
Long-Term Conservation Tillage Increases Soil Organic Carbon Stability by Modulating Microbial Nutrient Limitations and Aggregate Protection

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sampling and Laboratory Incubation
2.3. Aggregate Distribution
2.4. Soil Microbial Biomass Carbon and Nitrogen
2.5. Soil Enzyme Activity
2.6. Statistical Analysis
3. Results
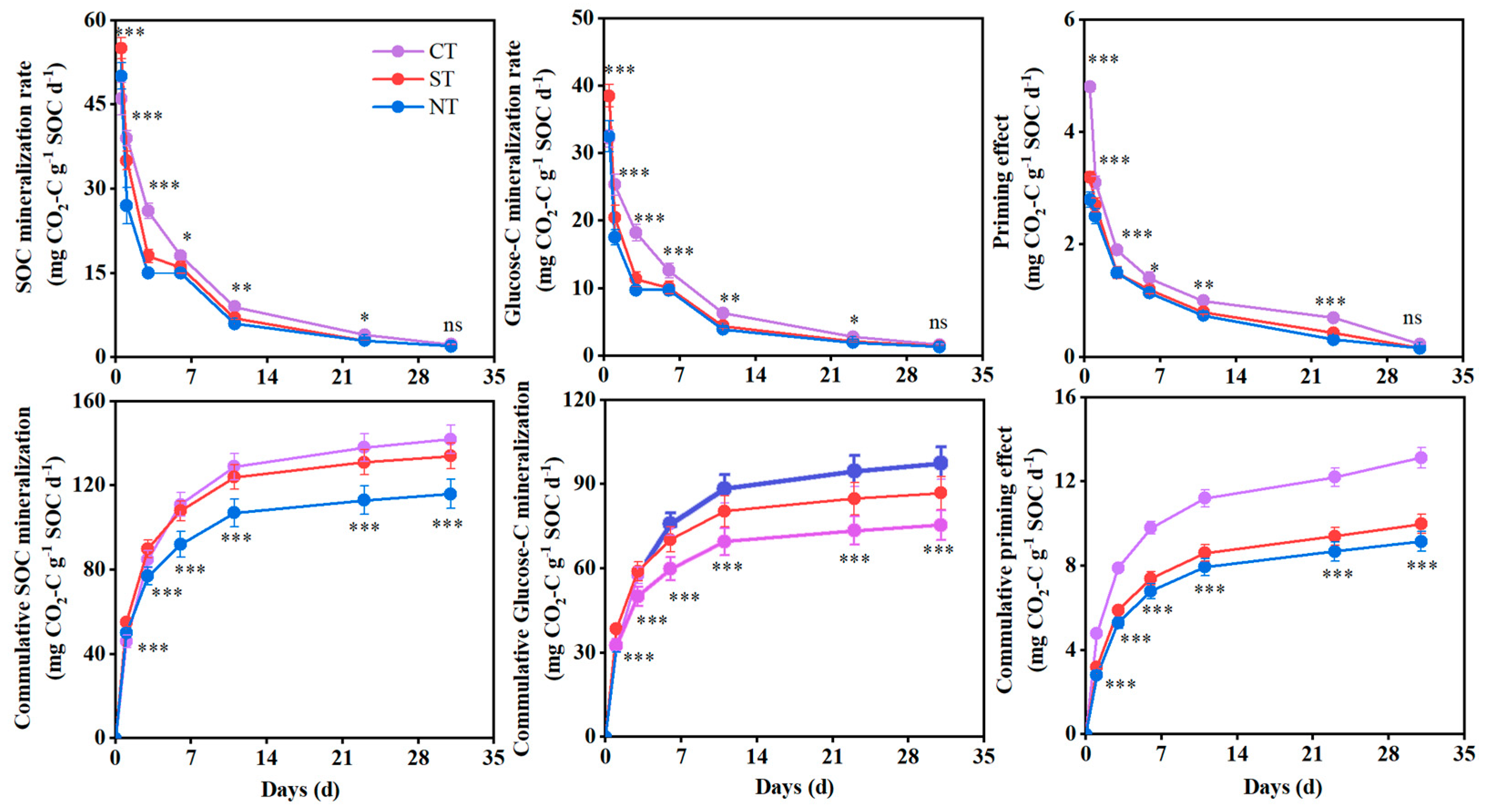
3.1. SOC Mineralization and Priming Effect
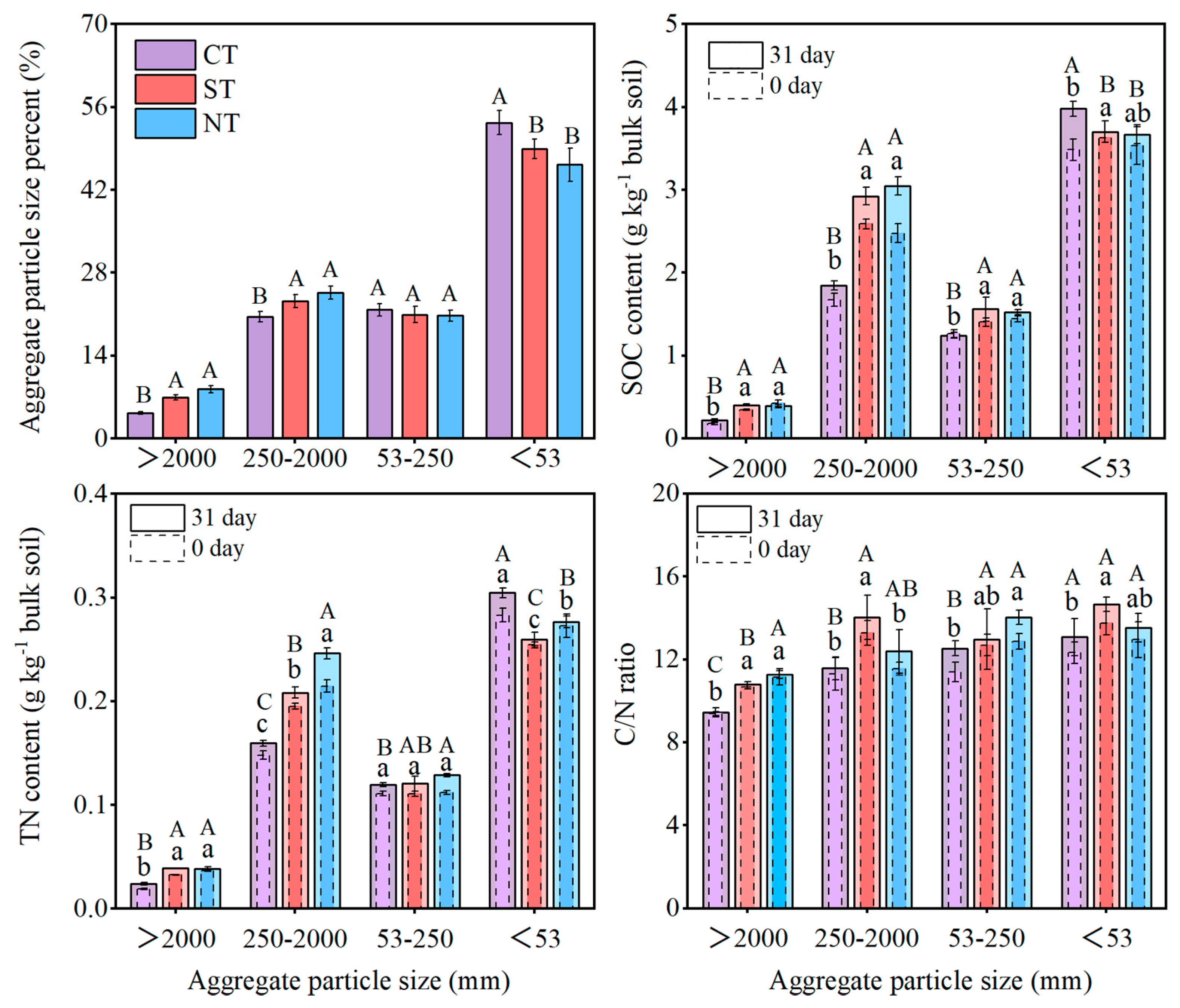
3.2. Aggregate Distribution and the Associated Carbon and Nitrogen Content
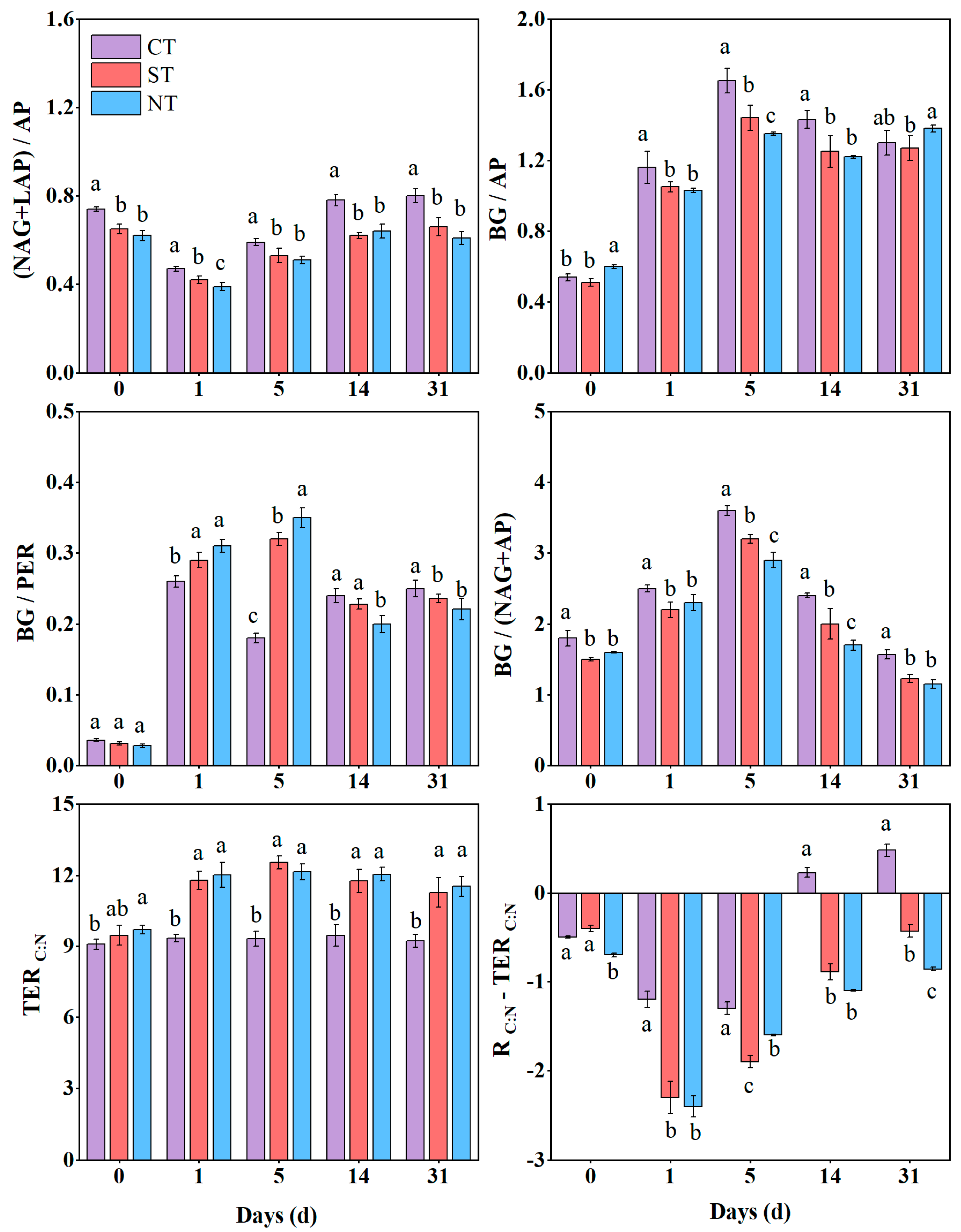
3.3. Soil Enzyme Stoichiometry
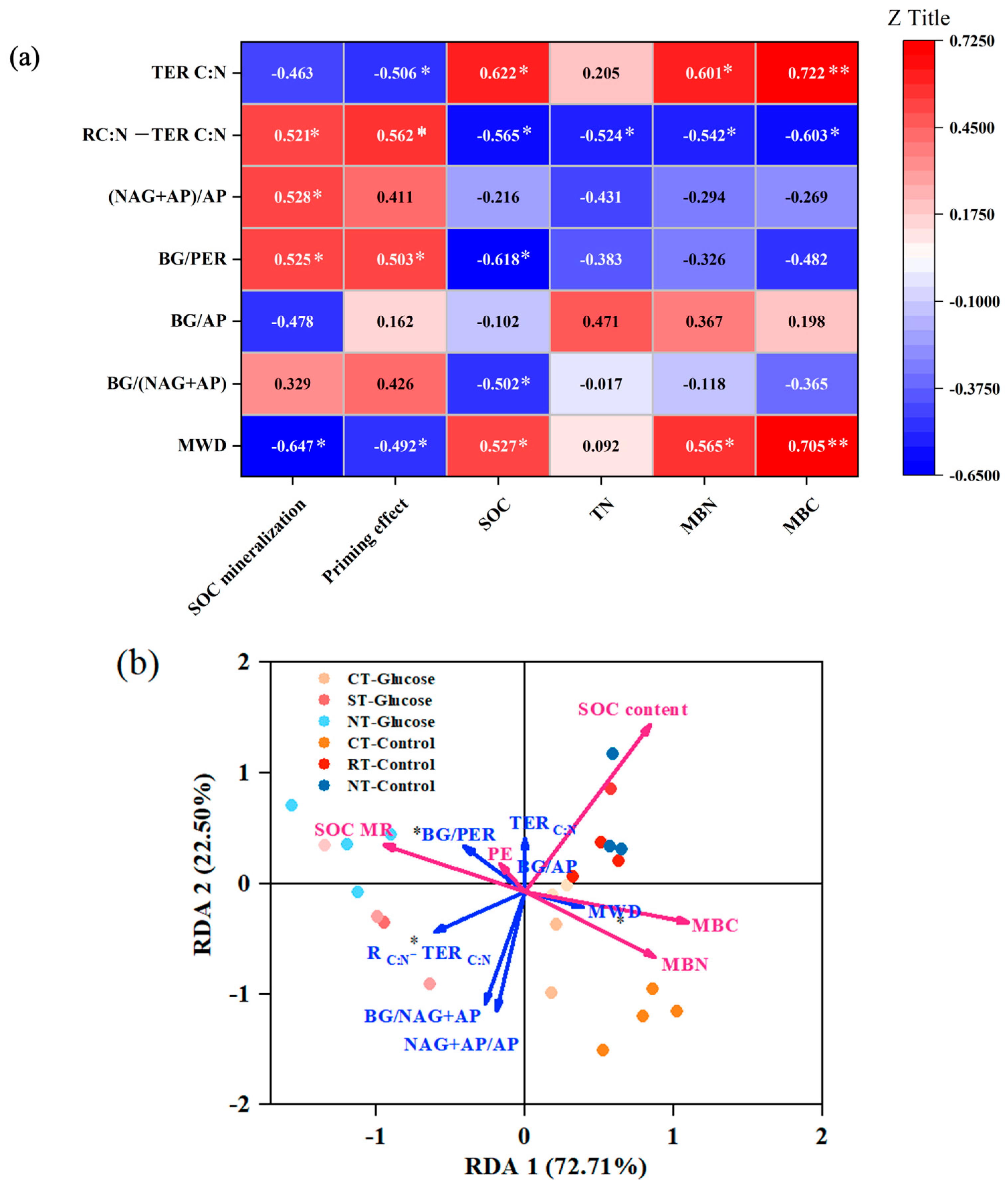
3.4. Key Factors Affecting SOC Sequestration and Decomposition
4. Discussion
4.1. Effects of Conservation Tillage on SOC Mineralization and Priming Response
4.2. The Regulation Mechanism of Priming Effect Under Conservation Tillage
4.3. Physical, Chemical and Microbial Nutrient Limitation Regulated SOC Stability Under Conservation Tillage
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SOC | Soil organic carbon |
| TN | Total nitrogen |
| CT | Conventional tillage with straw removed |
| ST | Subsoil tillage with straw mulching |
| NT | No tillage with straw mulching |
| BG | β-glucosidase |
| NAG | N-acetyl-β-D-glucosidase |
| AP | Phosphatase |
| PER | Peroxidase |
| TERC:N | Carbon–nitrogen imbalance of available resources |
| MBC | Microbial biomass carbon |
| MBN | Microbial biomass nitrogen |
| MWD | Mean weight diameter |
References
- Agnihotri, R.; Sharma, M.P.; Prakash, A.; Ramesh, A.; Bhattacharjya, S.; Patra, A.K.; Manna, M.C.; Kurganova, I.; Kuzyakov, Y. Glycoproteins of Arbuscular Mycorrhiza for Soil Carbon Sequestration: Review of Mechanisms and Controls. Sci. Total Environ. 2022, 806, 150571. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Zong, N.; Hartley, I.P.; He, N.; Zhang, J.; Powlson, D.; Zhou, J.; Kuzyakov, Y.; Zhang, F.; Yu, G.; et al. Microbial Metabolic Response to Winter Warming Stabilizes Soil Carbon. Glob. Change Biol. 2021, 27, 2011–2028. [Google Scholar] [CrossRef] [PubMed]
- Dimassi, B.; Mary, B.; Fontaine, S.; Perveen, N.; Revaillot, S.; Cohan, J.P. Effect of Nutrients Availability and Long-Term Tillage on Priming Effect and Soil C Mineralization. Soil Biol. Biochem. 2014, 78, 332–339. [Google Scholar] [CrossRef]
- Sauvadet, M.; Lashermes, G.; Alavoine, G.; Recous, S.; Chauvat, M.; Maron, P.-A.; Bertrand, I. High Carbon Use Efficiency and Low Priming Effect Promote Soil C Stabilization under Reduced Tillage. Soil Biol. Biochem. 2018, 123, 64–73. [Google Scholar] [CrossRef]
- Xu, P.; Zhu, J.; Wang, H.; Shi, L.; Zhuang, Y.; Fu, Q.; Chen, J.; Hu, H.; Huang, Q. Regulation of Soil Aggregate Size under Different Fertilizations on Dissolved Organic Matter, Cellobiose Hydrolyzing Microbial Community and Their Roles in Organic Matter Mineralization. Sci. Total Environ. 2021, 755, 142595. [Google Scholar] [CrossRef]
- Moreno, F.; Murillo, J.M.; Pelegrín, F.; Girón, I.F. Long-Term Impact of Conservation Tillage on Stratification Ratio of Soil Organic Carbon and Loss of Total and Active CaCO3. Soil Tillage Res. 2006, 85, 86–93. [Google Scholar] [CrossRef]
- Bell, J.M.; Smith, J.L.; Bailey, V.L.; Bolton, H. Priming Effect and C Storage in Semi-Arid No-till Spring Crop Rotations. Biol. Fertil. Soils 2003, 37, 237–244. [Google Scholar] [CrossRef]
- Nandan, R.; Singh, V.; Singh, S.S.; Kumar, V.; Hazra, K.K.; Nath, C.P.; Poonia, S.P.; Malik, R.K.; Bhattacharyya, R.; McDonald, A. Impact of Conservation Tillage in Rice–Based Cropping Systems on Soil Aggregation, Carbon Pools and Nutrients. Geoderma 2019, 340, 104–114. [Google Scholar] [CrossRef]
- Plaza, C.; Courtier-Murias, D.; Fernández, J.M.; Polo, A.; Simpson, A.J. Physical, Chemical, and Biochemical Mechanisms of Soil Organic Matter Stabilization under Conservation Tillage Systems: A Central Role for Microbes and Microbial by-Products in C Sequestration. Soil Biol. Biochem. 2013, 57, 124–134. [Google Scholar] [CrossRef]
- Gao, L.; Becker, E.; Liang, G.; Houssou, A.A.; Wu, H.; Wu, X.; Cai, D.; Degré, A. Effect of Different Tillage Systems on Aggregate Structure and Inner Distribution of Organic Carbon. Geoderma 2017, 288, 97–104. [Google Scholar] [CrossRef]
- Kan, Z.R.; Ma, S.T.; Liu, Q.Y.; Liu, B.Y.; Virk, A.L.; Qi, J.Y.; Zhao, X.; Lal, R.; Zhang, H.L. Carbon Sequestration and Mineralization in Soil Aggregates under Long-Term Conservation Tillage in the North China Plain. CATENA 2020, 188, 104428. [Google Scholar] [CrossRef]
- Schlüter, S.; Leuther, F.; Albrecht, L.; Hoeschen, C.; Kilian, R.; Surey, R.; Mikutta, R.; Kaiser, K.; Mueller, C.W.; Vogel, H.J. Microscale Carbon Distribution around Pores and Particulate Organic Matter Varies with Soil Moisture Regime. Nat. Commun. 2022, 13, 2098. [Google Scholar] [CrossRef]
- Mangalassery, S.; Mooney, S.J.; Sparkes, D.L.; Fraser, W.T.; Sjögersten, S. Impacts of Zero Tillage on Soil Enzyme Activities, Microbial Characteristics and Organic Matter Functional Chemistry in Temperate Soils. Eur. J. Soil Biol. 2015, 68, 9–17. [Google Scholar] [CrossRef]
- Pandey, D.; Agrawal, M.; Bohra, J.S. Effects of Conventional Tillage and No Tillage Permutations on Extracellular Soil Enzyme Activities and Microbial Biomass under Rice Cultivation. Soil Tillage Res. 2014, 136, 51–60. [Google Scholar] [CrossRef]
- Zheng, F.; Wu, X.; Zhang, M.; Liu, X.; Song, X.; Lu, J.; Wang, B.; Jan van Groenigen, K.; Li, S. Linking Soil Microbial Community Traits and Organic Carbon Accumulation Rate under Long-Term Conservation Tillage Practices. Soil Tillage Res. 2022, 220, 105360. [Google Scholar] [CrossRef]
- Li, Q.; Chen, J.; Feng, J.; Wu, J.; Zhang, Q.; Jia, W.; Lin, Q.; Cheng, X. How Do Biotic and Abiotic Factors Regulate Soil Enzyme Activities at Plot and Microplot Scales Under Afforestation? Ecosystems 2020, 23, 1408–1422. [Google Scholar] [CrossRef]
- Wang, C.; Lu, X.; Mori, T.; Mao, Q.; Zhou, K.; Zhou, G.; Nie, Y.; Mo, J. Responses of Soil Microbial Community to Continuous Experimental Nitrogen Additions for 13 Years in a Nitrogen-Rich Tropical Forest. Soil Biol. Biochem. 2018, 121, 103–112. [Google Scholar] [CrossRef]
- Engell, I.; Linsler, D.; Sandor, M.; Joergensen, R.G.; Meinen, C.; Potthoff, M. The Effects of Conservation Tillage on Chemical and Microbial Soil Parameters at Four Sites across Europe. Plants 2022, 11, 1747. [Google Scholar] [CrossRef]
- Liang, Z.; Rasmussen, J.; Poeplau, C.; Elsgaard, L. Priming Effects Decrease with the Quantity of Cover Crop Residues—Potential Implications for Soil Carbon Sequestration. Soil Biol. Biochem. 2023, 184, 109110. [Google Scholar] [CrossRef]
- Olsson, P.A.; Bååth, E.; Jakobsen, I.; Söderström, B. The Use of Phospholipid and Neutral Lipid Fatty Acids to Estimate Biomass of Arbuscular Mycorrhizal Fungi in Soil. Mycol. Res. 1995, 99, 623–629. [Google Scholar] [CrossRef]
- Song, X.; Liu, X.; Liang, G.; Li, S.; Li, J.; Zhang, M.; Zheng, F.; Ding, W.; Wu, X.; Wu, H. Positive Priming Effect Explained by Microbial Nitrogen Mining and Stoichiometric Decomposition at Different Stages. Soil Biol. Biochem. 2022, 175, 108852. [Google Scholar] [CrossRef]
- Bayer, C.; Mielniczuk, J.; Giasson, E.; Martin-Neto, L.; Pavinato, A. Tillage Effects on Particulate and Mineral-Associated Organic Matter in Two Tropical Brazilian Soils. Commun. Soil Sci. Plant Anal. 2006, 37, 389–400. [Google Scholar] [CrossRef]
- Liu, X.; Li, Q.; Tan, S.; Wu, X.; Song, X.; Gao, H.; Han, Z.; Jia, A.; Liang, G.; Li, S. Evaluation of Carbon Mineralization and Its Temperature Sensitivity in Different Soil Aggregates and Moisture Regimes: A 21-Year Tillage Experiment. Sci. Total Environ. 2022, 837, 155566. [Google Scholar] [CrossRef]
- Chen, C.; Meile, C.; Wilmoth, J.; Barcellos, D.; Thompson, A. Influence of pO2 on Iron Redox Cycling and Anaerobic Organic Carbon Mineralization in a Humid Tropical Forest Soil. Environ. Sci. Technol. 2018, 52, 7709–7719. [Google Scholar] [CrossRef]
- Feng, S.; Huang, Y.; Ge, Y.; Su, Y.; Xu, X.; Wang, Y.; He, X. Variations in the Patterns of Soil Organic Carbon Mineralization and Microbial Communities in Response to Exogenous Application of Rice Straw and Calcium Carbonate. Sci. Total Environ. 2016, 571, 615–623. [Google Scholar] [CrossRef]
- Nyamadzawo, G.; Nyamangara, J.; Nyamugafata, P.; Muzulu, A. Soil Microbial Biomass and Mineralization of Aggregate Protected Carbon in Fallow-Maize Systems under Conventional and No-Tillage in Central Zimbabwe. Soil Tillage Res. 2009, 102, 151–157. [Google Scholar] [CrossRef]
- Zheng, F.; Liu, X.; Zhang, M.; Li, S.; Song, X.; Wang, B.; Wu, X.; Van Groenigen, K.J. Strong Links between Aggregate Stability, Soil Carbon Stocks and Microbial Community Composition across Management Practices in a Chinese Dryland Cropping System. CATENA 2023, 233, 107509. [Google Scholar] [CrossRef]
- Margenot, A.J.; Nakayama, Y.; Parikh, S.J. Methodological Recommendations for Optimizing Assays of Enzyme Activities in Soil Samples. Soil Biol. Biochem. 2018, 125, 350–360. [Google Scholar] [CrossRef]
- Deng, S.; Popova, I.E.; Dick, L.; Dick, R. Bench Scale and Microplate Format Assay of Soil Enzyme Activities Using Spectroscopic and Fluorometric Approaches. Appl. Soil Ecol. 2013, 64, 84–90. [Google Scholar] [CrossRef]
- DeForest, J.L. The Influence of Time, Storage Temperature, and Substrate Age on Potential Soil Enzyme Activity in Acidic Forest Soils Using MUB-Linked Substrates and l-DOPA. Soil Biol. Biochem. 2009, 41, 1180–1186. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of Soil Enzyme Activity at Global Scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- Nannipieri, P.; Smalla, K. Soil biology. In Nucleic Acids and Proteins in Soil: With 42 Figures, 2 in Color; Springer: Berlin/Heidelberg, Germany, 2006; ISBN 978-3-540-29448-1. [Google Scholar]
- Zimmerman, A.R.; Gao, B.; Ahn, M.-Y. Positive and Negative Carbon Mineralization Priming Effects among a Variety of Biochar-Amended Soils. Soil Biol. Biochem. 2011, 43, 1169–1179. [Google Scholar] [CrossRef]
- Andruschkewitsch, R.; Geisseler, D.; Koch, H.-J.; Ludwig, B. Effects of Tillage on Contents of Organic Carbon, Nitrogen, Water-Stable Aggregates and Light Fraction for Four Different Long-Term Trials. Geoderma 2013, 192, 368–377. [Google Scholar] [CrossRef]
- Mo, F.; Zhang, Y.-Y.; Liu, Y.; Liao, Y.-C. Microbial Carbon-Use Efficiency and Straw-Induced Priming Effect within Soil Aggregates Are Regulated by Tillage History and Balanced Nutrient Supply. Biol. Fertil. Soils 2021, 57, 409–420. [Google Scholar] [CrossRef]
- Kan, Z.R.; Liu, W.X.; Liu, W.S.; Lal, R.; Dang, Y.P.; Zhao, X.; Zhang, H.L. Mechanisms of Soil Organic Carbon Stability and Its Response to No-till: A Global Synthesis and Perspective. Glob. Change Biol. 2022, 28, 693–710. [Google Scholar] [CrossRef]
- Song, X.; Li, J.; Liu, X.; Liang, G.; Li, S.; Zhang, M.; Zheng, F.; Wang, B.; Wu, X.; Wu, H. Altered Microbial Resource Limitation Regulates Soil Organic Carbon Sequestration Based on Ecoenzyme Stoichiometry under Long-Term Tillage Systems. Land Degrad. Dev. 2022, 33, 2795–2808. [Google Scholar] [CrossRef]
- Zhang, B.; Li, Y.; Ren, T.; Tian, Z.; Wang, G.; He, X.; Tian, C. Short-Term Effect of Tillage and Crop Rotation on Microbial Community Structure and Enzyme Activities of a Clay Loam Soil. Biol. Fertil. Soils 2014, 50, 1077–1085. [Google Scholar] [CrossRef]
- Han, S.; Delgado-Baquerizo, M.; Luo, X.; Liu, Y.; Van Nostrand, J.D.; Chen, W.; Zhou, J.; Huang, Q. Soil Aggregate Size-Dependent Relationships between Microbial Functional Diversity and Multifunctionality. Soil Biol. Biochem. 2021, 154, 108143. [Google Scholar] [CrossRef]
- Cai, Y.; Ma, T.; Wang, Y.; Jia, J.; Jia, Y.; Liang, C.; Feng, X. Assessing the Accumulation Efficiency of Various Microbial Carbon Components in Soils of Different Minerals. Geoderma 2022, 407, 115562. [Google Scholar] [CrossRef]
- Raiesi, F.; Kabiri, V. Carbon and Nitrogen Mineralization Kinetics as Affected by Tillage Systems in a Calcareous Loam Soil. Ecol. Eng. 2017, 106, 24–34. [Google Scholar] [CrossRef]
- Pramanick, B.; Kumar, M.; Naik, B.M.; Singh, S.K.; Kumar, M.; Singh, S.V. Soil Carbon-Nutrient Cycling, Energetics, and Carbon Footprint in Calcareous Soils with Adoption of Long-Term Conservation Tillage Practices and Cropping Systems Diversification. Sci. Total Environ. 2024, 912, 169421. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Duan, X.; Guo, X.; Gao, W.; Li, Y.; Zhou, P.; Zhu, Q.; O’Donnell, A.G.; Dai, K.; Wu, J. Microbial Metabolic Capacity Regulates the Accrual of Mineral-Associated Organic Carbon in Subtropical Paddy Soils. Soil Biol. Biochem. 2024, 195, 109457. [Google Scholar] [CrossRef]
- Han, Z.; Wu, X.; Liang, A.; Li, S.; Gao, H.; Song, X.; Liu, X.; Gao, H.; Degré, A. Conservation tillage enhances the sequestration and iron-mediated stabilization of aggregate-associated organic carbon in mollisols. Catena 2024, 243, 108197. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, Y.; Zhang, Y.; Huang, D.; Li, X.; Gregorich, E.; McLaughlin, N.; Zhang, X.; Chen, X.; Zhang, S.; et al. Effect of Long-Term Tillage and Cropping System on Portion of Fungal and Bacterial Necromass Carbon in Soil Organic Carbon. Soil Tillage Res. 2022, 218, 105307. [Google Scholar] [CrossRef]
- Chellappa, J.; Sagar, K.L.; Sekaran, U.; Kumar, S.; Sharma, P. Soil Organic Carbon, Aggregate Stability and Biochemical Activity under Tilled and No-Tilled Agroecosystems. J. Agric. Food Res. 2021, 4, 100139. [Google Scholar] [CrossRef]
- Jilling, A.; Keiluweit, M.; Gutknecht, J.L.M.; Grandy, A.S. Priming mechanisms providing plants and microbes access to mineral-associated organic matter. Soil Biol. Biochem. 2021, 158, 108265. [Google Scholar] [CrossRef]
- Qiu, H.; Ge, T.; Liu, J.; Chen, X.; Hu, Y.; Su, Y.; Wu, J.; Kuzyakov, Y. Effects of biotic and abiotic factors on soil organic matter mineralization: Experiments and structural modeling analysis. Eur. J. Soil Biol. 2018, 84, 27–34. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Tillage | MWD | SOC | TN | SOC/TN | MBC | MBN | MBC/MBN |
|---|---|---|---|---|---|---|---|---|
| 0 day | CT | 0.431 c | 6.69 b | 0.78 b | 8.58 b | 172.5 b | 30.17 b | 5.72 b |
| ST | 0.548 b | 8.70 a | 0.96 a | 9.06 ab | 247.5 a | 33.26 a | 7.44 a | |
| NT | 0.613 a | 8.92 a | 0.97 a | 9.19 a | 256.3 a | 35.85 a | 7.15 a | |
| 31 days | CT | 0.472 b | 8.24 b | 0.85 b | 9.74 b | 206.7 c | 24.52 c | 8.43 b |
| ST | 0.595 a | 11.12 a | 1.03 a | 10.85 a | 306.9 b | 30.31 b | 10.13 a | |
| NT | 0.610 a | 11.67 a | 1.09 a | 10.67 a | 367.7 a | 36.36 a | 10.11 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Z.; Wu, X.; Gao, H.; Jia, A.; Gao, Q. Long-Term Conservation Tillage Increases Soil Organic Carbon Stability by Modulating Microbial Nutrient Limitations and Aggregate Protection. Agronomy 2025, 15, 1571. https://doi.org/10.3390/agronomy15071571
Han Z, Wu X, Gao H, Jia A, Gao Q. Long-Term Conservation Tillage Increases Soil Organic Carbon Stability by Modulating Microbial Nutrient Limitations and Aggregate Protection. Agronomy. 2025; 15(7):1571. https://doi.org/10.3390/agronomy15071571
Chicago/Turabian StyleHan, Zixuan, Xueping Wu, Huizhou Gao, Angyuan Jia, and Qiqi Gao. 2025. "Long-Term Conservation Tillage Increases Soil Organic Carbon Stability by Modulating Microbial Nutrient Limitations and Aggregate Protection" Agronomy 15, no. 7: 1571. https://doi.org/10.3390/agronomy15071571
APA StyleHan, Z., Wu, X., Gao, H., Jia, A., & Gao, Q. (2025). Long-Term Conservation Tillage Increases Soil Organic Carbon Stability by Modulating Microbial Nutrient Limitations and Aggregate Protection. Agronomy, 15(7), 1571. https://doi.org/10.3390/agronomy15071571