
Extracellular Signalling Modulates Scar/WAVE Complex Activity through Abi Phosphorylation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Dictyostelium Cell Culture
2.2. Plasmid Constructs
2.3. Generation of Scar-/Abi- Double Knock out Cells
2.4. Generation of Scar-/Abi-/pir121-eGFP Cells
2.5. Transfection of Dictyostelium Cells
2.6. GFP-TRAP Pulldown
2.7. Western Blotting
2.8. Phosphatase Treatment
2.9. Under Agarose Chemotaxis
2.10. Stimulation of Cells with Folate and cAMP
2.11. Microscopy
2.12. Quantification of Data and Statistics
3. Results
3.1. Constitutive Phosphorylation of Abi
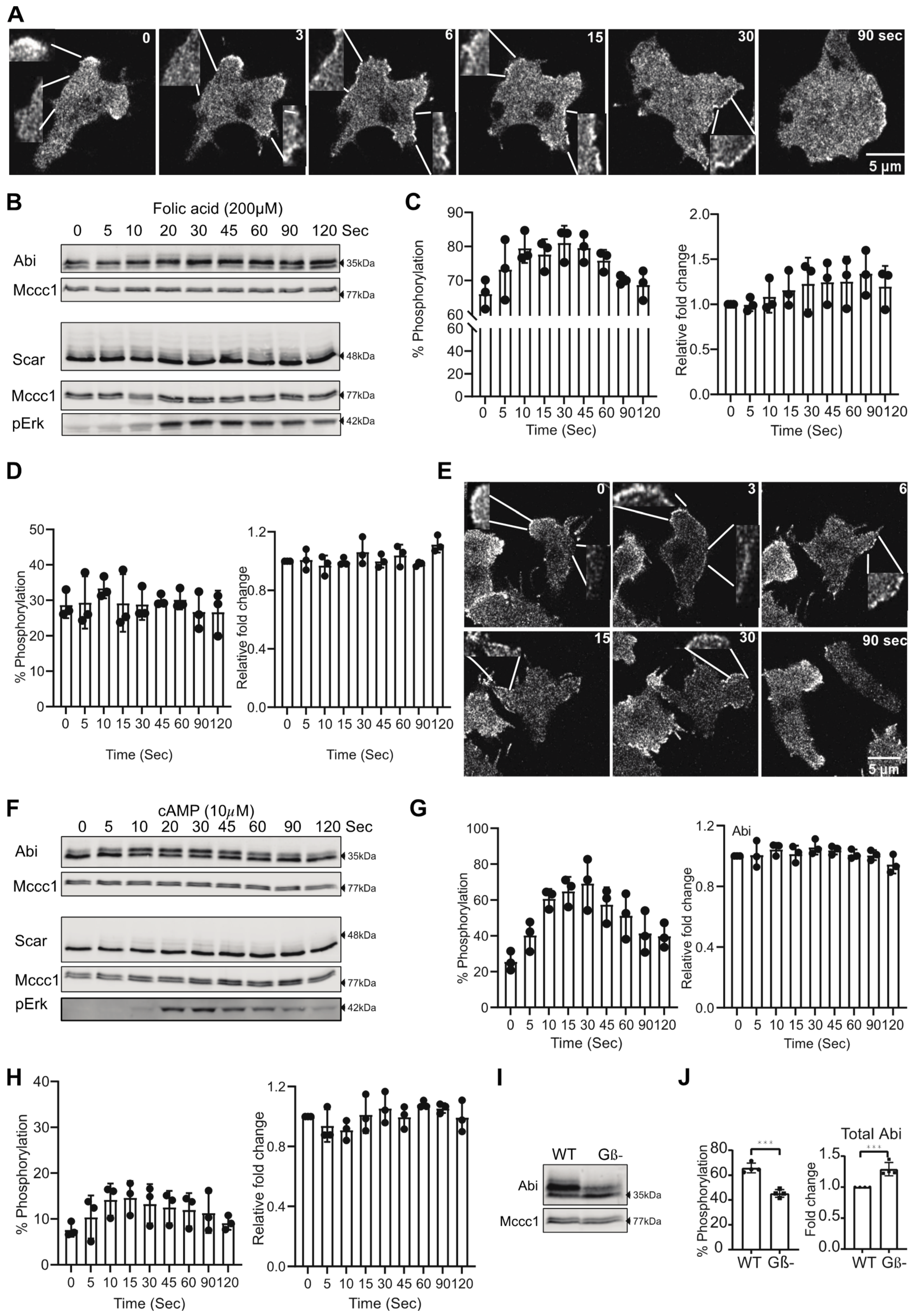
3.2. Chemoattractant Stimulation of Abi Phosphorylation
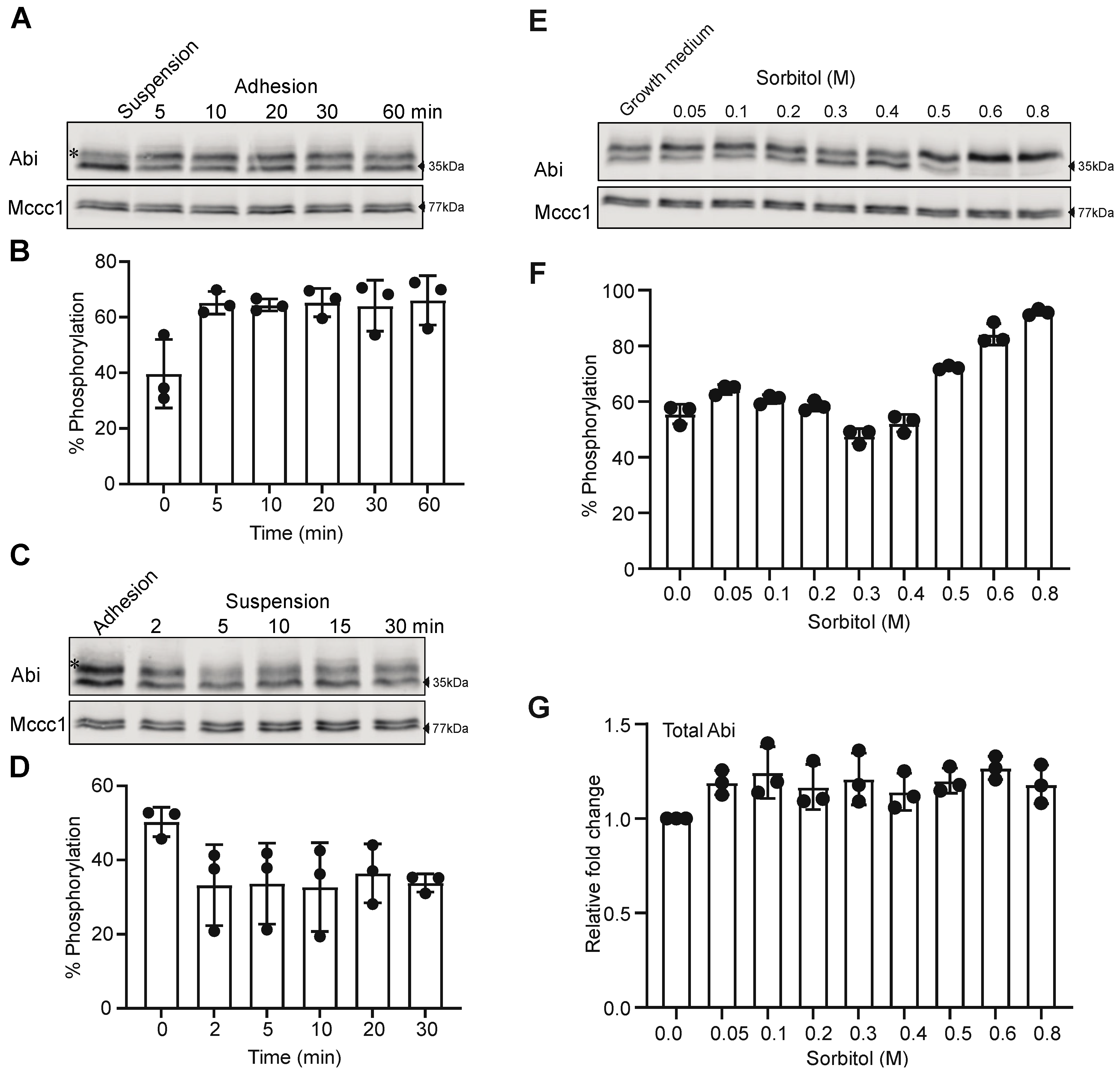
3.3. Cell-Substrate Adhesion and Osmotic Shock Alter Abi Phosphorylation
3.4. Abi Phosphorylation Is Activation-Dependent
3.5. Abi Phosphorylation Tunes Cell Migration and Pseudopod Formation
3.6. Double Abi/Scar Phospho-Mutants Still Rescue Migration in Scar-/Abi-Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krause, M.; Gautreau, A. Steering cell migration: Lamellipodium dynamics and the regulation of directional persistence. Nat. Rev. Mol. Cell Biol. 2014, 15, 577–590. [Google Scholar] [CrossRef]
- Insall, R.H.; Machesky, L.M. Actin dynamics at the leading edge: From simple machinery to complex networks. Dev. Cell 2009, 17, 310–322. [Google Scholar] [CrossRef]
- Chen, B.; Chou, H.T.; Brautigam, C.A.; Xing, W.; Yang, S.; Henry, L.; Doolittle, L.K.; Walz, T.; Rosen, M.K. Rac1 GTPase activates the WAVE regulatory complex through two distinct binding sites. Elife 2017, 6, e29795. [Google Scholar] [CrossRef] [PubMed]
- Schaks, M.; Singh, S.P.; Kage, F.; Thomason, P.; Klünemann, T.; Steffen, A.; Blankenfeldt, W.; Stradal, T.E.; Insall, R.H.; Rottner, K. Distinct Interaction Sites of Rac GTPase with WAVE Regulatory Complex Have Non-redundant Functions In Vivo. Curr. Biol. 2018, 28, 3674–3684.e3676. [Google Scholar] [CrossRef] [PubMed]
- Danson, C.M.; Pocha, S.M.; Bloomberg, G.B.; Cory, G.O. Phosphorylation of WAVE2 by MAP kinases regulates persistent cell migration and polarity. J. Cell Sci. 2007, 120, 4144–4154. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, M.C. Phosphoregulation of the WAVE regulatory complex and signal integration. Semin. Cell Dev. Biol. 2013, 24, 272–279. [Google Scholar] [CrossRef]
- Mendoza, M.C.; Er, E.E.; Zhang, W.; Ballif, B.A.; Elliott, H.L.; Danuser, G.; Blenis, J. ERK-MAPK drives lamellipodia protrusion by activating the WAVE2 regulatory complex. Mol. Cell 2011, 41, 661–671. [Google Scholar] [CrossRef]
- Nakanishi, O.; Suetsugu, S.; Yamazaki, D.; Takenawa, T. Effect of WAVE2 phosphorylation on activation of the Arp2/3 complex. J. Biochem. 2007, 141, 319–325. [Google Scholar] [CrossRef]
- Leng, Y.; Zhang, J.; Badour, K.; Arpaia, E.; Freeman, S.; Cheung, P.; Siu, M.; Siminovitch, K. Abelson-interactor-1 promotes WAVE2 membrane translocation and Abelson-mediated tyrosine phosphorylation required for WAVE2 activation. Proc. Natl. Acad. Sci. USA 2005, 102, 1098–1103. [Google Scholar] [CrossRef]
- Stuart, J.R.; Gonzalez, F.H.; Kawai, H.; Yuan, Z.M. c-Abl interacts with the WAVE2 signaling complex to induce membrane ruffling and cell spreading. J. Biol. Chem. 2006, 281, 31290–31297. [Google Scholar] [CrossRef]
- Li, Y.; Clough, N.; Sun, X.; Yu, W.; Abbott, B.L.; Hogan, C.J.; Dai, Z. Bcr-Abl induces abnormal cytoskeleton remodeling, beta1 integrin clustering and increased cell adhesion to fibronectin through the Abl interactor 1 pathway. J. Cell Sci. 2007, 120, 1436–1446. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, C.H.; Lin, T.Y.; Pan, R.L.; Juang, J.L. The involvement of Abl and PTP61F in the regulation of Abi protein localization and stability and lamella formation in Drosophila S2 cells. J. Biol. Chem. 2007, 282, 32442–32452. [Google Scholar] [CrossRef]
- Singh, S.P.; Thomason, P.A.; Lilla, S.; Schaks, M.; Tang, Q.; Goode, B.L.; Machesky, L.M.; Rottner, K.; Insall, R.H. Cell-substrate adhesion drives Scar/WAVE activation and phosphorylation by a Ste20-family kinase, which controls pseudopod lifetime. PLoS Biol. 2020, 18, e3000774. [Google Scholar] [CrossRef] [PubMed]
- Lebensohn, A.M.; Kirschner, M.W. Activation of the WAVE complex by coincident signals controls actin assembly. Mol. Cell 2009, 36, 512–524. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Tang, H.; Dissanaike, S.; Cobos, E.; Tao, Y.; Dai, Z. CDK1-mediated phosphorylation of Abi1 attenuates Bcr-Abl-induced F-actin assembly and tyrosine phosphorylation of WAVE complex during mitosis. J. Biol. Chem. 2011, 286, 38614–38626. [Google Scholar] [CrossRef]
- Dai, Z.; Kerzic, P.; Schroeder, W.G.; McNiece, I.K. Deletion of the Src homology 3 domain and C-terminal proline-rich sequences in Bcr-Abl prevents Abl interactor 2 degradation and spontaneous cell migration and impairs leukemogenesis. J. Biol. Chem. 2001, 276, 28954–28960. [Google Scholar] [CrossRef]
- Goldberg, J.M.; Manning, G.; Liu, A.; Fey, P.; Pilcher, K.E.; Xu, Y.; Smith, J.L. The dictyostelium kinome--analysis of the protein kinases from a simple model organism. PLoS Genet. 2006, 2, e38. [Google Scholar] [CrossRef]
- Insall, R.H.; Borleis, J.; Devreotes, P.N. The aimless RasGEF is required for processing of chemotactic signals through G-protein-coupled receptors in Dictyostelium. Curr. Biol. 1996, 6, 719–729. [Google Scholar] [CrossRef]
- Wilkins, A.; Khosla, M.; Fraser, D.J.; Spiegelman, G.B.; Fisher, P.R.; Weeks, G.; Insall, R.H. Dictyostelium RasD is required for normal phototaxis, but not differentiation. Genes Dev. 2000, 14, 1407–1413. [Google Scholar] [CrossRef]
- Fey, P.; Dodson, R.J.; Basu, S.; Chisholm, R.L. One stop shop for everything Dictyostelium: DictyBase and the Dicty Stock Center in 2012. Methods Mol. Biol. 2013, 983, 59–92. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.J.; Ura, S.; Thomason, P.A.; Kalna, G.; Insall, R.H. Abi is required for modulation and stability but not localization or activation of the SCAR/WAVE complex. Eukaryot. Cell 2013, 12, 1509–1516. [Google Scholar] [CrossRef][Green Version]
- Pollitt, A.Y.; Insall, R.H. Abi mutants in Dictyostelium reveal specific roles for the SCAR/WAVE complex in cytokinesis. Curr. Biol. 2008, 18, 203–210. [Google Scholar] [CrossRef]
- Davidson, A.J.; King, J.S.; Insall, R.H. The use of streptavidin conjugates as immunoblot loading controls and mitochondrial markers for use with Dictyostelium discoideum. Biotechniques 2013, 55, 39–41. [Google Scholar] [CrossRef]
- Singh, S.P.; Insall, R.H. Adhesion stimulates Scar/WAVE phosphorylation in mammalian cells. Commun. Integr. Biol. 2020, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Tang, M.; Shi, C.; Iglesias, P.A.; Devreotes, P.N. An excitable signal integrator couples to an idling cytoskeletal oscillator to drive cell migration. Nat. Cell Biol. 2013, 15, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.T.; Janetopoulos, C.; Lee, S.; Charest, P.G.; Takeda, K.; Sundheimer, L.W.; Meili, R.; Devreotes, P.N.; Firtel, R.A. G protein-independent Ras/PI3K/F-actin circuit regulates basic cell motility. J. Cell Biol. 2007, 178, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Cain, R.J.; Ridley, A.J. Phosphoinositide 3-kinases in cell migration. Biol. Cell 2009, 101, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Ura, S.; Pollitt, A.Y.; Veltman, D.M.; Morrice, N.A.; Machesky, L.M.; Insall, R.H. Pseudopod growth and evolution during cell movement is controlled through SCAR/WAVE dephosphorylation. Curr. Biol. 2012, 22, 553–561. [Google Scholar] [CrossRef] [PubMed]
- McRobbie, S.J.; Newell, P.C. Chemoattractant-mediated changes in cytoskeletal actin of cellular slime moulds. J. Cell Sci. 1984, 68, 139–151. [Google Scholar] [CrossRef]
- Xiong, Y.; Huang, C.H.; Iglesias, P.A.; Devreotes, P.N. Cells navigate with a local-excitation, global-inhibition-biased excitable network. Proc. Natl. Acad. Sci. USA 2010, 107, 17079–17086. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Valkema, R.; Van Haastert, P.J.; Devreotes, P.N. The G protein beta subunit is essential for multiple responses to chemoattractants in Dictyostelium. J. Cell Biol. 1995, 129, 1667–1675. [Google Scholar] [CrossRef]
- Yamada, Y.; Sameshima, M. Cell shape regulation and co-translocation of actin and adenosyl homocysteinase in response to intermediate hypertonicity. FEMS Microbiol. Lett. 2004, 238, 417–422. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rivero, F.; Koppel, B.; Peracino, B.; Bozzaro, S.; Siegert, F.; Weijer, C.J.; Schleicher, M.; Albrecht, R.; Noegel, A.A. The role of the cortical cytoskeleton: F-actin crosslinking proteins protect against osmotic stress, ensure cell size, cell shape and motility, and contribute to phagocytosis and development. J. Cell Sci. 1996, 109 Pt 11, 2679–2691. [Google Scholar] [CrossRef]
- Zischka, H.; Oehme, F.; Pintsch, T.; Ott, A.; Keller, H.; Kellermann, J.; Schuster, S.C. Rearrangement of cortex proteins constitutes an osmoprotective mechanism in Dictyostelium. EMBO J. 1999, 18, 4241–4249. [Google Scholar] [CrossRef] [PubMed]
- Kuwayama, H.; Ecke, M.; Gerisch, G.; Van Haastert, P.J. Protection against osmotic stress by cGMP-mediated myosin phosphorylation. Science 1996, 271, 207–209. [Google Scholar] [CrossRef]
- Coue, M.; Brenner, S.L.; Spector, I.; Korn, E.D. Inhibition of actin polymerization by latrunculin A. FEBS Lett. 1987, 213, 316–318. [Google Scholar] [CrossRef]
- Gerisch, G.; Bretschneider, T.; Muller-Taubenberger, A.; Simmeth, E.; Ecke, M.; Diez, S.; Anderson, K. Mobile actin clusters and traveling waves in cells recovering from actin depolymerization. Biophys. J. 2004, 87, 3493–3503. [Google Scholar] [CrossRef] [PubMed]
- Millius, A.; Dandekar, S.N.; Houk, A.R.; Weiner, O.D. Neutrophils establish rapid and robust WAVE complex polarity in an actin-dependent fashion. Curr. Biol. 2009, 19, 253–259. [Google Scholar] [CrossRef]
- Eden, S.; Rohatgi, R.; Podtelejnikov, A.V.; Mann, M.; Kirschner, M.W. Mechanism of regulation of WAVE1-induced actin nucleation by Rac1 and Nck. Nature 2002, 418, 790–793. [Google Scholar] [CrossRef]
- Faix, J.; Weber, I. A dual role model for active Rac1 in cell migration. Small GTPases 2013, 4, 110–115. [Google Scholar] [CrossRef]
- Veltman, D.M.; King, J.S.; Machesky, L.M.; Insall, R.H. SCAR knockouts in Dictyostelium: WASP assumes SCAR’s position and upstream regulators in pseudopods. J. Cell Biol. 2012, 198, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Soede, R.D.; Schaap, P.; Valkema, R.; Borleis, J.A.; Van Haastert, P.J.; Devreotes, P.N.; Hereld, D. Phosphorylation of chemoattractant receptors is not essential for chemotaxis or termination of G-protein-mediated responses. J. Biol. Chem. 1997, 272, 27313–27318. [Google Scholar] [CrossRef] [PubMed]
- Insall, R. The interaction between pseudopods and extracellular signalling during chemotaxis and directed migration. Curr. Opin. Cell Biol. 2013, 25, 526–531. [Google Scholar] [CrossRef]
- Insall, R.; Andrew, N. Chemotaxis in Dictyostelium: How to walk straight using parallel pathways. Curr. Opin. Microbiol. 2007, 10, 578–581. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.P.; Thomason, P.A.; Insall, R.H. Extracellular Signalling Modulates Scar/WAVE Complex Activity through Abi Phosphorylation. Cells 2021, 10, 3485. https://doi.org/10.3390/cells10123485
Singh SP, Thomason PA, Insall RH. Extracellular Signalling Modulates Scar/WAVE Complex Activity through Abi Phosphorylation. Cells. 2021; 10(12):3485. https://doi.org/10.3390/cells10123485
Chicago/Turabian StyleSingh, Shashi Prakash, Peter A. Thomason, and Robert H. Insall. 2021. "Extracellular Signalling Modulates Scar/WAVE Complex Activity through Abi Phosphorylation" Cells 10, no. 12: 3485. https://doi.org/10.3390/cells10123485
APA StyleSingh, S. P., Thomason, P. A., & Insall, R. H. (2021). Extracellular Signalling Modulates Scar/WAVE Complex Activity through Abi Phosphorylation. Cells, 10(12), 3485. https://doi.org/10.3390/cells10123485