
Heteromeric TRP Channels in Lung Inflammation


Abstract
1. Introduction
2. Methodological Approaches and Limitations in the Analysis of Heteromeric TRP Assemblies
3. Heteromeric TRP Channel Assemblies and Their Molecular Mechanisms
3.1. Introduction to TRP Channel Assemblies
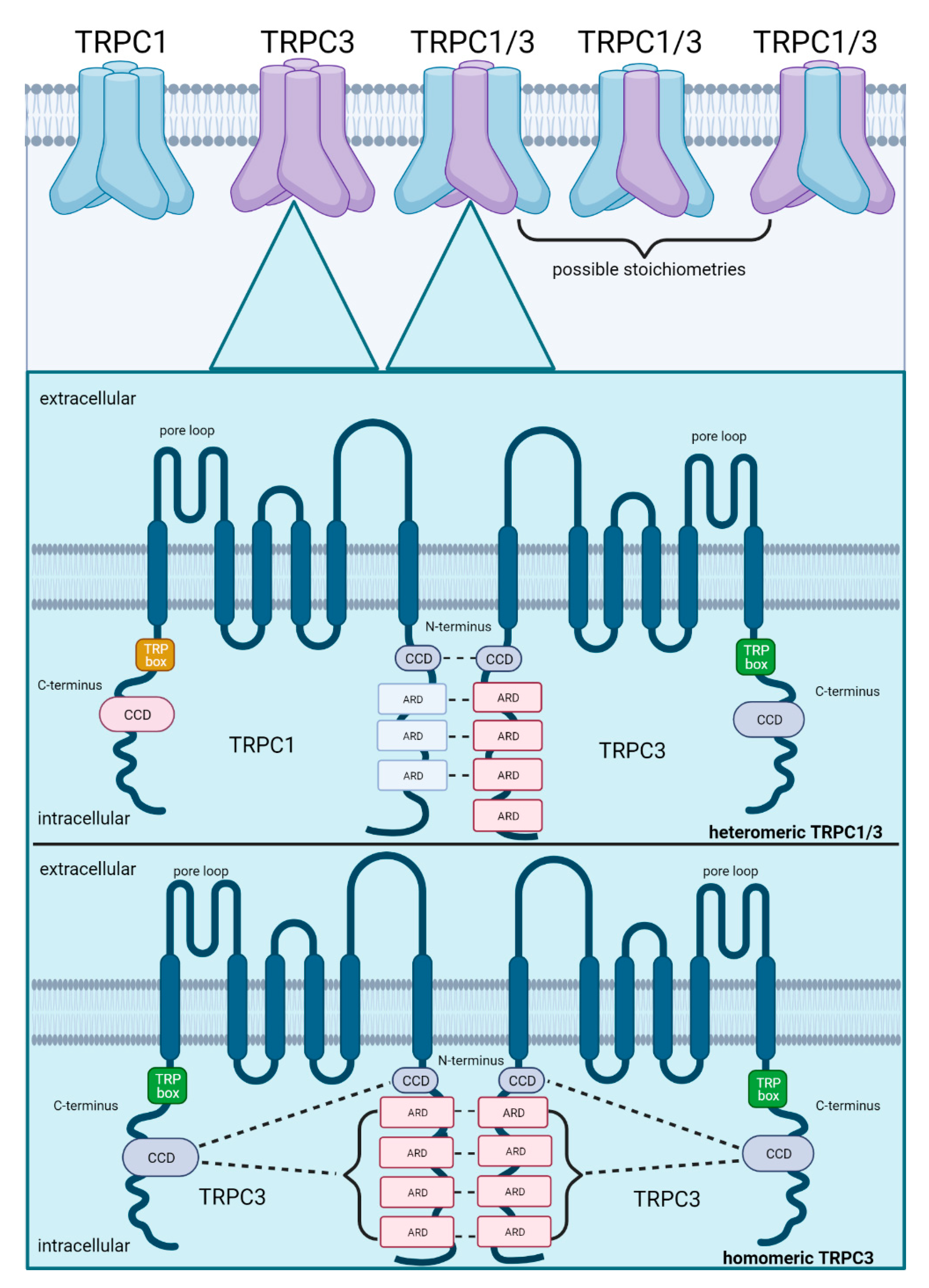
3.2. Heteromeric TRP Channel Assemblies
3.3. Molecular Mechanisms for Stabilization of Heteromeric TRP Channel Assemblies
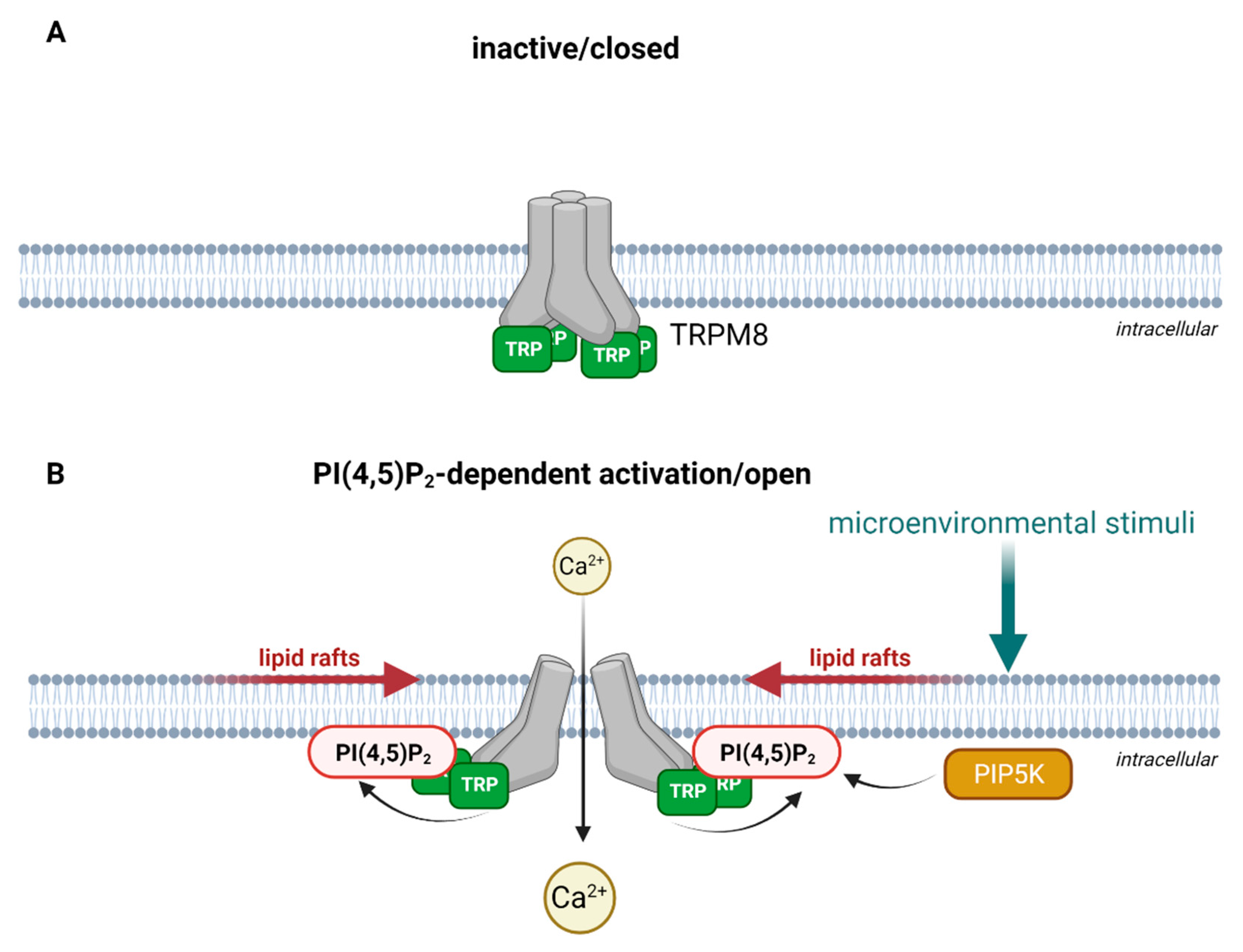
3.4. Properties of Heteromeric TRP Channel Assemblies
4. Heteromeric TRPC1/4 Assembly
4.1. Heteromeric TRPC1/4 Structure
4.2. TRPC1/4 Regulation
4.3. TRPC1/4 Function
5. Heteromeric TRPC3/6 Assembly
5.1. Heteromeric TRPC3/6 Structure
5.2. TRPC3/6 Regulation
5.3. TRPC3/6 Function
6. Heteromeric TRPV1/4 Assembly
6.1. TRPV1/4 Structure
6.2. TRPV1/4 Regulation
6.3. TRPV1/4 Function
7. Other Heteromeric TRP Assemblies
8. Role of Heteromeric TRP Assemblies in Lung Inflammation
8.1. Introduction to Lung Inflammation
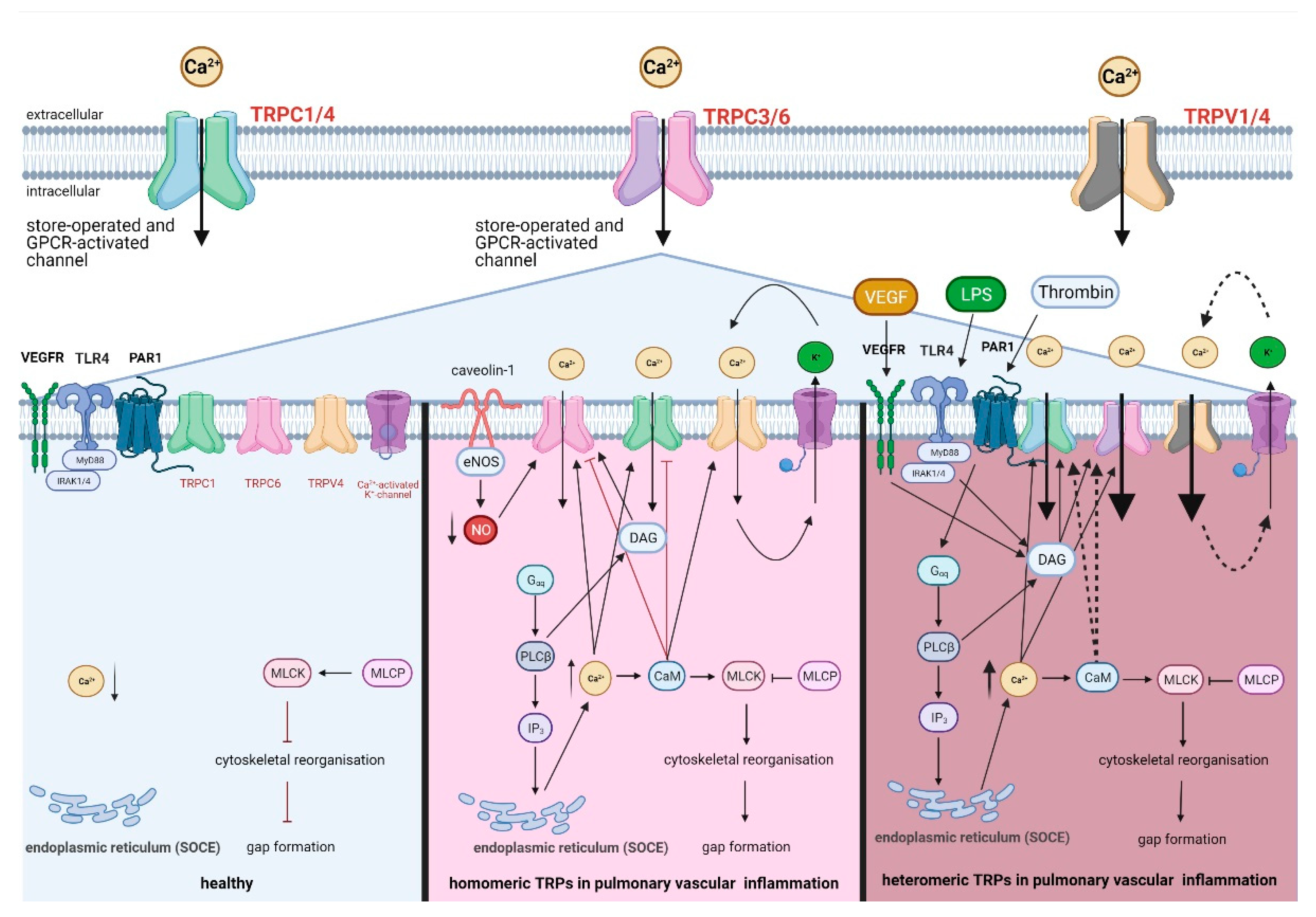
8.2. Activation of Heteromeric TRPC1/4 and TRPC3/6 in Lung Inflammation
8.3. Activation of Homomeric TRPC1, TRPC6, TRPV1, and TRPV4 in Lung Inflammation
8.4. Comparison of Heteromeric TRP Assemblies to Homomeric TRP Assemblies in Lung Inflammation
9. Summary
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AFM | atomic force microscopy |
| ARDS | Acute Respiratory Distress Syndrome |
| ARDs | ankyrin repeat domains |
| CaM | calmodulin |
| CCDs | coiled-coil domains |
| Co-IP | co-immunoprecipitation |
| DAG | diacylglycerol |
| eNOS | endothelial NO-synthase |
| FRET | Förster resonance energy transfer |
| GPCR | G-protein-coupled receptor |
| HEK293 | human embryonic kidney cells 293 |
| HMVECs | human microvascular endothelial cells |
| HUVECs | human umbilical vein endothelial cells |
| IK | intermediate conductance Ca2+-activated K+ channel |
| IP3 | inositol 1,4,5-trisphosphate |
| LPS | lipopolysaccharide |
| RMAECs | rat mesenteric artery endothelial cells |
| MLCII | myosin light chain II |
| MLCK | myosin light chain kinase |
| MLCP | myosin light chain phosphatase |
| MRLC | myosin II regulatory light chains |
| NF-κB | nuclear factor-kappa B |
| Orai1 | Ca2+ release-activated calcium channel protein |
| PAR1 | protease-activated receptor 1 |
| PI(4,5)P2 | phosphatidylinositol 4,5-bisphosphate |
| PLA | proximity ligation assay |
| PLCβ/γ | phospholipase C β/γ |
| PMNs | polymorphonuclear leukocytes |
| PRR | pathogen recognition receptors |
| RMECs | retinal microvascular endothelial cells |
| ROS | reactive oxygen species |
| SC-SMD | single channel single molecule detection |
| SK | small Ca2+-dependent K+ channel |
| SOCE | store-operated Ca2+ entry |
| STIM1 | stromal interaction molecule |
| TIRF | total internal reflection fluorescence microscopy |
| TLR | Toll-like receptors |
| TRP | transient receptor potential |
| VEGF | vascular endothelial growth factor |
| Y2H | yeast two-hybrid |
References
- Cioffi, D.L.; Lowe, K.; Alvarez, D.F.; Barry, C.; Stevens, T. TRPing on the lung endothelium: Calcium channels that regulate barrier function. Antioxid. Redox Signal. 2009, 11, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Kuebler, W.M. Mechanotransduction by TRP channels: General concepts and specific role in the vasculature. Cell Biochem. Biophys. 2010, 56, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Simmons, S.; Erfinanda, L.; Bartz, C.; Kuebler, W.M. Novel mechanisms regulating endothelial barrier function in the pulmonary microcirculation. J. Physiol. 2019, 597, 997–1021. [Google Scholar] [CrossRef]
- Villalta, P.C.; Townsley, M.I. Transient receptor potential channels and regulation of lung endothelial permeability. Pulm Circ. 2013, 3, 802–815. [Google Scholar] [CrossRef]
- Montell, C.; Birnbaumer, L.; Flockerzi, V.; Bindels, R.J.; Bruford, E.A.; Caterina, M.J.; Clapham, D.E.; Harteneck, C.; Heller, S.; Julius, D.; et al. A unified nomenclature for the superfamily of TRP cation channels. Mol. Cell 2002, 9, 229–231. [Google Scholar] [CrossRef]
- Paria, B.C.; Vogel, S.M.; Ahmmed, G.U.; Alamgir, S.; Shroff, J.; Malik, A.B.; Tiruppathi, C. Tumor necrosis factor-alpha-induced TRPC1 expression amplifies store-operated Ca2+ influx and endothelial permeability. Am. J. Physiol Lung Cell Mol. Physiol. 2004, 287, L1303–L1313. [Google Scholar] [CrossRef]
- Ma, X.; Cheng, K.T.; Wong, C.O.; O’Neil, R.G.; Birnbaumer, L.; Ambudkar, I.S.; Yao, X. Heteromeric TRPV4-C1 channels contribute to store-operated Ca(2+) entry in vascular endothelial cells. Cell Calcium 2011, 50, 502–509. [Google Scholar] [CrossRef]
- Yin, J.; Michalick, L.; Tang, C.; Tabuchi, A.; Goldenberg, N.; Dan, Q.; Awwad, K.; Wang, L.; Erfinanda, L.; Nouailles, G.; et al. Role of Transient Receptor Potential Vanilloid 4 in Neutrophil Activation and Acute Lung Injury. Am. J. Respir. Cell Mol. Biol. 2016, 54, 370–383. [Google Scholar] [CrossRef]
- Hecquet, C.M.; Zhang, M.; Mittal, M.; Vogel, S.M.; Di, A.; Gao, X.; Bonini, M.G.; Malik, A.B. Cooperative interaction of trp melastatin channel transient receptor potential (TRPM2) with its splice variant TRPM2 short variant is essential for endothelial cell apoptosis. Circ. Res. 2014, 114, 469–479. [Google Scholar] [CrossRef]
- Earley, S.; Brayden, J.E. Transient receptor potential channels in the vasculature. Physiol. Rev. 2015, 95, 645–690. [Google Scholar] [CrossRef]
- Moccia, F.; Guerra, G. Ca(2+) Signalling in Endothelial Progenitor Cells: Friend or Foe? J. Cell Physiol. 2016, 231, 314–327. [Google Scholar] [CrossRef]
- Smani, T.; Gomez, L.J.; Regodon, S.; Woodard, G.E.; Siegfried, G.; Khatib, A.M.; Rosado, J.A. TRP Channels in Angiogenesis and Other Endothelial Functions. Front. Physiol. 2018, 9, 1731. [Google Scholar] [CrossRef]
- Thomas, K.C.; Roberts, J.K.; Deering-Rice, C.E.; Romero, E.G.; Dull, R.O.; Lee, J.; Yost, G.S.; Reilly, C.A. Contributions of TRPV1, endovanilloids, and endoplasmic reticulum stress in lung cell death in vitro and lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 302, L111–L119. [Google Scholar] [CrossRef]
- Michalick, L.; Erfinanda, L.; Weichelt, U.; van der Giet, M.; Liedtke, W.; Kuebler, W.M. Transient Receptor Potential Vanilloid 4 and Serum Glucocorticoid-regulated Kinase 1 Are Critical Mediators of Lung Injury in Overventilated Mice In Vivo. Anesthesiology 2017, 126, 300–311. [Google Scholar] [CrossRef]
- Townsley, M.I. Permeability and calcium signaling in lung endothelium: Unpack the box. Pulm Circ. 2018, 8, 2045893217738218. [Google Scholar] [CrossRef]
- Kedei, N.; Szabo, T.; Lile, J.D.; Treanor, J.J.; Olah, Z.; Iadarola, M.J.; Blumberg, P.M. Analysis of the native quaternary structure of vanilloid receptor 1. J. Biol. Chem. 2001, 276, 28613–28619. [Google Scholar] [CrossRef]
- Hoenderop, J.G.; Voets, T.; Hoefs, S.; Weidema, F.; Prenen, J.; Nilius, B.; Bindels, R.J. Homo- and heterotetrameric architecture of the epithelial Ca2+ channels TRPV5 and TRPV6. EMBO J. 2003, 22, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ramirez, R.; Chen, Y.; Liedtke, W.B.; Morales-Lazaro, S.L. TRP Channels and Pain. In Neurobiology of TRP Channels; Emir, T.L.R., Ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 125–147. [Google Scholar]
- Cao, S.; Anishkin, A.; Zinkevich, N.S.; Nishijima, Y.; Korishettar, A.; Wang, Z.; Fang, J.; Wilcox, D.A.; Zhang, D.X. Transient receptor potential vanilloid 4 (TRPV4) activation by arachidonic acid requires protein kinase A-mediated phosphorylation. J. Biol. Chem. 2018, 293, 5307–5322. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, M. Homo- and heteromeric assembly of TRP channel subunits. Pflug. Arch. 2005, 451, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.X.; Giamarchi, A.; Rodat-Despoix, L.; Padilla, F.; Downs, T.; Tsiokas, L.; Delmas, P. Formation of a new receptor-operated channel by heteromeric assembly of TRPP2 and TRPC1 subunits. EMBO Rep. 2008, 9, 472–479. [Google Scholar] [CrossRef]
- Cheng, H.W.; James, A.F.; Foster, R.R.; Hancox, J.C.; Bates, D.O. VEGF activates receptor-operated cation channels in human microvascular endothelial cells. Arter. Thromb. Vasc. Biol. 2006, 26, 1768–1776. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Gross, C.M.; Kellner, M.; Wang, T.; Lu, Q.; Sun, X.; Zemskov, E.A.; Noonepalle, S.; Kangath, A.; Kumar, S.; Gonzalez-Garay, M.; et al. LPS-induced Acute Lung Injury Involves NF-kappaB-mediated Downregulation of SOX18. Am. J. Respir. Cell Mol. Biol. 2018, 58, 614–624. [Google Scholar] [CrossRef]
- Singh, A.K.; McGoldrick, L.L.; Sobolevsky, A.I. Expression, Purification, and Crystallization of the Transient Receptor Potential Channel TRPV6. Methods Mol. Biol. 2019, 1987, 23–37. [Google Scholar]
- Fredriksson, S.; Gullberg, M.; Jarvius, J.; Olsson, C.; Pietras, K.; Gustafsdottir, S.M.; Ostman, A.; Landegren, U. Protein detection using proximity-dependent DNA ligation assays. Nat. Biotechnol. 2002, 20, 473–477. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, C.; McGahon, M.K.; Ashraf, S.; McNaughten, J.; Friedel, T.; Cincola, P.; Barabas, P.; Fernandez, J.A.; Stitt, A.W.; McGeown, J.G.; et al. Involvement of TRPV1 and TRPV4 Channels in Retinal Angiogenesis. Invest. Ophthalmol Vis. Sci. 2019, 60, 3297–3309. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.; Schaefer, M.; Schultz, G.; Gudermann, T. Subunit composition of mammalian transient receptor potential channels in living cells. Proc. Natl. Acad. Sci. USA 2002, 99, 7461–7466. [Google Scholar] [CrossRef]
- Xu, X.Z.; Li, H.S.; Guggino, W.B.; Montell, C. Coassembly of TRP and TRPL produces a distinct store-operated conductance. Cell 1997, 89, 1155–1164. [Google Scholar] [CrossRef]
- Lentze, N.; Auerbach, D. Membrane-based yeast two-hybrid system to detect protein interactions. Curr. Protoc. Protein Sci. 2008, 52. [Google Scholar] [CrossRef]
- Das, S.; Chakrabarti, S. Classification and prediction of protein-protein interaction interface using machine learning algorithm. Sci. Rep. 2021, 11, 1761. [Google Scholar] [CrossRef]
- Singh, A.K.; McGoldrick, L.L.; Saotome, K.; Sobolevsky, A.I. X-ray crystallography of TRP channels. Channels (Austin) 2018, 12, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Hughes, T.E.T.; Moiseenkova-Bell, V.Y. Cryo Electron Microscopy of TRP Channels. Methods Mol. Biol. 2019, 1987, 39–50. [Google Scholar] [PubMed]
- Stewart, A.P.; Smith, G.D.; Sandford, R.N.; Edwardson, J.M. Atomic force microscopy reveals the alternating subunit arrangement of the TRPP2-TRPV4 heterotetramer. Biophys. J. 2010, 99, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Sinz, A.; Arlt, C.; Chorev, D.; Sharon, M. Chemical cross-linking and native mass spectrometry: A fruitful combination for structural biology. Protein Sci. 2015, 24, 1193–1209. [Google Scholar] [CrossRef]
- Silva, L.P. Imaging proteins with atomic force microscopy: An overview. Curr. Protein Pept. Sci. 2005, 6, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Takahashi, Y. Analysis of Protein-Protein Interaction by Co-IP in Human Cells. Methods Mol. Biol. 2018, 1794, 289–296. [Google Scholar] [PubMed]
- Hellwig, N.; Albrecht, N.; Harteneck, C.; Schultz, G.; Schaefer, M. Homo- and heteromeric assembly of TRPV channel subunits. J. Cell Sci. 2005, 118, 917–928. [Google Scholar] [CrossRef]
- Iacobucci, C.; Gotze, M.; Sinz, A. Cross-linking/mass spectrometry to get a closer view on protein interaction networks. Curr. Opin. Biotechnol. 2020, 63, 48–53. [Google Scholar] [CrossRef]
- Danev, R.; Yanagisawa, H.; Kikkawa, M. Cryo-Electron Microscopy Methodology: Current Aspects and Future Directions. Trends Biochem. Sci. 2019, 44, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, C.F.; Rees, E.J.; Schierle, G.S. A quantitative protocol for intensity-based live cell FRET imaging. Methods Mol. Biol. 2014, 1076, 445–454. [Google Scholar]
- Alam, M.S. Proximity Ligation Assay (PLA). Curr. Protoc. Immunol. 2018, 123, e58. [Google Scholar] [CrossRef]
- Jackson, M.B. Whole-cell voltage clamp recording. Curr. Protoc. Neurosci. 2001, 6, 6. [Google Scholar] [CrossRef]
- Storch, U.; Forst, A.L.; Philipp, M.; Gudermann, T.; Mederos y Schnitzler, M. Transient receptor potential channel 1 (TRPC1) reduces calcium permeability in heteromeric channel complexes. J. Biol. Chem. 2012, 287, 3530–3540. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, L.G.; Asanov, A.; Vaca, L. Single-Channel Single-Molecule Detection (SC-SMD) System. Methods Mol. Biol. 2018, 1843, 189–201. [Google Scholar]
- Smyth, M.S.; Martin, J.H. x ray crystallography. Mol. Pathol. 2000, 53, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Paiano, A.; Margiotta, A.; De Luca, M.; Bucci, C. Yeast Two-Hybrid Assay to Identify Interacting Proteins. Curr. Protoc. Protein Sci. 2019, 95, e70. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef]
- Phelps, C.B.; Procko, E.; Lishko, P.V.; Wang, R.R.; Gaudet, R. Insights into the roles of conserved and divergent residues in the ankyrin repeats of TRPV ion channels. Channels (Austin) 2007, 1, 148–151. [Google Scholar] [CrossRef]
- Li, J.; Mahajan, A.; Tsai, M.D. Ankyrin repeat: A unique motif mediating protein-protein interactions. Biochemistry 2006, 45, 15168–15178. [Google Scholar] [CrossRef]
- Mosavi, L.K.; Minor, D.L., Jr.; Peng, Z.Y. Consensus-derived structural determinants of the ankyrin repeat motif. Proc. Natl. Acad. Sci. USA 2002, 99, 16029–16034. [Google Scholar] [CrossRef]
- Li, M.; Yu, Y.; Yang, J. Structural biology of TRP channels. Adv. Exp. Med. Biol. 2011, 704, 1–23. [Google Scholar]
- Rohacs, T.; Lopes, C.M.; Michailidis, I.; Logothetis, D.E. PI(4,5)P2 regulates the activation and desensitization of TRPM8 channels through the TRP domain. Nat. Neurosci. 2005, 8, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Guo, W.; Zheng, L.; Wu, J.X.; Liu, M.; Zhou, X.; Zhang, X.; Chen, L. Structure of the receptor-activated human TRPC6 and TRPC3 ion channels. Cell Res. 2018, 28, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Tsuruda, P.R.; Julius, D.; Minor, D.L., Jr. Coiled coils direct assembly of a cold-activated TRP channel. Neuron 2006, 51, 201–212. [Google Scholar] [CrossRef]
- Liu, J.; Zheng, Q.; Deng, Y.; Cheng, C.S.; Kallenbach, N.R.; Lu, M. A seven-helix coiled coil. Proc. Natl. Acad. Sci. USA 2006, 103, 15457–15462. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Minor, D.L., Jr. X-ray crystal structure of a TRPM assembly domain reveals an antiparallel four-stranded coiled-coil. J. Mol. Biol. 2008, 383, 854–870. [Google Scholar] [CrossRef]
- Martinez, G.Q.; Gordon, S.E. Multimerization of Homo sapiens TRPA1 ion channel cytoplasmic domains. PLoS ONE 2019, 14, e0207835. [Google Scholar] [CrossRef] [PubMed]
- Myeong, J.; Ko, J.; Hong, C.; Yang, D.; Lee, K.P.; Jeon, J.H.; So, I. The interaction domains of transient receptor potential canonical (TRPC)1/4 and TRPC1/5 heteromultimeric channels. Biochem. Biophys. Res. Commun. 2016, 474, 476–481. [Google Scholar] [CrossRef]
- Schindl, R.; Fritsch, R.; Jardin, I.; Frischauf, I.; Kahr, H.; Muik, M.; Riedl, M.C.; Groschner, K.; Romanin, C. Canonical transient receptor potential (TRPC) 1 acts as a negative regulator for vanilloid TRPV6-mediated Ca2+ influx. J. Biol. Chem. 2012, 287, 35612–35620. [Google Scholar] [CrossRef]
- Guo, Z.; Grimm, C.; Becker, L.; Ricci, A.J.; Heller, S. A novel ion channel formed by interaction of TRPML3 with TRPV5. PLoS ONE 2013, 8, e58174. [Google Scholar] [CrossRef]
- Chen, X.; Lu, M.; He, X.; Ma, L.; Birnbaumer, L.; Liao, Y. TRPC3/6/7 Knockdown Protects the Brain from Cerebral Ischemia Injury via Astrocyte Apoptosis Inhibition and Effects on NF-small ka, CyrillicB Translocation. Mol. Neurobiol. 2017, 54, 7555–7566. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.G.; Wang, F.D.; Sun, Z.C.; Ma, S.B.; Wang, X.; Han, W.J.; Wang, F.; Bai, Z.T.; Wu, S.X.; Freichel, M.; et al. TRPC1/4/5 channels contribute to morphine-induced analgesic tolerance and hyperalgesia by enhancing spinal synaptic potentiation and structural plasticity. Faseb J. 2020, 34, 8526–8543. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Ma, X.; Shen, B.; Huang, Y.; Birnbaumer, L.; Yao, X. TRPV4, TRPC1, and TRPP2 assemble to form a flow-sensitive heteromeric channel. Faseb J. 2014, 28, 4677–4685. [Google Scholar] [CrossRef]
- Fonfria, E.; Murdock, P.R.; Cusdin, F.S.; Benham, C.D.; Kelsell, R.E.; McNulty, S. Tissue distribution profiles of the human TRPM cation channel family. J. Recept. Signal. Transduct. Res. 2006, 26, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Haber, N.; Dina, O.A.; Chen, X.; Levine, J.D. TRPC1 and TRPC6 channels cooperate with TRPV4 to mediate mechanical hyperalgesia and nociceptor sensitization. J. Neurosci. 2009, 29, 6217–6228. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Haber, N.; Yeh, J.J.; Boyd, A.E.; Parada, C.A.; Chen, X.; Reichling, D.B.; Levine, J.D. Hypotonicity induces TRPV4-mediated nociception in rat. Neuron 2003, 39, 497–511. [Google Scholar] [CrossRef]
- Liedtke, W.; Friedman, J.M. Abnormal osmotic regulation in trpv4-/- mice. Proc. Natl. Acad. Sci. USA 2003, 100, 13698–13703. [Google Scholar] [CrossRef]
- Phelan, K.D.; Mock, M.M.; Kretz, O.; Shwe, U.T.; Kozhemyakin, M.; Greenfield, L.J.; Dietrich, A.; Birnbaumer, L.; Freichel, M.; Flockerzi, V.; et al. Heteromeric canonical transient receptor potential 1 and 4 channels play a critical role in epileptiform burst firing and seizure-induced neurodegeneration. Mol. Pharm. 2012, 81, 384–392. [Google Scholar] [CrossRef]
- Thoppil, R.J.; Adapala, R.K.; Cappelli, H.C.; Kondeti, V.; Dudley, A.C.; Gary Meszaros, J.; Paruchuri, S.; Thodeti, C.K. TRPV4 channel activation selectively inhibits tumor endothelial cell proliferation. Sci. Rep. 2015, 5, 14257. [Google Scholar] [CrossRef]
- Wu, Q.Y.; Sun, M.R.; Wu, C.L.; Li, Y.; Du, J.J.; Zeng, J.Y.; Bi, H.L.; Sun, Y.H. Activation of calcium-sensing receptor increases TRPC3/6 expression in T lymphocyte in sepsis. Mol. Immunol. 2015, 64, 18–25. [Google Scholar] [CrossRef]
- Liu, X.; Bandyopadhyay, B.C.; Singh, B.B.; Groschner, K.; Ambudkar, I.S. Molecular analysis of a store-operated and 2-acetyl-sn-glycerol-sensitive non-selective cation channel. Heteromeric assembly of TRPC1-TRPC3. J. Biol. Chem. 2005, 280, 21600–21606. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zagranichnaya, T.K.; Gurda, G.T.; Eves, E.M.; Villereal, M.L. A TRPC1/TRPC3-mediated increase in store-operated calcium entry is required for differentiation of H19-7 hippocampal neuronal cells. J. Biol. Chem. 2004, 279, 43392–43402. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Yang, F.; Takanishi, C.L.; Zheng, J. Thermosensitive TRPV channel subunits coassemble into heteromeric channels with intermediate conductance and gating properties. J. Gen. Physiol. 2007, 129, 191–207. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.K.; Yeung, S.S.; Au, S.W.; Lam, L.S.; Dai, Z.Q.; Li, Y.H.; Yeung, E.W. Expression and association of TRPC1 with TRPC3 during skeletal myogenesis. Muscle Nerve 2011, 44, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Eder, P.; Groschner, K. TRPC3/6/7: Topical aspects of biophysics and pathophysiology. Channels (Austin) 2008, 2, 94–99. [Google Scholar] [CrossRef][Green Version]
- Tano, J.Y.; Smedlund, K.; Vazquez, G. Endothelial TRPC3/6/7 proteins at the edge of cardiovascular disease. Cardiovasc. Hematol. Agents Med. Chem. 2010, 8, 76–86. [Google Scholar] [CrossRef]
- Cioffi, D.L.; Wu, S.; Chen, H.; Alexeyev, M.; St Croix, C.M.; Pitt, B.R.; Uhlig, S.; Stevens, T. Orai1 determines calcium selectivity of an endogenous TRPC heterotetramer channel. Circ. Res. 2012, 110, 1435–1444. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, X.; Tian, J.; Xiao, Y.; Tian, T.; Xu, F.; Hong, X.; Zhu, M.X. TRPC channels: Structure, function, regulation and recent advances in small molecular probes. Pharm. Therap. 2020, 209, 107497. [Google Scholar] [CrossRef]
- Myeong, J.; Ko, J.; Kwak, M.; Kim, J.; Woo, J.; Ha, K.; Hong, C.; Yang, D.; Kim, H.J.; Jeon, J.H.; et al. Dual action of the Galphaq-PLCbeta-PI(4,5)P2 pathway on TRPC1/4 and TRPC1/5 heterotetramers. Sci. Rep. 2018, 8, 12117. [Google Scholar] [CrossRef]
- Ko, J.; Myeong, J.; Shin, Y.C.; So, I. Differential PI(4,5)P2 sensitivities of TRPC4, C5 homomeric and TRPC1/4, C1/5 heteromeric channels. Sci. Rep. 2019, 9, 1849. [Google Scholar] [CrossRef]
- Kelly, J.J.; Moore, T.M.; Babal, P.; Diwan, A.H.; Stevens, T.; Thompson, W.J. Pulmonary microvascular and macrovascular endothelial cells: Differential regulation of Ca2+ and permeability. Am. J. Physiol. 1998, 274, L810–L819. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, X.; Zeng, X.; Chen, S.; Yang, X.; Zhang, L. Galectin-3 mediates pulmonary vascular endothelial cell dynamics via TRPC1/4 under acute hypoxia. J. Biochem. Mol. Toxicol. 2020, 34, e22463. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, C.; Hichami, A.; Andreoletti, P.; Cherkaoui-Malki, M.; del Mar Cavia, M.; Abdoul-Azize, S.; Alonso-Torre, S.R.; Khan, N.A. Diacylglycerol-containing oleic acid induces increases in [Ca(2+)](i) via TRPC3/6 channels in human T-cells. Biochim. Biophys. Acta 2012, 1821, 618–626. [Google Scholar] [CrossRef]
- Bijli, K.M.; Fazal, F.; Slavin, S.A.; Leonard, A.; Grose, V.; Alexander, W.B.; Smrcka, A.V.; Rahman, A. Phospholipase C-epsilon signaling mediates endothelial cell inflammation and barrier disruption in acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2016, 311, L517–L524. [Google Scholar] [CrossRef] [PubMed]
- Futosi, K.; Fodor, S.; Mocsai, A. Reprint of Neutrophil cell surface receptors and their intracellular signal transduction pathways. Int. Immunopharmacol. 2013, 17, 1185–1197. [Google Scholar] [CrossRef]
- Thebault, S.; Zholos, A.; Enfissi, A.; Slomianny, C.; Dewailly, E.; Roudbaraki, M.; Parys, J.; Prevarskaya, N. Receptor-operated Ca2+ entry mediated by TRPC3/TRPC6 proteins in rat prostate smooth muscle (PS1) cell line. J. Cell Physiol. 2005, 204, 320–328. [Google Scholar] [CrossRef]
- Onohara, N.; Nishida, M.; Inoue, R.; Kobayashi, H.; Sumimoto, H.; Sato, Y.; Mori, Y.; Nagao, T.; Kurose, H. TRPC3 and TRPC6 are essential for angiotensin II-induced cardiac hypertrophy. EMBO J. 2006, 25, 5305–5316. [Google Scholar] [CrossRef]
- Meng, K.; Xu, J.; Zhang, C.; Zhang, R.; Yang, H.; Liao, C.; Jiao, J. Calcium sensing receptor modulates extracellular calcium entry and proliferation via TRPC3/6 channels in cultured human mesangial cells. PLoS ONE 2014, 9, e98777. [Google Scholar] [CrossRef]
- Hang, P.; Zhao, J.; Cai, B.; Tian, S.; Huang, W.; Guo, J.; Sun, C.; Li, Y.; Du, Z. Brain-derived neurotrophic factor regulates TRPC3/6 channels and protects against myocardial infarction in rodents. Int. J. Biol. Sci. 2015, 11, 536–545. [Google Scholar] [CrossRef]
- Hippenstiel, S.; Krull, M.; Ikemann, A.; Risau, W.; Clauss, M.; Suttorp, N. VEGF induces hyperpermeability by a direct action on endothelial cells. Am. J. Physiol. 1998, 274, L678–L684. [Google Scholar] [CrossRef]
- Rossman, M.J.; Kaplon, R.E.; Hill, S.D.; McNamara, M.N.; Santos-Parker, J.R.; Pierce, G.L.; Seals, D.R.; Donato, A.J. Endothelial cell senescence with aging in healthy humans: Prevention by habitual exercise and relation to vascular endothelial function. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H890–H895. [Google Scholar] [CrossRef]
- Chu, Y.; Cohen, B.E.; Chuang, H.H. A single TRPV1 amino acid controls species sensitivity to capsaicin. Sci. Rep. 2020, 10, 8038. [Google Scholar] [CrossRef] [PubMed]
- Min, J.W.; Liu, W.H.; He, X.H.; Peng, B.W. Different types of toxins targeting TRPV1 in pain. Toxicon 2013, 71, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Mazgaeen, L.; Gurung, P. Recent Advances in Lipopolysaccharide Recognition Systems. Int. J. Mol. Sci. 2020, 21, 379. [Google Scholar] [CrossRef]
- Chen, S.R.; Pan, H.L. Removing TRPV1-expressing primary afferent neurons potentiates the spinal analgesic effect of delta-opioid agonists on mechano-nociception. Neuropharmacology 2008, 55, 215–222. [Google Scholar] [CrossRef]
- Nozadze, I.; Tsiklauri, N.; Gurtskaia, G.; Tsagareli, M.G. Role of thermo TRPA1 and TRPV1 channels in heat, cold, and mechanical nociception of rats. Behav. Pharm. 2016, 27, 29–36. [Google Scholar] [CrossRef]
- Zheng, J.; Liu, F.; Du, S.; Li, M.; Wu, T.; Tan, X.; Cheng, W. Mechanism for Regulation of Melanoma Cell Death via Activation of Thermo-TRPV4 and TRPV2. J. Oncol. 2019, 2019, 7362875. [Google Scholar] [CrossRef]
- Porcari, C.Y.; Debarba, L.K.; Amigone, J.L.; Caeiro, X.E.; Reis, L.C.; Cunha, T.M.; Mecawi, A.S.; Elias, L.L.; Antunes-Rodrigues, J.; Vivas, L.; et al. Brain osmo-sodium sensitive channels and the onset of sodium appetite. Horm. Behav. 2020, 118, 104658. [Google Scholar] [CrossRef]
- Li, F.; Yang, W.; Jiang, H.; Guo, C.; Huang, A.J.W.; Hu, H.; Liu, Q. TRPV1 activity and substance P release are required for corneal cold nociception. Nat. Commun. 2019, 10, 5678. [Google Scholar] [CrossRef]
- Bagher, P.; Beleznai, T.; Kansui, Y.; Mitchell, R.; Garland, C.J.; Dora, K.A. Low intravascular pressure activates endothelial cell TRPV4 channels, local Ca2+ events, and IKCa channels, reducing arteriolar tone. Proc. Natl. Acad. Sci. USA 2012, 109, 18174–18179. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.T.; Jian, M.Y.; Taylor, M.S.; Cioffi, D.L.; Yap, F.C.; Liedtke, W.; Townsley, M.I. Functional coupling of TRPV4, IK, and SK channels contributes to Ca(2+)-dependent endothelial injury in rodent lung. Pulm Circ. 2015, 5, 279–290. [Google Scholar] [CrossRef]
- Minard, A.; Bauer, C.C.; Wright, D.J.; Rubaiy, H.N.; Muraki, K.; Beech, D.J.; Bon, R.S. Remarkable Progress with Small-Molecule Modulation of TRPC1/4/5 Channels: Implications for Understanding the Channels in Health and Disease. Cells 2018, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Broker-Lai, J.; Kollewe, A.; Schindeldecker, B.; Pohle, J.; Nguyen Chi, V.; Mathar, I.; Guzman, R.; Schwarz, Y.; Lai, A.; Weissgerber, P.; et al. Heteromeric channels formed by TRPC1, TRPC4 and TRPC5 define hippocampal synaptic transmission and working memory. EMBO J. 2017, 36, 2770–2789. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, J.; Wang, L.; Liang, Y.T.; Chen, Y.Q.; Lu, W.J.; Zhou, W.L. Remodeling of rat pulmonary artery induced by chronic smoking exposure. J. Thorac Dis. 2014, 6, 818–828. [Google Scholar] [PubMed]
- Kovalevskaya, N.V.; Bokhovchuk, F.M.; Vuister, G.W. The TRPV5/6 calcium channels contain multiple calmodulin binding sites with differential binding properties. J. Struct. Funct. Genom. 2012, 13, 91–100. [Google Scholar] [CrossRef][Green Version]
- Fan, H.; Shen, Y.X.; Yuan, Y.F. Expression and prognostic roles of TRPV5 and TRPV6 in non-small cell lung cancer after curative resection. Asian Pac. J. Cancer Prev. 2014, 15, 2559–2563. [Google Scholar] [CrossRef]
- Lehen’kyi, V.; Raphael, M.; Prevarskaya, N. The role of the TRPV6 channel in cancer. J. Physiol. 2012, 590, 1369–1376. [Google Scholar] [CrossRef]
- Muller-Redetzky, H.C.; Suttorp, N.; Witzenrath, M. Dynamics of pulmonary endothelial barrier function in acute inflammation: Mechanisms and therapeutic perspectives. Cell Tissue Res. 2014, 355, 657–673. [Google Scholar] [CrossRef]
- Ince, C.; Mayeux, P.R.; Nguyen, T.; Gomez, H.; Kellum, J.A.; Ospina-Tascon, G.A.; Hernandez, G.; Murray, P.; De Backer, D.; Workgroup, A.X. The Endothelium in Sepsis. Shock 2016, 45, 259–270. [Google Scholar] [CrossRef]
- Opitz, B.; van Laak, V.; Eitel, J.; Suttorp, N. Innate immune recognition in infectious and noninfectious diseases of the lung. Am. J. Respir. Crit Care Med. 2010, 181, 1294–1309. [Google Scholar] [CrossRef]
- Imai, Y.; Kuba, K.; Neely, G.G.; Yaghubian-Malhami, R.; Perkmann, T.; van Loo, G.; Ermolaeva, M.; Veldhuizen, R.; Leung, Y.H.; Wang, H.; et al. Identification of oxidative stress and Toll-like receptor 4 signaling as a key pathway of acute lung injury. Cell 2008, 133, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Glas, G.J.; Van Der Sluijs, K.F.; Schultz, M.J.; Hofstra, J.J.; Van Der Poll, T.; Levi, M. Bronchoalveolar hemostasis in lung injury and acute respiratory distress syndrome. J. Thromb. Haemost. 2013, 11, 17–25. [Google Scholar] [CrossRef]
- Cirino, G.; Cicala, C.; Bucci, M.R.; Sorrentino, L.; Maraganore, J.M.; Stone, S.R. Thrombin functions as an inflammatory mediator through activation of its receptor. J. Exp. Med. 1996, 183, 821–827. [Google Scholar] [CrossRef]
- Takahashi, T.; Yamaguchi, S.; Chida, K.; Shibuya, M. A single autophosphorylation site on KDR/Flk-1 is essential for VEGF-A-dependent activation of PLC-gamma and DNA synthesis in vascular endothelial cells. EMBO J. 2001, 20, 2768–2778. [Google Scholar] [CrossRef]
- Tiruppathi, C.; Ahmmed, G.U.; Vogel, S.M.; Malik, A.B. Ca2+ signaling, TRP channels, and endothelial permeability. Microcirculation 2006, 13, 693–708. [Google Scholar] [CrossRef]
- Uhlig, S.; Yang, Y. Sphingolipids in acute lung injury. Handb. Exp. Pharm. 2013, 216, 227–246. [Google Scholar]
- Polat, O.K.; Uno, M.; Maruyama, T.; Tran, H.N.; Imamura, K.; Wong, C.F.; Sakaguchi, R.; Ariyoshi, M.; Itsuki, K.; Ichikawa, J.; et al. Contribution of Coiled-Coil Assembly to Ca(2+)/Calmodulin-Dependent Inactivation of TRPC6 Channel and its Impacts on FSGS-Associated Phenotypes. J. Am. Soc. Nephrol. 2019, 30, 1587–1603. [Google Scholar] [CrossRef]
- Singh, B.B.; Liu, X.; Tang, J.; Zhu, M.X.; Ambudkar, I.S. Calmodulin regulates Ca(2+)-dependent feedback inhibition of store-operated Ca(2+) influx by interaction with a site in the C terminus of TrpC1. Mol. Cell 2002, 9, 739–750. [Google Scholar] [CrossRef]
- Antoniotti, S.; Lovisolo, D.; Fiorio Pla, A.; Munaron, L. Expression and functional role of bTRPC1 channels in native endothelial cells. Febs Lett. 2002, 510, 189–195. [Google Scholar] [CrossRef]
- Keeble, J.; Russell, F.; Curtis, B.; Starr, A.; Pinter, E.; Brain, S.D. Involvement of transient receptor potential vanilloid 1 in the vascular and hyperalgesic components of joint inflammation. Arthritis Rheum. 2005, 52, 3248–3256. [Google Scholar] [CrossRef] [PubMed]
- Strotmann, R.; Schultz, G.; Plant, T.D. Ca2+-dependent potentiation of the nonselective cation channel TRPV4 is mediated by a C-terminal calmodulin binding site. J. Biol. Chem. 2003, 278, 26541–26549. [Google Scholar] [CrossRef]
- Tauseef, M.; Knezevic, N.; Chava, K.R.; Smith, M.; Sukriti, S.; Gianaris, N.; Obukhov, A.G.; Vogel, S.M.; Schraufnagel, D.E.; Dietrich, A.; et al. TLR4 activation of TRPC6-dependent calcium signaling mediates endotoxin-induced lung vascular permeability and inflammation. J. Exp. Med. 2012, 209, 1953–1968. [Google Scholar] [CrossRef]
- Ufret-Vincenty, C.A.; Klein, R.M.; Hua, L.; Angueyra, J.; Gordon, S.E. Localization of the PIP2 sensor of TRPV1 ion channels. J. Biol. Chem. 2011, 286, 9688–9698. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.T.; Liu, X.; Ong, H.L.; Swaim, W.; Ambudkar, I.S. Local Ca(2)+ entry via Orai1 regulates plasma membrane recruitment of TRPC1 and controls cytosolic Ca(2)+ signals required for specific cell functions. PLoS Biol. 2011, 9, e1001025. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhang, Y.; Wang, M.; Zhang, H.; Chen, Y.; Adcock, I.M.; Chung, K.F.; Mo, J.; Zhang, Y.; Li, F. TRPV1 and TRPA1 in Lung Inflammation and Airway Hyperresponsiveness Induced by Fine Particulate Matter (PM2.5). Oxid. Med. Cell Longev. 2019, 2019, 7450151. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Li, Z.P.; Li, M.X.; Li, D.W.; Ye, H.B.; Su, K.M.; Lin, H.; Zhang, W.T. Pro-inflammatory role of transient receptor potential canonical channel 6 in the pathogenesis of chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2018, 8, 1334–1341. [Google Scholar] [CrossRef] [PubMed]
- Sel, S.; Rost, B.R.; Yildirim, A.O.; Sel, B.; Kalwa, H.; Fehrenbach, H.; Renz, H.; Gudermann, T.; Dietrich, A. Loss of classical transient receptor potential 6 channel reduces allergic airway response. Clin. Exp. Allergy 2008, 38, 1548–1558. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Method | Application | Limitation | Example |
|---|---|---|---|
| AFM (atomic force microscopy) [36] | for stoichiometric determination in cells expressing tagged channels; allows for detection at the single channel level as it yields three dimensional structures in an Ångström (Å) range | not applicable for electrophysiological analysis | TRPP2/V4 [34] |
| Co-IP (co-immuno- precipitation) [37] | proof of interactions between TRP channel subunits in cells expressing tagged channels; allows for differentiation of heteromeric or homomeric assemblies | not applicable for structural and electrophysiological analysis | TRPV1/4 [38] |
| cross-linking in combination with mass spectrometry [39] | proof of interactions between TRP channel subunits and for structural and stoichiometric determination in native cells; provides a 3D-structure of interacting proteins, the distance between TRP channel subunits in the cross-linking reaction depends on the length of the crosslinker | not applicable for electrophysiological analysis | no applications made for heteromeric TRP channels yet |
| cryo-electron microscopy [40] | for structural analysis in cells overexpressing channels; allows for detection at the single channel level as it shows three dimensional structures, but sensitivity depends on the investigated protein’s electron irradiation and scattering | not applicable for electrophysiological analysis | no applications made for heteromeric TRP channels yet |
| FRET (Förster resonance energy transfer) [41] | proof of interactions between fluorescently tagged TRP channel subunits by live cell imaging; quantifies interaction over short distances (10–100 Å), yet fluorophores can alter binding properties | not applicable for structural, stoichiometric and electrophysiological analysis | TRPV1/4 [38] |
| molecular docking [31] | for prediction of structural domains that stabilize the TRP heteromer; method is based on acquired input data | not applicable for electrophysiological analysis | no applications made for heteromeric TRP channels yet |
| PLA (proximity ligation assay) [42] | proof of interactions between TRP channel subunits in native cells; quantifies interactions in a 30-nm range, allows quantification of heteromeric channel assemblies, but not at the single channel level; strongly depends on antibody specificity | not applicable for structural, stoichiometric and electrophysiological analysis | TRPV1/4 [27] |
| whole-cell patch-clamp recordings [43] | for electrophysiological analysis in native cells or cells expressing tagged channels; measures single ion channel current in a picoampere range | not applicable for structural and stoichiometric determinations | TRPC1/4 [44] |
| SC-SMD (single channel single molecule determinations) [45] | proof of interactions between TRP channel subunits and for structural and stoichiometric analysis in cells expressing tagged channels; detection at the single channel level as it yields three dimensional structures of proteins in a nanometer range | not applicable for electrophysiological analysis | no applications made for heteromeric TRP channels yet |
| x-ray crystallography [46] | for structural and stoichiometric analysis in cells overexpressing channels; detection at the single channel level as it yields three dimensional structures of molecules in an (Å) range, but is difficult to perform for transmembrane proteins | not applicable for electrophysiological analysis | no applications made for heteromeric TRP channels |
| Y2H (yeast two-hybrid technique) [47] | proof of interactions between TRP channel subunits in living native cells; detection of protein interactions by yeast reproduction, but not on a single channel level, difficult application for transmembrane proteins, and high rate of false positive results | not applicable for structural, stoichiometric and electrophysiological analysis | Drosophila retinal-specific TRP/L [29] |
| Receptor | Function in Lung Inflammation | Involved TRP Channels |
|---|---|---|
| TLR4 (Toll-like receptor) | activation of NF-κB signaling pathway and thus initiation of cytokine production; DAG production [111] | DAG production activates TRPC1/4, TRPC3/6, and TRPC6 [1,81,84] |
| PAR1 (protease-activated receptor 1) | secretion of VEGF and formation of IP3 [91,114] | activation of TRPC1/4 and TRPC3/6 via IP3-dependent Ca2+ release from the endoplasmic reticulum [84,115] |
| VEGFR (vascular endothelial growth factor receptor) | DAG production [115] | DAG production activates TRPC1/4, TRPC3/6, and TRPC6 [84,104,123] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zergane, M.; Kuebler, W.M.; Michalick, L. Heteromeric TRP Channels in Lung Inflammation. Cells 2021, 10, 1654. https://doi.org/10.3390/cells10071654
Zergane M, Kuebler WM, Michalick L. Heteromeric TRP Channels in Lung Inflammation. Cells. 2021; 10(7):1654. https://doi.org/10.3390/cells10071654
Chicago/Turabian StyleZergane, Meryam, Wolfgang M. Kuebler, and Laura Michalick. 2021. "Heteromeric TRP Channels in Lung Inflammation" Cells 10, no. 7: 1654. https://doi.org/10.3390/cells10071654
APA StyleZergane, M., Kuebler, W. M., & Michalick, L. (2021). Heteromeric TRP Channels in Lung Inflammation. Cells, 10(7), 1654. https://doi.org/10.3390/cells10071654