
Immunomodulatory Properties of Polysaccharides from the Coral Pseudopterogorgia americana in Macrophages

 ,
,  ,
, 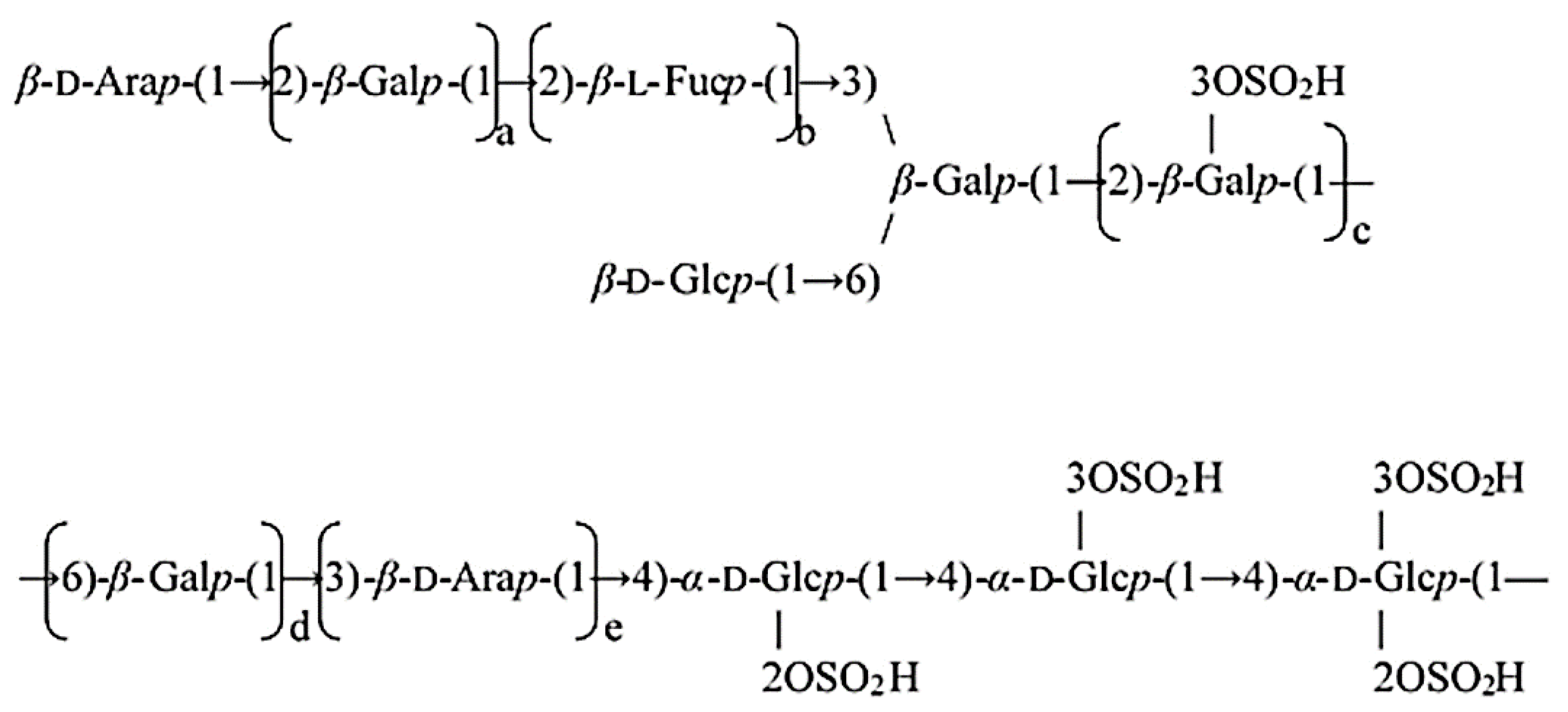{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Cell Cultures
2.3. Isolation of PPA
2.4. Activation of Macrophages by PPA and Bacteria
2.5. Detection of Cytokines
2.6. Detection of Cytokines and NO
2.7. Analysis of NF-κB Activity
2.8. Detection of ROS Generation
2.9. Phagocytosis Assay
2.10. Analysis of mRNA Levels of PKC
2.11. Statistical Analysis
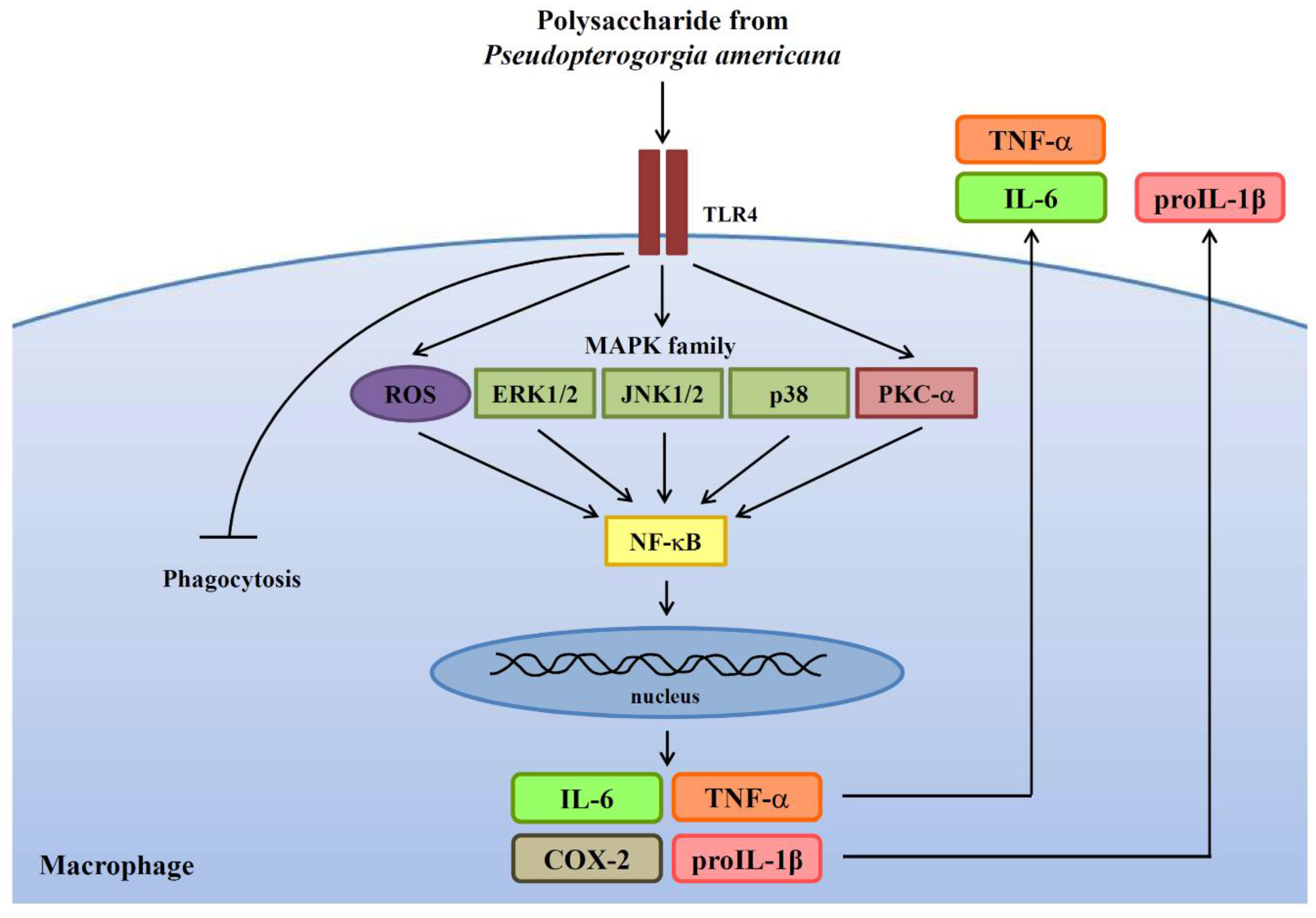
3. Results
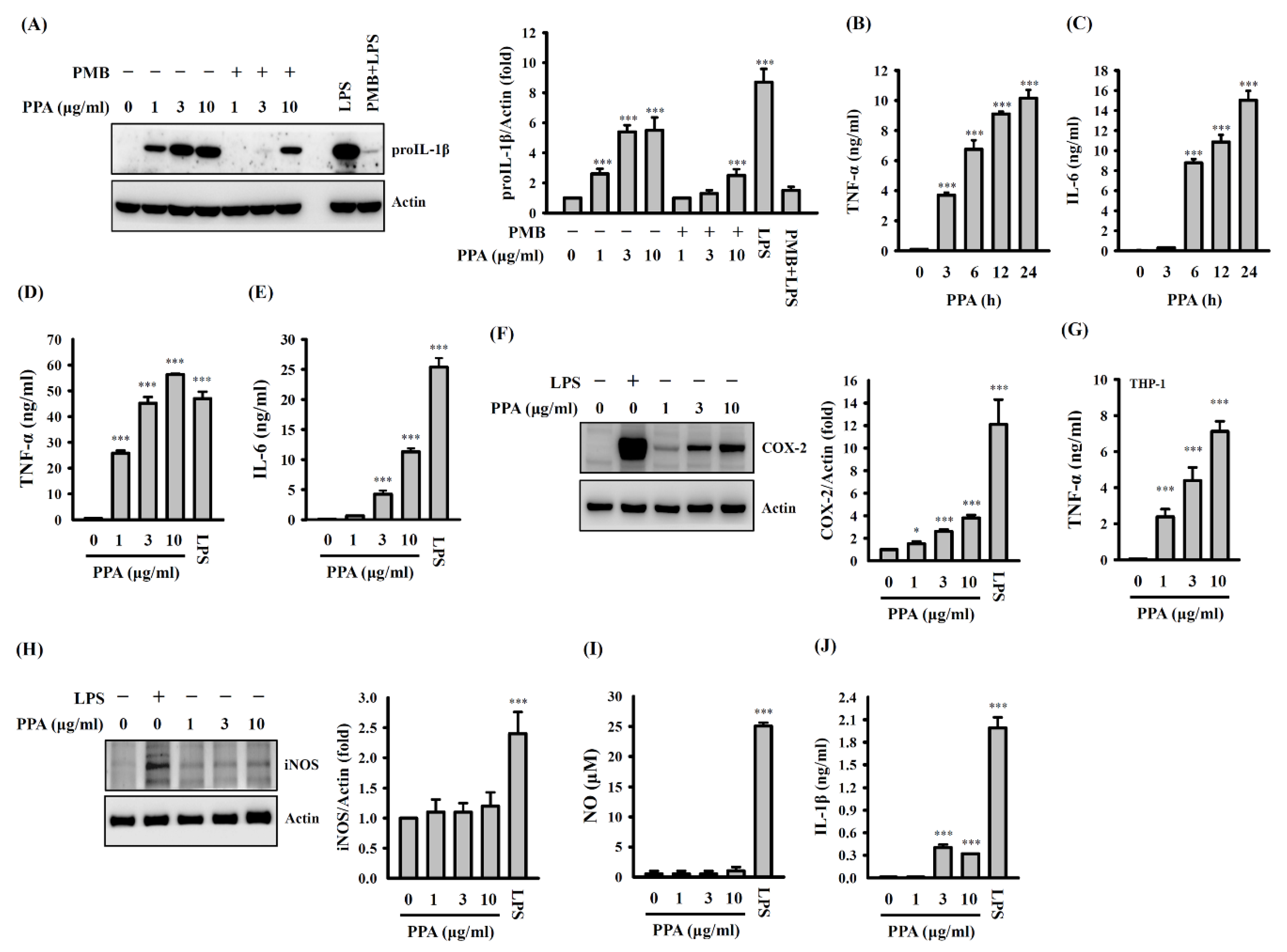
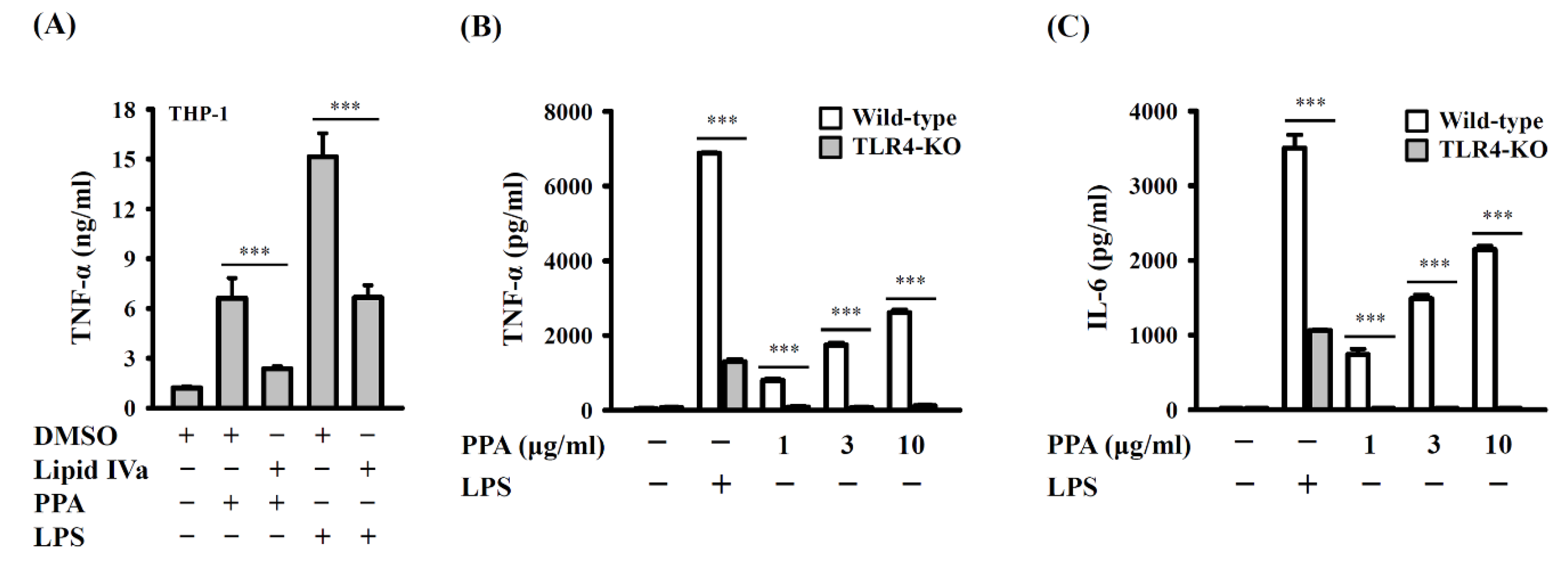
3.1. PPA Activates Macrophages Independent of LPS Contamination
3.2. PPA Activates Macrophages through TLR4
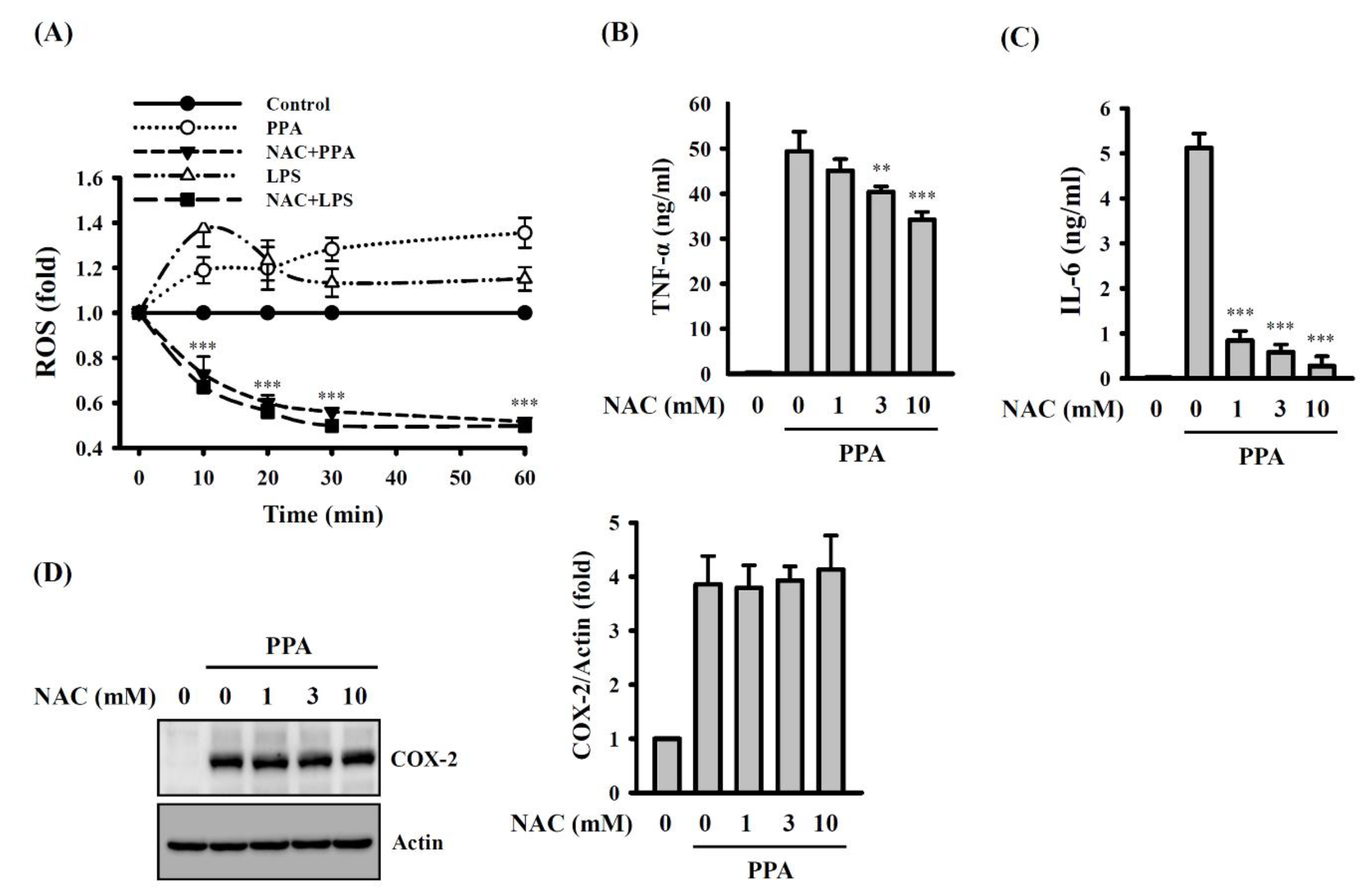
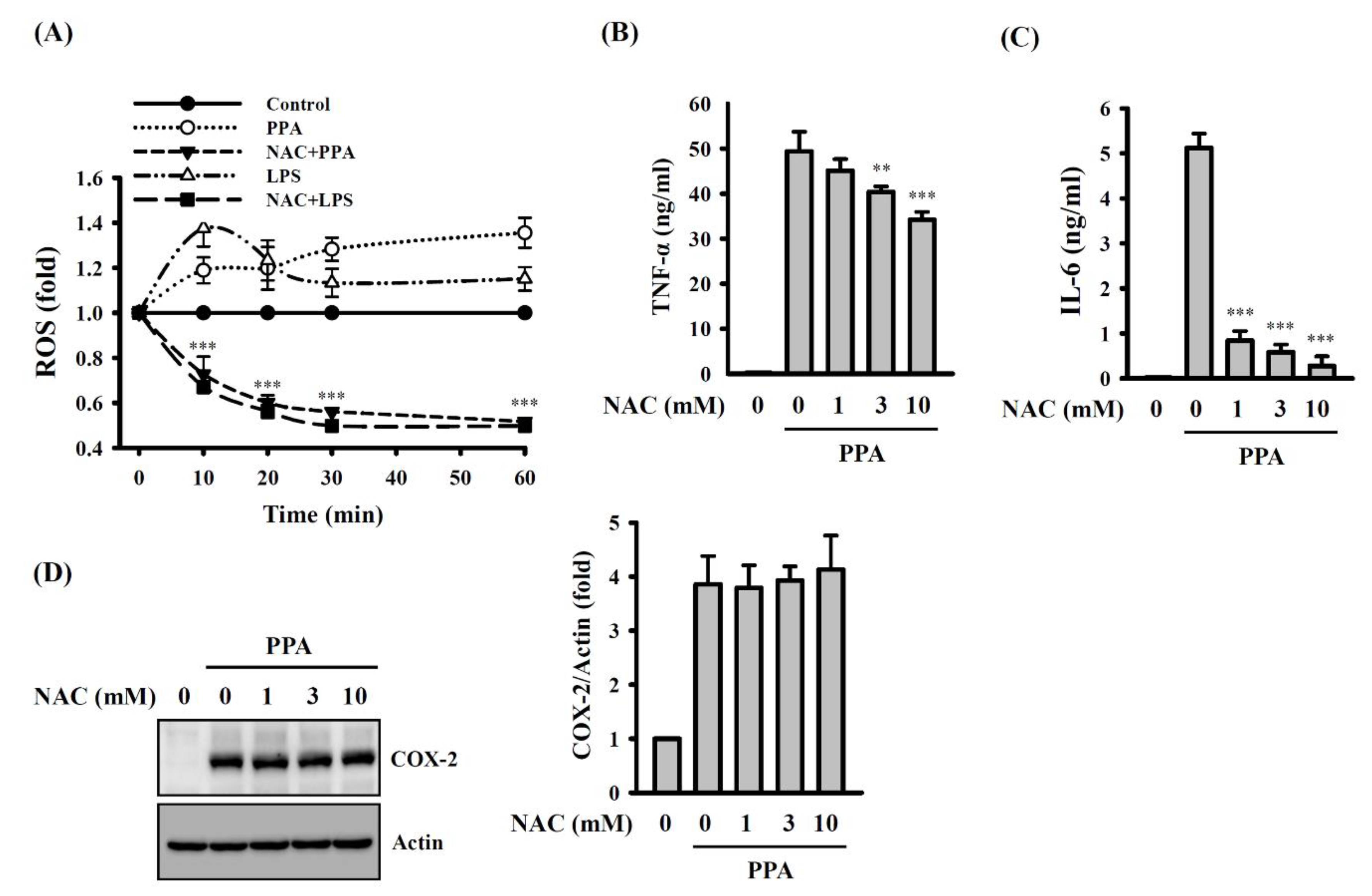
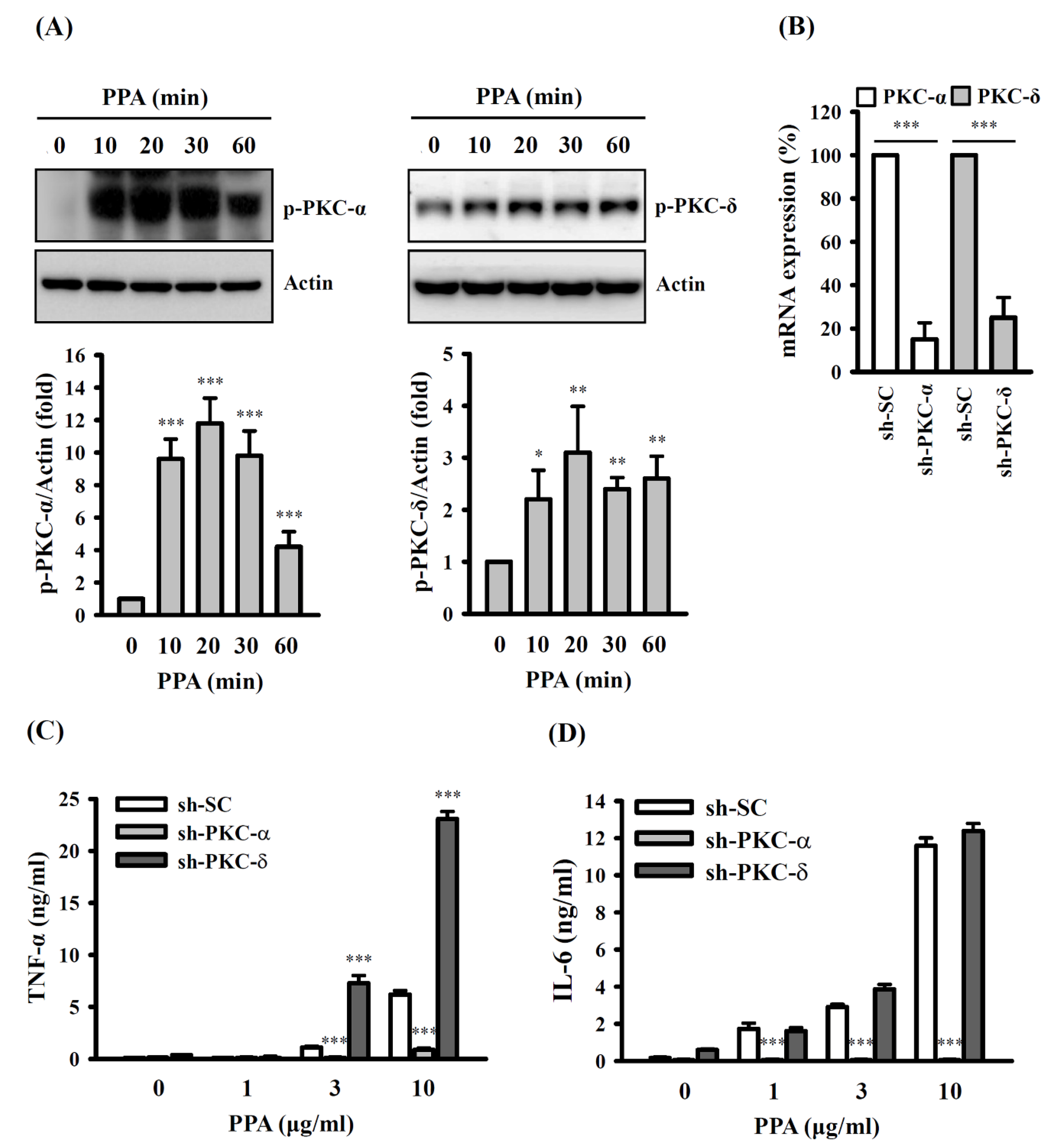
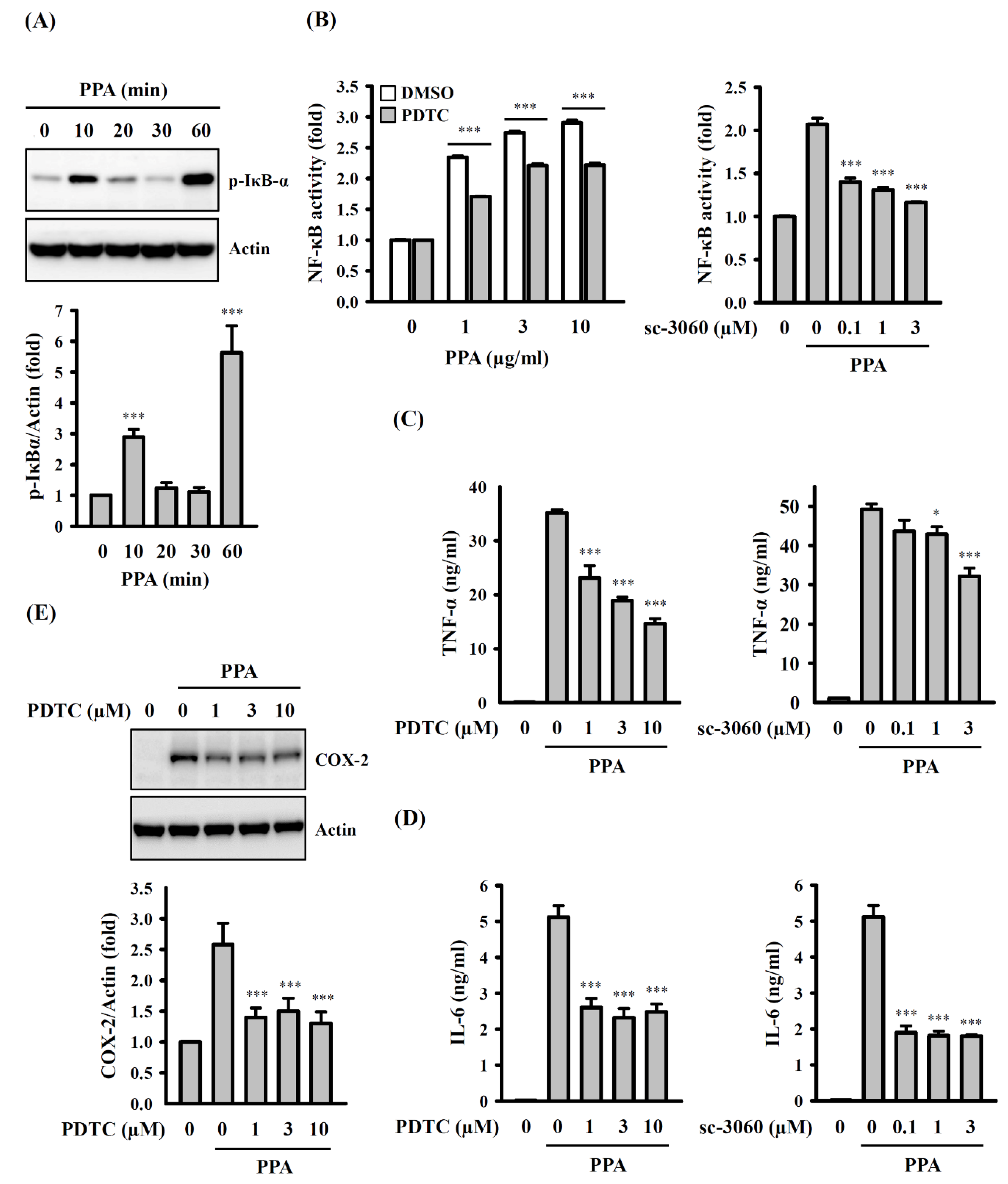
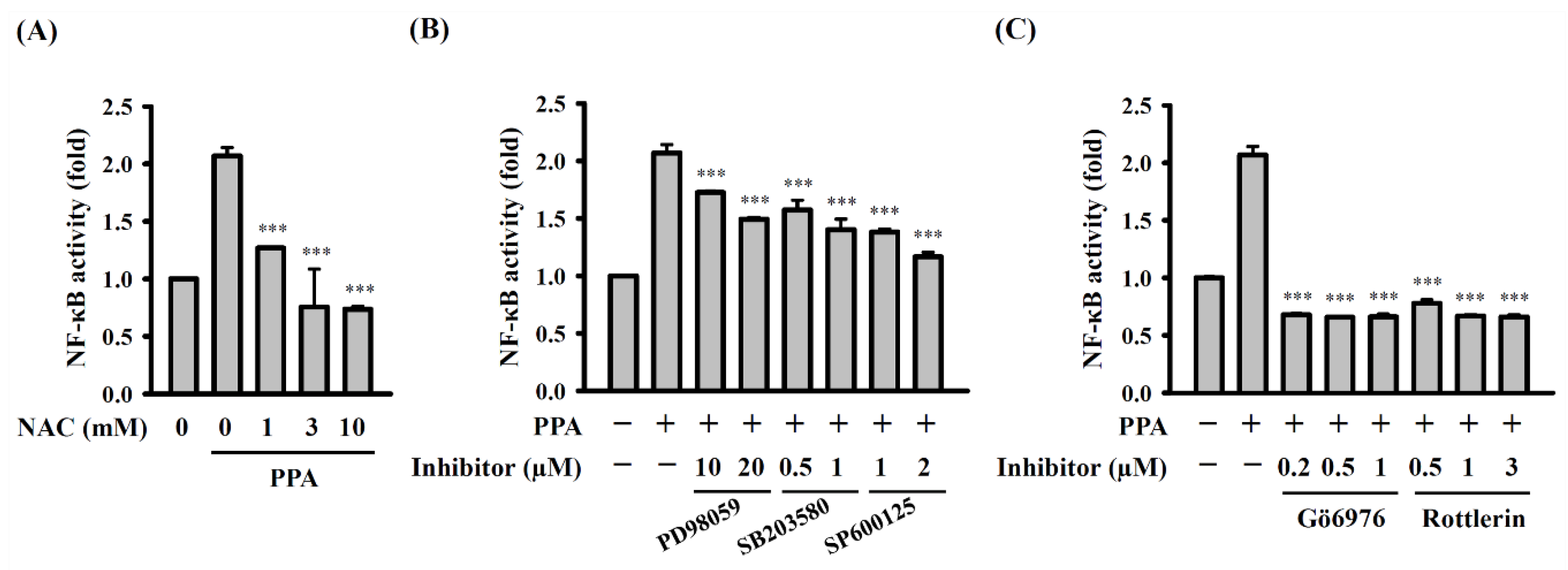
3.3. PPA-Induced TNF-A and IL-6 Expression through Reactive Oxygen Species (ROS) and Protein Kinase C (PKC)-A
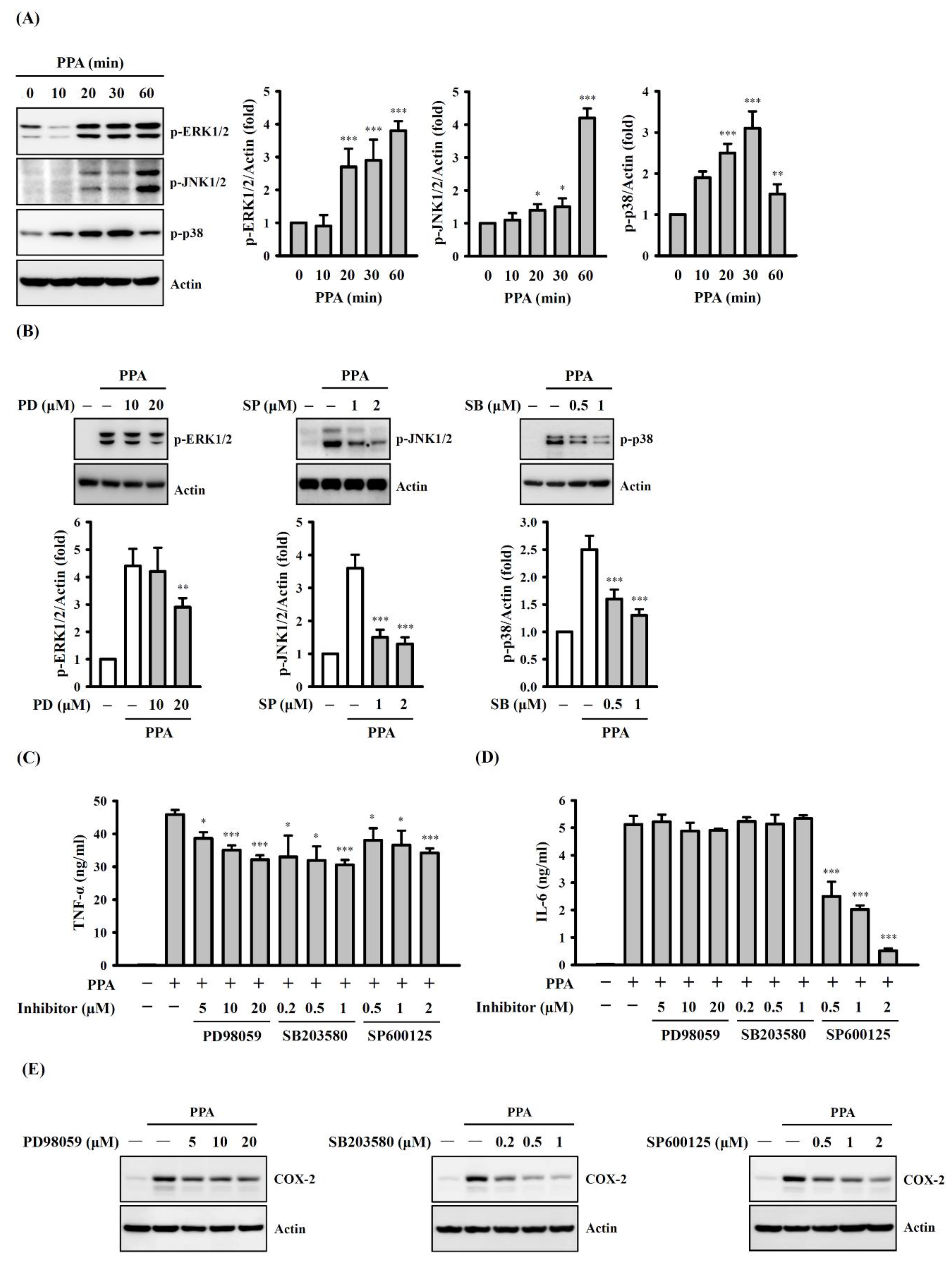
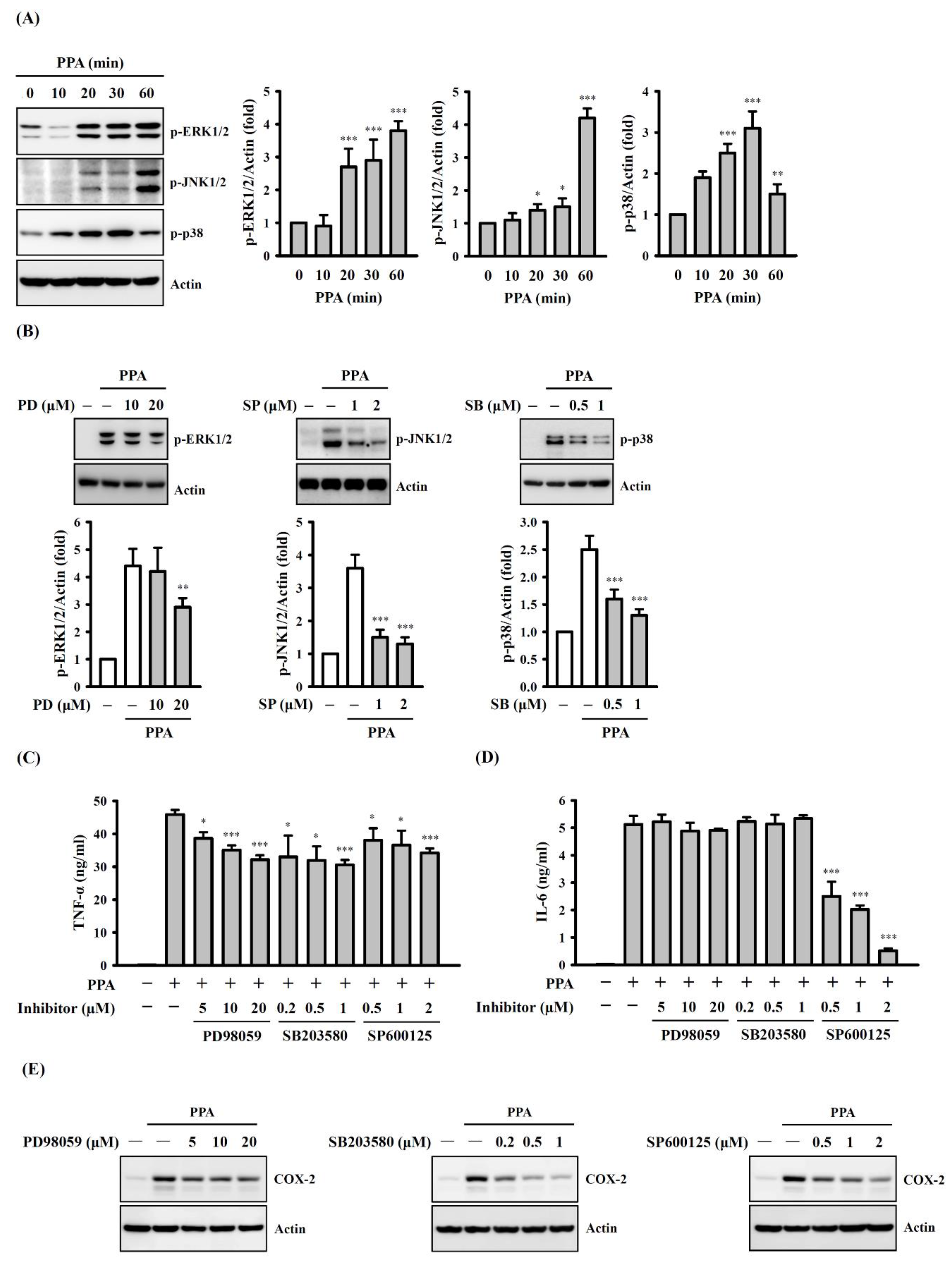
3.4. The Role of Mitogen-Activated Protein Kinases (MAPK) and NF-Κb on TNF-A, IL-6 and COX-2 Expression in PPA-Activated Macrophages
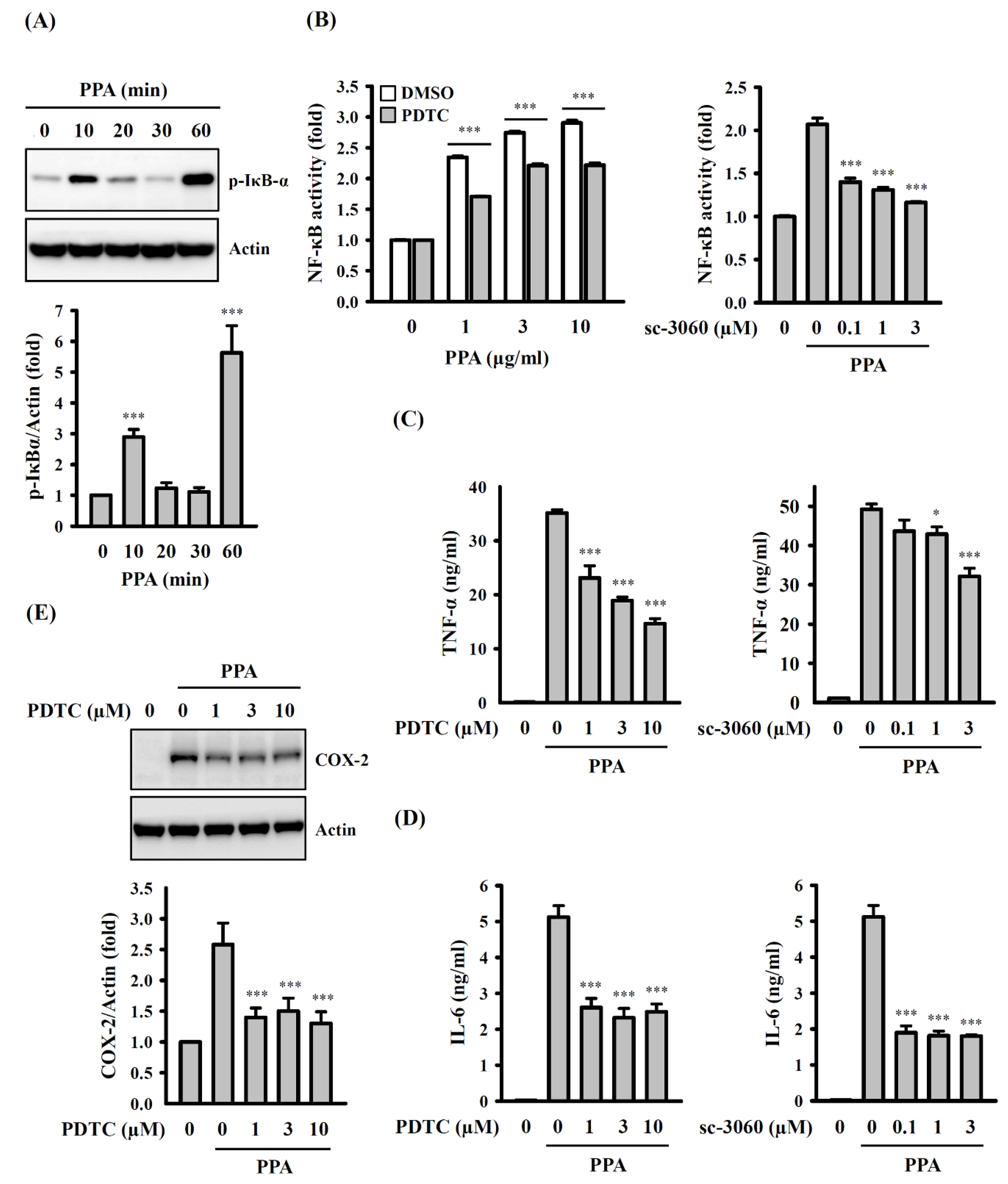
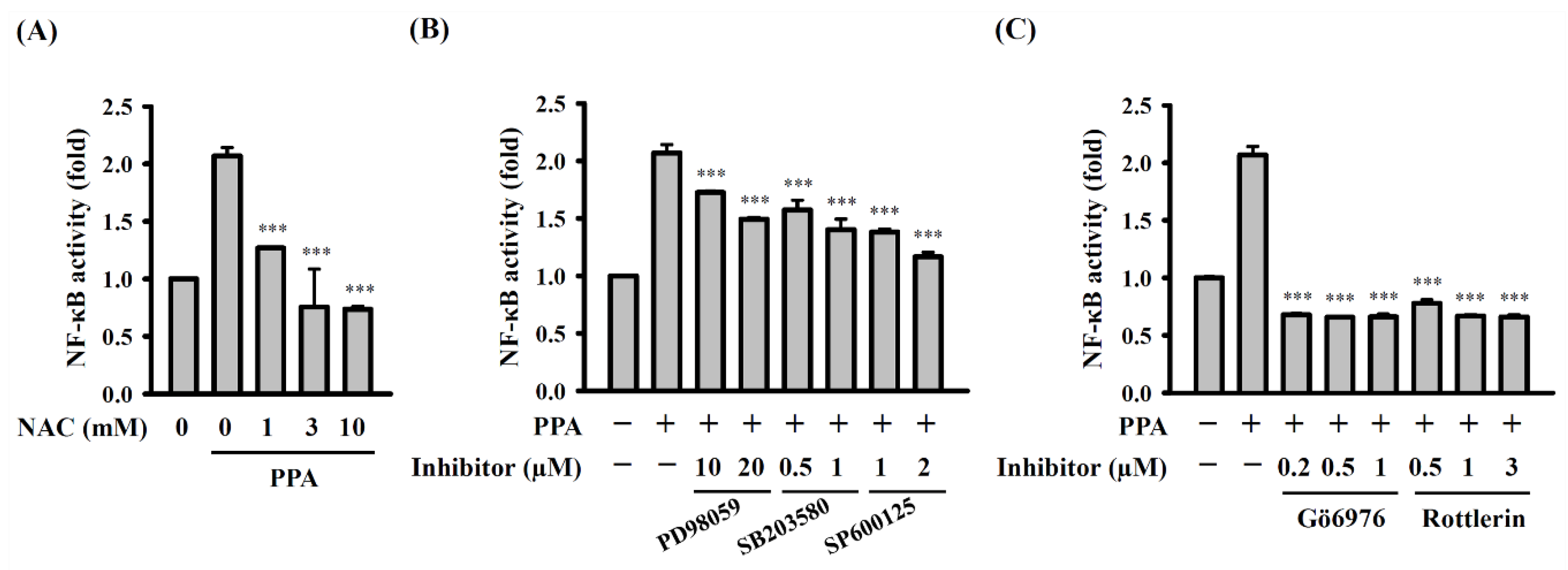
3.5. ROS, MAPK, and PKC Function Upstream of NF-κB in PPA-Activated Macrophages
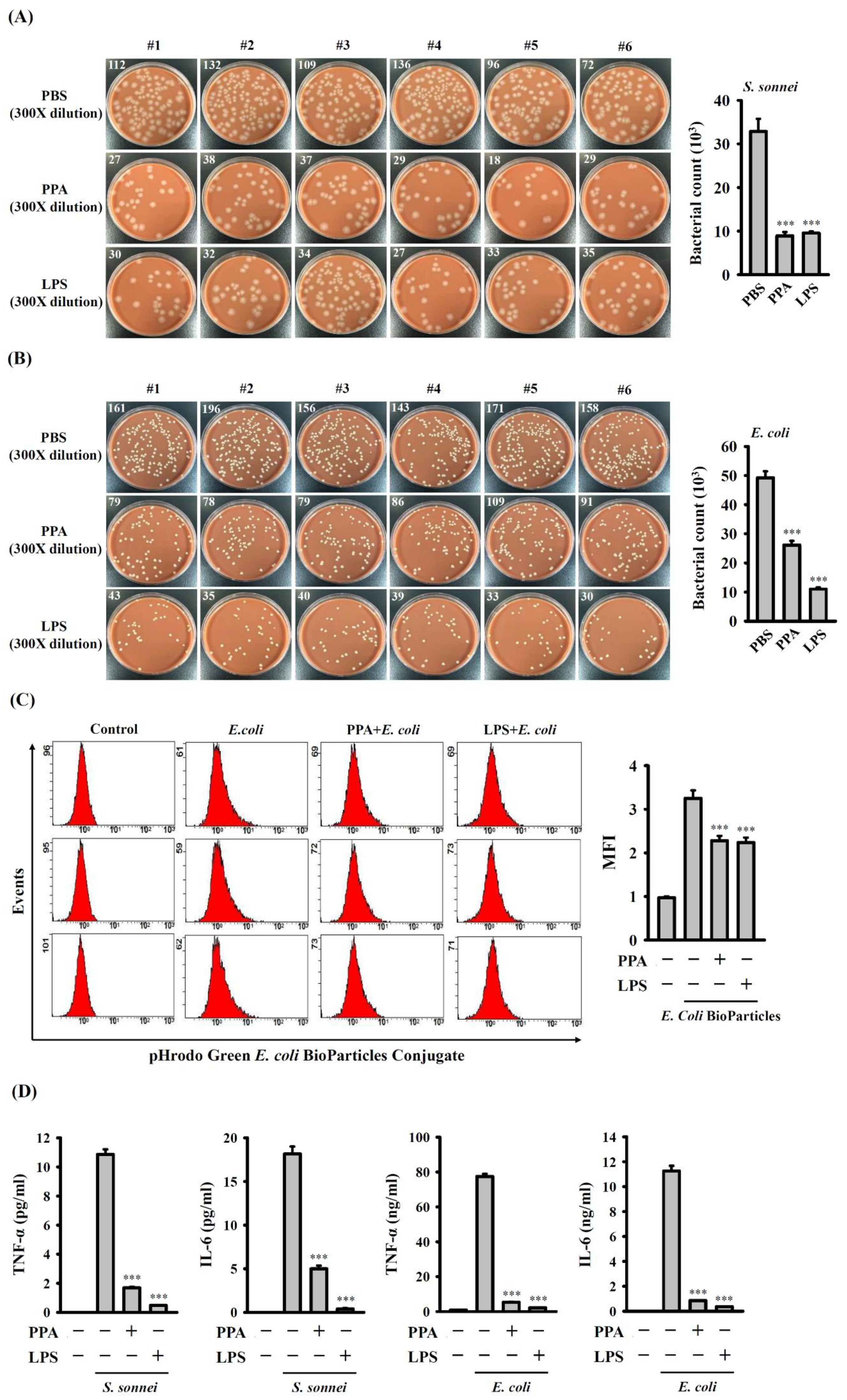
3.6. PPA Reduces the Phagocytic Activity and Cytokine Production of Macrophages in Response to Bacterial Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ameen, F.; AlNadhari, S.; Al-Homaidan, A.A. Marine microorganisms as an untapped source of bioactive compounds. Saudi J. Biol. Sci. 2021, 28, 224–231. [Google Scholar] [CrossRef]
- Xu, S.-Y.; Huang, X.; Cheong, K.-L. Recent Advances in Marine Algae Polysaccharides: Isolation, Structure, and Activities. Mar. Drugs 2017, 15, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besednova, N.N.; Zaporozhets, T.S.; Kuznetsova, T.A.; Makarenkova, I.D.; Kryzhanovsky, S.P.; Fedyanina, L.N.; Ermakova, S.P. Extracts and Marine Algae Polysaccharides in Therapy and Prevention of Inflammatory Diseases of the Intestine. Mar. Drugs 2020, 18, 289. [Google Scholar] [CrossRef] [PubMed]
- Riccio, G.; Lauritano, C. Microalgae with Immunomodulatory Activities. Mar. Drugs 2019, 18, 2. [Google Scholar] [CrossRef] [Green Version]
- Hans, N.; Malik, A.; Naik, S. Antiviral activity of sulfated polysaccharides from marine algae and its application in combating COVID-19: Mini review. Bioresour. Technol. Rep. 2021, 13, 100623. [Google Scholar] [CrossRef]
- Chen, X.; Han, W.; Wang, G.; Zhao, X. Application prospect of polysaccharides in the development of anti-novel coronavirus drugs and vaccines. Int. J. Biol. Macromol. 2020, 164, 331–343. [Google Scholar] [CrossRef]
- Song, S.; Peng, H.; Wang, Q.; Liu, Z.; Dong, X.; Wen, C.; Ai, C.; Zhang, Y.; Wang, Z.; Zhu, B. Inhibitory activities of marine sulfated polysaccharides against SARS-CoV-2. Food Funct. 2020, 11, 7415–7420. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Palanisamy, S.; Talapphet, N.; Cho, M.; You, S. Preparation and characterization of folic acid conjugated sulfated polysaccharides on NK cell activation and cellular uptake in HeLa cells. Carbohydr. Polym. 2021, 254, 117250. [Google Scholar] [CrossRef]
- Shi, F.; Liu, Z.; Liu, Y.; Cheong, K.-L.; Teng, B.; Khan, B.M. Comparison of Physicochemical Characteristics and Macrophage Immunostimulatory Activities of Polysaccharides from Chlamys farreri. Mar. Drugs 2020, 18, 429. [Google Scholar] [CrossRef]
- Yang, D.; Lin, F.; Huang, Y.; Ye, J.; Xiao, M. Separation, purification, structural analysis and immune-enhancing activity of sulfated polysaccharide isolated from sea cucumber viscera. Int. J. Biol. Macromol. 2020, 155, 1003–1018. [Google Scholar] [CrossRef]
- Tabarsa, M.; You, S.; Yelithao, K.; Palanisamy, S.; Prabhu, N.M.; Nan, M. Isolation, structural elucidation and immuno-stimulatory properties of polysaccharides from Cuminum cyminum. Carbohydr. Polym. 2020, 230, 115636. [Google Scholar] [CrossRef]
- Hong, W.; Wei, B.; Wang, S.; Ke, S.; Chen, J.; Zhang, H.; Wang, H. The Antioxidant Activity of Polysaccharides Derived from Marine Organisms: An Overview. Mar. Drugs 2019, 17, 674. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, G.; Liu, R.; Wei, M.; Zhang, J.; Sun, C. EPS364, a Novel Deep-Sea Bacterial Exopolysaccharide, Inhibits Liver Cancer Cell Growth and Adhesion. Mar. Drugs 2021, 19, 171. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.-X.; Chen, X.-Q.; Cheong, K.-L. Current trends in marine algae polysaccharides: The digestive tract, microbial catabolism, and prebiotic potential. Int. J. Biol. Macromol. 2020, 151, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Zhang, L.; Yan, Z.; Shen, J.; Chang, Y.; Wang, J. ι-Carrageenan Tetrasaccharide from ι-Carrageenan Inhibits Islet β Cell Apoptosis Via the Upregulation of GLP-1 to Inhibit the Mitochondrial Apoptosis Pathway. J. Agric. Food Chem. 2021, 69, 212–222. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Li, S.; Chen, Z.; Tan, J.; Yao, J.; Duan, D. Low molecular weight fucoidan alleviates diabetic nephropathy by binding fibronectin and inhibiting ECM-receptor interaction in human renal mesangial cells. Int. J. Biol. Macromol. 2020, 150, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Zhao, J.; Xing, M.; Xiao, H.; Zhang, Q.; Liang, H.; Ji, A.; Song, S. Current Research Landscape of Marine-Derived Anti-Atherosclerotic Substances. Mar. Drugs 2020, 18, 440. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Sulfated polysaccharides of some seaweeds exhibit neuroprotection via mitigation of oxidative stress, cholinergic dysfunction and inhibition of Zn—Induced neuronal damage in HT-22 cells. BMC Complement. Med. Ther. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Wright, R.M.; Strader, M.E.; Genuise, H.M.; Matz, M. Effects of thermal stress on amount, composition, and antibacterial properties of coral mucus. PeerJ 2019, 7, e6849. [Google Scholar] [CrossRef] [Green Version]
- Meikle, P.; Richards, G.N.; Yellowlees, D. Structural determination of the oligosaccharide side chains from a glycoprotein isolated from the mucus of the coral Acropora formosa. J. Biol. Chem. 1987, 262, 16941–16947. [Google Scholar] [CrossRef]
- Krediet, C.J.; Ritchie, K.B.; Cohen, M.; Lipp, E.; Sutherland, K.P.; Teplitski, M. Utilization of Mucus from the Coral Acropora palmata by the Pathogen Serratia marcescens and by Environmental and Coral Commensal Bacteria. Appl. Environ. Microbiol. 2009, 75, 3851–3858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molchanova, V.I.; Ovodova, R.G.; Perera, I.; Ovodov, Y.S. Carbohydrate-protein biopolymers from Pseudopterogorgia americana. Chem. Nat. Compd. 1982, 18, 639–642. [Google Scholar] [CrossRef]
- Molchanova, V.I.; Ovodova, R.G.; Ovodov, Y.S.; Elkin, Y.N.; Santana, V.F. Studies of the polysaccharide moiety of corallan, a glycoprotein from Pseudopterogorgia americana. Carbohydr. Res. 1985, 141, 289–293. [Google Scholar] [CrossRef]
- Bergman, P.; Raqib, R.; Rekha, R.S.; Agerberth, B.; Gudmundsson, G.H. Host Directed Therapy Against Infection by Boosting Innate Immunity. Front. Immunol. 2020, 11, 1209. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nat. Cell Biol. 2017, 542, 177–185. [Google Scholar] [CrossRef]
- Mangan, M.S.J.; Olhava, E.J.; Roush, W.R.; Seidel, H.M.; Glick, G.D.; Latz, E. Targeting the NLRP3 inflammasome in inflammatory diseases. Nat. Rev. Drug Discov. 2018, 17, 588–606. [Google Scholar] [CrossRef] [PubMed]
- Cui, J. Natural COX-2 Inhibitors as Promising Anti-inflammatory Agents: An Update. Curr. Med. Chem. 2021, 28, 3622–3646. [Google Scholar] [CrossRef]
- Hua, K.-F.; Yang, F.-L.; Chiu, H.-W.; Chou, J.-C.; Dong, W.-C.; Lin, C.-N.; Lin, C.-Y.; Wang, J.-T.; Li, L.-H.; Chiu, H.-W.; et al. Capsular Polysaccharide Is Involved in NLRP3 Inflammasome Activation by Klebsiella pneumoniae Serotype K1. Infect. Immun. 2015, 83, 3396–3409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loegering, D.J.; Lennartz, M.R. Protein Kinase C and Toll-Like Receptor Signaling. Enzym. Res. 2011, 2011, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Arthur, J.S.C.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef]
- Su, S.-C.; Hua, K.-F.; Lee, H.; Chao, L.K.; Tan, S.-K.; Yang, S.-F.; Hsu, H.-Y. LTA and LPS mediated activation of protein kinases in the regulation of inflammatory cytokines expression in macrophages. Clin. Chim. Acta 2006, 374, 106–115. [Google Scholar] [CrossRef]
- Chao, L.K.; Hua, K.-F.; Hsu, H.-Y.; Cheng, S.-S.; Lin, I.-F.; Chen, C.-J.; Chen, S.-T.; Chang, S.-T. Cinnamaldehyde inhibits pro-inflammatory cytokines secretion from monocytes/macrophages through suppression of intracellular signaling. Food Chem. Toxicol. 2008, 46, 220–231. [Google Scholar] [CrossRef]
- Lu, Y.-C.; Yeh, W.-C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Lye, E.; Mirtsos, C.; Suzuki, N.; Suzuki, S.; Yeh, W.-C. The Role of Interleukin 1 Receptor-associated Kinase-4 (IRAK-4) Kinase Activity in IRAK-4-mediated Signaling. J. Biol. Chem. 2004, 279, 40653–40658. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Sanjo, H.; Takeda, K.; Ninomiya-Tsuji, J.; Yamamoto, M.; Kawai, T.; Matsumoto, K.; Takeuchi, O.; Akira, S. Essential function for the kinase TAK1 in innate and adaptive immune responses. Nat. Immunol. 2005, 6, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Kaisho, T.; Sanjo, H.; Takeuchi, O.; Sugiyama, M.; Okabe, M.; Takeda, K.; et al. Role of Adaptor TRIF in the MyD88-Independent Toll-Like Receptor Signaling Pathway. Science 2003, 301, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Meylan, E.; Burns, K.; Hofmann, K.; Blancheteau, V.; Martinon, F.; A Kelliher, M.; Tschopp, J. RIP1 is an essential mediator of Toll-like receptor 3–induced NF-κB activation. Nat. Immunol. 2004, 5, 503–507. [Google Scholar] [CrossRef]
- Liao, P.-C.; Chao, L.K.; Chou, J.-C.; Dong, W.-C.; Lin, C.-N.; Lin, C.-Y.; Chen, A.; Ka, S.-M.; Ho, C.-L.; Hua, K.-F. Lipopolysaccharide/adenosine triphosphate-mediated signal transduction in the regulation of NLRP3 protein expression and caspase-1-mediated interleukin-1β secretion. Inflamm. Res. 2013, 62, 89–96. [Google Scholar] [CrossRef]
- Fujihara, M.; Wakamoto, S.; Ito, T.; Muroi, M.; Suzuki, T.; Ikeda, H.; Ikebuchi, K. Lipopolysaccharide-triggered desensitization of TNF-alpha mRNA expression involves lack of phosphorylation of IkappaBalpha in a murine macrophage-like cell line, P388D1. J. Leukoc. Biol. 2000, 68, 267–276. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Wen, M.-H. Lipopolysaccharide-mediated Reactive Oxygen Species and Signal Transduction in the Regulation of Interleukin-1 Gene Expression. J. Biol. Chem. 2002, 277, 22131–22139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.-L.; Yang, Y.-L.; Liao, P.-C.; Chou, J.-C.; Tsai, K.-C.; Yang, A.-S.; Sheu, F.; Lin, T.-L.; Hsieh, P.-F.; Wang, J.-T.; et al. Structure and Immunological Characterization of the Capsular Polysaccharide of a Pyrogenic Liver Abscess Caused by Klebsiella pneumoniae. J. Biol. Chem 2011, 286, 21041–21051. [Google Scholar] [CrossRef] [Green Version]
- Dupré, S.; Erard, M.; Nüβe, O. ROS production in phagocytes: Why, when, and where? J. Leukoc. Biol. 2013, 94, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Perera, N.; Yang, F.-L.; Chang, C.-M.; Lu, Y.-T.; Zhan, S.-H.; Tsai, Y.-T.; Hsieh, J.-F.; Li, L.-H.; Hua, K.-F.; Wu, S.-H. Galactomannan from Antrodia cinnamomea Enhances the Phagocytic Activity of Macrophages. Org. Lett. 2017, 19, 3486–3489. [Google Scholar] [CrossRef]
- Perera, N.; Yang, F.-L.; Chern, J.; Chiu, H.-W.; Hsieh, C.-Y.; Li, L.-H.; Zhang, Y.-L.; Hua, K.-F.; Wu, S.-H. Carboxylic and O-acetyl moieties are essential for the immunostimulatory activity of glucuronoxylomannan: A novel TLR4 specific immunostimulator from Auricularia auricula-judae. Chem. Commun. 2018, 54, 6995–6998. [Google Scholar] [CrossRef]
- Perera, N.; Yang, F.-L.; Lu, Y.-T.; Li, L.-H.; Hua, K.-F.; Wu, S.-H. Antrodia cinnamomea Galactomannan Elicits Immuno-stimulatory Activity Through Toll-like Receptor 4. Int. J. Biol. Sci. 2018, 14, 1378–1388. [Google Scholar] [CrossRef] [Green Version]
- Hua, K.-F.; Hsu, H.-Y.; Chao, L.K.; Chen, S.-T.; Yang, W.-B.; Hsu, J.; Wong, C.-H. Ganoderma lucidum polysaccharides enhance CD14 endocytosis of LPS and promote TLR4 signal transduction of cytokine expression. J. Cell. Physiol. 2007, 212, 537–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uribe-Querol, E.; Rosales, C. Control of Phagocytosis by Microbial Pathogens. Front. Immunol. 2017, 8, 1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Li, Y.; Pei, G. Polysaccharides from Ganoderma lucidum attenuate microglia-mediated neuroinflammation and modulate microglial phagocytosis and behavioural response. J. Neuroinflamm. 2017, 14, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hult, A.; Toss, F.; Malm, C.; Oldenborg, P. In vitro phagocytosis of liquid-stored red blood cells requires serum and can be inhibited with fucoidan and dextran sulphate. Vox Sang. 2020, 115, 647–654. [Google Scholar] [CrossRef]
- de Lima, T.M.; Sampaio, S.C.; Petroni, R.; Brigatte, P.; Velasco, I.T.; Soriano, F.G. Phagocytic activity of LPS tolerant macrophages. Mol. Immunol. 2014, 60, 8–13. [Google Scholar] [CrossRef]
- Pena, O.M.; Pistolic, J.; Raj, D.; Fjell, C.D.; Hancock, R.E.W. Endotoxin tolerance represents a distinctive state of alternative polarization (M2) in human mononuclear cells. J. Immunol. 2011, 186, 7243–7254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
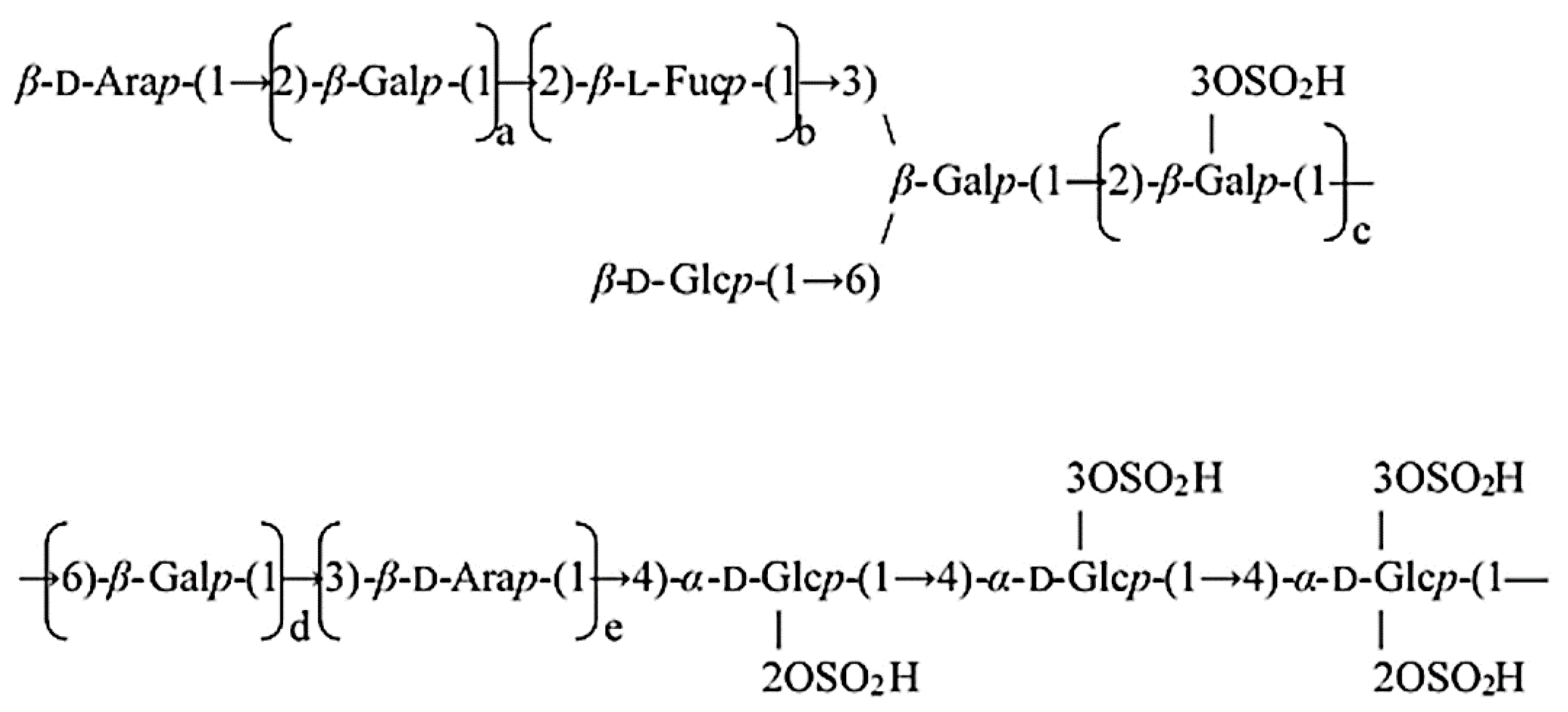

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernikov, O.V.; Chiu, H.-W.; Li, L.-H.; Kokoulin, M.S.; Molchanova, V.I.; Hsu, H.-T.; Ho, C.-L.; Hua, K.-F. Immunomodulatory Properties of Polysaccharides from the Coral Pseudopterogorgia americana in Macrophages. Cells 2021, 10, 3531. https://doi.org/10.3390/cells10123531
Chernikov OV, Chiu H-W, Li L-H, Kokoulin MS, Molchanova VI, Hsu H-T, Ho C-L, Hua K-F. Immunomodulatory Properties of Polysaccharides from the Coral Pseudopterogorgia americana in Macrophages. Cells. 2021; 10(12):3531. https://doi.org/10.3390/cells10123531
Chicago/Turabian StyleChernikov, Oleg V., Hsiao-Wen Chiu, Lan-Hui Li, Maxim S. Kokoulin, Valentina I. Molchanova, Hsien-Ta Hsu, Chen-Lung Ho, and Kuo-Feng Hua. 2021. "Immunomodulatory Properties of Polysaccharides from the Coral Pseudopterogorgia americana in Macrophages" Cells 10, no. 12: 3531. https://doi.org/10.3390/cells10123531
APA StyleChernikov, O. V., Chiu, H.-W., Li, L.-H., Kokoulin, M. S., Molchanova, V. I., Hsu, H.-T., Ho, C.-L., & Hua, K.-F. (2021). Immunomodulatory Properties of Polysaccharides from the Coral Pseudopterogorgia americana in Macrophages. Cells, 10(12), 3531. https://doi.org/10.3390/cells10123531