
Hyaluronan-Based Gel Promotes Human Dental Pulp Stem Cells Bone Differentiation by Activating YAP/TAZ Pathway

 ,
,  , ,
, ,  ,
,  ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Dental Pulp Extraction and Culture
2.3. hDPSCs Isolation and Osteogenic Differentiation
2.4. YAP/TAZ Inhibitor-1 Treatment and Cell Viability Assay
2.5. RNA Extraction and Quantitative Real-Time PCR Analyses
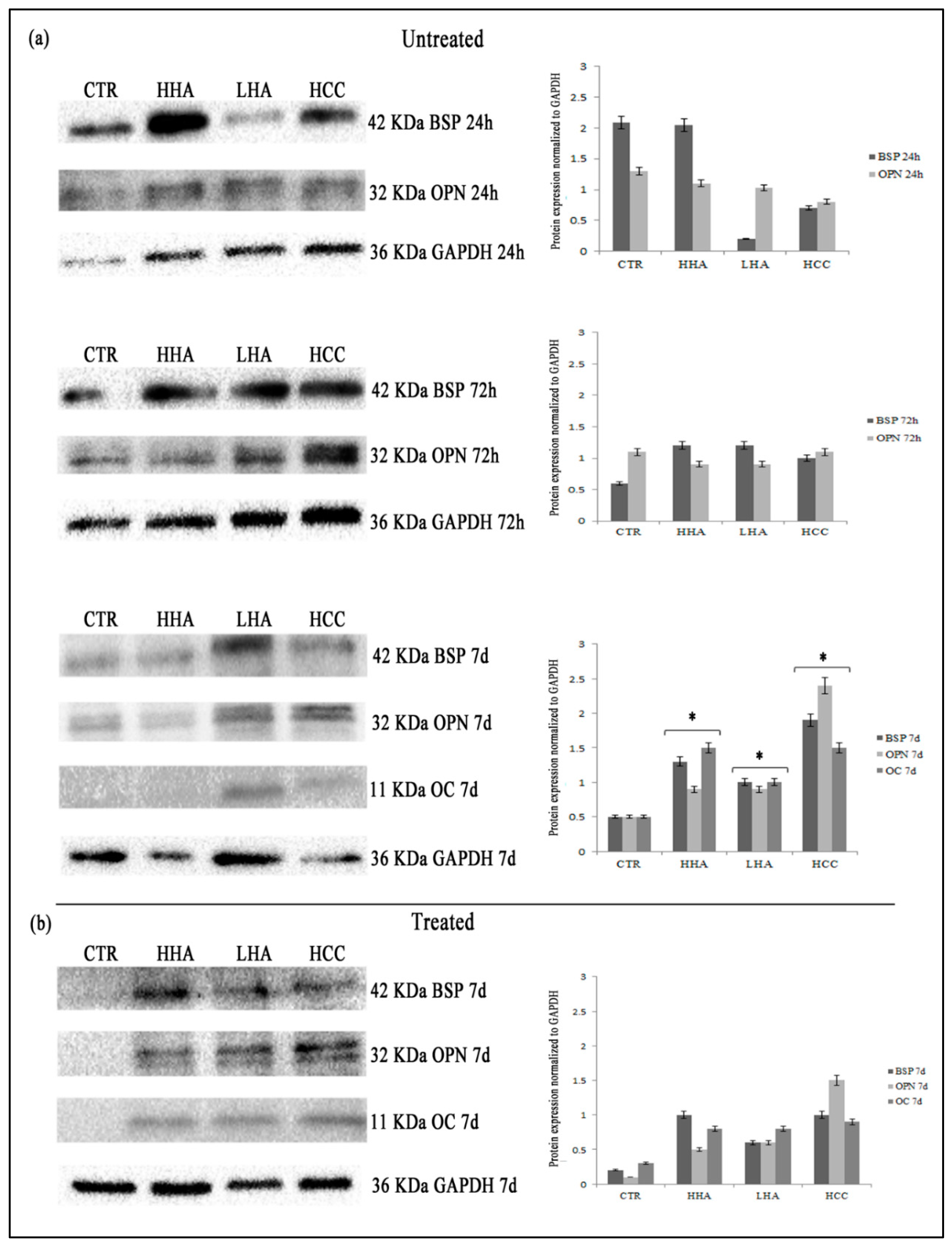
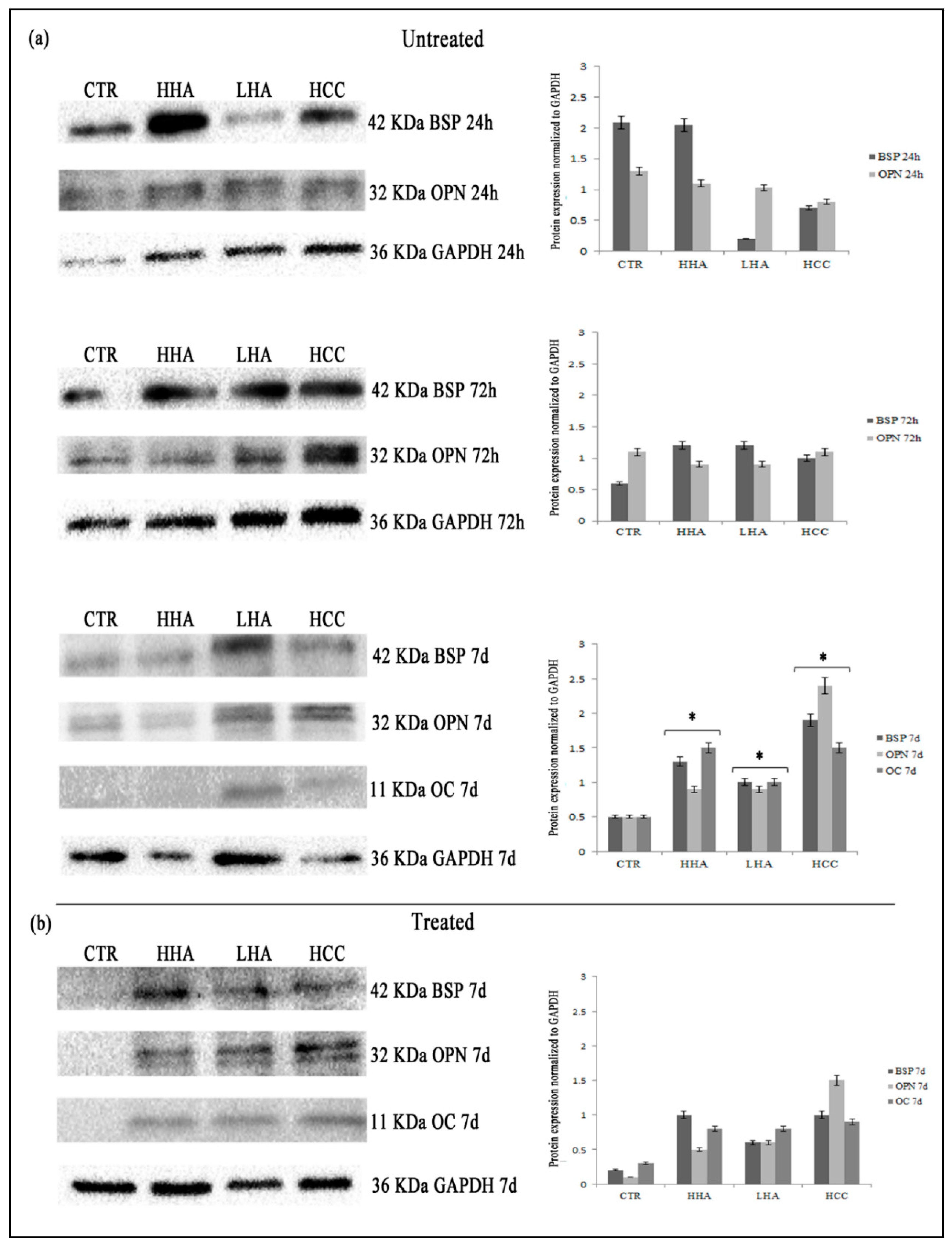
2.6. Protein Levels Evaluation of OC, OPN, and BSP by Western Blotting Analyses
2.7. CD44 Expression in hDPSCs by Immunofluorescence Staining
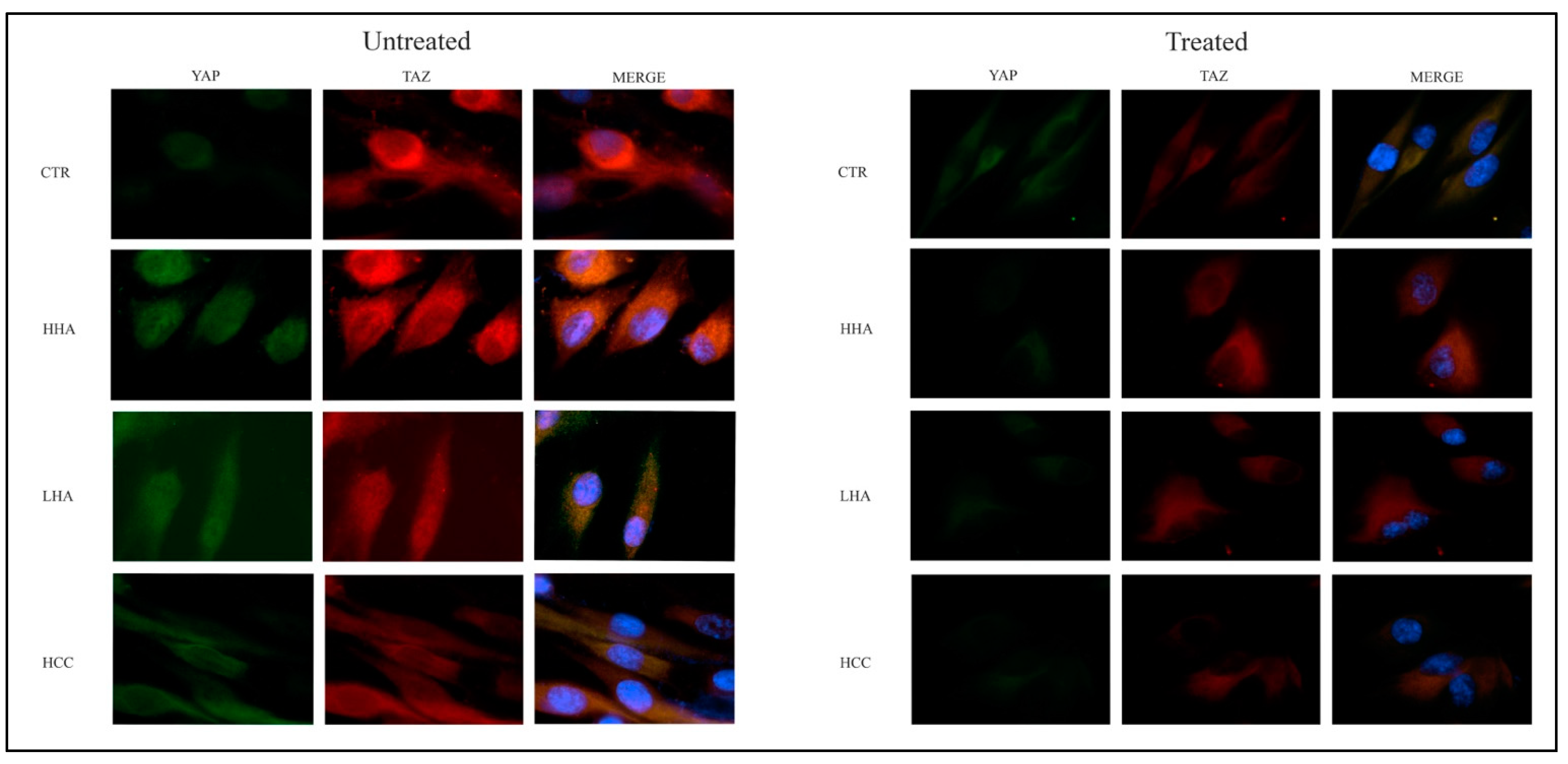
2.8. YAP-TAZ Expression in hDPSCs by Immunofluorescence Staining
2.9. Statistical Analysis
3. Results
3.1. hDPSCs Isolation and Cell Culture
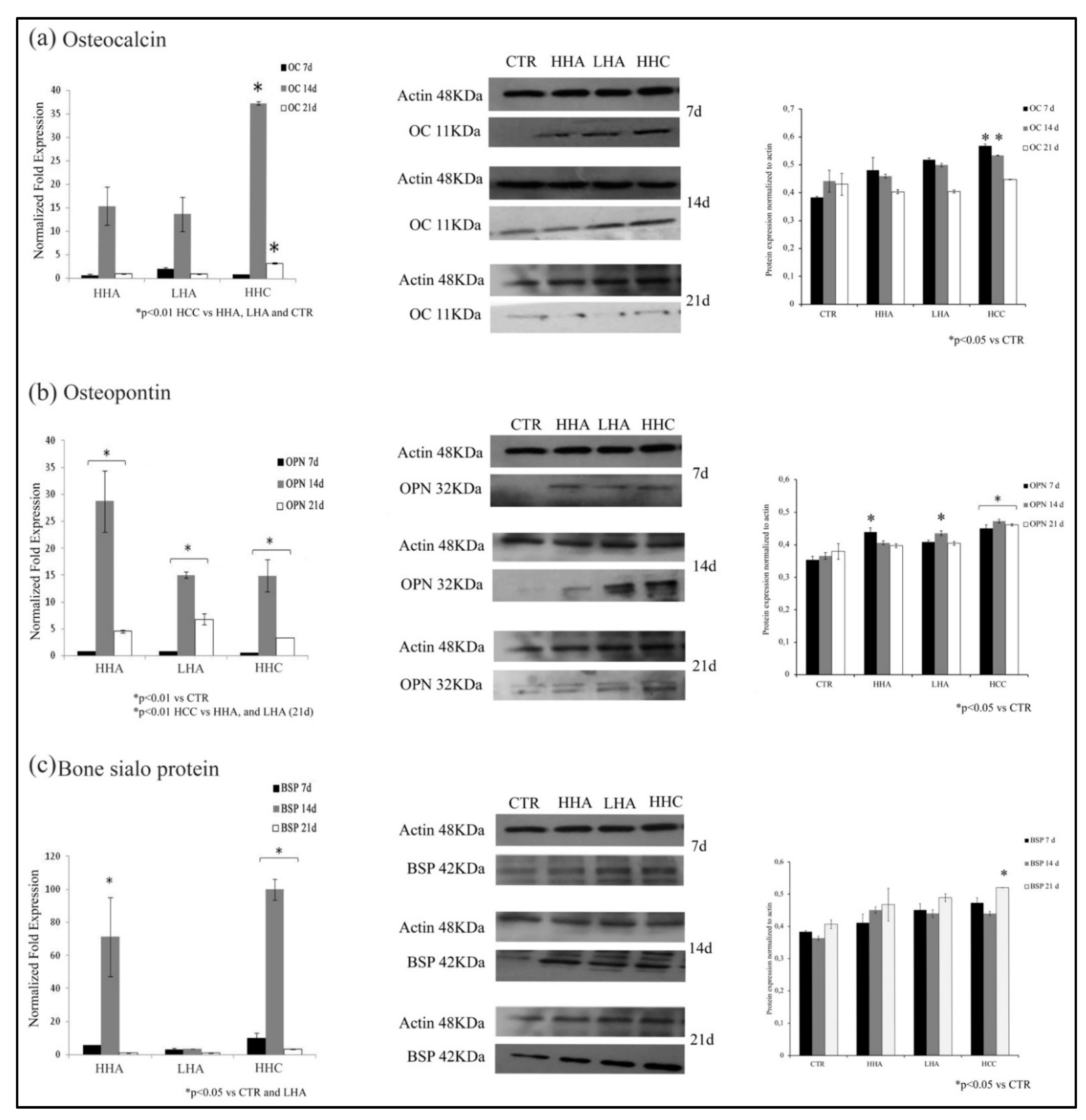
3.2. Hyaluronan-Based Gels Positively Affected the Expression of the Osteogenic Markers
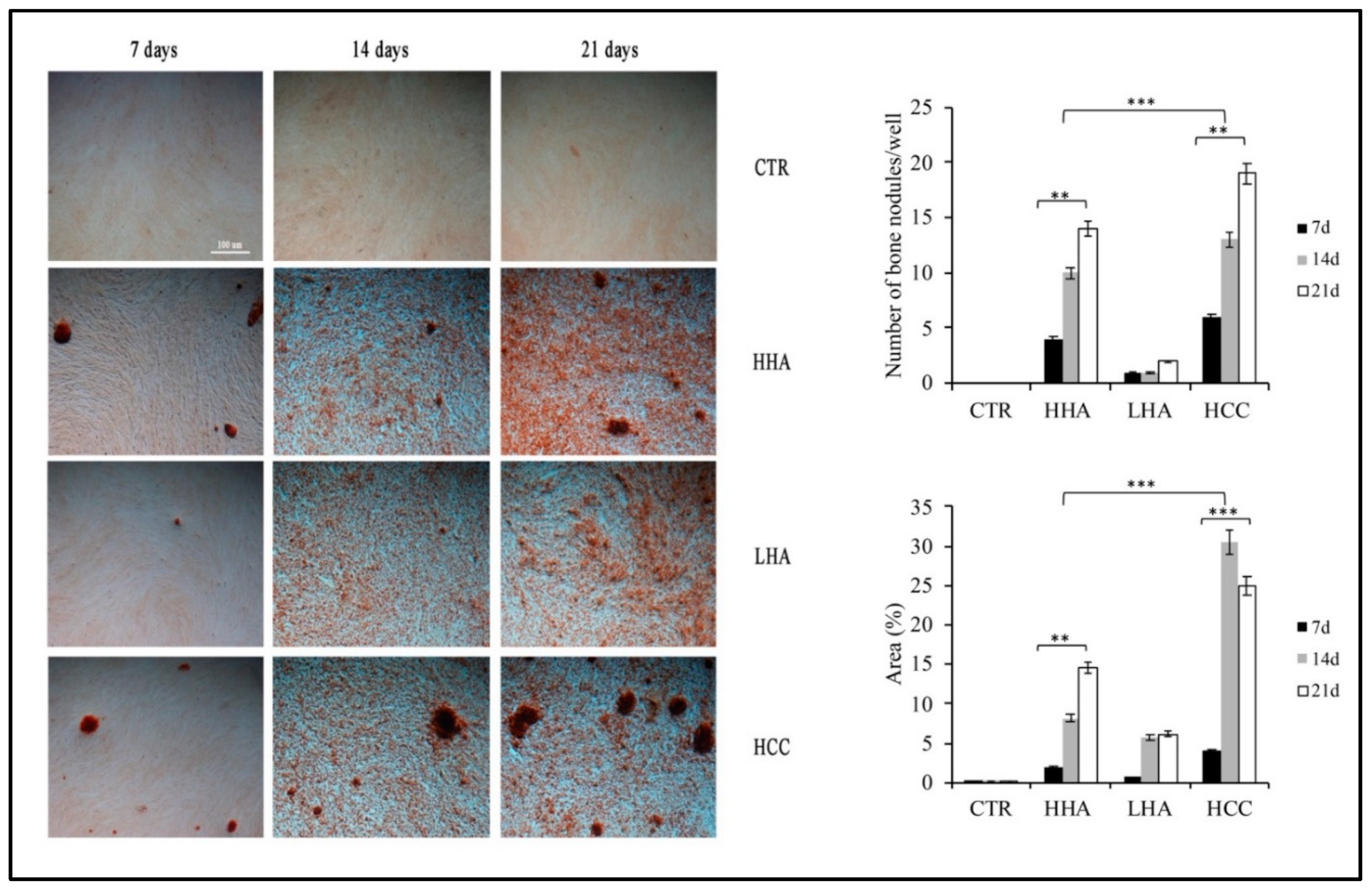
3.3. Hyaluronan-Based Gels Induced an Increase of Calcification Nodules in hDPSCs
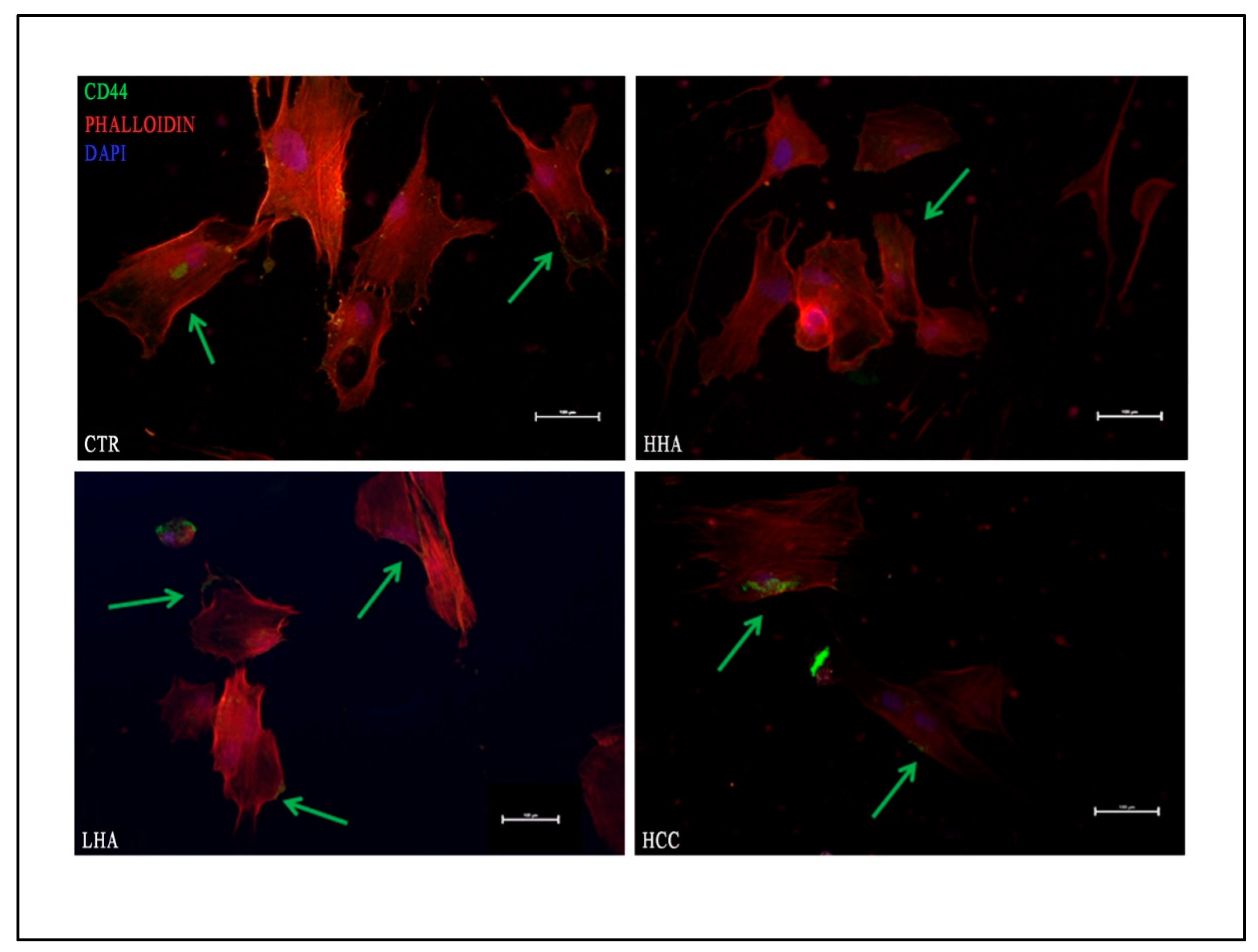
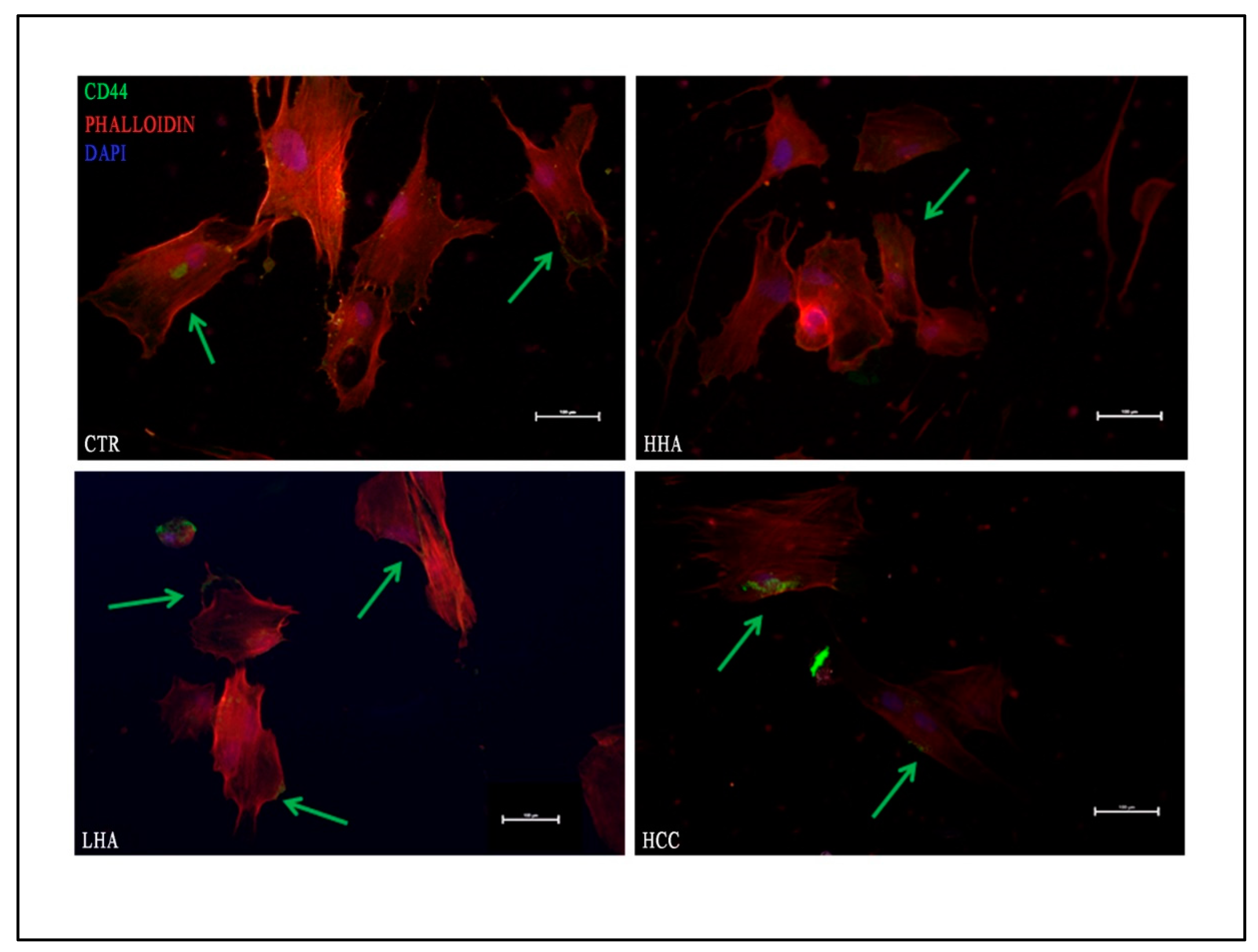
3.4. Hyaluronan-Based Gels Promoted Expression of CD44 in hDPSCs
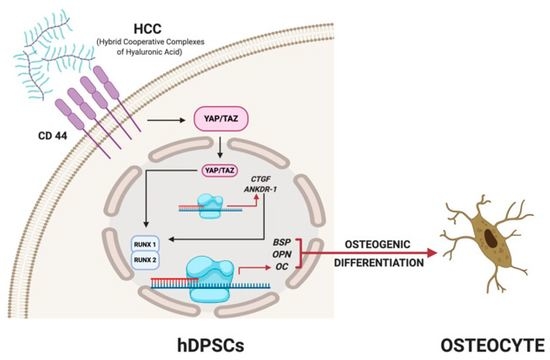
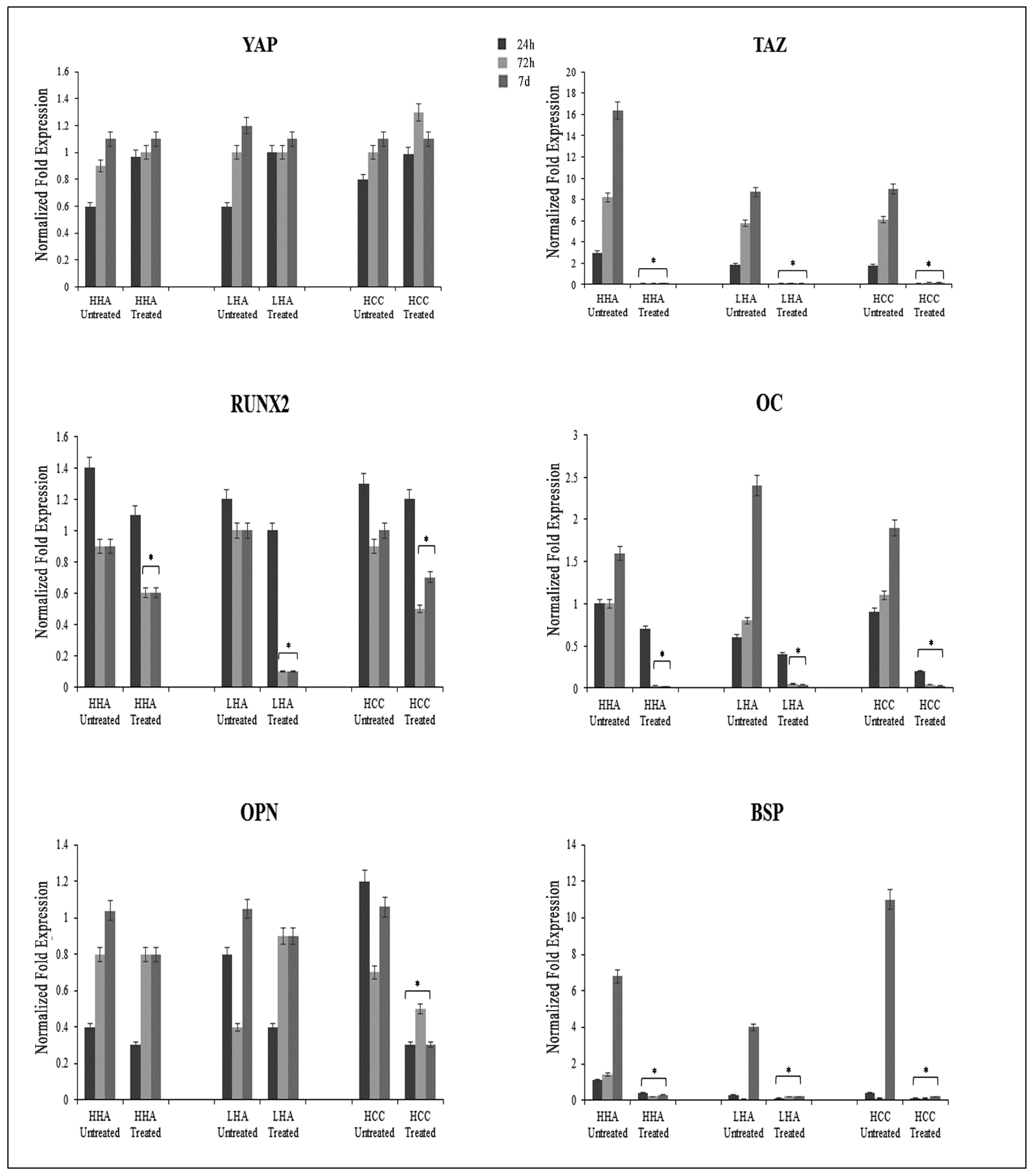
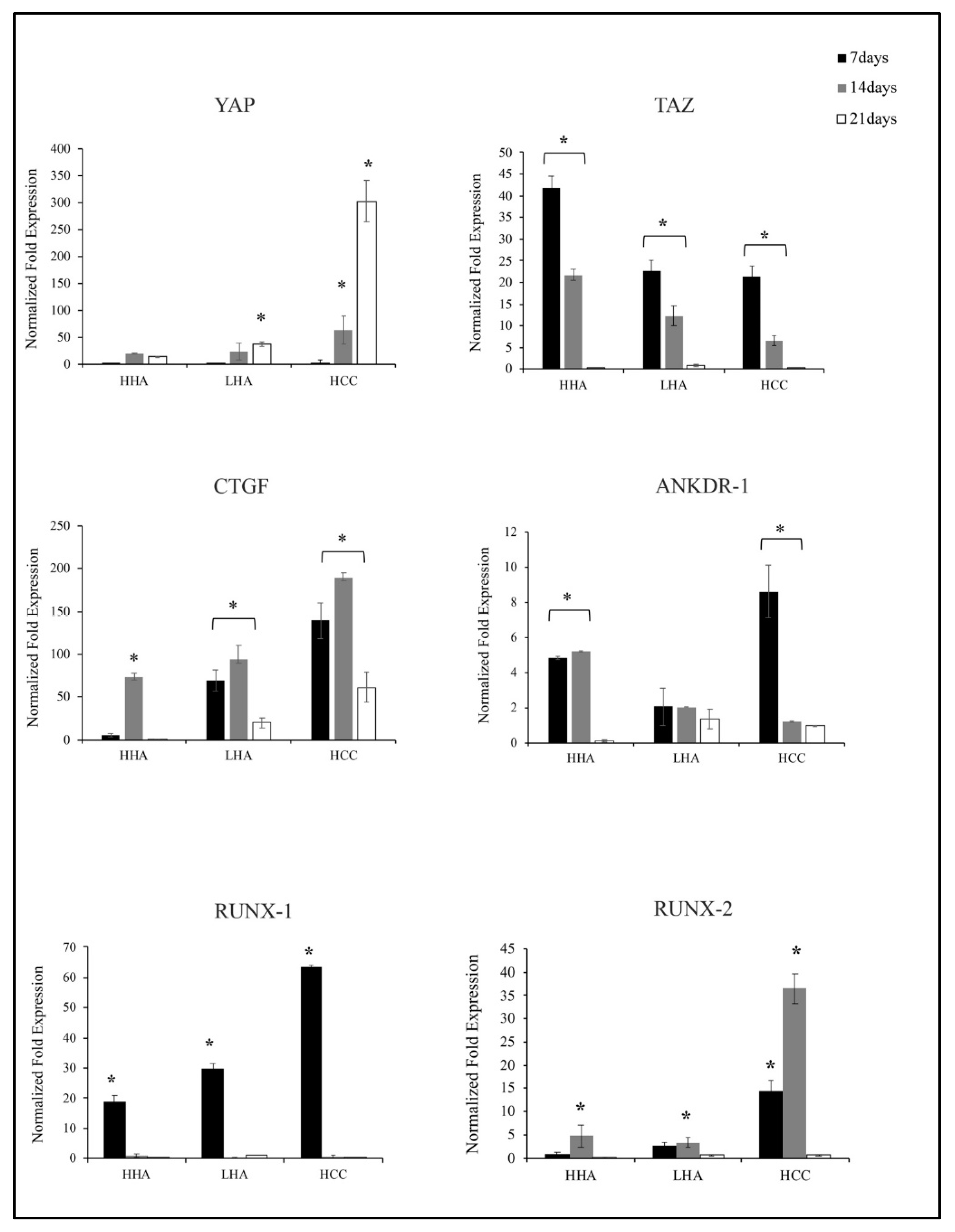
3.5. Hyaluronan-Based Gels Positively Regulated Gene Expression of YAP/TAZ, CTGF, ANKDR-1 and RUNX-1 and RUNX-2 in hDPSCs
3.6. Inhibition of YAP/TAZ Pathway Is Associated with a Reduction of Osteogenic Differentiation Exercised by Hyaluronans
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Toole, B.P. Hyaluronan in morphogenesis. Semin. Cell Dev. Biol. 2001, 12, 79–87. [Google Scholar] [CrossRef]
- Knudson, C.B.; Knudson, W. Hyaluronan-binding proteins in development, tissue homeostasis, and disease. FASEB J. 1993, 7, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Ashry, E.S.H.E.; Aly, M.R.E. Synthesis and biological relevance of N-acetylglucosamine-containing oligosaccharides. Pure Appl. Chem. 2007, 12, 2229–2242. [Google Scholar] [CrossRef]
- Huang, L.; Cheng, Y.Y.; Koo, P.L.; Lee, K.M.; Qin, L.; Cheng, J.C.Y.; Kumta, S.M. The effect of hyaluronan on osteoblast proliferation and differentiation in rat calvarial-derived cell cultures. J. Biomed. Mater. Res. 2003, 66A, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Stellavato, A.; Corsuto, L.; D’Agostino, A.; la Gatta, A.; Diana, P.; Bernini, P.; de Rosa, M.; Schiraldi, C. Hyaluronan hybrid cooperative complexes as a novel frontier for cellular bioprocesses re-activation. PLoS ONE 2016, 11, e0163510. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.L.H.; Chen, Y.-H.A.; Zhuo, Z.-Y.; Hsieh, Y.-T.; Yang, C.-L.; Chen, W.-T.; Lin, J.-Y.; Lin, Y.-X.; Jiang, J.-T.; Zhuang, C.-H.; et al. Medical applications of collagen and hyaluronan in regenerative medicine. Adv. Exp. Med. Biol. 2018, 1077, 285–306. [Google Scholar] [CrossRef] [PubMed]
- Stellavato, A.; la Noce, M.; Corsuto, L.; Pirozzi, A.V.A.; de Rosa, M.; Papaccio, G.; Schiraldi, C.; Tirino, V. Hybrid complexes of high and low molecular weight hyaluronans highly enhance HASCs differentiation: Implication for facial bioremodelling. Cell. Physiol. Biochem. 2017, 44, 1078–1092. [Google Scholar] [CrossRef] [Green Version]
- Alessio, N.; Stellavato, A.; Squillaro, T.; del Gaudio, S.; di Bernardo, G.; Peluso, G.; de Rosa, M.; Schiraldi, C.; Galderisi, U. Hybrid complexes of high and low molecular weight hyaluronan delay in vitro replicative senescence of mesenchymal stromal cells: A pilot study for future therapeutic application. Aging 2018, 10, 1575–1585. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise review: Multifaceted characterization of human mesenchymal stem cells for use in regenerative medicine. Stem Cells Transl. Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef] [Green Version]
- D’Aquino, R.; Graziano, A.; Sampaolesi, M.; Laino, G.; Pirozzi, G.; de Rosa, A.; Papaccio, G. Human postnatal dental pulp cells co-differentiate into osteoblasts and endotheliocytes: A pivotal synergy leading to adult bone tissue formation. Cell Death Differ. 2007, 14, 1162–1171. [Google Scholar] [CrossRef] [Green Version]
- Laino, G.; D’Aquino, R.; Graziano, A.; Lanza, V.; Carinci, F.; Naro, F.; Pirozzi, G.; Papaccio, G. A new population of human adult dental pulp stem cells: A useful source of living autologous fibrous bone tissue (LAB). J. Bone Miner. Res. 2005, 20, 1394–1402. [Google Scholar] [CrossRef]
- Yamada, Y.; Fujimoto, A.; Ito, A.; Yoshimi, R.; Ueda, M. Cluster analysis and gene expression profiles: A cDNA microarray system-based comparison between human dental pulp stem cells (hDPSCs) and human mesenchymal stem cells (hMSCs) for tissue engineering cell therapy. Biomaterials 2006, 27, 3766–3781. [Google Scholar] [CrossRef]
- Tirino, V.; Paino, F.; de Rosa, A.; Papaccio, G. Identification, Isolation, characterization, and banking of human dental pulp stem cells. Methods Mol. Biol. 2012, 879, 443–463. [Google Scholar] [CrossRef]
- Zhang, W.; Walboomers, X.F.; Shi, S.; Fan, M.; Jansen, J.A. Multilineage differentiation potential of stem cells derived from human dental pulp after cryopreservation. Tissue Eng. 2006, 12, 2813–2823. [Google Scholar] [CrossRef]
- Caliari, S.; Vega, S.L.; Kwon, M.; Soulas, E.M.; Burdick, J.A. Dimensionality and spreading influence MSC YAP/TAZ signaling in hydrogel environments. Biomaterials 2016, 103, 314–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragona, M.; Panciera, T.; Manfrin, A.; Giulitti, S.; Michielin, F.; Elvassore, N.; Dupont, S.; Piccolo, S. A mechanical checkpoint controls multicellular growth through YAP/TAZ regulation by actin-processing factors. Cell 2013, 154, 1047–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef]
- Low, B.C.; Pan, C.Q.; Shivashankar, G.; Bershadsky, A.; Sudol, M.; Sheetz, M. YAP/TAZ as mechanosensors and mechanotransducers in regulating organ size and tumor growth. FEBS Lett. 2014, 588, 2663–2670. [Google Scholar] [CrossRef] [Green Version]
- Stellavato, A.; Pirozzi, A.V.A.; Diana, P.; Reale, S.; Vassallo, V.; Fusco, A.; Donnarumma, G.; de Rosa, M.; Schiraldi, C. Hyaluronic acid and chondroitin sulfate, alone or in combination, efficiently counteract induced bladder cell damage and inflammation. PLoS ONE 2019, 14, e0218475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heng, B.C.; Zhang, X.; Aubel, D.; Bai, Y.; Li, X.; Wei, Y.; Fussenegger, M.; Deng, X. Role of YAP/TAZ in cell lineage fate determination and related signaling pathways. Front. Cell Dev. Biol. 2020, 8, 735. [Google Scholar] [CrossRef]
- Yi, T.; Weiss, S.J. Snail/slug-YAP/TAZ complexes cooperatively regulate mesenchymal stem cell function and bone formation. Cell Cycle 2017, 16, 399–405. [Google Scholar]
- Piccolo, S.; Dupont, S.; Cordenonsi, M. The biology of YAP/TAZ: Hippo signaling and beyond. Physiol. Rev. 2014, 94, 1287–1312. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.X.; Xiong, L.; Zhao, K.; Zeng, P.; Wang, B.; Tang, F.L.; Sun, D.; Guo, H.H.; Yang, X.; Cui, S.; et al. YAP promotes osteogenesis and suppresses adipogenic differentiation by regulating β-catenin signaling. Bone Res. 2018, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Song, R.; Murphy, M.; Li, C.; Ting, K.; Soo, C.; Zheng, Z. Current development of biodegradable polymeric materials for biomedical applications. Drug Des. Dev. Ther. 2018, 12, 3117–3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, C.; Peng, S.; Feng, P.; Shuai, C. Bone biomaterials and interactions with stem cells. Bone Res. 2017, 5, 17059. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.; Manescu, A.; Langer, M.; Rustichelli, F.; Desiderio, V.; Paino, F.; de Rosa, A.; Laino, L.; D’Aquino, R.; Tirino, V.; et al. Three years after transplants in human mandibles, histological and in-line holotomography revealed that stem cells regenerated a compact rather than a spongy bone: Biological and clinical implications. Stem Cells Transl. Med. 2013, 2, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; McKee, M.D.; Nanci, A.; Sodek, J. Bone sialoprotein mRNA expression and ultrastructural localization in fetal porcine calvarial bone: Comparisons with osteopontin. Histochem. J. 1994, 26, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Oldberg, A.; Franzén, A.; Heinegård, D. The primary structure of a cell-binding bone sialoprotein. J. Biol. Chem. 1988, 263, 19430–19432. [Google Scholar] [CrossRef]
- Gordon, J.A.; Tye, C.; Sampaio, A.V.; Underhill, T.M.; Hunter, G.K.; Goldberg, H.A. Bone sialoprotein expression enhances osteoblast differentiation and matrix mineralization in vitro. Bone 2007, 41, 462–473. [Google Scholar] [CrossRef]
- Chen, T.L.; Bates, R.L.; Dudley, A.; Hammonds, R.G., Jr.; Amento, E.P. Bone morphogenetic protein-2b stimulation of growth and osteogenic phenotypes in rat osteoblast-like cells: Comparison with TGF-β1. J. Bone Miner. Res. 1991, 6, 1387–1393. [Google Scholar] [CrossRef]
- Hunter, G.K.; Goldberg, H.A. Modulation of crystal formation by bone phosphoproteins: Role of glutamic acid-rich sequences in the nucleation of hydroxyapatite by bone sialoprotein. Biochem. J. 1994, 302, 175–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, J.; Wang, C.; Zhang, D.; Wang, B.; Hou, W.; Zhou, Y. Osteopontin in bone metabolism and bone diseases. Med Sci. Monit. 2020, 26, e919159-1. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, S.; Karnad, A.; Freeman, J.W. The biology and role of CD44 in cancer progression: Therapeutic implications. J. Hematol. Oncol. 2018, 11, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Stamenkovic, I.; Yu, Q. CD44 attenuates activation of the hippo signaling pathway and is a prime therapeutic target for glioblastoma. Cancer Res. 2010, 70, 2455–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Name | Sequence | Ta |
|---|---|---|---|
| OC | Osteocalcin | Forward CTCCACATCCTCGCCCTATTG Reverse CTTGGACACAAAGGCTGCAC | 58 °C |
| OPN | Osteopontin | Forward GCCGAGGTGATAAGTGTGGTT Reverse TGAGGTGATGTCCTCGTTCTG | 58 °C |
| BSP | Bone sialo protein | Forward CTGGCACAGGGTATACAGGGTAG Reverse ACTGGTGCCGTTTATGCCTTG | 60 °C |
| YAP | Yes-associated protein 1 | Forward CAACTCCAACCAGCAGCAAC Reverse TTGGTAACTGGCTACGCAGG | 55 °C |
| TAZ | Transcriptional co-activator with PDZ-binding motif | Forward TGGACCAAGTACATGAACCACC Reverse TGCCTTCTATGCTCCCTCCT | 55 °C |
| hCTGF | human-Connective tissue growth factor | Forward AGGAGTGGGTGTGTGACGA Reverse CCAGGCAGTTGGCTCTAATC | 57 °C |
| hANKDR1 | human-Ankyrin repeat domain-containing protein 1 | Forward AGTAGAGGAACTGGTCACTGG Reverse TGGGCTAGAAGTGTCTTCAGAT | 57 °C |
| RUNX-1 | Runt domain transcription factors 1 | Forward AACCCAGCATAGTGGTCAGC Reverse CATGGCTGCGGTAGCATTTC | 57 °C |
| RUNX-2 | Runt domain transcription factors 2 | Forward ACCAGCAGCACTCCATATCTCTAC Reverse CTTCCATCAGCGTCAACACCATC | 57 °C |
| HRPT | Hypoxanthine-guanine phosphoribosyltransferase | Forward TGACCTTGATTTATTTTGCATACC Reverse CGAGCAAGACGTTCAGTCCT | 60 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
La Noce, M.; Stellavato, A.; Vassallo, V.; Cammarota, M.; Laino, L.; Desiderio, V.; Del Vecchio, V.; Nicoletti, G.F.; Tirino, V.; Papaccio, G.; et al. Hyaluronan-Based Gel Promotes Human Dental Pulp Stem Cells Bone Differentiation by Activating YAP/TAZ Pathway. Cells 2021, 10, 2899. https://doi.org/10.3390/cells10112899
La Noce M, Stellavato A, Vassallo V, Cammarota M, Laino L, Desiderio V, Del Vecchio V, Nicoletti GF, Tirino V, Papaccio G, et al. Hyaluronan-Based Gel Promotes Human Dental Pulp Stem Cells Bone Differentiation by Activating YAP/TAZ Pathway. Cells. 2021; 10(11):2899. https://doi.org/10.3390/cells10112899
Chicago/Turabian StyleLa Noce, Marcella, Antonietta Stellavato, Valentina Vassallo, Marcella Cammarota, Luigi Laino, Vincenzo Desiderio, Vitale Del Vecchio, Giovanni Francesco Nicoletti, Virginia Tirino, Gianpaolo Papaccio, and et al. 2021. "Hyaluronan-Based Gel Promotes Human Dental Pulp Stem Cells Bone Differentiation by Activating YAP/TAZ Pathway" Cells 10, no. 11: 2899. https://doi.org/10.3390/cells10112899
APA StyleLa Noce, M., Stellavato, A., Vassallo, V., Cammarota, M., Laino, L., Desiderio, V., Del Vecchio, V., Nicoletti, G. F., Tirino, V., Papaccio, G., Schiraldi, C., & Ferraro, G. A. (2021). Hyaluronan-Based Gel Promotes Human Dental Pulp Stem Cells Bone Differentiation by Activating YAP/TAZ Pathway. Cells, 10(11), 2899. https://doi.org/10.3390/cells10112899