Genetic Diversification and Selection Strategies for Improving Sorghum Grain Yield Under Phosphorous-Deficient Conditions in West Africa

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
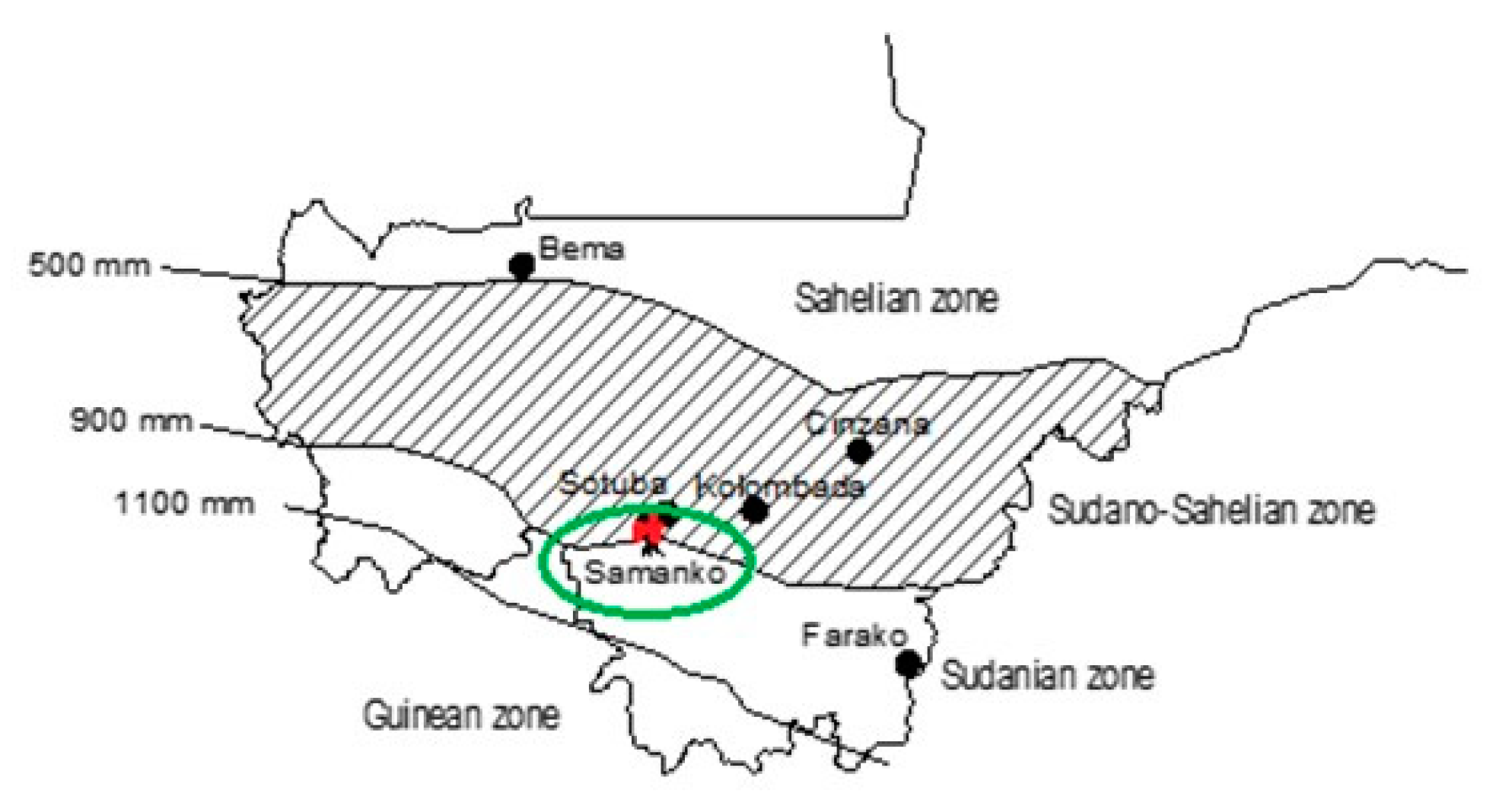
2.1. Plant Materials
2.2. Phenotyping
2.3. Statistical Analysis
2.3.1. Individual Trial Analysis
2.3.2. Combined Analysis
3. Results
3.1. Performance for Grain Yield and Related Traits Uunder LP and HP Field Conditions
3.2. Genetic Parameters
3.2.1. Repeatability, Heritability, and Genetic Variance Estimates
3.2.2. Predicted Responses to Direct and Indirect Selection for Grain Yield Under P-Limited Conditions
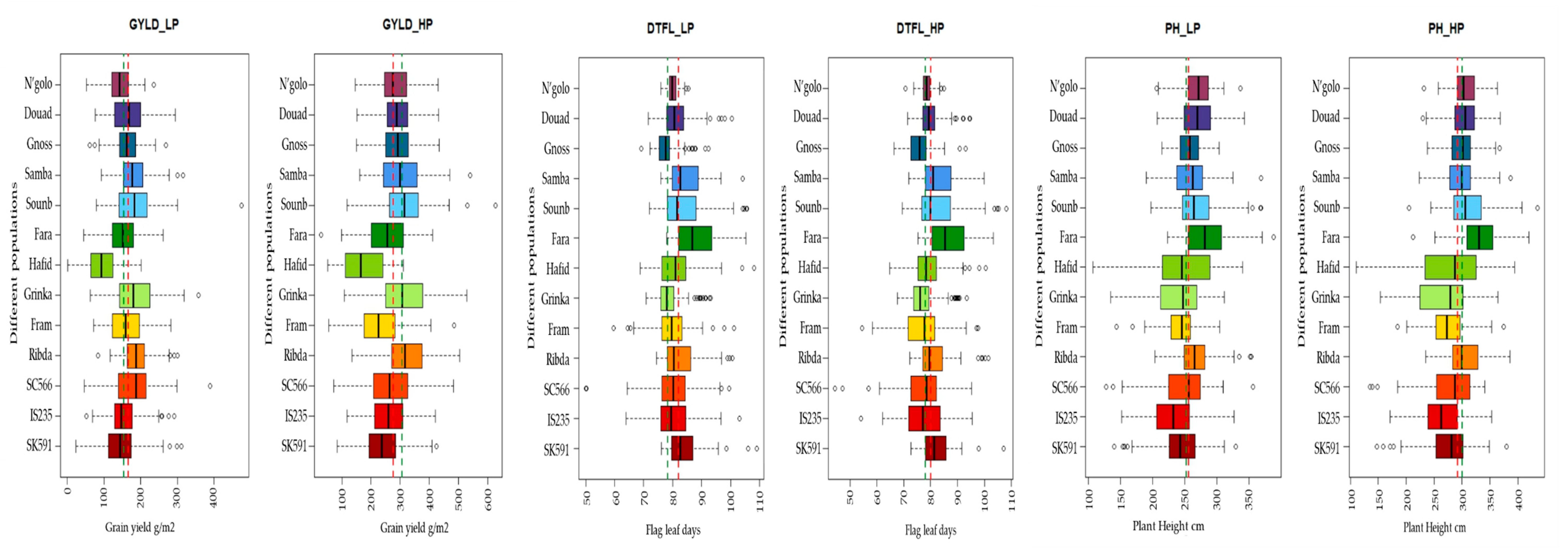
3.3. Performance of Specific Populations
4. Discussion
4.1. Genetic Parameters
4.2. Usefulness of BCNAM Populations
5. Implications and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. FAOSTAT. 2017. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 17 December 2017).
- Breman, H. Amélioration de la fertilité des sols en Afrique de l’Ouest: Contraintes et perspectives. In Soil Fertility Management in West African Land Use Systems; Renard, G., Neef, A., Becker, K., Oppen, M.V., Eds.; Margraf Verlag: Weikersheim, Germany, 1998; pp. 7–20. [Google Scholar]
- Rattunde, H.F.W. Farmer Participatory Early-Generation Yield Testing of Sorghum in West Africa: Possibilities to Optimize Genetic Gains for Yield in Farmers’ Fields. Crop Sci. 2016, 56, 2493. [Google Scholar] [CrossRef]
- vom Brocke, K.; Trouche, G.; Weltzien, E.; Barro-Kondombo, C.P.; Gozé, E.; Chantereau, J. Participatory variety development for sorghum in Burkina Faso: Farmers’ selection and farmers’ criteria. Field Crops Res. 2010, 119, 183–194. [Google Scholar] [CrossRef]
- Buerkert, A.; Bationo, A.; Piepho, H.-P. Efficient phosphorus application strategies for increased crop production in sub-Saharan West Africa. Field Crops Res. 2001, 72, 1–15. [Google Scholar] [CrossRef]
- Leiser, W.L. Selection Strategy for Sorghum Targeting Phosphorus-limited Environments in West Africa: Analysis of Multi-environment Experiments. Crop Sci. 2012, 52, 2517. [Google Scholar] [CrossRef]
- Weltzien, E.; Rattunde, H.F.W.; van Mourik, T.A.; Ajeigbe, H.A. Sorghum cultivation and improvement in West and Central Africa. In Burleigh Dodds Series in Agricultural Science; Rooney, W., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; pp. 217–240. [Google Scholar]
- Leiser, W.L. Phosphorous Efficiency and Tolerance Traits for Selection of Sorghum for Performance in Phosphorous-Limited Environments. Crop Sci. 2015, 55, 1152. [Google Scholar] [CrossRef]
- Leiser, W.L.; Weltzien-Rattunde, H.F.; Weltzien-Rattunde, E.; Haussmann, B.I.G. Sorghum tolerance to low-phosphorus soil conditions. In Burleigh Dodds Series in Agricultural Science; Texas A&M University, USA, Rooney, W., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; pp. 247–272. [Google Scholar]
- Rattunde, H.F.W. Population Improvement of Pearl Millet and Sorghum: Current Research, Impact and Issues for Implementation. In Proceedings of the International Conference on Genetic Improvement of Sorghum and Pearl Millet, Holiday Inn Plaza, Lubbock, TX, USA, 22–27 September 1996; p. 188. [Google Scholar]
- Morris, G.P. Population genomic and genome-wide association studies of agroclimatic traits in sorghum. Proc. Natl. Acad. Sci. USA 2016, 110, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Gore, M.; Buckler, E.S.; Yu, J. Status and Prospects of Association Mapping in Plants. Plant Genome J. 2008, 1, 5. [Google Scholar] [CrossRef]
- Jordan, D.R.; Mace, E.S.; Cruickshank, A.W.; Hunt, C.H.; Henzell, R.G. Exploring and Exploiting Genetic Variation from Unadapted Sorghum Germplasm in a Breeding Program. Crop Sci. 2011, 51, 1444. [Google Scholar] [CrossRef]
- Mace, E.S.; Jordan, D.R. Integrating sorghum whole genome sequence information with a compendium of sorghum QTL studies reveals uneven distribution of QTL and of gene-rich regions with significant implications for crop improvement. Theor. Appl. Genet. 2011, 123, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Kante, M. Sorghum Hybrids for Low-Input Farming Systems in West Africa: Quantitative Genetic Parameters to Guide Hybrid Breeding. Crop Sci. 2019, 59, 1–18. [Google Scholar] [CrossRef]
- Weltzien, E. Enhancing farmers’ access to sorghum varieties through scaling-up participatory plant breeding in Mali, West Africa. In Bringing Farmers Back into Breeding. Experiences with Participatory Plant Breding and Challenges for Institutionalisation; Almekinders, C., Hardon, J., Eds.; Agromisa Foundation: Wageningen, The Netherlands, 2006; pp. 58–69. [Google Scholar]
- Weltzien, E.; Vom Brocke, K.; Touré, A.; Rattunde, F.; Chantereau, J. Revue et Tendances pour la Recherche en Sélection Participative en Afrique de l’Ouest [Review and outlook for participatory plant breeding research in West Africa]. Cah. Agric. 2008, 17, 165–171. [Google Scholar]
- Piepho, H.-P.; Mohring, J. Computing Heritability and Selection Response From Unbalanced Plant Breeding Trials. Genetics 2007, 177, 1881–1888. [Google Scholar] [CrossRef] [PubMed]
- Atlin, G.N.; Frey, K.J. Predicting the relative effectiveness of direct versus indirect selection for oat yield in three types of stress environments. Euphytica 1989, 44, 137–142. [Google Scholar] [CrossRef]
- Chen, J.; Xu, L.; Cai, Y.; Xu, J. QTL mapping of phosphorus efficiency and relative biologic characteristics in maize (Zea mays L.) at two sites. Plant Soil 2008, 313, 251–266. [Google Scholar] [CrossRef]
- Cichy, K.A.; Snapp, S.S.; Blair, M.W. Plant growth habit, root architecture traits and tolerance to low soil phosphorus in an Andean bean population. Euphytica 2009, 165, 257–268. [Google Scholar] [CrossRef]
- Parentoni, S. Inheritance and breeding strategies for phosphorous efficiency in tropical maize (Zea mays L). Maydica 2010, 55, 1. [Google Scholar]
- Rossiter, R.C. Phosphorus deficiency and flowering in subterranean clover (T. subterraneum L.). Ann. Bot. 1978, 42, 325–329. [Google Scholar] [CrossRef]
- Wissuwa, M.; Ae, N. Further characterization of two QTLs that increase phosphorus uptake of rice (Oryza sativa L.) under phosphorus deficiency. Plant Soil 2001, 237, 275–286. [Google Scholar] [CrossRef]
- Gelli, M. Mapping QTLs and association of differentially expressed gene transcripts for multiple agronomic traits under different nitrogen levels in sorghum. BMC Plant Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Kante, M. Can Tall Guinea-Race Sorghum Hybrids Deliver Yield Advantage to Smallholder Farmers in West and Central Africa? Crop Sci. 2017, 57, 833. [Google Scholar] [CrossRef]
- Rattunde, H.F.W. Yield of Photoperiod-sensitive Sorghum Hybrids Based on Guinea-race Germplasm under Farmers’ Field Conditions in Mali. Crop Sci. 2013, 53, 2454. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor | Race | Pop Names | Origin | Progenies | Specific Advantages |
|---|---|---|---|---|---|
| N’golofing | Guinea (Guineense) | N’golo | Mali | 80 | Grain and panicle traits |
| Douadjè | Guinea (Guineense) | Douad | Mali | 80 | Low-P adaptation and also allele for Al tolerance. |
| Gnossiconi | Guinea (Guineense) | Gnoss | Burkina Faso | 71 | Grain and panicle traits |
| Sambalma | Guinea (Conspicuum) | Samba | Nigeria | 102 | Grain and panicle traits, Al tolerance |
| IS15401 | Guinea (Conspicuum) | Soumb | Cameroon | 101 | Striga, aluminum, low-P, and midge tolerance/resistance |
| Fara-Fara | Guinea (Conspicuum) | Fara | Nigeria | 80 | Geographic and intra-racial diversity |
| IS23645 | Guinea (Margaritiferum) | Hafid | Gambia | 75 | Intra-racial diversity and vitreous grain |
| Grinkan | Caudatum-Guinea | Grinka | Mali | 100 | Productivity, Stover quality |
| Framida | Caudatum | Fram | Burkina Faso | 80 | Striga tolerance |
| Ribdahu | Caudatum | Ribda | Nigeria | 80 | Midge resistance, racial diversity |
| SC566-14 | Caudatum | SC566 | Brazil | 80 | Aluminum tolerance |
| IS 23540 | Caudatum | IS235 | Ethiopia | 80 | Sweet stem |
| SK 5912 | Durra-Caudatum | SK591 | Nigeria | 80 | Geographic and racial diversity |
| Traits | Abbreviation | Units | Method |
|---|---|---|---|
| Seedling vigor | GV | Score (1–9) | Visual score of seedling growth 35 d after sowing from lowest (1) to highest (9) |
| Date to flag leaf appearance | DTFL | Julian day | Number of days from sowing to flag leaf appearance |
| Plant height | PH | cm | Length of the main stalk recorded on 3 random plants from the base of the stalk to the tip of the panicle (maturity). |
| Panicle length | PANL | cm | Length from the first whorl of panicle branches to the tip of the rachis (maturity) |
| Grain yield/m2 | GYLD | g m−2 | Threshed grain weight per unit area(maturity) |
| Weight of 100 grains | HGW | g | Weight of 100 grains with grain humidity less than or equal to 12% |
| Traits | HP | LP | Minimum | Maximum | Mean | Repeatability | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| σ²G | s.e. | σ²G | s.e. | LP | HP | LP | HP | LP | HP | LP | HP | |
| GYLD | 4918 *** | 313 | 1801 *** | 135 | 20 | 40 | 509 | 703 | 166 | 277 | 0.60 | 0.69 |
| DTFL | 56.5 *** | 2.6 | 50.9 *** | 2.3 | 49.5 | 46.5 | 129 | 133 | 82.0 | 80.1 | 0.95 | 0.94 |
| PH | 1801.4 *** | 87.7 | 1185.4 *** | 65.6 | 103 | 115 | 420 | 485 | 256 | 292 | 0.79 | 0.88 |
| HGW | 0.081 *** | 0.004 | 0.091 *** | 0.004 | 0.49 | 0.60 | 3.19 | 3.20 | 2.15 | 2.31 | 0.90 | 0.91 |
| PANL | 11.44 *** | 0.63 | 9.76 *** | 0.57 | 16.0 | 13.5 | 45.5 | 50.0 | 30.0 | 29.3 | 0.76 | 0.79 |
| GV | 0.17 *** | 0.03 | 0.07 *** | 0.02 | 2 | 2 | 8 | 9 | 5.17 | 5.92 | 0.03 | 0.00 |
| Traits | Combined | ||||
|---|---|---|---|---|---|
| σ²G | s.e. | σ²G × P | s.e. | h2 | |
| GYLD | 2460 *** | 171.00 | 932 *** | 118.00 | 0.67 |
| DTFL | 52.63 *** | 2.34 | 1.18 *** | 0.19 | 0.96 |
| PH | 1463.2 *** | 69.60 | 39 *** | 15.60 | 0.91 |
| HGW | 0.08 *** | 0.00 | 0.01 *** | 0.00 | 0.91 |
| PANL | 10.12 *** | 0.51 | 0.48 *** | 0.18 | 0.86 |
| GV | 0.10 *** | 0.01 | 0.02 ns | 0.02 | 0.35 |
| Env | Sources of Variation | σ² | s.e. | h2 |
|---|---|---|---|---|
| HP | Genotype | 6319 *** | 1126 | 0.59 |
| Genotype × Year | 11,570 *** | 1218 | ||
| LP | Genotype | 1866 *** | 431 | 0.57 |
| Genotype × Year | 3726 *** | 478 | ||
| Combined | Genotype | 3997 *** | 661 | 0.79 |
| Genotype × P | 1202 *** | 370 | ||
| Genotype × Year | 4515 *** | 608 | ||
| Genotype × P × Year | 2223 *** | 481 |
| Traits | Correlation (r) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DTFL_HP | DTFL_LP | GV_HP | GV_LP | GYLD_HP | GYLD_LP | PANL_HP | PANL_LP | PH_HP | PH_LP | HGW_HP | HGW_LP | |
| DTFL_HP | ||||||||||||
| DTFL_LP | 0.91 *** | |||||||||||
| GV_HP | −0.16 *** | −0.09 ** | ||||||||||
| GV_LP | −0.11 *** | −0.25 *** | 0.20 *** | |||||||||
| GYLD_HP | −0.23 *** | −0.25 *** | 0.44 *** | 0.24 *** | ||||||||
| GYLD_LP | −0.03 ns | −0.13 *** | 0.12 *** | 0.37 *** | 0.54 *** | |||||||
| PANL_HP | 0.05 ns | 0.03 ns | 0.09 ** | 0.06 ns | 0.23 *** | 0.06 ns | ||||||
| PANL_LP | 0.19 *** | 0.14 *** | 0.04 ns | 0.05 ns | 0.12 *** | 0.10 ** | 0.73 *** | |||||
| PH_HP | 0.29 *** | 0.28 *** | 0.34 *** | 0.12 *** | 0.31 *** | 0.15 *** | 0.44 *** | 0.37 *** | ||||
| PH_LP | 0.27 *** | 0.19 *** | 0.19 *** | 0.32 *** | 0.22 *** | 0.27 *** | 0.36 *** | 0.43 *** | 0.82 *** | |||
| HGW_HP | −0.13 *** | −0.14 *** | 0.21 *** | 0.12 | 0.30 *** | 0.25 *** | 0.06 ns | −0.03 ns | 0.13 *** | 0.11 *** | ||
| HGW_LP | −0.05 ns | −0.07 * | 0.14 *** | 0.15 | 0.20 *** | 0.37 *** | 0.07 * | 0.01 ns | 0.18 *** | 0.21 *** | 0.83 *** | |
| Population | 2013 | 2014 | 2015 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LP | HP | # Progenies Selected | LP | HP | LP | HP | |||||||
| Overall | Top Quartile | Overall | Top Quartile | Overall | Top Quartile | Overall | Top Quartile | Overall | Top Quartile | Overall | Top Quartile | ||
| N’golo | 143 | 186 | 284 | 358 | 26 | 128 | 159 | 300 | 385 | 109 | 148 | 209 | 258 |
| Douad | 165 | 227 | 292 | 369 | 29 | 131 | 158 | 280 | 350 | 102 | 149 | 205 | 221 |
| Gnoss | 164 | 212 | 291 | 369 | 16 | 103 | 126 | 292 | 361 | 96 | 126 | 191 | 245 |
| Samba | 184 | 240 | 304 | 399 | 39 | 105 | 146 | 220 | 324 | 85 | 133 | 191 | 255 |
| Soumb | 186 | 260 | 315 | 421 | 36 | 129 | 170 | 286 | 378 | 109 | 152 | 225 | 276 |
| Fara | 152 | 202 | 257 | 352 | 11 | 114 | 143 | 251 | 324 | 85 | 115 | 194 | 240 |
| Hafid | 100 | 150 | 176 | 264 | 0 | - | - | - | - | - | - | - | |
| Grinka | 186 | 268 | 308 | 426 | 47 | 123 | 171 | 304 | 401 | 104 | 159 | 214 | 289 |
| Fram | 162 | 230 | 236 | 343 | 15 | 116 | 149 | 332 | 458 | 88 | 123 | 170 | 218 |
| Ribda | 190 | 241 | 318 | 419 | 32 | 121 | 159 | 311 | 416 | 92 | 132 | 209 | 277 |
| SC566 | 179 | 249 | 263 | 375 | 22 | 122 | 159 | 243 | 328 | 106 | 141 | 181 | 217 |
| IS235 | 155 | 215 | 261 | 350 | 12 | 132 | 191 | 319 | 427 | 79 | 117 | 199 | 259 |
| SK591 | 151 | 226 | 241 | 332 | 13 | 105 | 131 | 252 | 347 | 101 | 141 | 176 | 213 |
| Lata3 | 155 | 308 | 114 | 375 | 106 | 202 | |||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diallo, C.; Rattunde, H.F.W.; Gracen, V.; Touré, A.; Nebié, B.; Leiser, W.; Dzidzienyo, D.K.; Sissoko, I.; Danquah, E.Y.; Diallo, A.G.; et al. Genetic Diversification and Selection Strategies for Improving Sorghum Grain Yield Under Phosphorous-Deficient Conditions in West Africa. Agronomy 2019, 9, 742. https://doi.org/10.3390/agronomy9110742
Diallo C, Rattunde HFW, Gracen V, Touré A, Nebié B, Leiser W, Dzidzienyo DK, Sissoko I, Danquah EY, Diallo AG, et al. Genetic Diversification and Selection Strategies for Improving Sorghum Grain Yield Under Phosphorous-Deficient Conditions in West Africa. Agronomy. 2019; 9(11):742. https://doi.org/10.3390/agronomy9110742
Chicago/Turabian StyleDiallo, Chiaka, H. Frederick W. Rattunde, Vernon Gracen, Aboubacar Touré, Baloua Nebié, Willmar Leiser, Daniel K. Dzidzienyo, Ibrahima Sissoko, Eric Y. Danquah, Abdoulaye G. Diallo, and et al. 2019. "Genetic Diversification and Selection Strategies for Improving Sorghum Grain Yield Under Phosphorous-Deficient Conditions in West Africa" Agronomy 9, no. 11: 742. https://doi.org/10.3390/agronomy9110742
APA StyleDiallo, C., Rattunde, H. F. W., Gracen, V., Touré, A., Nebié, B., Leiser, W., Dzidzienyo, D. K., Sissoko, I., Danquah, E. Y., Diallo, A. G., Sidibé, B., Sidibé, M., & Weltzien, E. (2019). Genetic Diversification and Selection Strategies for Improving Sorghum Grain Yield Under Phosphorous-Deficient Conditions in West Africa. Agronomy, 9(11), 742. https://doi.org/10.3390/agronomy9110742