
Dual Impact of a Benzimidazole Resistant β-Tubulin on Microtubule Behavior in Fission Yeast



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Genetics
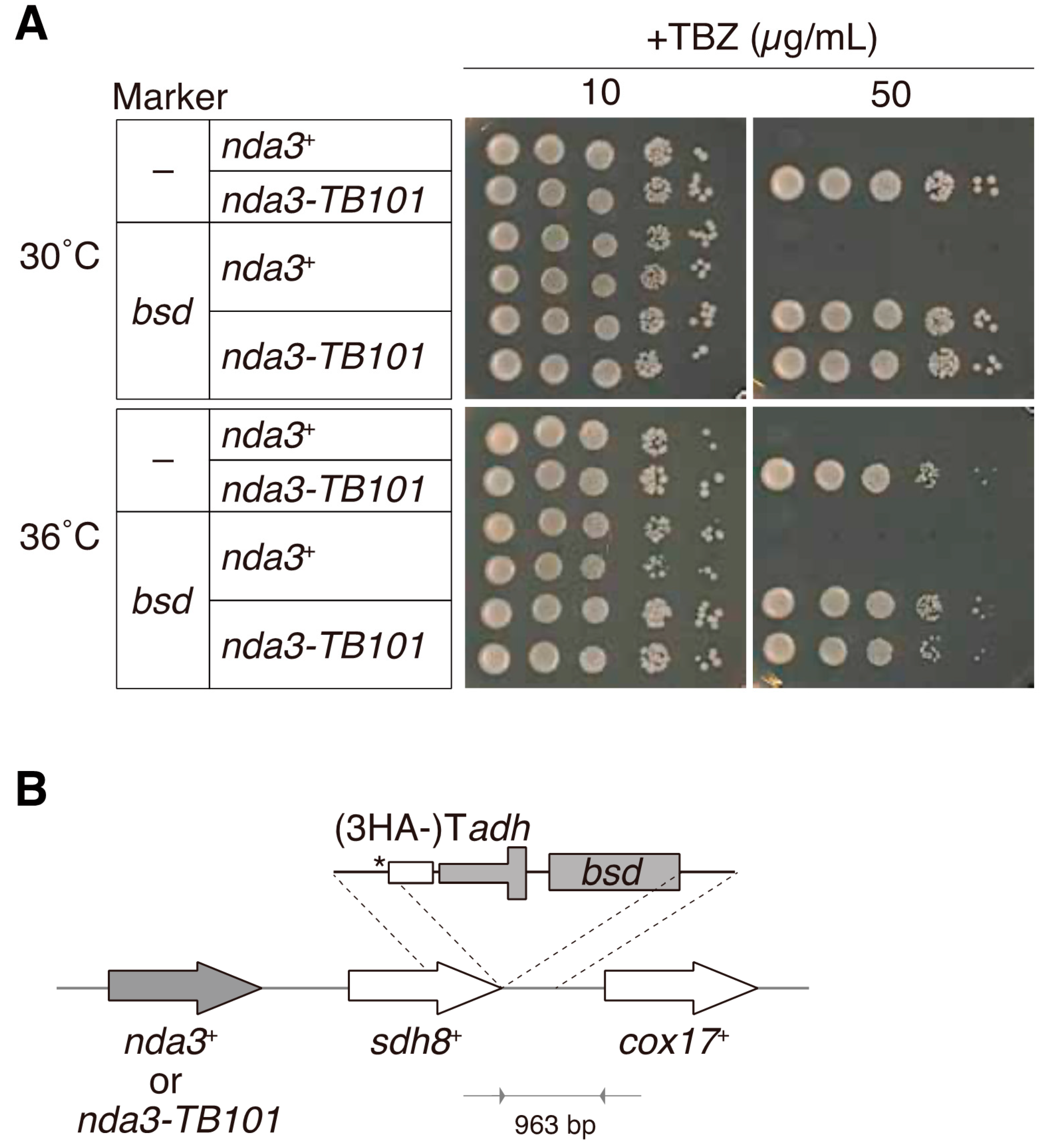
2.2. Insertion of the Blasticidin S Resistance Gene
2.3. Prediction and Modelling of the Protein Structure
2.4. Microscopy
2.5. Drug Assays
2.6. Cold-Shock Assays
2.7. In Vitro TBZ Binding Assays
2.8. Statistical Analysis
3. Results
3.1. The nda3-TB101 Mutant Is Resistant to TBZ and MBC
3.2. Nda3-TB101 Has a Mutation Near the H1′-S2 Loop of β-Tubulin
3.3. Two Possible Modes for Y50S β-Tubulin
3.4. Microtubules Containing Nda3-TB101 Are Unaffected by TBZ and MBC
3.5. The nda3-TB101 Mutant Is Resistant to Nocodazole
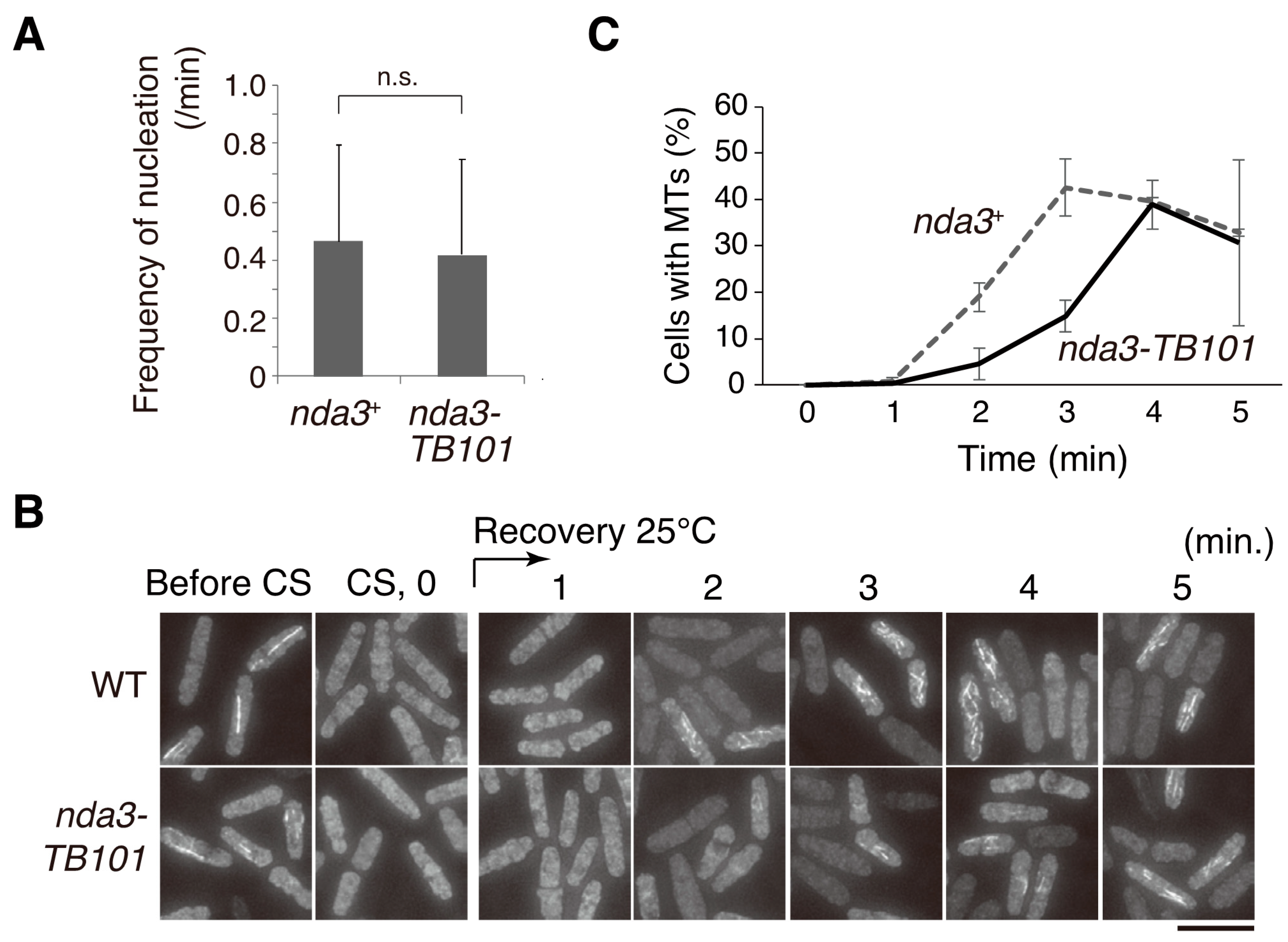
3.6. Microtubule Dynamics in nda3-TB101 Were Moderately Stabilized in the Absence of Drugs
3.7. Microtubule Nucleation Was Not Affected by the nda3-TB101 Mutation
3.8. The nda3-TB101 Mutation Suppressed the TBZ Sensitivity of the alp14 Mutant
3.9. Phenotypic Differences in klp5/6 and nda3-TB101 Mutants
3.10. The Y50S Mutation of β-Tubulin Interferes Binding of TBZ
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nogales, E.; Wolf, S.G.; Downing, K.H. Structure of the αβ tubulin dimer by electron crystallography. Nature 1998, 391, 199–203. [Google Scholar] [CrossRef]
- Borisy, G.; Heald, R.; Howard, J.; Janke, C.; Musacchio, A.; Nogales, E. microtubules: 50 years on from the discovery of tubulin. Nat. Rev. Mol. Cell Biol. 2016, 17, 322–328. [Google Scholar] [CrossRef]
- Desai, A.; Mitchison, T.J. Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. 1997, 13, 83–117. [Google Scholar] [CrossRef]
- Maiato, H.; Sampaio, P.; Sunkel, C.E. Microtubule-associated proteins and their essential roles during mitosis. Int. Rev. Cytol. 2004, 241, 53–153. [Google Scholar] [PubMed]
- Maurer, S.P.; Bieling, P.; Cope, J.; Hoenger, A.; Surrey, T. GTPγS microtubules mimic the growing microtubule end structure recognized by end-binding proteins (EBs). Proc. Natl. Acad. Sci. USA 2011, 108, 3988–3993. [Google Scholar] [CrossRef] [PubMed]
- Maurer, S.P.; Fourniol, F.J.; Bohner, G.; Moores, C.A.; Surrey, T. EBs recognize a nucleotide-dependent structural cap at growing microtubule ends. Cell 2012, 149, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Brouhard, G.J.; Stear, J.H.; Noetzel, T.L.; Al-Bassam, J.; Kinoshita, K.; Harrison, S.C.; Howard, J.; Hyman, A.A. XMAP215 is a processive microtubule polymerase. Cell 2008, 132, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, K.; Kurooka, H.; Takeuchi, M.; Kinoshita, K.; Nakaseko, Y.; Yanagida, M. P93dis1, which is required for sister chromatid separation, is a novel microtubule and spindle pole body-associating protein phosphorylated at the Cdc2 target sites. Genes Dev. 1995, 9, 1572–1585. [Google Scholar] [CrossRef]
- Nakaseko, Y.; Goshima, G.; Morishita, J.; Yanagida, M. M phase-specific kinetochore proteins in fission yeast: Microtu-bule-associating Dis1 and Mtc1 display rapid separation and segregation during anaphase. Curr. Biol. 2001, 11, 537–549. [Google Scholar] [CrossRef]
- Garcia, M.A.; Vardy, L.; Koonrugsa, N.; Toda, T. Fission yeast ch-TOG/XMAP215 homologue Alp14 connects mitotic spindles with the kinetochore and is a component of the Mad2-dependent spindle checkpoint. EMBO J. 2001, 20, 3389–3401. [Google Scholar] [CrossRef]
- Al-Bassam, J.; Kim, H.; Flor-Parra, I.; Lal, N.; Velji, H.; Chang, F. Fission yeast Alp14 is a dose-dependent plus end–tracking microtubule polymerase. Mol. Biol. Cell 2012, 23, 2878–2890. [Google Scholar] [CrossRef]
- Walczak, C.E.; Mitchison, T.J.; Desai, A. XKCM1: A xenopus kinesin-related protein that regulates microtubule dynamics during mitotic spindle assembly. Cell 1996, 84, 37–47. [Google Scholar] [CrossRef]
- Desai, A.; Verma, S.; Mitchison, T.J.; Walczak, C.E. Kin I kinesins are microtubule-destabilizing enzymes. Cell 1999, 96, 69–78. [Google Scholar] [CrossRef]
- Howard, J.; Hyman, A.A. Microtubule polymerases and depolymerases. Curr. Opin. Cell Biol. 2007, 19, 31–35. [Google Scholar] [CrossRef] [PubMed]
- West, R.R.; Malmstrom, T.; Troxell, C.L.; McIntosh, J.R. Two related kinesins, klp5+andklp6+, foster microtubule disassembly and are required for meiosis in fission yeast. Mol. Biol. Cell 2001, 12, 3919–3932. [Google Scholar] [CrossRef]
- Garcia, M.A.; Koonrugsa, N.; Toda, T. Two kinesin-like kin I family proteins in fission yeast regulate the establishment of metaphase and the onset of anaphase A. Curr. Biol. 2002, 12, 610–621. [Google Scholar] [CrossRef]
- Grissom, P.M.; Fiedler, T.; Grishchuk, E.L.; Nicastro, D.; West, R.R.; McIntosh, J.R. Kinesin-8 from fission yeast: A hetero-dimeric, plus-end-directed motor that can couple microtubule depolymerization to cargo movement. Mol. Biol. Cell 2009, 20, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Davidse, L.C. Benzimidazole fungicides: Mechanism of action and biological impact. Annu. Rev. Phytopathol. 1986, 24, 43–65. [Google Scholar] [CrossRef]
- Iwasaki, S. Antimitotic agents: Chemistry and recognition of tubulin molecule. Med. Res. Rev. 1993, 13, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.K.; Oakley, B.R. Identification of an amino acid substitution in the benA, β-tubulin gene of Aspergillus nidulans that confers thiabendazole resistance and benomyl supersensitivity. Cell Motil. Cytoskelet. 1990, 17, 87–94. [Google Scholar] [CrossRef]
- Jung, M.K.; Wilder, I.B.; Oakley, B.R. Amino acid alterations in the benA (β-tubulin) gene of Aspergillus nidulans that confer benomyl resistance. Cell Motil. Cytoskelet. 1992, 22, 170–174. [Google Scholar] [CrossRef]
- Fujimura, M.; Oeda, K.; Inoue, H.; Kato, T. A single amino-acid substitution in the beta-tubulin gene of Neurospora confers both carbendazim resistance and diethofencarb sensitivity. Curr. Genet. 1992, 21, 399–404. [Google Scholar] [CrossRef]
- Vela-Corcía, D.; Bellón-Gómez, D.; López-Ruiz, F.; Torés, J.A.; Pérez-García, A. The Podosphaera fusca TUB2 gene, a molecular “Swiss Army knife” with multiple applications inpowdery mildew research. Fungal Biol. 2014, 118, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Yarden, O.; Katan, T. Mutations leading to substitutions at amino acids 198 and 200 of beta-tubulin that correlate with be-nomyl-resistance phenotypes of field strains of Botrytis cinerea. Phytopathology 1993, 83, 1478–1483. [Google Scholar] [CrossRef]
- Reijo, R.A.; Cooper, E.M.; Beagle, G.J.; Huffaker, T.C. Systematic mutational analysis of the yeast β-tubulin gene. Mol. Biol. Cell 1994, 5, 29–43. [Google Scholar] [CrossRef][Green Version]
- Albertini, C.; Gredt, M.; Leroux, P. Mutations of the β-Tubulin gene associated with different phenotypes of benzimidazole resistance in the cereal eyespot fungi Tapesia yallundae and Tapesia acuformis. Pestic. Biochem. Physiol. 1999, 64, 17–31. [Google Scholar] [CrossRef]
- Kwa, M.S.; Veenstra, J.G.; Roos, M.H. Benzimidazole resistance in Haemonchus contortus is correlated with a conserved mutation at amino acid 200 in β-tubulin isotype 1. Mol. Biochem. Parasitol. 1994, 63, 299–303. [Google Scholar] [CrossRef]
- Prichard, R. Genetic variability following selection of Haemonchus contortus with anthelmintics. Trends Parasitol. 2001, 17, 445–453. [Google Scholar] [CrossRef]
- Toda, T.; Adachi, Y.; Hiraoka, Y.; Yanagida, M. Identification of the pleiotropic cell division cycle gene NDA2 as one of two different α-tubulin genes in schizosaccharomyces pombe. Cell 1984, 37, 233–241. [Google Scholar] [CrossRef]
- Adachi, Y.; Toda, T.; Niwa, O.; Yanagida, M. Differential expressions of essential and nonessential α-tubulin genes in Schiz-osaccharomyces pombe. Mol. Cell Biol. 1986, 6, 2168–2178. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, Y.; Toda, T.; Yanagida, M. The NDA3 gene of fission yeast encodes β-tubulin: A cold-sensitive nda3 mutation re-versibly blocks spindle formation and chromosome movement in mitosis. Cell 1984, 39, 349–358. [Google Scholar] [CrossRef]
- Yamamoto, M. Genetic analysis of resistant mutants to antimitotic benzimidazole compounds in Schizosaccharomyces pombe. Mol. Genet. Genom. 1980, 180, 231–234. [Google Scholar] [CrossRef]
- Umesono, K.; Toda, T.; Hayashi, S.; Yanagida, M. Two cell division cycle genes NDA2 and NDA3 of the fission yeast Schiz-osaccharomyces pombe control microtubular organization and sensitivity to anti-mitotic benzimidazole compounds. J. Mol. Biol. 1983, 168, 271–284. [Google Scholar] [CrossRef]
- Vela-Corcía, D.; Romero, D.; De Vicente, A.; Pérez-García, A. Analysis of β-tubulin-carbendazim interaction reveals that binding site for MBC fungicides does not include residues involved in fungicide resistance. Sci. Rep. 2018, 8, 7161. [Google Scholar] [CrossRef] [PubMed]
- Moreno, S.; Klar, A.; Nurse, P. [56] Molecular genetic analysis of fission yeast Schizosaccharomyces pombe. Methods Enzymol. 1991, 194, 795–823. [Google Scholar] [CrossRef]
- Bähler, J.; Wu, J.-Q.; Longtine, M.S.; Shah, N.G.; McKenzie, A.; Steever, A.B.; Wach, A.; Philippsen, P.; Pringle, J.R. Heter-ologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 1998, 14, 943–951. [Google Scholar] [CrossRef]
- Sato, M.; Dhut, S.; Toda, T. New drug-resistant cassettes for gene disruption and epitope tagging inSchizosaccharomyces pombe. Yeast 2005, 22, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Toya, M.; Toda, T. Visualization of fluorescence-tagged proteins in fission yeast: The analysis of mitotic spindle dynamics using GFP-tubulin under the native promoter. Methods Mol. Biol. 2009, 545, 185–203. [Google Scholar] [PubMed]
- Kelley, L.A.; Sternberg, M.J. Protein structure prediction on the Web: A case study using the Phyre server. Nat. Protoc. 2009, 4, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Ohta, M.; Sato, M.; Yamamoto, M. Spindle pole body components are reorganized during fission yeast meiosis. Mol. Biol. Cell 2012, 23, 1799–1811. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for bio-logical-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Aoi, Y.; Sato, M.; Sutani, T.; Shirahige, K.; Kapoor, T.M.; Kawashima, S.A. Dissecting the first and the second meiotic divisions using a marker-less drug-hypersensitive fission yeast. Cell Cycle 2014, 13, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Jalali, F.; Dorraji, P.S. Interaction of anthelmintic drug (thiabendazole) with DNA: Spectroscopic and molecular modeling studies. Arab. J. Chem. 2017, 10, S3947–S3954. [Google Scholar] [CrossRef]
- Nogales, E.; Whittaker, M.; Milligan, R.A.; Downing, K.H. High-Resolution Model of the Microtubule. Cell 1999, 96, 79–88. [Google Scholar] [CrossRef]
- Sui, H.; Downing, K.H. Structural basis of interprotofilament interaction and lateral deformation of microtubules. Structure 2010, 18, 1022–1031. [Google Scholar] [CrossRef]
- Zhang, R.; Alushin, G.M.; Brown, A.; Nogales, E. Mechanistic origin of microtubule dynamic instability and its modulation by eb proteins. Cell 2015, 162, 849–859. [Google Scholar] [CrossRef]
- Robinson, M.W.; McFerran, N.; Trudgett, A.; Hoey, L.; Fairweather, I. A possible model of benzimidazole binding to β-tubulin disclosed by invoking an inter-domain movement. J. Mol. Graph. Model. 2004, 23, 275–284. [Google Scholar] [CrossRef]
- Sawin, K.E.; Snaith, H.A. Role of microtubules and tea1p in establishment and maintenance of fission yeast cell polarity. J. Cell Sci. 2004, 117, 689–700. [Google Scholar] [CrossRef]
- Kawashima, S.A.; Takemoto, A.; Nurse, P.; Kapoor, T.M. Analyzing Fission Yeast Multidrug Resistance Mechanisms to Develop a Genetically Tractable Model System for Chemical Biology. Chem. Biol. 2012, 19, 893–901. [Google Scholar] [CrossRef]
- Tran, P.; Marsh, L.; Doye, V.; Inoué, S.; Chang, F. A Mechanism for nuclear positioning in fission yeast based on microtubule pushing. J. Cell Biol. 2001, 153, 397–412. [Google Scholar] [CrossRef]
- Busch, K.E.; Brunner, D. The microtubule plus end-tracking proteins mal3p and tip1p cooperate for cell-end targeting of interphase microtubules. Curr. Biol. 2004, 14, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Beinhauer, J.D.; Hagan, I.M.; Hegemann, J.H.; Fleig, U. Mal3, the fission yeast homologue of the human APC-interacting protein EB-1 is required for microtubule integrity and the maintenance of cell form. J. Cell Biol. 1997, 139, 717–728. [Google Scholar] [CrossRef]
- Kikkawa, M.; Ishikawa, T.; Nakata, T.; Wakabayashi, T.; Hirokawa, N. Direct visualization of the microtubule lattice seam both in vitro and in vivo. J. Cell Biol. 1994, 127, 1965–1971. [Google Scholar] [CrossRef]
- Linse, K.; Mandelkow, E.M. The GTP-binding peptide of β-tubulin. Localization by direct photoaffinity labeling and com-parison with nucleotide-binding proteins. J. Biol. Chem. 1988, 263, 15205–15210. [Google Scholar] [CrossRef]
- Des Georges, A.; Katsuki, M.; Drummond, D.R.; Osei, M.; Cross, R.A.; Amos, L.A. Mal3, the Schizosaccharomyces pombe homolog of EB1, changes the microtubule lattice. Nat. Struct. Mol. Biol. 2008, 15, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Zanic, M.; Stear, J.H.; Hyman, A.A.; Howard, J. EB1 recognizes the nucleotide state of tubulin in the microtubule lattice. PLoS ONE 2009, 4, e7585. [Google Scholar] [CrossRef] [PubMed]
- Downing, K.H. Structural basis for the action of drugs that affect microtubule dynamics. Emerg. Ther. Targets 2000, 4, 219–237. [Google Scholar] [CrossRef]
- Guzmán-Ocampo, D.C.; Aguayo-Ortiz, R.; Cano-González, L.; Castillo, R.; Hernández-Campos, A.; Dominguez, L. Effects of the Protonation State of Titratable Residues and the Presence of Water Molecules on Nocodazole Binding to β-Tubulin. Chem. Med. Chem. 2018, 13, 20–24. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minagawa, M.; Shirato, M.; Toya, M.; Sato, M. Dual Impact of a Benzimidazole Resistant β-Tubulin on Microtubule Behavior in Fission Yeast. Cells 2021, 10, 1042. https://doi.org/10.3390/cells10051042
Minagawa M, Shirato M, Toya M, Sato M. Dual Impact of a Benzimidazole Resistant β-Tubulin on Microtubule Behavior in Fission Yeast. Cells. 2021; 10(5):1042. https://doi.org/10.3390/cells10051042
Chicago/Turabian StyleMinagawa, Mamika, Minamo Shirato, Mika Toya, and Masamitsu Sato. 2021. "Dual Impact of a Benzimidazole Resistant β-Tubulin on Microtubule Behavior in Fission Yeast" Cells 10, no. 5: 1042. https://doi.org/10.3390/cells10051042
APA StyleMinagawa, M., Shirato, M., Toya, M., & Sato, M. (2021). Dual Impact of a Benzimidazole Resistant β-Tubulin on Microtubule Behavior in Fission Yeast. Cells, 10(5), 1042. https://doi.org/10.3390/cells10051042