
Response of Typical Artificial Forest Soil Microbial Community to Revegetation in the Loess Plateau, China


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. Soil Sample Determination
2.2.1. Sequencing of Soil Microbial Genes
2.2.2. Soil Microbial Carbon and Nitrogen Determination
2.2.3. Determination of Soil Physical and Chemical Properties
2.3. Data Processing and Analysis
2.3.1. Basic Data Processing
2.3.2. Operational Taxonomic Units (OTU)Analysis
2.3.3. Alpha Diversity Analysis and Rarefaction Curve
2.3.4. Statistical Analysis
3. Results
3.1. Analysis of Soil Microbial Carbon and Nitrogen
3.1.1. Characteristics of Soil Microbial Carbon and Nitrogen
3.1.2. Regression Analysis of Soil Microbial Carbon and Nitrogen Contents and Soil Carbon and Nitrogen Contents
3.2. Soil Bacterial Community Diversity
3.3. Soil Bacterial Community Structure
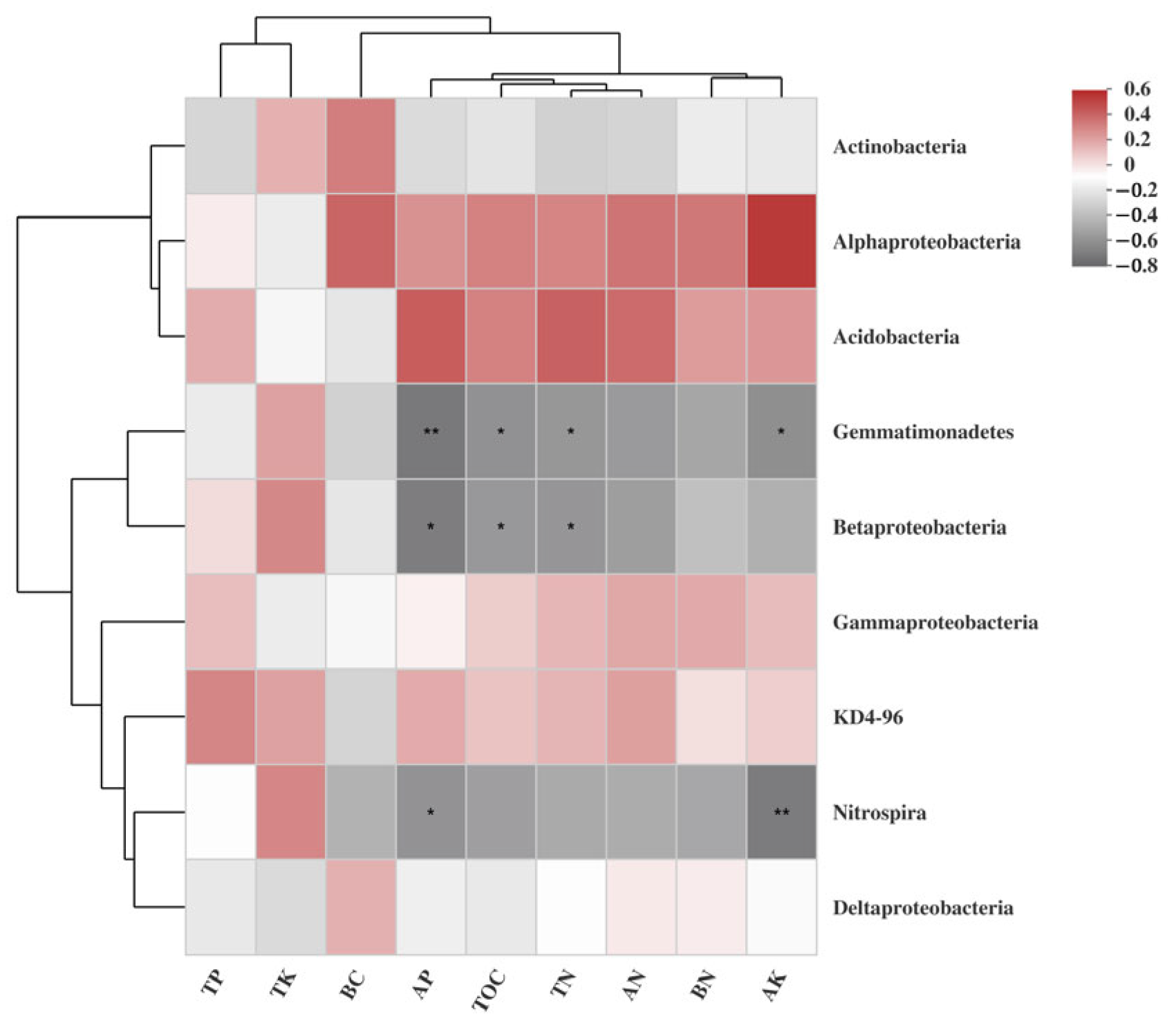
3.4. Relationships Among Soil Properties Factor and Bacterial Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, S.J.; Li, B.Y.; Gao, D.X.; Fu, S.Y.; Lu, Y.F.; Xu, M.P.; Ren, C.J.; Han, X.H. Response of Ecoenzymatic Stoichiometry to Soil Physicochemical Properties after Afforestation on Loess Hilly Region. Eurasian Soil Sci. 2020, 53, 1669–1675. [Google Scholar] [CrossRef]
- Sun, L.; Li, J.; Wang, Q.; Zhang, Y.; Xu, Z.; Wang, R.; Wang, X.; Jia, G.; Zhang, X. The effects of eight years of conservation tillage on the soil physicochemical properties and bacterial communities in a rain-fed agroecosystem of the loess plateau, China. Land Degrad. Dev. 2020, 31, 2475–2489. [Google Scholar] [CrossRef]
- Liu, S.; Ng, G.C. The role of spatiotemporal plant trait variability in model predictions of ecohydrological responses to climate change in a desert shrubland. J. Hydrol. 2020, 588, 125088. [Google Scholar] [CrossRef]
- Ali, A.; Dai, D.; Akhtar, K.; Teng, M.; Yan, Z.; Urbina-Cardona, N.; Mullerova, J.; Zhou, Z. Response of understory vegetation, tree regeneration, and soil quality to manipulated stand density in a Pinus massoniana plantation. Glob. Ecol. Conserv. 2019, 20, e775. [Google Scholar] [CrossRef]
- Xu, M.; Jian, J.; Wang, J.; Zhang, Z.; Yang, G.; Han, X.; Ren, C. Response of root nutrient resorption strategies to rhizosphere soil microbial nutrient utilization along Robinia pseudoacacia plantation chronosequence. For. Ecol. Manag. 2021, 489, 119053. [Google Scholar] [CrossRef]
- Islam, M.R.; Singh, B.; Dijkstra, F.A. Microbial carbon use efficiency of glucose varies with soil clay content: A meta-analysis. Appl. Soil Ecol. 2023, 181, 104636. [Google Scholar] [CrossRef]
- Leewis, M.; Lawrence, C.R.; Schulz, M.S.; Tfaily, M.M.; Ayala-Ortiz, C.O.; Flores, G.E.; Mackelprang, R.; McFarland, J.W. The influence of soil development on the depth distribution and structure of soil microbial communities. Soil Biol. Biochem. 2022, 174, 108808. [Google Scholar] [CrossRef]
- Guo, Z.; Huang, X.; Wang, T.; Su, J.; Li, S. Altitudinal influences on soil microbial diversity and community assembly in topsoil and subsoil layers: Insights from the jinsha river basin, southwest China. Forests 2024, 15, 1746. [Google Scholar] [CrossRef]
- Seaton, F.M.; Griffiths, R.I.; Goodall, T.; Lebron, I.; Norton, L.R. Soil bacterial and fungal communities show within field heterogeneity that varies by land management and distance metric. Soil Biol. Biochem. 2023, 177, 108920. [Google Scholar] [CrossRef]
- Pan, J.; Wu, H.; Xiang, W.; Ouyang, S.; Chen, L.; Zeng, Y.; Deng, X.; Zhao, Z.; Zeng, W.; Kuzyakov, Y. Soil microbial richness and community composition are primarily mediated by functional trait diversity of fine roots in subtropical forests. Plant Soil 2024, 497, 485–501. [Google Scholar] [CrossRef]
- You, N.; Dong, J.; Xiao, T.; Liu, J.; Xiao, X. The effects of the “Grain for Green” project on gross primary productivity in the Loess Plateau. Sci. Geogr. Sin. 2020, 40, 315–323. [Google Scholar] [CrossRef]
- Shan, L.; Xu, B. Discussion on some issues about returning farmland to forest or grassland on Loess Plateau in new era. Bull. Soil Water Conserv. 2019, 39, 295–297. [Google Scholar] [CrossRef]
- Wu, D.; Zou, C.; Cao, W.; Xiao, T.; Gong, G. Ecosystem services changes between 2000 and 2015 in the Loess Plateau, China: A response to ecological restoration. PLoS ONE 2019, 14, e0209483. [Google Scholar] [CrossRef]
- Wen, X.; Zhen, L. Soil erosion control practices in the Chinese Loess Plateau: A systematic review. Environ. Dev. 2020, 34, 100493. [Google Scholar] [CrossRef]
- Chen, Y.; Wei, T.; Sha, G.; Zhu, Q.; Liu, Z.; Ren, K.; Yang, C. Soil enzyme activities of typical plant communities after vegetation restoration on the Loess Plateau, China. Appl. Soil Ecol. 2022, 170, 104292. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Ren, C.J.; Zhang, L.; Han, X.H.; Yang, G.H.; Wang, J. Changes in soil microbial community are linked to soil carbon fractions after afforestation. Eur. J. Soil Sci. 2018, 69, 370–379. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, G.; Li, P.; Xue, S. Dynamics of soil specific enzyme activities and temperature sensitivities during grassland succession after farmland abandonment. Catena 2021, 199, 105081. [Google Scholar] [CrossRef]
- Xu, H.W.; Qu, Q.; Chen, Y.H.; Liu, G.B.; Xue, S. Responses of soil enzyme activity and soil organic carbon stability over time after cropland abandonment in different vegetation zones of the Loess Plateau of China. Catena 2021, 196, 104812. [Google Scholar] [CrossRef]
- Liu, X.H.; Wei, T.X. High-throughput sequencing analysis of soil bacterial community in the grain for green project areas of the loess plateau. Environ. Sci. 2021, 43, 4489–4499. [Google Scholar]
- Mei, D.; Ma, X.; Fu, F.; Cao, F. Research Status and Development Prospects of Sea buckthorn (Hippophae rhamnoides L.) Resources in China. Forests 2023, 14, 2461. [Google Scholar] [CrossRef]
- Wu, R.; Wang, Y.; Huo, X.; Chen, W.; Wang, D. Drought and vegetation restoration patterns shape soil enzyme activity and nutrient limitation dynamics in the loess plateau. J. Environ. Manag. 2025, 374, 123846. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Guo, X.; Wang, X.; Wang, Y.; Li, P.; Zhang, Y.; Zhang, X. Responses of soil microbial communities to nutrient limitation in the desert-grassland ecological transition zone. Sci. Total Environ. 2018, 642, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yu, Y.J. Differences of soil microbial diversity and community composition between reclaimed woodland and grassland in coal waste pile and their influencing factors. Chin. J. Ecol. 2018, 37, 1662–1668. [Google Scholar]
- Ding, J.N. Effect of cultivation and natural restoration on soil microbial functional structure in coldregion wetlands. Appl. Ecol. Environ. Res. 2023, 21, 1471–1484. [Google Scholar] [CrossRef]
- Chen, X.; Feng, J.; Ding, Z.; Tang, M.; Zhu, B. Changes in soil total, microbial and enzymatic C-N-P contents and stoichiometry with depth and latitude in forest ecosystems. Sci. Total Environ. 2022, 816, 151583. [Google Scholar] [CrossRef]
- Pankratov, T.A.; Ivanova, A.O.; Dedysh, S.N.; Liesack, W. Bacterial populations and environmental factors controlling cellulose degradation in an acidic Sphagnum peat. Environ. Microbiol. 2011, 13, 1800–1814. [Google Scholar] [CrossRef]
- Lin, Y.; Yang, J.; Ye, Y. Analysis on diversity of soil bacterial community under different land use patterns in saline-alkali land. Acta Sci. Circumstantiae 2019, 39, 1266–1273. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.; Song, Z.; Wang, J.; Guo, L. Interactions of soil bacteria and fungi with plants during long-term grazing exclusion in semiarid grasslands. Soil Biol. Biochem. 2018, 124, 47–58. [Google Scholar] [CrossRef]
- Xu, Y.; Zhong, Z.; Zhang, W.; Han, X.; Yang, G.; Ren, C.; Feng, Y.; Ren, G.; Wang, X. Responses of soil nosZ-type denitrifying microbial communities to the various land-use types of the Loess Plateau, China. Soil Tillage Res. 2019, 195, 104378. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, X.; Mao, Z.; Yang, L.; Jiang, Z.; Chen, X.; Aubrey, D.P. Changes in soil microbial community structure following different tree species functional traits afforestation. Forests 2021, 12, 1018. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Code | Forest Types | Geographical Location | Stand Age/Year | Altitude/m | Slope Aspect | Slope/° | Main Vegetation |
|---|---|---|---|---|---|---|---|
| HrN | H. rhamnoides pure forest (shady slope) | 108°14′16.132″ E 36°53′45.027″ N | 20 | 1504 | North | 24 | Carex lanceolata Boott, Carduus nutans L., Buddleja lindleyana Fortune |
| HrS | H.rhamnoides pure forest (Sunny slope) | 108°14′16.132″ E 36°53′45.027″ N | 20 | 1521 | South | 15 | Carex lanceolata Boott, Potentilla strigosa Pall ex Pursh ex Pursh, Artemisia capillaris, Saussurea japonica (Thunb.) DC. |
| HrBo | H.rhamnoides and Biota orientalis mixed forest | 108°14′16.132″ E 36°53′45.027″ N | 20 | 1433 | South | 28 | Carex lanceolata Boott, Kalimeris indica (Linn.) Sch., Phragmites communis, Deyeuxia arundinacea (L.) Beauv., Hedyotis auricularia L. |
| HrPt | H.rhamnoides and Pinus tabulaeformis mixed forest | 108°14′16.132″ E 36°53′45.027″ N | 20 | 1408 | South | 18 | Carex lanceolata Boott, Potentilla strigosa Pall ex Pursh ex Pursh, Artemisia sacrorum Ledeb., Lespedeza bicolor Turcz. |
| GL | Grassland | 108°14′16.132″ E 36°53′45.027″ N | — | 1408 | South | 18 | Kalimeris indica (Linn.) Sch., Potentilla strigosa Pall ex Pursh ex Pursh, Lespedeza bicolor Turcz., Pterocypsela indica (L.) Shih, Artemisia annua |
| Plot Code | Soil Layer (cm) | BC (mg/kg) | BN (mg/kg) | BC/TOC (%) | BN/TN (%) | BN/AN (%) |
|---|---|---|---|---|---|---|
| HrN | 0–10 | 135.83 | 40.01 | 1.17 | 3.96 | 31.02 |
| 10–20 | 113.42 | 27.93 | 1.53 | 4.67 | 42.00 | |
| 20–40 | 86.05 | 16.90 | 2.05 | 3.01 | 29.03 | |
| 40–60 | 59.58 | 12.95 | 1.53 | 3.59 | 23.67 | |
| 60–80 | 78.15 | 11.56 | 3.13 | 3.23 | 22.41 | |
| 80–100 | 57.87 | 7.40 | 2.89 | 2.36 | 17.20 | |
| HrS | 0–10 | 157.72 | 79.00 | 1.04 | 6.32 | 72.48 |
| 10–20 | 154.14 | 32.54 | 1.59 | 3.97 | 39.64 | |
| 20–40 | 142.83 | 20.37 | 4.08 | 3.84 | 32.54 | |
| 40–60 | 100.77 | 8.45 | 4.20 | 2.14 | 16.64 | |
| 60–80 | 76.73 | 7.58 | 3.65 | 2.22 | 24.21 | |
| 80–100 | 86.49 | 6.80 | 4.12 | 2.12 | 22.83 | |
| HrBo | 0–10 | 159.40 | 40.04 | 0.50 | 3.15 | 31.04 |
| 10–20 | 95.12 | 28.96 | 1.29 | 3.88 | 37.03 | |
| 20–40 | 82.78 | 13.22 | 1.15 | 1.83 | 19.88 | |
| 40–60 | 53.73 | 6.92 | 0.78 | 1.12 | 13.61 | |
| 60–80 | 43.06 | 5.09 | 0.67 | 0.96 | 9.31 | |
| 80–100 | 59.58 | 3.10 | 1.22 | 0.67 | 6.60 | |
| HrPt | 0–10 | 179.02 | 18.03 | 8.95 | 6.22 | 65.80 |
| 10–20 | 127.62 | 21.17 | 9.82 | 9.53 | 60.13 | |
| 20–40 | 128.60 | 17.69 | 6.43 | 7.76 | 75.28 | |
| 40–60 | 125.45 | 29.59 | 6.27 | 13.33 | 54.09 | |
| 60–80 | 175.02 | 22.54 | 8.33 | 10.25 | 78.82 | |
| 80–100 | 114.30 | 15.84 | 8.16 | 7.27 | 76.52 |
| Sample Plot Type | Regression Model | BC and TOC | BN and TN | BN and AN | |||
|---|---|---|---|---|---|---|---|
| Regression Equation | R2 | Regression Equation | R2 | Regression Equation | R2 | ||
| HrN | Exponential | y = 0.7818e0.0195x | 0.83 | y = 0.2493e0.0347x | 0.93 | y = 0.4803e0.0219x | 0.87 |
| Linear | y = 0.1103x − 4.4926 | 0.88 | y = 0.0205x + 0.134 | 0.93 | y = 0.0177x + 0.4375 | 0.85 | |
| HrS | Exponential | y = 0.471e0.0173x | 0.92 | y = 0.3343e0.0183x | 0.89 | y = 36.321e0.0158x | 0.73 |
| Linear | y = 0.1317x − 10.314 | 0.86 | y = 0.0128x + 0.2789 | 0.97 | y = 1.0253x + 34.49 | 0.87 | |
| HrBo | Exponential | y = 2.5924e0.0144x | 0.84 | y = 0.4803e0.0219x | 0.87 | y = 45.007e0.0242x | 0.94 |
| Linear | y = 0.2201x − 7.3738 | 0.83 | y = 0.0177x + 0.4375 | 0.85 | y = 1.9374x + 39.59 | 0.90 | |
| HrPt | Exponential | y = 0.9366e0.0045x | 0.35 | y = 0.2637e−0.006x | 0.08 | y = 7.5664e0.0662x | 0.90 |
| Linear | y = 0.0076x + 0.7217 | 0.36 | y = −0.0016x + 0.2659 | 0.08 | y = 2.3702x − 17.641 | 0.91 | |
| Plot Code | Mean Length | Sobs | Shannon | Simpson | Ace | Chao1 | Coverage |
|---|---|---|---|---|---|---|---|
| HrN | 438.57 ± 0.89 a | 1398 ± 72 a | 6.20 ± 0.11 a | 0.004 ± 0.001 a | 1596.33 ± 73.19 a | 1632.10 ± 65.23 a | 0.986 ± 0.000 bc |
| HrS | 439.01 ± 0.34 a | 1428 ± 149 a | 6.27 ± 0.19 a | 0.004 ± 0.001 a | 1619.04 ± 49.26 a | 1633.50 ± 15.69 a | 0.986 ± 0.001 ac |
| HrBo | 438.48 ± 0.68 a | 1486 ± 23 a | 6.35 ± 0.01 a | 0.003 ± 0.000 a | 1653.70 ± 12.35 a | 1681.85 ± 18.58 a | 0.987 ± 0.001 ac |
| HrPt | 438.15 ± 0.77 ab | 1500 ± 69 a | 6.37 ± 0.09 a | 0.003 ± 0.001 a | 1660.69 ± 79.06 a | 1683.02 ± 11.59 a | 0.987 ± 0.001 ac |
| GL | 436.81 ± 0.92 a | 1394 ± 64 a | 6.19 ± 0.15 a | 0.005 ± 0.002 a | 1559.06 ± 56.21 a | 1577.40 ± 49.96 a | 0.987 ± 0.000 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Wei, T.; Fan, D.; Bi, H.; Zhu, Q. Response of Typical Artificial Forest Soil Microbial Community to Revegetation in the Loess Plateau, China. Agronomy 2025, 15, 1821. https://doi.org/10.3390/agronomy15081821
Liu X, Wei T, Fan D, Bi H, Zhu Q. Response of Typical Artificial Forest Soil Microbial Community to Revegetation in the Loess Plateau, China. Agronomy. 2025; 15(8):1821. https://doi.org/10.3390/agronomy15081821
Chicago/Turabian StyleLiu, Xiaohua, Tianxing Wei, Dehui Fan, Huaxing Bi, and Qingke Zhu. 2025. "Response of Typical Artificial Forest Soil Microbial Community to Revegetation in the Loess Plateau, China" Agronomy 15, no. 8: 1821. https://doi.org/10.3390/agronomy15081821
APA StyleLiu, X., Wei, T., Fan, D., Bi, H., & Zhu, Q. (2025). Response of Typical Artificial Forest Soil Microbial Community to Revegetation in the Loess Plateau, China. Agronomy, 15(8), 1821. https://doi.org/10.3390/agronomy15081821