
Taste Receptors: The Gatekeepers of the Airway Epithelium


Abstract
:1. Introduction
2. Bitter and Sweet Taste Receptors in the Respiratory Tract
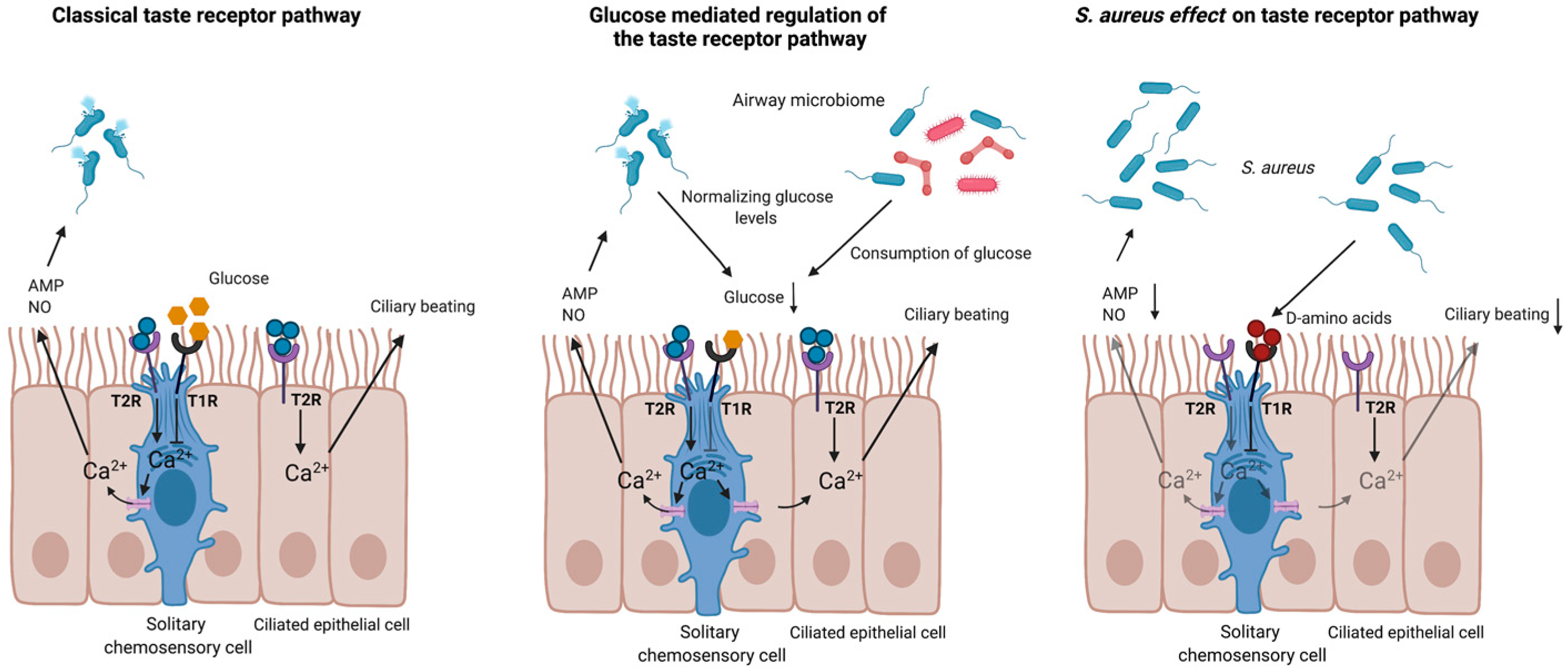
2.1. Taste Receptors in the Upper Respiratory Tract
2.2. Taste Receptor Polymorphisms in the Upper Respiratory Tract
2.3. Taste Receptors in the Lower Airways
2.4. Taste Receptor Polymorphisms in the Diseased Lower Respiratory Tract
3. Clinical Application of Taste Receptors for the Airways
Challenges of T1R and T2R Therapies
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Chandrashekar, J.; Hoon, M.A.; Ryba, N.J.; Zuker, C.S. The receptors and cells for mammalian taste. Nature 2006, 444, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Harmon, C.P.; Deng, D.; Breslin, P.A.S. Bitter Taste Receptors (T2Rs) are Sentinels that Coordinate Metabolic and Immunological Defense Responses. Curr. Opin. Physiol. 2021, 20, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Tizzano, M.; Redding, K.; He, J.; Peng, X.; Jiang, P.; Xu, X.; Zhou, X.; Margolskee, R.F. Gingival solitary chemosensory cells are immune sentinels for periodontitis. Nat. Commun. 2019, 10, 4496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, J.M.; Margolskee, R.F. Taste cells of the gut and gastrointestinal chemosensation. Mol. Interv. 2008, 8, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Margolskee, R.F.; Dyer, J.; Kokrashvili, Z.; Salmon, K.S.; Ilegems, E.; Daly, K.; Maillet, E.L.; Ninomiya, Y.; Mosinger, B.; Shirazi-Beechey, S.P. T1R3 and gustducin in gut sense sugars to regulate expression of Na+-glucose cotransporter 1. Proc. Natl. Acad. Sci. USA 2007, 104, 15075–15080. [Google Scholar] [CrossRef] [Green Version]
- San Gabriel, A.; Uneyama, H. Amino acid sensing in the gastrointestinal tract. Amino Acids. 2013, 45, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Steensels, S.; Depoortere, I. Chemoreceptors in the Gut. Annu Rev. Physiol. 2018, 10, 117–141. [Google Scholar] [CrossRef] [PubMed]
- Roura, E.; Depoortere, I.; Navarro, M. Review: Chemosensing of nutrients and non-nutrients in the human and porcine gastrointestinal tract. Animal 2019, 13, 2714–2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modvig, I.M.; Kuhre, R.E.; Holst, J.J. Peptone-mediated glucagon-like peptide-1 secretion depends on intestinal absorption and activation of basolaterally located Calcium-Sensing Receptors. Physiol. Rep. 2019, 7, e14056. [Google Scholar] [CrossRef]
- Wu, S.V.; Rozengurt, N.; Yang, M.; Young, S.H.; Sinnett-Smith, J.; Rozengurt, E. Expression of bitter taste receptors of the T2R family in the gastrointestinal tract and enteroendocrine STC-1 cells. Proc. Natl. Acad. Sci. USA 2002, 99, 2392–2397. [Google Scholar] [CrossRef] [Green Version]
- Glendinning, J.I.; Yiin, Y.M.; Ackroff, K.; Sclafani, A. Intragastric infusion of denatonium conditions flavor aversions and delays gastric emptying in rodents. Physiol. Behav. 2008, 93, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Janssen, S.; Laermans, J.; Verhulst, P.J.; Thijs, T.; Tack, J.; Depoortere, I. Bitter taste receptors and α-gustducin regulate the secretion of ghrelin with functional effects on food intake and gastric emptying. Proc. Natl. Acad. Sci. USA 2011, 108, 2094–2099. [Google Scholar] [CrossRef] [Green Version]
- Howitt, M.R.; Lavoie, S.; Michaud, M.; Blum, A.M.; Tran, S.V.; Weinstock, J.V.; Gallini, C.A.; Redding, K.; Margolskee, R.F.; Osborne, L.C.; et al. Tuft cells, taste-chemosensory cells, orchestrate parasite type 2 immunity in the gut. Science 2016, 351, 1329–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, J.; Casanova-Martí, À.; Depoortere, I.; Blay, M.T.; Terra, X.; Pinent, M.; Ardévol, A. Subchronic treatment with grape-seed phenolics inhibits ghrelin production despite a short-term stimulation of ghrelin secretion produced by bitter-sensing flavanols. Mol. Nutr. Food Res. 2016, 60, 2554–2564. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhou, M. Depletion of bitter taste transduction leads to massive spermatid loss in transgenic mice. Mol. Hum. Reprod. 2012, 18, 289–297. [Google Scholar] [CrossRef]
- Deckmann, K.; Filipski, K.; Krasteva-Christ, G.; Fronius, M.; Althaus, M.; Rafiq, A.; Papadakis, T.; Renno, L.; Jurastow, I.; Wessels, L.; et al. Bitter triggers acetylcholine release from polymodal urethral chemosensory cells and bladder reflexes. Proc. Natl. Acad. Sci. USA 2014, 111, 8287–8292. [Google Scholar] [CrossRef] [Green Version]
- Foster, S.R.; Porrello, E.R.; Purdue, B.; Chan, H.W.; Voigt, A.; Frenzel, S.; Hannan, R.D.; Moritz, K.M.; Simmons, D.G.; Molenaar, P.; et al. Expression, regulation and putative nutrient-sensing function of taste GPCRs in the heart. PLoS ONE 2013, 8, e64579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Vrontakis, M.; Parkinson, F.; Chelikani, P. Functional bitter taste receptors are expressed in brain cells. Biochem. Biophys. Res. Commun. 2011, 406, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Dehkordi, O.; Rose, J.E.; Fatemi, M.; Allard, J.S.; Balan, K.V.; Young, J.K.; Fatima, S.; Millis, R.M.; Jayam-Trouth, A. Neuronal expression of bitter taste receptors and downstream signaling molecules in the rat brainstem. Brain Res. 2012, 1475, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tomás, J.; Santos, C.R.; Quintela, T.; Gonçalves, I. “Tasting” the cerebrospinal fluid: Another function of the choroid plexus? Neuroscience 2016, 320, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Kofonow, J.M.; Rosen, P.L.; Siebert, A.P.; Chen, B.; Doghramji, L.; Xiong, G.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; et al. Bitter and sweet taste receptors regulate human upper respiratory innate immunity. J. Clin. Investig. 2014, 124, 1393–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, R.M.; Lee, R.J. Taste Receptors in Upper Airway Innate Immunity. Nutrients 2019, 11, 2017. [Google Scholar] [CrossRef] [Green Version]
- Triantafillou, V.; Workman, A.D.; Patel, N.N.; Maina, I.W.; Tong, C.C.L.; Kuan, E.C.; Kennedy, D.W.; Palmer, J.N.; Adappa, N.D.; Waizel-Haiat, S.; et al. Broncho-Vaxom® (OM-85 BV) soluble components stimulate sinonasal innate immunity. Int. Forum. Allergy Rhinol. 2019, 9, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Devillier, P.; Naline, E.; Grassin-Delyle, S. The pharmacology of bitter taste receptors and their role in human airways. Pharmacol. Ther. 2015, 155, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Investig. 2012, 122, 4145–4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsmark-Pietras, C.; James, A.; Konradsen, J.R.; Nordlund, B.; Söderhäll, C.; Pulkkinen, V.; Pedroletti, C.; Daham, K.; Kupczyk, M.; Dahlén, B.; et al. Transcriptome analysis reveals upregulation of bitter taste receptors in severe asthmatics. Eur. Respir. J. 2013, 42, 65–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Ishimaru, Y. Oral and extra-oral taste perception. Semin. Cell Dev. Biol. 2013, 24, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Conte, C.; Ebeling, M.; Marcuz, A.; Nef, P.; Andres-Barquin, P.J. Identification and characterization of human taste receptor genes belonging to the TAS2R family. Cytogenet. Genome Res. 2002, 98, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Brockhoff, A.; Behrens, M.; Massarotti, A.; Appendino, G.; Meyerhof, W. Broad tuning of the human bitter taste receptor hTAS2R46 to various sesquiterpene lactones, clerodane and labdane diterpenoids, strychnine, and denatonium. J. Agric. Food Chem. 2007, 55, 6236–6243. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Larson, E.D.; Anderson, C.B.; Agarwal, P.; Frank, D.N.; Kinnamon, S.C.; Ramakrishnan, V.R. Expression of Bitter Taste Receptors and Solitary Chemosensory Cell Markers in the Human Sinonasal Cavity. Chem. Sens. 2019, 44, 483–495. [Google Scholar] [CrossRef]
- Saunders, C.J.; Christensen, M.; Finger, T.E.; Tizzano, M. Cholinergic neurotransmission links solitary chemosensory cells to nasal inflammation. Proc. Natl. Acad. Sci. USA 2014, 111, 6075–6080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, D.A.; Wang, W.C.; McIlmoyle, E.L.; Robinett, K.S.; Schillinger, R.M.; An, S.S.; Sham, J.S.; Liggett, S.B. Bitter taste receptors on airway smooth muscle bronchodilate by localized calcium signaling and reverse obstruction. Nat. Med. 2010, 16, 1299–1304. [Google Scholar] [CrossRef] [Green Version]
- Bourke, J.E.; Donovan, C.; FitzPatrick, M.; Simoons, M. Bitter taste receptor agonists are novel bronchodilators of small airways in mouse lung slices. Am. J. Respir. Crit. Care Med. 2013, 187, A2288. [Google Scholar]
- Pulkkinen, V.; Manson, M.L.; Säfholm, J.; Adner, M.; Dahlén, S.E. The bitter taste receptor (TAS2R) agonists denatonium and chloroquine display distinct patterns of relaxation of the guinea pig trachea. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 303, L956–L966. [Google Scholar] [CrossRef] [PubMed]
- Morice, A.H.; Bennett, R.T.; Chaudhry, M.A.; Cowen, M.E.; Griffin, S.C.; Loubani, M. Effect of bitter tastants on human bronchi. Nat. Med. 2011, 17, 775. [Google Scholar] [CrossRef] [PubMed]
- Belvisi, M.G.; Dale, N.; Birrell, M.A.; Canning, B.J. Bronchodilator activity of bitter tastants in human tissue. Nat. Med. 2011, 17, 776. [Google Scholar] [CrossRef] [PubMed]
- Grassin-Delyle, S.; Abrial, C.; Fayad-Kobeissi, S.; Brollo, M.; Faisy, C.; Alvarez, J.C.; Naline, E.; Devillier, P. The expression and relaxant effect of bitter taste receptors in human bronchi. Respir. Res. 2013, 14, 134. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.S.; Ben-Shahar, Y.; Moninger, T.O.; Kline, J.N.; Welsh, M.J. Motile cilia of human airway epithelia are chemosensory. Science 2009, 325, 1131–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekoff, M.; Choi, J.H.; James, A.; Dahlén, B.; Nilsson, G.; Dahlén, S.E. Bitter taste receptor (TAS2R) agonists inhibit IgE-dependent mast cell activation. J. Allergy Clin. Immunol. 2014, 134, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Vrančić, M.; Banjanac, M.; Nujić, K.; Bosnar, M.; Murati, T.; Munić, V.; Stupin Polančec, D.; Belamarić, D.; Parnham, M.J.; Eraković Haber, V. Azithromycin distinctively modulates classical activation of human monocytes in vitro. Br. J. Pharmacol. 2012, 165, 1348–1360. [Google Scholar] [CrossRef] [Green Version]
- Ogino, H.; Fujii, M.; Ono, M.; Maezawa, K.; Hori, S.; Kizu, J. In vivo and in vitro effects of fluoroquinolones on lipopolysaccharide-induced pro-inflammatory cytokine production. J. Infect. Chemother. 2009, 15, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.H.; Choi, J.H.; Byun, M.S.; Jue, D.M. Chloroquine inhibits production of TNF-alpha, IL-1beta and IL-6 from lipopolysaccharide-stimulated human monocytes/macrophages by different modes. Rheumatology 2006, 45, 703–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasutomi, M.; Ohshima, Y.; Omata, N.; Yamada, A.; Iwasaki, H.; Urasaki, Y.; Mayumi, M. Erythromycin differentially inhibits lipopolysaccharide- or poly(I:C)-induced but not peptidoglycan-induced activation of human monocyte-derived dendritic cells. J. Immunol. 2005, 175, 8069–8076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.Y.; Jue, D.M. Chloroquine inhibits processing of tumor necrosis factor in lipopolysaccharide-stimulated RAW 264.7 macrophages. J. Immunol. 1997, 158, 4901–4907. [Google Scholar] [PubMed]
- Harrington, E.O.; Vang, A.; Braza, J.; Shil, A.; Chichger, H. Activation of the sweet taste receptor, T1R3, by the artificial sweetener sucralose regulates the pulmonary endothelium. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 314, L165–L176. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Cohen, N.A. Bitter and sweet taste receptors in the respiratory epithelium in health and disease. J. Mol. Med. 2014, 92, 1235–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, A.P.; Shah, S.D.; Michael, J.V.; Deshpande, D.A. Bitter Taste Receptors for Asthma Therapeutics. Front. Physiol. 2019, 10, 884. [Google Scholar] [CrossRef] [Green Version]
- Grassin-Delyle, S.; Naline, E.; Devillier, P. Taste receptors in asthma. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Triantafillou, V.; Workman, A.D.; Kohanski, M.A.; Cohen, N.A. Taste Receptor Polymorphisms and Immune Response: A Review of Receptor Genotypic-Phenotypic Variations and Their Relevance to Chronic Rhinosinusitis. Front. Cell Infect. Microbiol. 2018, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Maina, I.W.; Workman, A.D.; Cohen, N.A. The role of bitter and sweet taste receptors in upper airway innate immunity: Recent advances and future directions. World J. Otorhinolaryngol. Head Neck Surg. 2018, 4, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Hariri, B.M.; McMahon, D.B.; Chen, B.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; Jiang, P.; Margolskee, R.F.; et al. Bacterial d-amino acids suppress sinonasal innate immunity through sweet taste receptors in solitary chemosensory cells. Sci. Signal. 2017, 10, eaam7703. [Google Scholar] [CrossRef] [Green Version]
- Kim, U.; Wooding, S.; Ricci, D.; Jorde, L.B.; Drayna, D. Worldwide haplotype diversity and coding sequence variation at human bitter taste receptor loci. Hum. Mutat. 2005, 26, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Fokkens, W.J.; Lund, V.J.; Mullol, J.; Bachert, C.; Alobid, I.; Baroody, F.; Cohen, N.; Cervin, A.; Douglas, R.; Gevaert, P.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2012. Rhinology 2012, 50, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bufe, B.; Breslin, P.A.; Kuhn, C.; Reed, D.R.; Tharp, C.D.; Slack, J.P.; Kim, U.K.; Drayna, D.; Meyerhof, W. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr. Biol. 2005, 15, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Kim, U.K.; Drayna, D. Genetics of individual differences in bitter taste perception: Lessons from the PTC gene. Clin Genet. 2005, 67, 275–280, Erratum in 2005, 67, 534. [Google Scholar] [CrossRef] [PubMed]
- Adappa, N.D.; Truesdale, C.M.; Workman, A.D.; Doghramji, L.; Mansfield, C.; Kennedy, D.W.; Palmer, J.N.; Cowart, B.J.; Cohen, N.A. Correlation of T2R38 taste phenotype and in vitro biofilm formation from nonpolypoid chronic rhinosinusitis patients. Int. Forum. Allergy Rhinol. 2016, 6, 783–791. [Google Scholar] [CrossRef]
- Adappa, N.D.; Howland, T.J.; Palmer, J.N.; Kennedy, D.W.; Doghramji, L.; Lysenko, A.; Reed, D.R.; Lee, R.J.; Cohen, N.A. Genetics of the taste receptor T2R38 correlates with chronic rhinosinusitis necessitating surgical intervention. Int. Forum. Allergy Rhinol. 2013, 3, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Adappa, N.D.; Zhang, Z.; Palmer, J.N.; Kennedy, D.W.; Doghramji, L.; Lysenko, A.; Reed, D.R.; Scott, T.; Zhao, N.W.; Owens, D.; et al. The bitter taste receptor T2R38 is an independent risk factor for chronic rhinosinusitis requiring sinus surgery. Int. Forum. Allergy Rhinol. 2014, 4, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Mfuna Endam, L.; Filali-Mouhim, A.; Boisvert, P.; Boulet, L.P.; Bossé, Y.; Desrosiers, M. Genetic variations in taste receptors are associated with chronic rhinosinusitis: A replication study. Int. Forum. Allergy Rhinol. 2014, 4, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopez, O.; Panduro, A.; Martinez-Lopez, E.; Roman, S. Sweet Taste Receptor TAS1R2 Polymorphism (Val191Val) Is Associated with a Higher Carbohydrate Intake and Hypertriglyceridemia among the Population of West Mexico. Nutrients 2016, 8, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, S.; Wabnitz, G.H.; Kahle, N.A.; Stegmaier, S.; Prior, B.; Giese, T.; Gaida, M.M.; Samstag, Y.; Hänsch, G.M. Tasting Pseudomonas aeruginosa Biofilms: Human Neutrophils Express the Bitter Receptor T2R38 as Sensor for the Quorum Sensing Molecule N-(3-Oxododecanoyl)-l-Homoserine Lactone. Front. Immunol. 2015, 6, 369. [Google Scholar] [CrossRef] [Green Version]
- Tran, H.T.T.; Herz, C.; Ruf, P.; Stetter, R.; Lamy, E. Human T2R38 Bitter Taste Receptor Expression in Resting and Activated Lymphocytes. Front. Immunol. 2018, 9, 2949. [Google Scholar] [CrossRef]
- Sbarbati, A.; Merigo, F.; Benati, D.; Tizzano, M.; Bernardi, P.; Crescimanno, C.; Osculati, F. Identification and characterization of a specific sensory epithelium in the rat larynx. J. Comp. Neurol. 2004, 475, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Tizzano, M.; Cristofoletti, M.; Sbarbati, A.; Finger, T.E. Expression of taste receptors in solitary chemosensory cells of rodent airways. BMC Pulm. Med. 2011, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Perniss, A.; Liu, S.; Boonen, B.; Keshavarz, M.; Ruppert, A.L.; Timm, T.; Pfeil, U.; Soultanova, A.; Kusumakshi, S.; Delventhal, L.; et al. Chemosensory Cell-Derived Acetylcholine Drives Tracheal Mucociliary Clearance in Response to Virulence-Associated Formyl Peptides. Immunity 2020, 52, 683–699.e11. [Google Scholar] [CrossRef] [PubMed]
- Avau, B.; Rotondo, A.; Thijs, T.; Andrews, C.N.; Janssen, P.; Tack, J.; Depoortere, I. Targeting extra-oral bitter taste receptors modulates gastrointestinal motility with effects on satiation. Sci. Rep. 2015, 5, 15985. [Google Scholar] [CrossRef] [PubMed]
- Lasconi, C.; Pifferi, S.; Hernandez-Clavijo, A.; Merigo, F.; Cecchini, M.P.; Gonzalez-Velandia, K.Y.; Agostinelli, E.; Sbarbati, A.; Menini, A. Bitter tastants and artificial sweeteners activate a subset of epithelial cells in acute tissue slices of the rat trachea. Sci. Rep. 2019, 9, 8834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaida, M.M.; Dapunt, U.; Hänsch, G.M. Sensing developing biofilms: The bitter receptor T2R38 on myeloid cells. Pathog. Dis. 2016, 74, ftw004. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Yi, R.; Nayak, A.P.; Wang, N.; Tang, F.; Knight, M.J.; Pan, S.; Oliver, B.; Deshpande, D.A. Bitter Taste Receptor Agonists Mitigate Features of Allergic Asthma in Mice. Sci. Rep. 2017, 7, 46166. [Google Scholar] [CrossRef]
- Adappa, N.D.; Workman, A.D.; Hadjiliadis, D.; Dorgan, D.J.; Frame, D.; Brooks, S.; Doghramji, L.; Palmer, J.N.; Mansfield, C.; Reed, D.R.; et al. T2R38 genotype is correlated with sinonasal quality of life in homozygous ΔF508 cystic fibrosis patients. Int. Forum Allergy Rhinol. 2016, 6, 356–361. [Google Scholar] [CrossRef] [Green Version]
- Piatti, G.; Ambrosetti, U.; Robino, A.; Girotto, G.; Gasparini, P. Primary Ciliary Dyskinesia: The Impact of Taste Receptor (TAS2R38) Gene Polymorphisms on Disease Outcome and Severity. Int. Arch. Allergy Immunol. 2020, 181, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Hume, D.A.; Underhill, D.M.; Sweet, M.J.; Ozinsky, A.O.; Liew, F.Y.; Aderem, A. Macrophages exposed continuously to lipopolysaccharide and other agonists that act via toll-like receptors exhibit a sustained and additive activation state. BMC Immunol. 2001, 2, 11. [Google Scholar] [CrossRef]
- Patel, N.N.; Kohanski, M.A.; Maina, I.W.; Workman, A.D.; Herbert, D.R.; Cohen, N.A. Sentinels at the wall: Epithelial-derived cytokines serve as triggers of upper airway type 2 inflammation. Int. Forum Allergy Rhinol. 2019, 9, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Workman, A.D.; Palmer, J.N.; Adappa, N.D.; Cohen, N.A. The Role of Bitter and Sweet Taste Receptors in Upper Airway Immunity. Curr. Allergy Asthma Rep. 2015, 15, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekstedt, S.; Kumlien Georén, S.; Cardell, L.O. Effects of MP-AzeFlu enhanced by activation of bitter taste receptor TAS2R. Allergy Asthma Clin. Immunol. 2020, 16, 45. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.S.; Evans, C.T.; Tan, B.K.; Chandra, R.K.; Smith, S.B.; Kern, R.C. National burden of antibiotic use for adult rhinosinusitis. J. Allergy Clin. Immunol. 2013, 132, 1230–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnett, J.P.; Nguyen, T.T.; Moffatt, J.D.; Pelham, E.R.; Kalsi, K.K.; Baker, E.H.; Baines, D.L. Proinflammatory mediators disrupt glucose homeostasis in airway surface liquid. J. Immunol. 2012, 189, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Kalsi, K.K.; Baker, E.H.; Fraser, O.; Chung, Y.L.; Mace, O.J.; Tarelli, E.; Philips, B.J.; Baines, D.L. Glucose homeostasis across human airway epithelial cell monolayers: Role of diffusion, transport and metabolism. Pflugers Arch. 2009, 457, 1061–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzulo, A.A.; Gutiérrez, J.; Duschner, K.S.; McConnell, K.S.; Taft, P.J.; Ernst, S.E.; Yahr, T.L.; Rahmouni, K.; Klesney-Tait, J.; Stoltz, D.A.; et al. Glucose depletion in the airway surface liquid is essential for sterility of the airways. PLoS ONE 2011, 6, e16166. [Google Scholar] [CrossRef] [Green Version]
- Steelant, B.; Seys, S.F.; Van Gerven, L.; Van Woensel, M.; Farré, R.; Wawrzyniak, P.; Kortekaas Krohn, I.; Bullens, D.M.; Talavera, K.; Raap, U.; et al. Histamine and T helper cytokine-driven epithelial barrier dysfunction in allergic rhinitis. J. Allergy Clin. Immunol. 2018, 141, 951–963.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Civantos, A.M.; Maina, I.W.; Arnold, M.; Lin, C.; Stevens, E.M.; Tan, L.H.; Gleeson, P.K.; Colquitt, L.R.; Cowart, B.J.; Bosso, J.V.; et al. Denatonium benzoate bitter taste perception in chronic rhinosinusitis subgroups. Int. Forum. Allergy Rhinol. 2021, 11, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Civantos, A.M.; Arnold, M.; Stevens, E.M.; Cowart, B.J.; Colquitt, L.R.; Mansfield, C.; Kennedy, D.W.; Brooks, S.G.; Workman, A.D.; et al. Divergent bitter and sweet taste perception intensity in chronic rhinosinusitis patients. Int. Forum. Allergy Rhinol. 2021, 11, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Kook, J.H.; Kim, H.K.; Kim, H.J.; Kim, K.W.; Kim, T.H.; Kang, K.R.; Oh, D.J.; Lee, S.H. Increased expression of bitter taste receptors in human allergic nasal mucosa and their contribution to the shrinkage of human nasal mucosa. Clin. Exp. Allergy 2016, 46, 584–601. [Google Scholar] [CrossRef] [PubMed]
- Weichelt, U.; Cay, R.; Schmitz, T.; Strauss, E.; Sifringer, M.; Bührer, C.; Endesfelder, S. Prevention of hyperoxia-mediated pulmonary inflammation in neonatal rats by caffeine. Eur. Respir. J. 2013, 41, 966–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sainz, E.; Cavenagh, M.M.; Gutierrez, J.; Battey, J.F.; Northup, J.K.; Sullivan, S.L. Functional characterization of human bitter taste receptors. Biochem. J. 2007, 403, 537–543. [Google Scholar] [CrossRef]
- Bukowskyj, M.; Nakatsu, K. The bronchodilator effect of caffeine in adult asthmatics. Am. Rev. Respir Dis. 1987, 135, 173–175. [Google Scholar] [CrossRef] [PubMed]
- Gong, H., Jr.; Simmons, M.S.; Tashkin, D.P.; Hui, K.K.; Lee, E.Y. Bronchodilator effects of caffeine in coffee. A dose-response study of asthmatic subjects. Chest 1986, 89, 335–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kivity, S.; Ben Aharon, Y.; Man, A.; Topilsky, M. The effect of caffeine on exercise-induced bronchoconstriction. Chest 1990, 97, 1083–1085. [Google Scholar] [CrossRef] [PubMed]
- Welsh, E.J.; Bara, A.; Barley, E.; Cates, C.J. Caffeine for asthma. Cochrane Database Syst Rev. 2010, 2010, CD001112. [Google Scholar] [CrossRef] [Green Version]
- Miyatake, H.; Taki, F.; Taniguchi, H.; Suzuki, R.; Takagi, K.; Satake, T. Erythromycin reduces the severity of bronchial hyperresponsiveness in asthma. Chest 1991, 99, 670–673. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.B.; Simons, K.J.; Gillespie, C.A.; Simons, F.E. The bronchodilator effects and pharmacokinetics of caffeine in asthma. N. Engl. J. Med. 1984, 310, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Kreutzer, K.; Bassler, D. Caffeine for apnea of prematurity: A neonatal success story. Neonatology 2014, 105, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Jaggupilli, A.; Singh, N.; De Jesus, V.C.; Gounni, M.S.; Dhanaraj, P.; Chelikani, P. Chemosensory bitter taste receptors (T2Rs) are activated by multiple antibiotics. FASEB J. 2019, 33, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.G.; Yang, I.A.; Upham, J.W.; Reynolds, P.N.; Hodge, S.; James, A.L.; Jenkins, C.; Peters, M.J.; Marks, G.B.; Baraket, M.; et al. Effect of azithromycin on asthma exacerbations and quality of life in adults with persistent uncontrolled asthma (AMAZES): A randomised, double-blind, placebo-controlled trial. Lancet 2017, 390, 659–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaman, R.; Nowak, S.; Di Pizio, A.; Kitaneh, H.; Abu-Jaish, A.; Meyerhof, W.; Niv, M.Y.; Behrens, M. Probing the Binding Pocket of the Broadly Tuned Human Bitter Taste Receptor TAS2R14 by Chemical Modification of Cognate Agonists. Chem. Biol. Drug Des. 2016, 88, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Di Pizio, A.; Niv, M.Y. Promiscuity and selectivity of bitter molecules and their receptors. Bioorg. Med. Chem. 2015, 23, 4082–4091. [Google Scholar] [CrossRef] [PubMed]
- Di Pizio, A.; Levit, A.; Slutzki, M.; Behrens, M.; Karaman, R.; Niv, M.Y. Comparing Class A GPCRs to bitter taste receptors: Structural motifs, ligand interactions and agonist-to-antagonist ratios. Methods Cell Biol. 2016, 132, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Bachmanov, A.A.; Bosak, N.P.; Lin, C.; Matsumoto, I.; Ohmoto, M.; Reed, D.R.; Nelson, T.M. Genetics of taste receptors. Curr. Pharm. Des. 2014, 20, 2669–2683. [Google Scholar] [CrossRef] [Green Version]
- Mennella, J.A.; Pepino, M.Y.; Reed, D.R. Genetic and environmental determinants of bitter perception and sweet preferences. Pediatrics 2005, 115, e216–e222. [Google Scholar] [CrossRef] [Green Version]
- Fushan, A.A.; Simons, C.T.; Slack, J.P.; Manichaikul, A.; Drayna, D. Allelic polymorphism within the TAS1R3 promoter is associated with human taste sensitivity to sucrose. Curr. Biol. 2009, 19, 1288–1293. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Cell Type (Organ) | Taste Receptor | Process Regulated by Taste Receptor | Ref. |
|---|---|---|---|
| Oral cavity | |||
| Taste cells | T1R1, T1R2, T1R3 | Tasting sweet and umami nutrients | [1] |
| Taste cells | T2Rs | Tasting bitter nutrients | [1] |
| Taste cells | Sour and salty tastants | Modulation of taste cell’s function | [1] |
| Gastrointestinal tract | |||
| Enteroendocrine cells | T2Rs | Release of hormones as paracrine factors, neurotransmitters, and neuromodulators Motility of the gastrointestinal tract Innate immune response | [4,10,11,12,13,14] |
| Enteroendocrine cells | T1R and T1R3 | Glucose sensing Energy balancing | [5] |
| Enteroendocrine cells | Umami and other amino acid receptors | Modulation of protein digestion Modulation of exocrine and endocrine secretion Metabolism and nutrient utilization | [6,7,8,9] |
| Genitourinary system | |||
| Testis | T2R | Spermatogenesis | [13] |
| Bladder | T2R | Bladder contraction | [14] |
| Cardiovascular system | |||
| Cardiac monocytes | T2R and T1R | Nutrient sensors | [15] |
| Central Nervous system | |||
| Brain stem | T2R | Food intake | [19] |
| Brain cells | T2R | Food intake Other unknown physiological functions | [18] |
| parabrachial nucleus and horoid plexus | T2R | Assessment of cerebrospinal fluid | [20] |
| Upper airways | |||
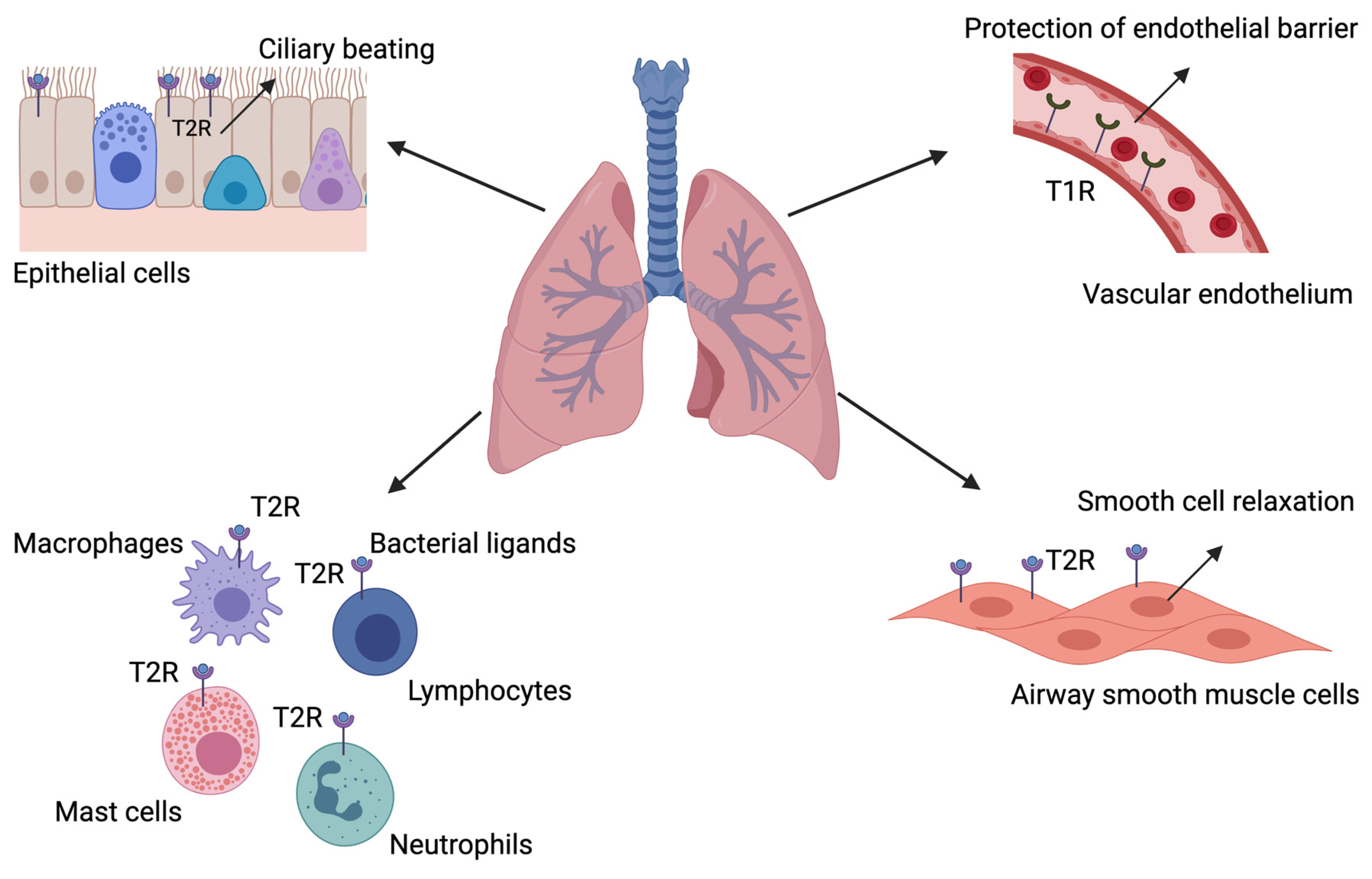
| Epithelial cells | T2R | Neurogenic inflammation Modulation of ciliary beat frequency Bactericidal clearance | [21,22,23] |
| Solitary chemosensory cells | T1R | Modulation of glucose levels Downregulation of T2R activation | [21,24] |
| Solitary chemosensory cells | T2R | Neurogenic inflammation Modulation of ciliary beat frequency Bactericidal clearance | [21,22,23,25,26,27,28,29,30,31] |
| Lower airways | |||
| Airway smooth muscle cells | T2R | Bronchial smooth muscle cell relaxation Ciliary beat frequency | [32,33,34,35,36,37] |
| Epithelial cells | T2R | Ciliary beat frequency | [38] |
| Immune cells | T2R | Recognition of microbial products and induction of immune response | [26,39,40,41,42,43,44] |
| Microvascular endothelial cells | T1R | Protection against edemagenic agent-induced barrier disruption | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martens, K.; Steelant, B.; Bullens, D.M.A. Taste Receptors: The Gatekeepers of the Airway Epithelium. Cells 2021, 10, 2889. https://doi.org/10.3390/cells10112889
Martens K, Steelant B, Bullens DMA. Taste Receptors: The Gatekeepers of the Airway Epithelium. Cells. 2021; 10(11):2889. https://doi.org/10.3390/cells10112889
Chicago/Turabian StyleMartens, Katleen, Brecht Steelant, and Dominique M. A. Bullens. 2021. "Taste Receptors: The Gatekeepers of the Airway Epithelium" Cells 10, no. 11: 2889. https://doi.org/10.3390/cells10112889
APA StyleMartens, K., Steelant, B., & Bullens, D. M. A. (2021). Taste Receptors: The Gatekeepers of the Airway Epithelium. Cells, 10(11), 2889. https://doi.org/10.3390/cells10112889