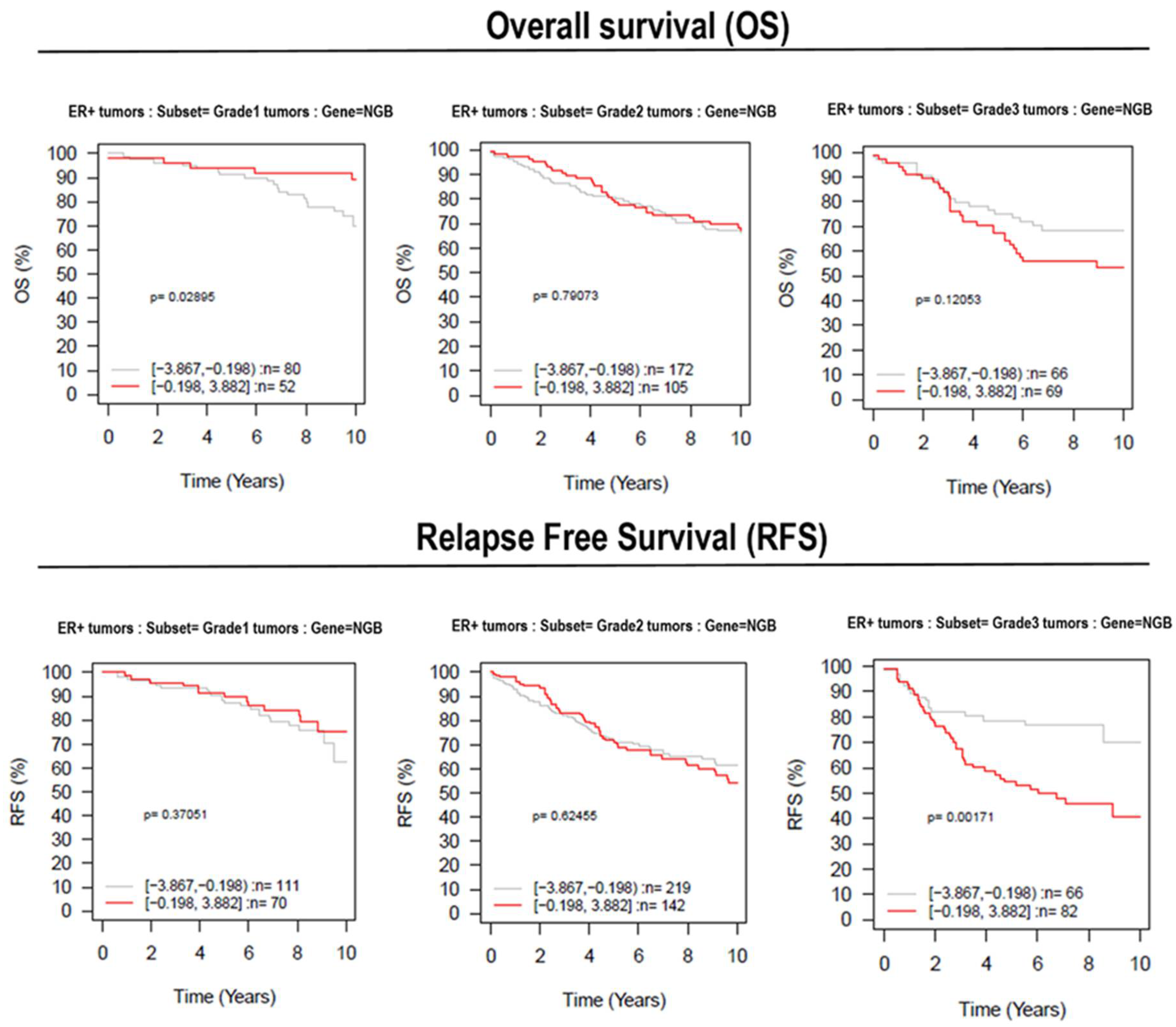
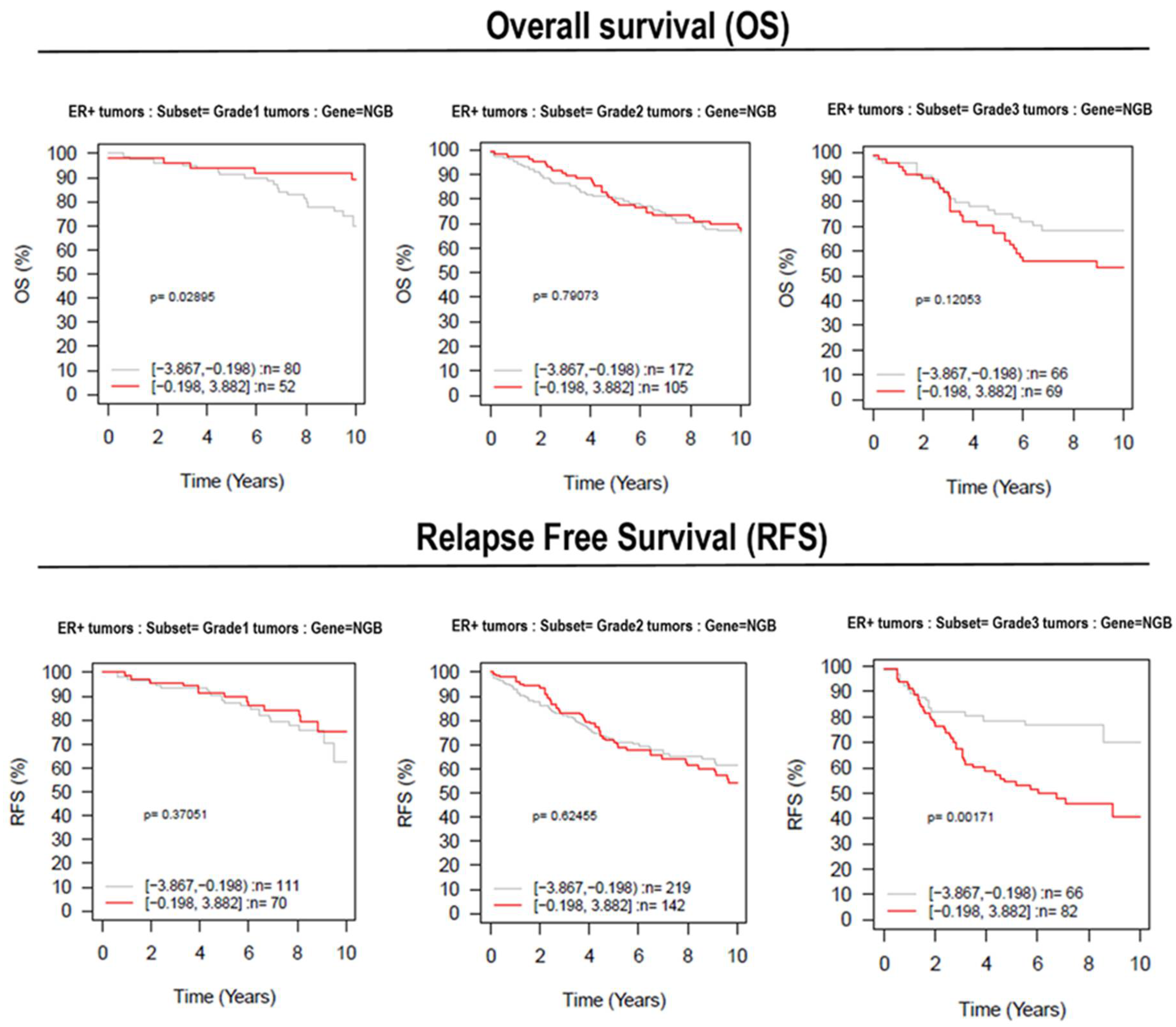
3.5. Effect of NGB Expression in Breast Cancer Patient Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, C.E.; Ma, J.; Gaudet, M.M.; Newman, L.A.; Miller, K.D.; Sauer, A.G.; Jemal, A.; Siegel, R.L. Breast Cancer Statistics, 2019. CA A Cancer J. Clin. 2019, 69, 438–451. [Google Scholar] [CrossRef]
- Acconcia, F.; Marino, M. The Effects of 17b-Estradiol in Cancer Are Mediated by Estrogen Receptor Signaling at the Plasma Membrane. Front. Physiol. 2011, 2, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skandalis, S.S.; Afratis, N.; Smirlaki, G.; Nikitovic, D.; Theocharis, A.D.; Tzanakakis, G.N.; Karamanos, N.K. Cross-Talk between Estradiol Receptor and EGFR/IGF-IR Signaling Pathways in Estrogen-Responsive Breast Cancers: Focus on the Role and Impact of Proteoglycans. Matrix Biol. 2014, 35, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Vuong, D.; Simpson, P.T.; Green, B.; Cummings, M.C.; Lakhani, S.R. Molecular Classification of Breast Cancer. Virchows Arch. 2014, 465, 1–14. [Google Scholar] [CrossRef]
- Lei, J.T.; Anurag, M.; Haricharan, S.; Gou, X.; Ellis, M.J. Endocrine Therapy Resistance: New Insights1. Breast 2019, 48, S26–S30. [Google Scholar] [CrossRef] [Green Version]
- Murphy, L.C.; Seekallu, S.V.; Watson, P.H. Clinical Significance of Estrogen Receptor Phosphorylation. Endocr. Relat. Cancer 2011, 18, R1–R14. [Google Scholar] [CrossRef] [PubMed]
- Fiocchetti, M.; Nuzzo, M.T.; Totta, P.; Acconcia, F.; Ascenzi, P.; Marino, M. Neuroglobin, a pro-Survival Player in Estrogen Receptor α -Positive Cancer Cells. Cell Death Dis. 2014, 5, e1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fordel, E.; Thijs, L.; Martinet, W.; Lenjou, M.; Laufs, T.; Van Bockstaele, D.; Moens, L.; Dewilde, S. Neuroglobin and Cytoglobin Overexpression Protects Human SH-SY5Y Neuroblastoma Cells against Oxidative Stress-Induced Cell Death. Neurosci. Lett. 2006, 410, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, D.A.; Jin, K.; Khan, A.A. Neuroglobin: An Endogenous Neuroprotectant. Curr. Opin. Pharmacol. 2008, 8, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.A.; Wang, Y.; Sun, Y.; Mao, X.O.; Xie, L.; Miles, E.; Graboski, J.; Chen, S.; Ellerby, L.M.; Jin, K.; et al. Neuroglobin-Overexpressing Transgenic Mice Are Resistant to Cerebral and Myocardial Ischemia. Proc. Natl. Acad. Sci. USA 2006, 103, 17944–17948. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Jin, K.; Mao, X.O.; Zhu, Y.; Greenberg, D.A. Neuroglobin Is Up-Regulated by and Protects Neurons from Hypoxic-Ischemic Injury. Proc. Natl. Acad. Sci. USA 2001, 98, 15306–15311. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Liu, N.; Liu, J.; Yang, K.; Wang, X. Neuroglobin, a Novel Target for Endogenous Neuroprotection against Stroke and Neurodegenerative Disorders. Int. J. Mol. Sci. 2012, 13, 6995–7014. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; di Masi, A.; Leboffe, L.; Fiocchetti, M.; Nuzzo, M.T.; Brunori, M.; Marino, M. Neuroglobin: From Structure to Function in Health and Disease. Mol. Asp. Med. 2016, 52, 1–48. [Google Scholar] [CrossRef]
- Fiocchetti, M.; Cipolletti, M.; Leone, S.; Ascenzi, P.; Marino, M. Neuroglobin Overexpression Induced by the 17β-Estradiol–Estrogen Receptor-α Pathway Reduces the Sensitivity of MCF-7 Breast Cancer Cell to Paclitaxel. IUBMB Life 2016, 68, 645–651. [Google Scholar] [CrossRef]
- Fiocchetti, M.; Cipolletti, M.; Ascenzi, P.; Marino, M. Dissecting the 17β-Estradiol Pathways Necessary for Neuroglobin Anti-Apoptotic Activity in Breast Cancer. J. Cell. Physiol. 2018, 233, 5087–5103. [Google Scholar] [CrossRef] [PubMed]
- Emara, M.; Turner, A.R.; Allalunis-Turner, J. Hypoxic Regulation of Cytoglobin and Neuroglobin Expression in Human Normal and Tumor Tissues. Cancer Cell Int. 2010, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Gorr, T.A.; Wichmann, D.; Pilarsky, C.; Theurillat, J.-P.; Fabrizius, A.; Laufs, T.; Bauer, T.; Koslowski, M.; Horn, S.; Burmester, T.; et al. Old Proteins–New Locations: Myoglobin, Haemoglobin, Neuroglobin and Cytoglobin in Solid Tumours and Cancer Cells. Acta Physiol. 2011, 202, 563–581. [Google Scholar] [CrossRef]
- Oleksiewicz, U.; Daskoulidou, N.; Liloglou, T.; Tasopoulou, K.; Bryan, J.; Gosney, J.R.; Field, J.K.; Xinarianos, G. Neuroglobin and Myoglobin in Non-Small Cell Lung Cancer: Expression, Regulation and Prognosis. Lung Cancer 2011, 74, 411–418. [Google Scholar] [CrossRef]
- Zhang, J.; Lan, S.J.; Liu, Q.R.; Liu, J.M.; Chen, X.Q. Neuroglobin, a Novel Intracellular Hexa-Coordinated Globin, Functions as a Tumor Suppressor in Hepatocellular Carcinoma via Raf/MAPK/Erk. Mol. Pharm. 2013, 83, 1109–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, L.-A.; Dowsett, M. BCL-2: A New Therapeutic Target in Estrogen Receptor-Positive Breast Cancer? Cancer Cell 2013, 24, 7–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acconcia, F.; Kumar, R. Signaling Regulation of Genomic and Nongenomic Functions of Estrogen Receptors. Cancer Lett. 2006, 238, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Medunjanin, S.; Hermani, A.; De Servi, B.; Grisouard, J.; Rincke, G.; Mayer, D. Glycogen Synthase Kinase-3 Interacts with and Phosphorylates Estrogen Receptor α and Is Involved in the Regulation of Receptor Activity. J. Biol. Chem. 2005, 280, 33006–33014. [Google Scholar] [CrossRef] [Green Version]
- Fiocchetti, M.; Cipolletti, M.; Marino, M. Compensatory Role of Neuroglobin in Nervous and Non-Nervous Cancer Cells in Response to the Nutrient Deprivation. PLoS ONE 2017, 12, e0189179. [Google Scholar] [CrossRef] [Green Version]
- Ringnér, M.; Fredlund, E.; Häkkinen, J.; Borg, Å.; Staaf, J. GOBO: Gene Expression-Based Outcome for Breast Cancer Online. PLoS ONE 2011, 6, e17911. [Google Scholar] [CrossRef] [PubMed]
- Fiocchetti, M.; Cracco, P.; Montalesi, E.; Solar Fernandez, V.; Stuart, J.A.; Marino, M. Neuroglobin and Mitochondria: The Impact on Neurodegenerative Diseases. Arch. Biochem. Biophys. 2021, 701, 108823. [Google Scholar] [CrossRef]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.-P.; Tong, F.; et al. A Collection of Breast Cancer Cell Lines for the Study of Functionally Distinct Cancer Subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Ali, H.; Rousseau, J.; Gantchev, T.G.; van Lier, J.E. 2- and 4-Fluorinated 16.Alpha.-[125I]Iodoestradiol Derivatives: Synthesis and Effect on Estrogen Receptor Binding and Receptor-Mediated Target Tissue Uptake. J. Med. Chem. 1993, 36, 4255–4263. [Google Scholar] [CrossRef]
- Geisler, J. Breast Cancer Tissue Estrogens and Their Manipulation with Aromatase Inhibitors and Inactivators. J. Steroid Biochem. Mol. Biol. 2003, 86, 245–253. [Google Scholar] [CrossRef]
- Savolainen-Peltonen, H.; Vihma, V.; Leidenius, M.; Wang, F.; Turpeinen, U.; Hämäläinen, E.; Tikkanen, M.J.; Mikkola, T.S. Breast Adipose Tissue Estrogen Metabolism in Postmenopausal Women With or Without Breast Cancer. J. Clin. Endocrinol. Metab. 2014, 99, E2661–E2667. [Google Scholar] [CrossRef] [Green Version]
- Khatpe, A.S.; Adebayo, A.K.; Herodotou, C.A.; Kumar, B.; Nakshatri, H. Nexus between PI3K/AKT and Estrogen Receptor Signaling in Breast Cancer. Cancers 2021, 13, 369. [Google Scholar] [CrossRef] [PubMed]
- Acconcia, F.; Totta, P.; Ogawa, S.; Cardillo, I.; Inoue, S.; Leone, S.; Trentalance, A.; Muramatsu, M.; Marino, M. Survival versus Apoptotic 17β-Estradiol Effect: Role of ERα and ERβ Activated Non-Genomic Signaling. J. Cell. Physiol. 2005, 203, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, A.V.; George, J.; Senko, O.; Mow, B.; Putti, T.C.; Smeds, J.; Lindahl, T.; Pawitan, Y.; Hall, P.; Nordgren, H.; et al. Genetic Reclassification of Histologic Grade Delineates New Clinical Subtypes of Breast Cancer. Cancer Res. 2006, 66, 10292–10301. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Poppe, J.L.; Wang, X. Mitochondrial Mechanisms of Neuroglobin’s Neuroprotection. Oxidative Med. Cell. Longev. 2013, 2013, e756989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiocchetti, M.; Solar Fernandez, V.; Segatto, M.; Leone, S.; Cercola, P.; Massari, A.; Cavaliere, F.; Marino, M. Extracellular Neuroglobin as a Stress-Induced Factor Activating Pre-Adaptation Mechanisms against Oxidative Stress and Chemotherapy-Induced Cell Death in Breast Cancer. Cancers 2020, 12, 2451. [Google Scholar] [CrossRef] [PubMed]
- Shivapurkar, N.; Stastny, V.; Okumura, N.; Girard, L.; Xie, Y.; Prinsen, C.; Thunnissen, F.B.; Wistuba, I.I.; Czerniak, B.; Frenkel, E.; et al. Cytoglobin, the Newest Member of the Globin Family, Functions as a Tumor Suppressor Gene. Cancer Res. 2008, 68, 7448–7456. [Google Scholar] [CrossRef] [Green Version]
- Bholah, T.C.; Neergheen-Bhujun, V.S.; Hodges, N.J.; Dyall, S.D.; Bahorun, T. Cytoglobin as a Biomarker in Cancer: Potential Perspective for Diagnosis and Management. BioMed Res. Int. 2015, 2015, e824514. [Google Scholar] [CrossRef] [Green Version]
- Rowland, L.K.; Campbell, P.S.; Mavingire, N.; Wooten, J.V.; McLean, L.; Zylstra, D.; Thorne, G.; Daly, D.; Boyle, K.; Whang, S.; et al. Putative Tumor Suppressor Cytoglobin Promotes Aryl Hydrocarbon Receptor Ligand–Mediated Triple Negative Breast Cancer Cell Death. J. Cell. Biochem. 2019, 120, 6004–6014. [Google Scholar] [CrossRef]
- Flonta, S.E.; Arena, S.; Pisacane, A.; Michieli, P.; Bardelli, A. Expression and Functional Regulation of Myoglobin in Epithelial Cancers. Am. J. Pathol. 2009, 175, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Bicker, A.; Nauth, T.; Gerst, D.; Aboouf, M.A.; Fandrey, J.; Kristiansen, G.; Gorr, T.A.; Hankeln, T. The Role of Myoglobin in Epithelial Cancers: Insights from Transcriptomics. Int. J. Mol. Med. 2020, 45, 385–400. [Google Scholar] [CrossRef] [Green Version]
- Kristiansen, G.; Rose, M.; Geisler, C.; Fritzsche, F.R.; Gerhardt, J.; Lüke, C.; Ladhoff, A.-M.; Knüchel, R.; Dietel, M.; Moch, H.; et al. Endogenous Myoglobin in Human Breast Cancer Is a Hallmark of Luminal Cancer Phenotype. Br. J. Cancer 2010, 102, 1736–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, D.A.; Cirrincione, C.; Henderson, I.C.; Citron, M.L.; Budman, D.R.; Goldstein, L.J.; Martino, S.; Perez, E.A.; Muss, H.B.; Norton, L.; et al. Estrogen-Receptor Status and Outcomes of Modern Chemotherapy for Patients With Node-Positive Breast Cancer. JAMA 2006, 295, 1658–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faneyte, I.F.; Schrama, J.G.; Peterse, J.L.; Remijnse, P.L.; Rodenhuis, S.; van de Vijver, M.J. Breast Cancer Response to Neoadjuvant Chemotherapy: Predictive Markers and Relation with Outcome. Br. J. Cancer 2003, 88, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Solar Fernandez, V.; Cipolletti, M.; Ascenzi, P.; Marino, M.; Fiocchetti, M. Neuroglobin As Key Mediator in the 17β-Estradiol-Induced Antioxidant Cell Response to Oxidative Stress. Antioxid. Redox Signal. 2019, 32, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Fiocchetti, M.; Cipolletti, M.; Leone, S.; Naldini, A.; Carraro, F.; Giordano, D.; Verde, C.; Ascenzi, P.; Marino, M. Neuroglobin in Breast Cancer Cells: Effect of Hypoxia and Oxidative Stress on Protein Level, Localization, and Anti-Apoptotic Function. PLoS ONE 2016, 11, e0154959. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Number of Patients (NTOT = 53) | % on Total Patients | |
|---|---|---|---|
| Menopausal status | Yes (Age ≥ 51 years) | 38 | 71.7 |
| No (Age < 51 years) | 15 | 28.3 | |
| Hystological type | Ductal carcinoma | 53 | 100 |
| ER/PR Status | Positive | 53 | 100 |
| Grade | G2 | 38 (26 Menopause Y; 12 Menopause N) | 71.7 |
| G3 | 15 (12 Menopause Y; 3 Menopause N) | 28.3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solar Fernandez, V.; Fiocchetti, M.; Cipolletti, M.; Segatto, M.; Cercola, P.; Massari, A.; Ghinassi, S.; Cavaliere, F.; Marino, M. Neuroglobin: A New Possible Marker of Estrogen-Responsive Breast Cancer. Cells 2021, 10, 1986. https://doi.org/10.3390/cells10081986
Solar Fernandez V, Fiocchetti M, Cipolletti M, Segatto M, Cercola P, Massari A, Ghinassi S, Cavaliere F, Marino M. Neuroglobin: A New Possible Marker of Estrogen-Responsive Breast Cancer. Cells. 2021; 10(8):1986. https://doi.org/10.3390/cells10081986
Chicago/Turabian StyleSolar Fernandez, Virginia, Marco Fiocchetti, Manuela Cipolletti, Marco Segatto, Paolo Cercola, Annalisa Massari, Sabrina Ghinassi, Francesco Cavaliere, and Maria Marino. 2021. "Neuroglobin: A New Possible Marker of Estrogen-Responsive Breast Cancer" Cells 10, no. 8: 1986. https://doi.org/10.3390/cells10081986
APA StyleSolar Fernandez, V., Fiocchetti, M., Cipolletti, M., Segatto, M., Cercola, P., Massari, A., Ghinassi, S., Cavaliere, F., & Marino, M. (2021). Neuroglobin: A New Possible Marker of Estrogen-Responsive Breast Cancer. Cells, 10(8), 1986. https://doi.org/10.3390/cells10081986