Interactions between Cellulose and (1,3;1,4)-β-glucans and Arabinoxylans in the Regenerating Wall of Suspension Culture Cells of the Ryegrass Lolium multiflorum

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Lolium SCC Growth and Maintenance
2.2. Isolation and Regeneration of Lolium SCC Protoplasts
2.3. Wall Labeling
2.4. Laser Scanning Microscopy (LSM) and Superresolution Microscopy
2.5. Transmission Electron Microscopy (TEM)
2.6. Scanning Electron Microscopy (SEM)
3. Results
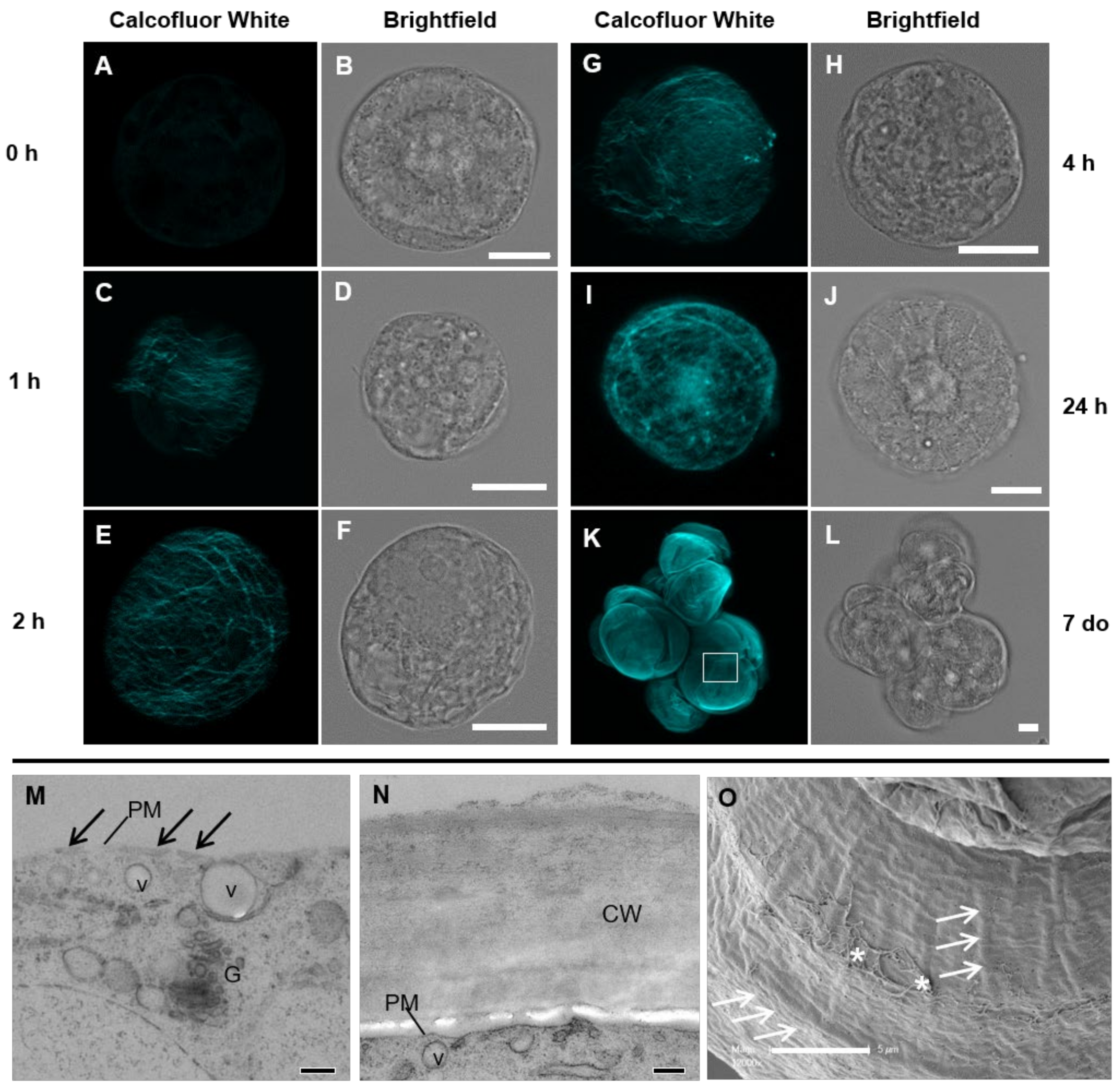
3.1. Spatial and Temporal Cell Wall Development
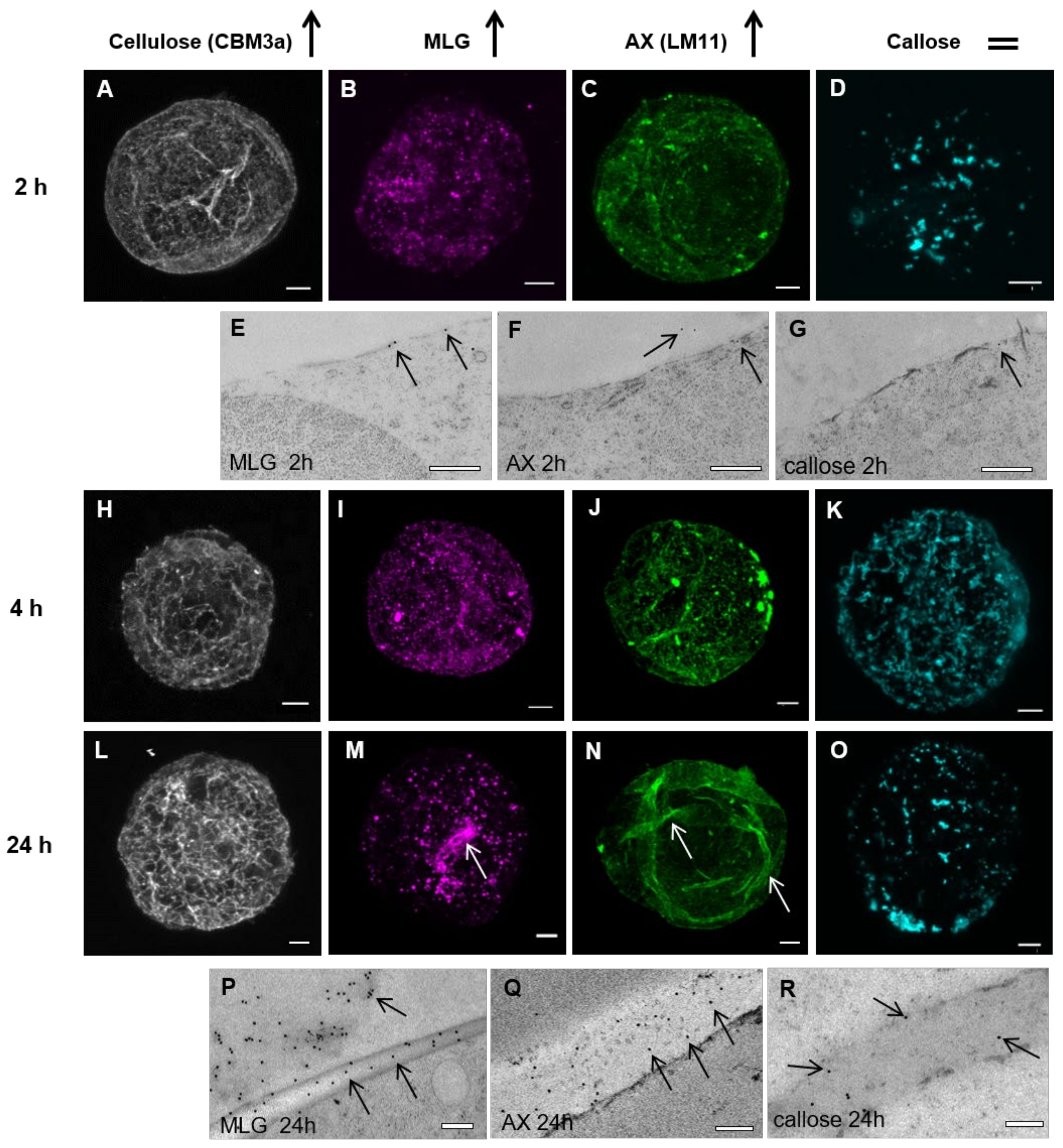
3.1.1. Spatial and Temporal Deposition of the Major Cell Wall Polysaccharides
3.1.2. Cell Wall Maturation
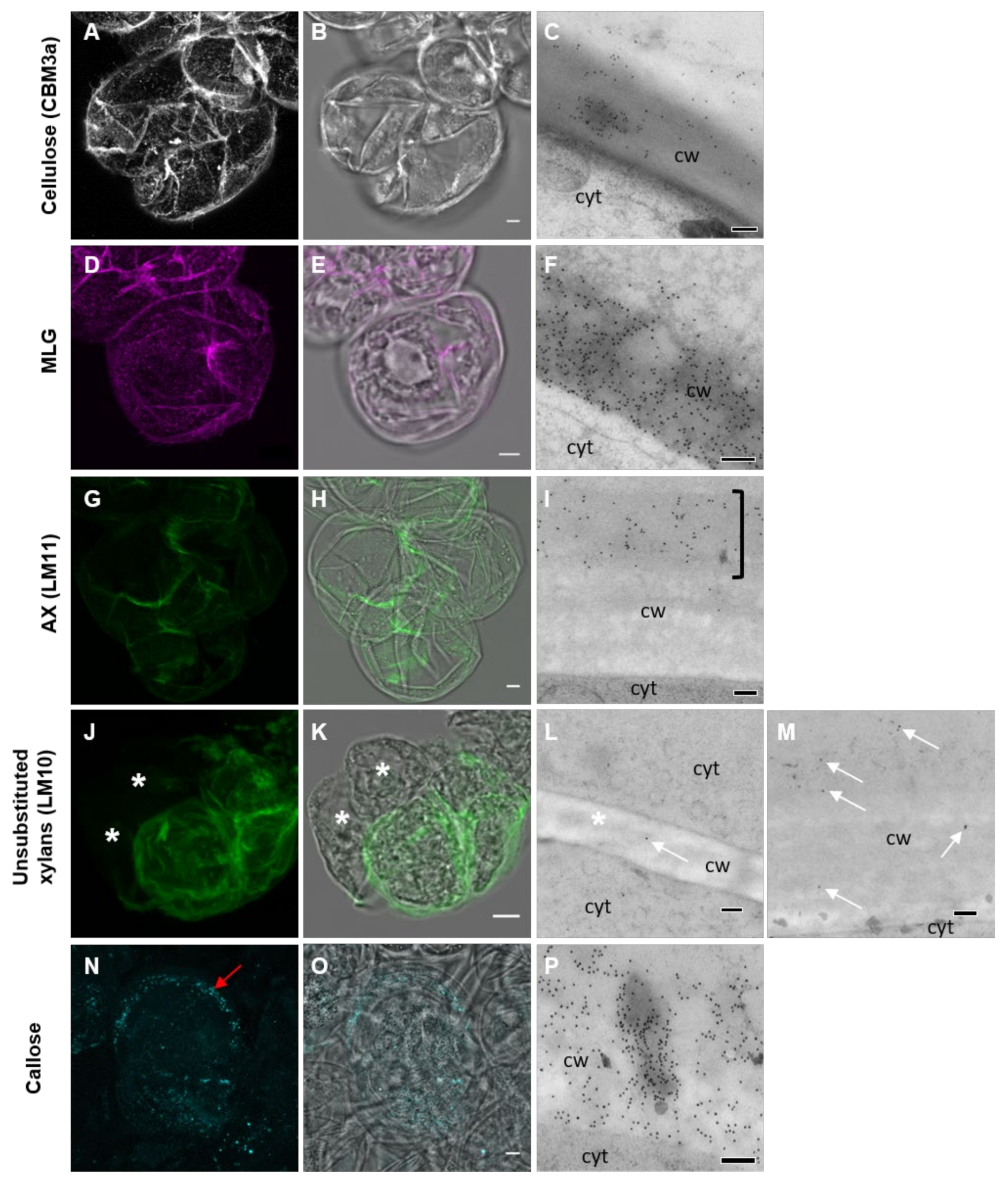
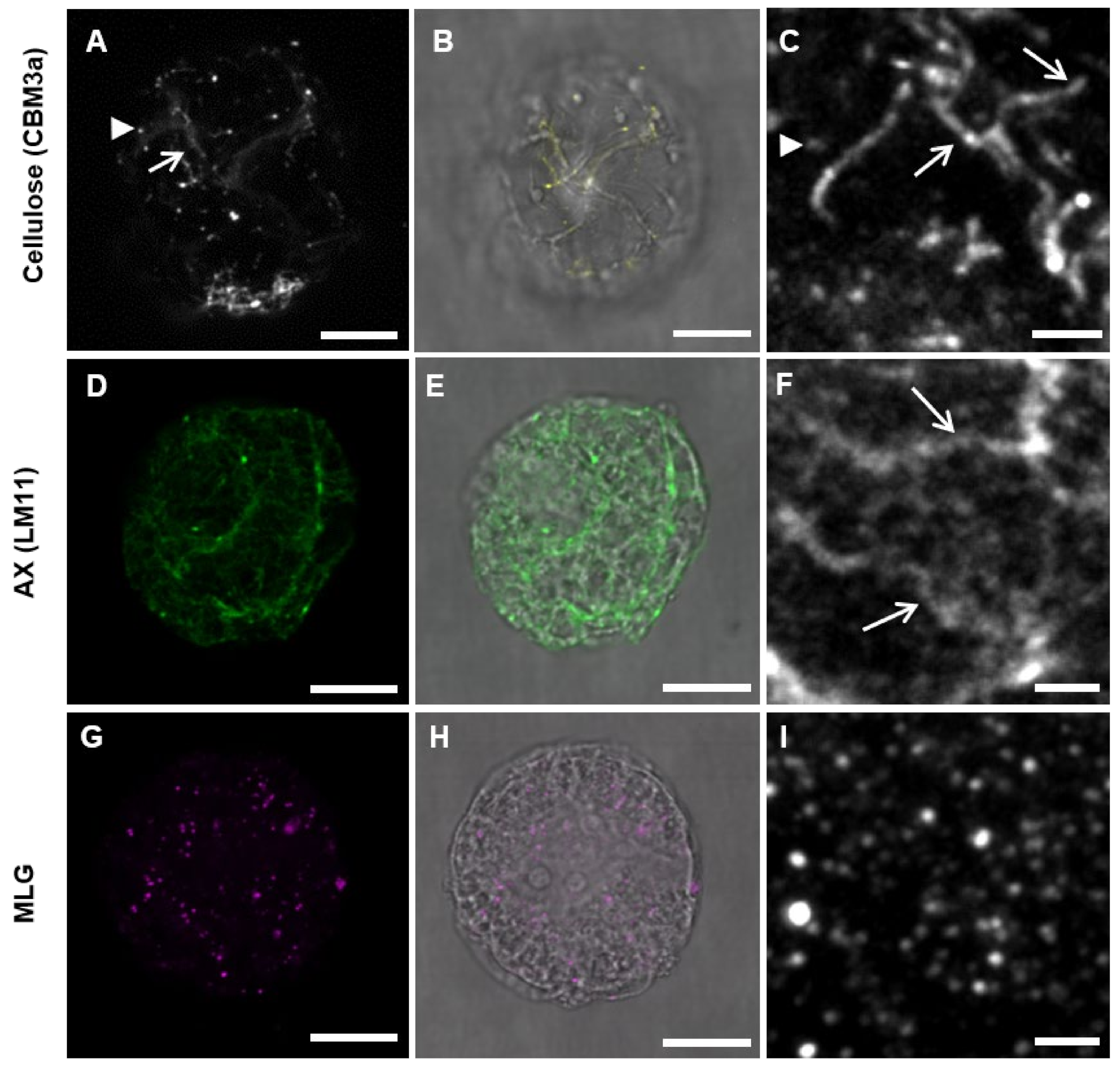
3.2. Pattern of Labeling of Cellulose, MLG, and AX in the Cell Wall
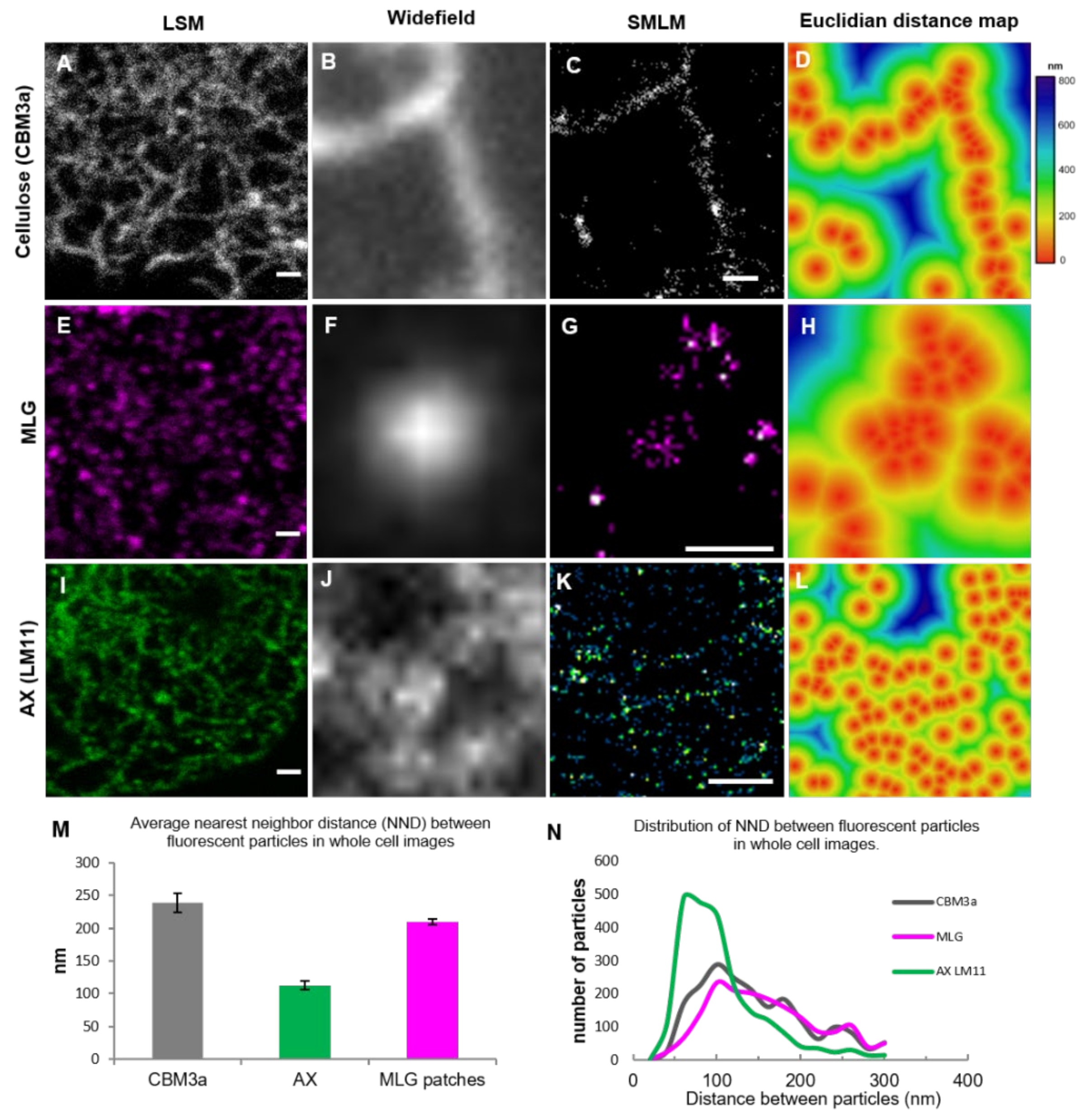
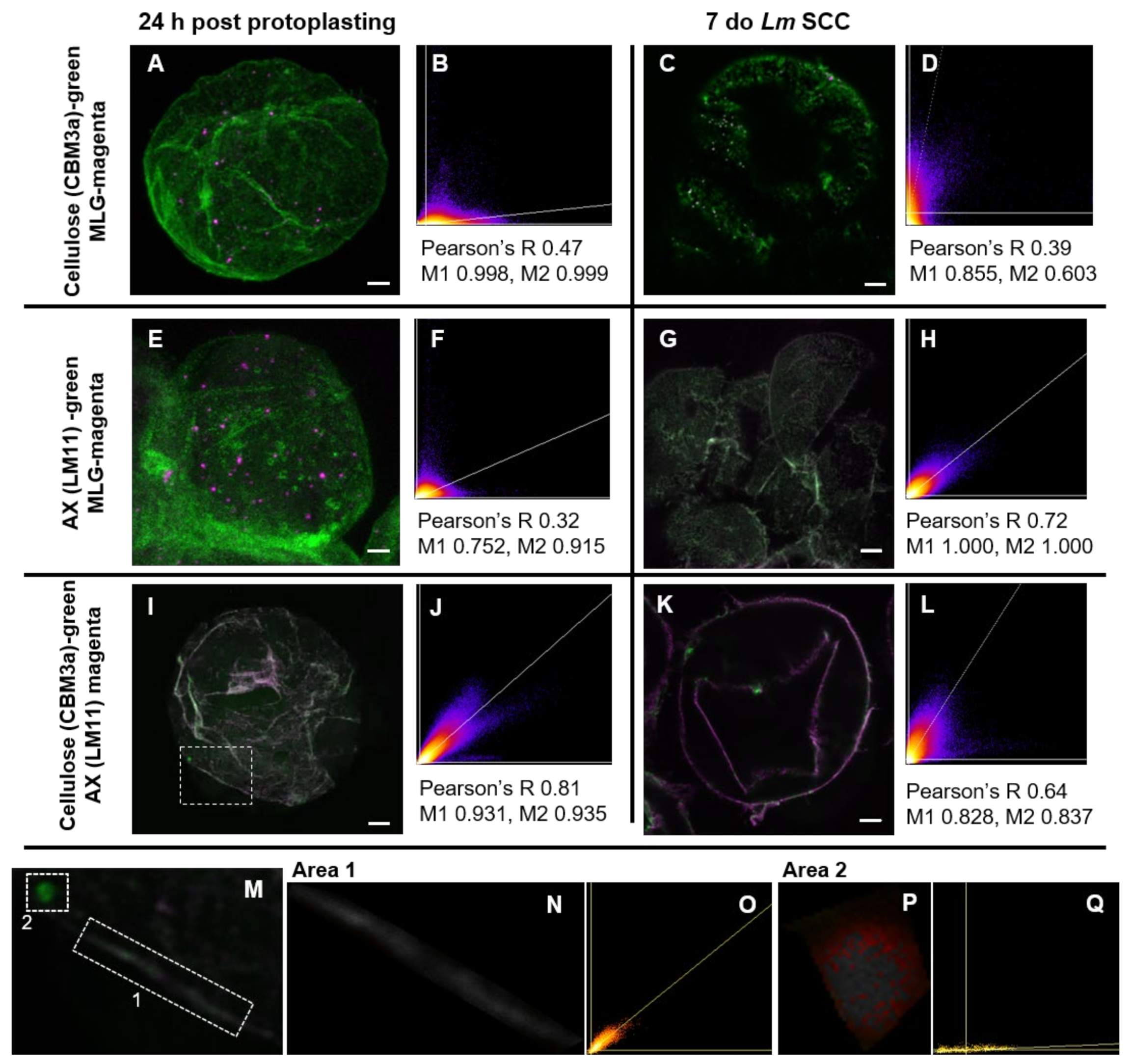
3.3. Association between Polysaccharides in the Developing Cell Wall
3.4. Wall Architecture after the Disruption of Cellulose
4. Discussion
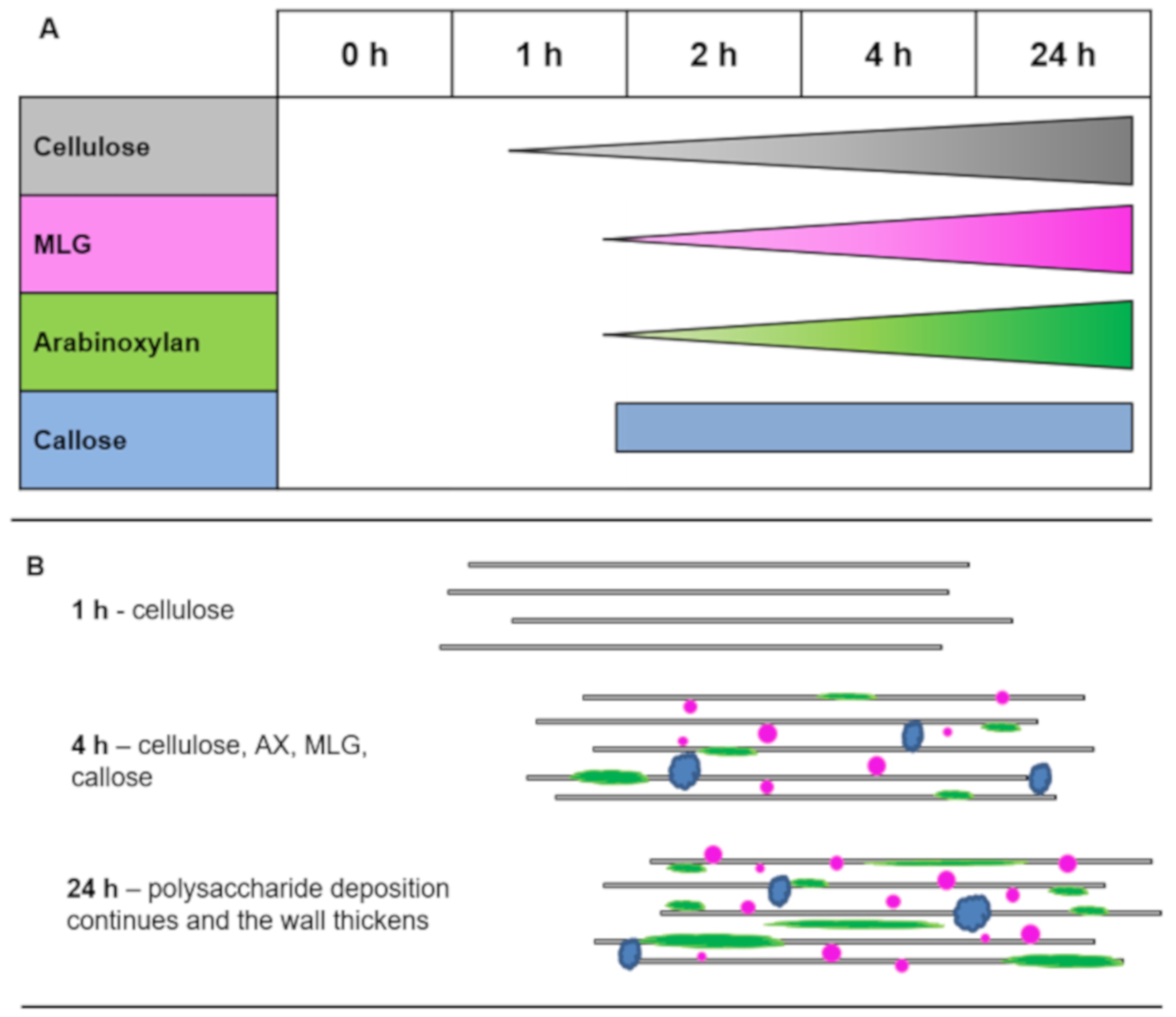
4.1. Timing of Polysaccharide Deposition
4.2. Callose Is Not Deposited First in the Regenerating Cell Wall
4.3. Cell Wall Architecture and Heterogeneity
4.4. Physico-Chemical Heterogeneity of the Cell Wall
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doblin, M.S.; Pettolino, F.; Bacic, A. Plant cell walls: The skeleton of the plant world. Func. Plant Biol. 2010, 37, 357–381. [Google Scholar] [CrossRef]
- Bacic, A.; Harris, P.J.; Stone, B.A. Structure and function of plant cell walls. In The Biochemistry of Plants; Priess, J., Ed.; Academic Press: New York, NY, USA, 1988; pp. 297–371. [Google Scholar]
- Carpita, N.C.; Gibeaut, D.M. Structural models of primary cell walls in flowering plants: Consistency of molecular structure with the physical properties of the walls during growth. Plant J. 1993, 3, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Fincher, G.B.; Stone, B.A. Cell walls and their components in cereal grain technology. In Encyclopedia of Plant Physiology. New Series, Plant. Carbohydrates II; Tanner, W.W., Loewus, F.A., Eds.; Springer: Berlin, Germany, 1986; pp. 68–132. [Google Scholar]
- McCann, M.; Roberts, K. Changes in cell wall architecture during cell elongation. J. Exp. Bot. 1994, 45, 1683–1691. [Google Scholar] [CrossRef]
- Smith, B.G.; Harris, P.J. The polysacchride composition of Poales cell walls: Poaceae cell walls are not unique. Biochem. Syst. Ecol. 1999, 27, 33–53. [Google Scholar] [CrossRef]
- Yokoyama, R.; Nishitani, K. Genomic basis for cell-wall diversity in plants. A comparative approach to gene families in rice and Arabidopsis. Plant Cell Phys. 2004, 45, 1111–1121. [Google Scholar] [CrossRef]
- York, W.S.; O’Neill, M.A. Biochemical control of xylan biosyntheis—Which end is up? Curr. Opt. Plant Biol. 2008, 11, 258–265. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.; Bush, M.; Milioni, D.; Sado, P.; Stacey, N.J.; Catchpole, G.; Defernez, M.; Carpita, N.C.; Hofte, H.; Ulvskov, P.; et al. Approaches to understanding the functional architecture of the plant cell wall. Phytochem 2001, 57, 811–821. [Google Scholar] [CrossRef]
- Tucker, M.R.; Lou, H.; Aubert, M.K.; Wilkinson, L.G.; Little, A.; Houston, K.; Pinto, S.C.; Shirley, N.J. Exploring the role of cell wall-related genes and polysaccharides during plant development. Plants 2018, 7, 42. [Google Scholar] [CrossRef]
- Velickovic, D.; Ropartz, D.; Guillon, F.; Saulnier, L.; Rogniaux, H. New insights into the structural and spaital variability of cell wall polysaccharides during wheat grain development as revealed through MALDI mass spectrometry imaging. J. Exp. Bot. 2014, 65, 2079–2091. [Google Scholar] [CrossRef]
- Burton, R.A.; Gidley, M.J.; Fincher, G.B. Heterogeneity in the chemistry, structure and function of plant cell walls. Nat. Chem. Biol. 2010, 6, 724–732. [Google Scholar] [CrossRef]
- McCann, M.; Chen, L.; Roberts, K.; Kemsley, E.K.; Sene, C.; Carpita, N.C.; Stacey, N.J.; Wilson, R.H. Infrared microspectroscopy: Sampling heterogeneity in plant cell wall composition and architecture. Phys. Plant 1997, 100, 729–738. [Google Scholar] [CrossRef]
- van de Meene, A.M.L.; Doblin, M.S.; Bacic, A. The plant secretory pathway seen through the lens of the cell wall. Protoplasma 2017, 254, 75–94. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Diffuse Growth of Plant Cell Walls. Plant Phys. 2018, 176, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Yakubov, G.E.; Bonilla, M.R.; Chen, H.; Doblin, M.S.; Bacic, A.; Gidley, M.J.; Stokes, J.R. Mapping nano-scale mechanical heterogeneity of primary plant cell walls. J. Exp. Bot. 2016, 67, 2799–2816. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zheng, Y.; Cosgrove, D.J. Spatial organization of cellulose microfibrils and matrix polysaccharides in primary plant cell walls as imaged by multichannel atomic force microscopy. Plant J. 2016, 85, 179–192. [Google Scholar] [CrossRef]
- Levesque-Tremblay, G.; Pelloux, J.; Braybrook, S.A.; Muller, K. Tuning of pectin methylesterification: Consequences for cell wall biomechanics and development. Planta 2015, 242, 791–811. [Google Scholar] [CrossRef]
- Nakamura, A.; Furuta, H.; Maeda, H.; Takao, T.; Naganatsu, Y. Structural studies by stepwise enzymatic degradation of the main backbone of soybean soluble polysaccharides consisting of galacturonan and rhamnogalacturonan. Biosci. Biotechnol. Biochem. 2002, 66, 1301–1313. [Google Scholar] [CrossRef]
- Tan, L.; Eberhard, S.; Pattathil, S.; Warder, C.; Glushka, J.; Yuan, C.; Hao, Z.; Zhu, X.; Avci, U.; Miller, J.S.; et al. An Arabidopsis cell wall proteoglycan consists of pectin and arabinoxylan covalently linked to an arabinogalactan protein. Plant Cell 2013, 25, 270–287. [Google Scholar] [CrossRef]
- Willats, W.G.T.; McCartney, L.; Mackie, W.; Knox, J.P. Pectin: Cell biology and prospects for functional analysis. Plant Mol. Biol. 2001, 47, 9–27. [Google Scholar] [CrossRef]
- De Lorenzo, G.; Ferrari, S.; Giovannoni, M.; Mattei, B.; Cervone, F. Cell wall traits that influence plant development, immunity, and bioconversion. Plant J. 2019, 97, 134–147. [Google Scholar] [CrossRef]
- Wu, H.-C.; Bulgakov, V.P.; Jinn, T.-L. Pectin Methylesterases: Cell wall remodeling proteins are required for plant response to heat stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef] [PubMed]
- Atalla, R.H.; Hackney, J.M.; Uhlin, I.; Thompson, N.S. Hemicelluloses as structure regulators in the aggregation of native cellulose. Int. J. Biol. Macromol. 1993, 15, 109–112. [Google Scholar] [CrossRef]
- Ebringerová, A.; Hromádková, Z.; Heinze, T. Hemicellulose. In Polysaccharides I; Heinze, T., Ed.; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Gunness, P.; Michiels, J.; Vanhaecke, L.; de Smet, S.; Kravchuk, O.; van de Meene, A.M.L.; Gidley, M.J. Reduction in circulating bile acid and restricted diffusion across the intestinal epithelium are associated with a decrease in blood cholesterol in the presence of oat β-glucan. FASEB J. 2016, 30, 4227–4238. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.X.; Walker, K.Z.; Muir, J.G.; O’Dea, K. Arabinoxylan fibre improves metabolic control in people with Type II diabetes. Eur. J. Clin. Nut. 2004, 58, 621–628. [Google Scholar] [CrossRef]
- Smith, M.M.; Stone, B.A. Chemical composition of the cell walls of Lolium multiflorum endosperm. Phytochemistry 1973, 12, 1361–1367. [Google Scholar] [CrossRef]
- Moscatiello, R.; Baldan, B.; Navazio, L. Plant cell suspension cultures. Methods Mol. Biol. 2013, 953, 77–93. [Google Scholar]
- Kuki, H.; Higaki, T.; Yokoyama, R.; Kuroha, T.; Shinohara, N.; Hasezawa, S.; Nishitani, K. Quantitative confocal imaging method for analyzing cellulose dynamics during cell wall regeneration in Arabidopsis mesophyll protoplasts. Plant Direct. 2017, 1, e00021. [Google Scholar] [CrossRef]
- Rico, H.; Carrillo, C.; Aguado, C.; Mormeneo, S.; Sentandreu, R. Initial steps of wall protoplast regeneration in Candida albicans. Res. Microbiol. 1997, 148, 593–603. [Google Scholar] [CrossRef]
- van Amstel, T.N.M.; Kengen, H.M.P. Callose deposition in the primary wall of suspension cells and regenerating protoplasts, and its relationship to patterned cellulose synthesis. Can. J. Bot. 1996, 74, 1040–1049. [Google Scholar] [CrossRef]
- Yang, X.; Tu, L.; Zhu, L.; Fu, L.; Min, L.; Zhang, X. Expression profile analysis of genes involved in cell wall regeneration during protoplast culture in cotton by suppression subtractive hybridization and macroarray. J. Exp. Bot. 2008, 59, 3661–3674. [Google Scholar] [CrossRef]
- Smith, M.M.; Stone, B.A. Studies on Lolium multiflorum endosperm in tissue culture I. Nutrition. Aust. J. Biol. Sci. 1973, 26, 123–134. [Google Scholar] [CrossRef]
- Wilson, S.M.; Bacic, A. Preparation of plant cells for transmission electron microscopy to optimize immunogold labeling of carbohydrate and protein epitopes. Nat. Prot. 2012, 7, 1716–1727. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.M.; Ho, Y.Y.; Lampugnani, E.R.; van de Meene, A.M.L.; Bain, M.P.; Bacic, A.; Doblin, M.S. Determining the subcellular location of synthesis and assembly of the cell wall polysaccharide (1,3;1,4)-β-D-glucan in grasses. Plant Cell 2015, 27, 754–771. [Google Scholar] [CrossRef] [PubMed]
- Nicovich, P.R.; Owen, D.M.; Gaus, K. Turning single-molecule localization microscopy into a quantitative bioanalytical tool. Nat. Prot. 2017, 12, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Halls, M.L.; Yeatman, H.R.; Nowell, C.J.; Thompson, G.L.; Gondin, A.B.; Civcirstov, S.; Bunnett, N.W.; Lambert, N.A.; Poole, D.P.; Canals, M. Plasma membrane localisation of the m-opioid receptor controls spatiotemporal signalling. Sci. Sig. 2016, 9, ra16. [Google Scholar] [CrossRef]
- Danielsson, P.-E. Euclidean distance mapping. Comp. Graph. Image Proc. 1980, 14, 227–248. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganada-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. FIJI: An open-source platform for biological-image analysis. Nat. Meth. 2012, 9, 676–682. [Google Scholar] [CrossRef]
- McCartney, L.; Steele-King, C.G.; Jordan, E.; Knox, J.P. Cell wall pectic (1-4)-β-D-galactan marks the acceleration ofcell elongation in the Arabidopsis seedling root meristem. Plant J. 2003, 33, 447–454. [Google Scholar] [CrossRef]
- Blake, A.W.; McCartney, L.; Flint, J.E.; Bolam, D.N.; Boraston, A.B.; Gilbert, H.J.; Knox, J.P. Understanding the biological rationale for the diversity of cellulose-directed carbohydrate-binding modules in prokaryotic enzymes. J. Biol. Chem. 2006, 2006, 29321–29329. [Google Scholar] [CrossRef]
- Meikle, P.J.; Hoogenraad, N.J.; Bonig, I.; Clarke, A.E.; Stone, B.A. A (1-3;1-4)-β-glucan specific monoclonal antibody and its use in the quantitation and immunocytochemical location of (1-3;1-4)-β-glucans. Plant J. 1994, 51, 1–9. [Google Scholar] [CrossRef]
- McCartney, L.; Marcus, S.E.; Knox, J.P. Monoclonal antibodies to plant cell wall xylans and arabinoxylans. J. Histo. Cytochem. 2005, 53, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Meikle, P.J.; Bonig, I.; Hoogenraad, N.J.; Clarke, A.E.; Stone, B.A. The location of (1→3)-β-glucans in the walls of pollen tubes of Nicotiana alata using a (1→3)-β-glucan-specific monoclonal antibody. Planta 1991, 185, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Samuels, L.A.; Giddings, T.H.J.; Staehelin, L.A. Cytokinesis in tobacco BY-2 and root tip cells: A new model of cell plate formation in higher plants. J. Cell Biol. 1995, 130, 1345–1357. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-Y.; Kim, J.-Y. Callose synthesis in higher plants. Plant Sig. Behav. 2009, 4, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.K.; Lipka, V.; Burton, R.A.; Panstruga, R.; Strizhov, N.; Schulze-Lefert, P.; Fincher, G.B. An Arabidopsis callose synthase, GSL5, is required for wound and papillary callose formation. Plant Cell 2003, 15, 2503–2513. [Google Scholar] [CrossRef]
- Hernandez-Gomez, M.C.; Rydahl, M.G.; Rogowski, A.; Morland, C.; Cartmell, A.; Crouch, L.; Labourel, A.; Fontes, C.M.G.A.; Willats, W.G.T.; Gilbert, H.J.; et al. Recognition of xyloglucan by the crystalline cellulose-binding site of a family 3a carbohydrate-binding module. FEBS Lett. 2015, 589, 2297–2303. [Google Scholar] [CrossRef]
- Marcus, S.E.; Verhertbruggen, Y.; Herve, C.; Ordaz-Ortiz, J.J.; Farkas, V.; Pedersen, H.L.; Willats, W.G.T.; Knox, J.P. Pectic homogalacturonan masks abundant sets of xyloglucan epitopes in plant cell walls. BMC Plant Biol. 2008, 8, 60. [Google Scholar] [CrossRef]
- Pettolino, F.; Hoogenraad, N.J.; Ferguson, C.; Bacic, A.; Johnson, E.; Stone, B.A. A (1-4)β-mannan-specific monoclonal antibody and its use in the immunocytochemical location of galactomannans. Planta 2001, 214, 235–242. [Google Scholar] [CrossRef]
- Knox, J.P.; Linstead, P.J.; King, J.; Cooper, C.; Roberts, K. Pectin esterification is spatially regulated both within cell walls and between developing tissues of root apices. Planta 1990, 181, 512–521. [Google Scholar] [CrossRef]
- Pennell, R.I.; Knox, J.P.; Scofield, G.N.; Selvendran, R.; Roberts, K. A family of abundant plasma membrane-associated glycoproteins related to the arabinogalactan proteins is unique to flowering plants. J. Cell Biol. 1989, 108, 1967–1977. [Google Scholar] [CrossRef]
- Harris, P.J.; Hartley, R.D. Detection of bound ferulic acid in cell-walls of Gramineae by ultraviolet fluorescence microscopy. Nature 1976, 259, 508–510. [Google Scholar] [CrossRef]
- Ruprecht, C.; Bartetzko, M.P.; Senf, D.; Dallabernadina, P.; Books, I.; Andersen, M.C.F.; Kotake, T.; Knox, J.P.; Hahn, M.G.; Clausen, M.H.; et al. A synthetic glycan microarray enables epitope mapping of plant cell wall glycan-directed antibodies. Plant Phys. 2017, 175, 1094–1104. [Google Scholar] [CrossRef]
- Desprez, T.; Vernhettes, S.; Fagard, M.; Refregier, G.; Desnos, T.; Aletti, E.; Py, N.; Pelletier, S.; Hofte, H. Resistance against herbicide isoxaben cellulose deficiency caused by distinct mutations in same cellulose synthase isoform CESA6. Plant Phys. 2002, 128, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Scheible, W.R.; Eshed, R.; Richmond, T.; Delmber, D.; Somerville, C. Modifications of cellulose synthase confer resistance to isoxaben and thiazolidinone herbicides in Arabidopsis ixr1 mutants. Proc. Natl. Acad. Sci. USA 2001, 98, 10079–10084. [Google Scholar] [CrossRef] [PubMed]
- Nebenfuhr, A.; Gallagher, L.A.; Dunahay, T.G.; Frohlick, J.A.; Mazurkiewicz, A.M.; Meehl, J.B.; Staehelin, L.A. Stop-and-go movements of plant Golgi stacks are mediated by the acto-myosin system. Plant Phys. 1999, 121, 1127–1141. [Google Scholar] [CrossRef]
- Chevalier, L.; Bernard, S.; Ramdani, Y.; Lamour, R.; Bardor, M.; Lerouge, P.; Follet-Gueye, M.-L.; Driouich, A. Subcompartment localization of the side chain xyloglucan-synthesizing enzymes within Golgi stacks of tobacco suspension-cultured cells. Plant J. 2010, 64, 977–989. [Google Scholar] [CrossRef]
- Kim, S.-J.; Zemelis, S.; Keegstra, K.; Brandizzi, F. The cytoplasmic localization of the catalytic site of CSLF6 supports a channeling model for the biosynthesis of mixed-linkage glucan. Plant J. 2015, 81, 537–547. [Google Scholar] [CrossRef]
- McFarlane, H.E.; Doring, A.; Persson, S. The cell biology of cellulose synthesis. Ann. Rev. Plant Biol. 2014, 65, 69–94. [Google Scholar] [CrossRef]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Ann. Rev. Plant Biol. 2010, 263–289. [Google Scholar] [CrossRef]
- Zhang, G.F.; Staehelin, L.A. Functional compartmentation of the Golgi apparatus of plant cells. Plant Phys. 1992, 99, 1070–1083. [Google Scholar] [CrossRef]
- Malinsky, J.; Opekarova, M.; Grossman, G.; Tanner, W. Membrane microdomains, rafts and detergent-resistant membranes in plants and fungi. Ann. Rev. Plant Biol. 2013, 64, 501–529. [Google Scholar] [CrossRef] [PubMed]
- Martiniere, A.; Lavagi, I.; Nageswaran, G.G.; Rolfe, D.J.; Maneta-Peyret, L.; Luu, D.-T.; Botchway, S.W.; Webb, S.E.D.; Mongrand, S.; Maurel, C.; et al. Cell wall constrains lateral diffusion of plant plasma-membrane proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 12805–12810. [Google Scholar] [CrossRef]
- Shim, I.; Law, R.; Kileeg, Z.; Stronghill, P.; Northey, J.G.B.; Strap, J.L.; Bonetta, D.T. Alleles causing resistance to isoxaben and flupoxam highlight the significance of transmembrane domains for CESA protein function. Front. Plant Sci. 2018, 9, 1152. [Google Scholar]
- Tateno, M.; Brabham, C.; DeBolt, S. Cellulose biosynthesis inhibitors—A multifunctional toolbox. J. Exp. Bot. 2016, 67, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Fugelstad, J.; Bulone, V. What Do We Really Know about Cellulose Biosynthesis in Higher Plants? J. Integ. Plant Biol. 2010, 52, 161–175. [Google Scholar] [CrossRef]
- Shedletzky, E.; Shmuel, M.; Delmer, D.; Lamport, D.T.A. Adaptation and Growth of Tomato Cells on the Herbicide 2,6-Dichlorobenzonitrile Leads to Production of Unique Cell Walls Virtually Lacking a Cellulose-Xyloglucan Network. Plant Phys. 1990, 94, 980–987. [Google Scholar] [CrossRef]
- Langenaeken, N.A.; Ieven, P.; Hedlund, E.G.; Kyomugasho, C.; van de Walle, D.; Dewettinck, K.; van Loey, A.M.; Roeffaers, M.B.J.; Courtin, C.M. Arabinoxylan, b-glucan and pectin in barley and malt endosperm cell walls: A microstructure study using CLSM and cryo-SEM. Plant J. 2020, 103, 1477–1489. [Google Scholar] [CrossRef]
- Guillon, F.; Tranquet, O.; Quillien, L.; Utille, J.-P.; Ortiz, J.J.O.; Sualnier, L. Generation of polyclonal and monoclonal antibodies against arabinoxylans and their use for immunocytochemical location of arabinoxylans in cell walls of endosperm of wheat. J. Cereal Sci. 2004, 40, 167–182. [Google Scholar] [CrossRef]
- Brown, R.C.; Lemmon, B.E.; Stone, B.A.; Olsen, O.A. Cell wall (1-3)- and (1-3;1-4)-β-glucans during early grain development in rice (Oryza sativa L). Planta 1997, 202, 414–426. [Google Scholar] [CrossRef]
- Philippe, S.; Saulnier, L.; Guillon, F. Arabinoxylan and (1-3;1-4)-β-glucan deposition in cell walls during wheat endosperm development. Planta 2006, 224, 449–461. [Google Scholar] [CrossRef]
- Wilson, S.M.; Burton, R.A.; Collins, H.M.; Doblin, M.S.; Pettolino, F.A.; Shirley, N.; Fincher, G.B.; Bacic, A. Pattern of deposition of cell wall polysaccharides and transcript abundance of related cell wall synthesis genes during differentiation in barley endosperm. Plant Phys. 2012, 159, 655–670. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.M.; Burton, R.A.; Doblin, M.S.; Stone, B.A.; Newbigin, E.J.; Fincher, G.B.; Bacic, A. Temporal and spatial appearance of wall polysaccharides during cellularization of barley (Hordeum vulgare) endosperm. Planta 2006, 224, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Beeckman, T.; Przemeck, G.K.H.; Stamatiou, G.; Lau, R.; Terryn, N.; de Rycke, R.; Inze, D.; Berleth, T. Genetic complexity of Cellulose Synthase A gene function in Arabidopsis embryogenesis. Plant Phys. 2002, 130, 1883–1893. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-Y.; Liu, L.; Lee, E.; Han, X.; Rim, Y.; Chu, H.; Kim, S.-W.; Sack, F.; Kim, J.-Y. The Arabidopsis callose synthase gene GSL8 is required for cytokinesis and cell patterning. Plant Phys. 2009, 150, 105–113. [Google Scholar] [CrossRef]
- Nakashima, J.; Laosinchai, W.; Cui, X.; Brown, R.M.J. New insight into the mechanism of cellulose and callose biosynthesis: Proteases may regulate callose biosynthesis upon wounding. Cellulose 2003, 10, 369–389. [Google Scholar] [CrossRef]
- Anderson, C.T.; Kieber, J.J. Dynamic construction, perception, and remodeling of plant cell walls. Ann. Rev. Plant Biol. 2020, 71, 39–69. [Google Scholar] [CrossRef]
- Barnes, W.J.; Anderson, C.T. Release, recycle, rebuild: Cell-wall remodeling, autodegradation, and sugar salvage for new wall biosynthesis during plant development. Mol. Plant 2018, 11, 31–46. [Google Scholar] [CrossRef]
- Grantham, N.J.; Wurman-Rodrich, J.; Terrett, O.M.; Lyczakowski, J.J.; Stott, K.; Iuga, D.U.; Simmons, T.J.; Durand-Tardif, M.; Brown, S.P.; Duree, R.; et al. An even pattern of xylan substitution is critical for interaction with cellulose in plant cell walls. Nat. Plants 2017, 3, 859–865. [Google Scholar] [CrossRef]
- Mikkelsen, D.; Flanagan, B.M.; Wilson, S.M.; Bacic, A.; Gidley, M.J. Interactions of arabinoxylan and (1,3)(1,4)-β-glucan with cellulose networks. Biomacromolecules 2015, 16, 1232–1239. [Google Scholar] [CrossRef]
- Simmons, T.J.; Mortimer, J.C.; Bernardinellli, O.D.; Poppler, A.-C.; Brown, S.P.; deAzevedo, E.R.; Dupree, R.; Dupree, P. Folding of xylan onto cellulose fibrils in plant cell walls revealed by solid-state NMR. Nat. Comm. 2016, 7, 13902. [Google Scholar] [CrossRef]
- Daher, F.B.; Chen, Y.; Bozorg, B.; Clough, J.; Jonsson, H.; Braybrook, S.A. Anisotropic growth is achieved through the additive mechanical effect of material anisotropy and elastic asymmetry. eLIFE 2018, 7, e38161. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Wall Component | Name of Stain/Antibody | Specific Epitope | Source | Ref | Working Dilution |
|---|---|---|---|---|---|
| General cell wall stain | Calcofluor White | (1,3)-, (1,3;1,4)-, and (1,4)-β-glucans | Sigma | [41] | 0.01% |
| Cellulose | CBM3a | crystalline cellulose | Plant Probes | [42] | 1:50 |
| Mixed-linkage glucan (MLG) | (1-3;1-4)-β-glucan-directed monoclonal antibody | linear (1,3;1,4)-β-oligo-saccharide segments in (1,3;1,4)-β-glucans | Biosupplies | [43] | 1:500 |
| Callose | (1-3)-β-glucan-directed monoclonal antibody | linear (1,3)-β-oligosaccharide segments in (1,3)-β-glucan | Biosupplies | [45] | 1:300 |
| Xylan | LM10 | (1→4)-β-D-xylan | Plant Probes | [44] | 1:20 |
| Arabinoxylan | LM11 | (1→4)-β-D-xylan/arabinoxylan | Plant Probes | [44] | 1:20 |
| Xyloglucan | LM15 | XXXG motif of xyloglucans | Plant Probes | [50] | 1:20 |
| Mannan | (1-4)-β-mannan-directed monoclonal antibody | (1,4)-β-manno-oligosaccharides in (1,4)-β-mannans and galactomannans | Biosupplies | [51] | 1:300 |
| Pectin | JIM 7 | homogalacturonan partially Me-HG, general pectin | Plant Probes | [52] | 1:20 |
| Arabinogalactan protein (AGP) | MAC207 | AGP glycan | CCRC | [53] | 1:10 |
| Ferulic acid | NH4OH bathychromatic shift | Ferulic acid | Sigma | [54] | 0.1 M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van de Meene, A.; McAloney, L.; Wilson, S.M.; Zhou, J.; Zeng, W.; McMillan, P.; Bacic, A.; Doblin, M.S. Interactions between Cellulose and (1,3;1,4)-β-glucans and Arabinoxylans in the Regenerating Wall of Suspension Culture Cells of the Ryegrass Lolium multiflorum. Cells 2021, 10, 127. https://doi.org/10.3390/cells10010127
van de Meene A, McAloney L, Wilson SM, Zhou J, Zeng W, McMillan P, Bacic A, Doblin MS. Interactions between Cellulose and (1,3;1,4)-β-glucans and Arabinoxylans in the Regenerating Wall of Suspension Culture Cells of the Ryegrass Lolium multiflorum. Cells. 2021; 10(1):127. https://doi.org/10.3390/cells10010127
Chicago/Turabian Stylevan de Meene, Allison, Lauren McAloney, Sarah M. Wilson, JiZhi Zhou, Wei Zeng, Paul McMillan, Antony Bacic, and Monika S. Doblin. 2021. "Interactions between Cellulose and (1,3;1,4)-β-glucans and Arabinoxylans in the Regenerating Wall of Suspension Culture Cells of the Ryegrass Lolium multiflorum" Cells 10, no. 1: 127. https://doi.org/10.3390/cells10010127
APA Stylevan de Meene, A., McAloney, L., Wilson, S. M., Zhou, J., Zeng, W., McMillan, P., Bacic, A., & Doblin, M. S. (2021). Interactions between Cellulose and (1,3;1,4)-β-glucans and Arabinoxylans in the Regenerating Wall of Suspension Culture Cells of the Ryegrass Lolium multiflorum. Cells, 10(1), 127. https://doi.org/10.3390/cells10010127