
Double Mutations Drive Multiple Resistances to Herbicides in Greek Rigid Ryegrass (Lolium rigidum Gaudin)


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Whole-Plant Dose–Response Assays for the 14 Putative Resistance Populations
2.3. Amplification and Sequencing of Als and Accase Gene Fragments
3. Results
3.1. Lolium rigidum Seed Germination Ability of 40 Populations
3.2. Whole-Plant Dose–Response Assays
3.3. Amplification and Sequencing of Accace and Als Gene Fragments
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vahamidis, P.; Stefopoulou, A.; Lagogianni, C.S.; Economou, G.; Dercas, N.; Kotoulas, V.; Kalivas, D.; Tsitsigiannis, D.I. Pyrenophora teres and Rhynchosporium secalis Establishment in a Mediterranean Malt Barley Field: Assessing Spatial, Temporal and Management Effects. Agriculture 2020, 10, 553. [Google Scholar] [CrossRef]
- Briggs, D.E. Malt modification—A century of evolving views. J. Inst. Brew. 2002, 108, 395–405. [Google Scholar] [CrossRef]
- Mornhinweg, D.W. Biotic stress in barley: Insects problems and solutions. In Barley: Production, Improvement, and Uses; Ulrich, S.E., Ed.; Wiley-Blackwell Pubblishing: Hoboken, NJ, USA, 2011; pp. 355–390. [Google Scholar]
- FAO. FAOSTAT Database; Food and Agriculture Organization of the United Nation: Rome, Italy, 2022. [Google Scholar]
- Watson, P.R.; Driksen, D.A.; Van Acker, R.C. The ability of 29 cultivars to compete and withstand competition. Weed Sci. 2006, 57, 783–792. [Google Scholar] [CrossRef]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Papapanagiotou, A.P.; Paresidou, M.I.; Kaloumenos, N.S.; Eleftherohorinos, I.G. ACCase mutations in Avena sterilis populations and their impact on plant fitness. Pestic. Biochem. Physiol. 2015, 123, 40–48. [Google Scholar] [CrossRef]
- Kaloumenos, N.S.; Tsioni, V.C.; Daliani, E.G.; Papavassileiou, S.E.; Vassileiou, A.G.; Laoutidou, P.N.; Eleftherohorinos, I.G. Multiple Pro-197 substitutions in the acetolactate synthase of rigid ryegrass (Lolium rigidum) and their impact on chlorsulfuron activity and plant growth. Crop Prot. 2012, 38, 35–43. [Google Scholar] [CrossRef]
- Anthimidou, E.; Ntoanidou, S.; Madesis, P.; Eleftherohorinos, I.G. Mechanisms of Lolium rigidum multiple resistance to ALS- and ACCase-inhibiting herbicides and their impact on plant fitness. Pestic. Biochem. Physiol. 2019, 164, 65–72. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S.B. Metabolism-based herbicide resistance and cross- resistance in crop weeds: A threat to herbocode sustainability and global crop production. Plant Physiol. 2014, 166, 1106–1118. [Google Scholar] [CrossRef]
- Spoor, W.; McCraw, J. Self incomptability in lolium: A reply. Heredity 1984, 53, 239–240. [Google Scholar] [CrossRef]
- Travlos, I.; Gazoulis, I.; Kanatas, P.; Tsekoura, A.; Zannopoulos, S.; Papastylianou, P. Key factors affecting weed seeds’ germination, weed Emergence, and their possible role for the efficacy of false seedbed technique as weed management practice. Front. Agron. 2020, 2, 1. [Google Scholar] [CrossRef]
- Kotoula-Syka, E.; Tal, A.; Rubin, B. Diclofop-resistant Lolium rigidum from northern Greece with cross-resistance to ACCase inhibitors and multiple resistance to chlorsulfuron. Pest Manag. Sci. 2000, 56, 1054–1058. [Google Scholar] [CrossRef]
- Scarabel, L.; Panozzo, S.; Loddo, D.; Mathiassen, S.K.; Kristensen, M.; Kudsk, P.; Gitsopoulos, T.; Travlos, I.; Tani, E.; Chachalis, D.; et al. Diversified Resistance Mechanisms in Multi-Resistant Lolium spp. in Three European Countries. Front. Plant Sci. 2020, 11, 608845. [Google Scholar] [CrossRef] [PubMed]
- Goggin, D.E.; Powles, S.B.; Steadman, K.J. Understanding Lolium rigidum Seeds: The Key to Managing a Problem Weed. Agronomy 2012, 2, 222–239. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Latif, S.; Borger, C.; Iqbal, N.; Asaduzzaman, M.D.; Wu, H.; Walsh, M. The Remarkable Journey of a Weed: Biology and Management of Annual Ryegrass (Lolium rigidum) in Conservation Cropping Systems of Australia. Plants 2021, 10, 1505. [Google Scholar] [CrossRef]
- Izquierdo, J.; Recasens, J.; Fernández-Quintanilla, C.; Gill, G. Effects of crop and weed densities on the interactions between barley and Lolium rigidum in several Mediterranean locations. Agronomie 2003, 23, 529–536. [Google Scholar] [CrossRef]
- Lemerle, D.; Verbeek, B.; Coombes, N. Losses in grain yield of winter crops from Lolium rigidum competition depend on crop species, cultivar and season. Weed Res. 1995, 35, 503–509. [Google Scholar] [CrossRef]
- Menegat, A.; Bailly, G.C.; Aponte, R.; Heinrich, G.M.T.; Sievernich, B.; Gerhards, R. Acetohydroxyacid synthase (AHAS) amino acid substitution Asp376Glu in Lolium perenne: Effect on herbicide efficacy and plant growth. J. Plant Dis. Prot. 2016, 123, 145–153. [Google Scholar] [CrossRef]
- Fernández, P.; Alcántara, R.; Osuna, M.D.; Vila-Aiub, M.M.; De Prado, R. Forward selection for multiple resistance across the non-selective glyphosate, glufosinate and oxyfluorfen herbicides in Lolium weed species. Pest Manag. Sci. 2017, 73, 936–944. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org/Summary/Country.aspx (accessed on 22 June 2025).
- Délye, C.; Jasieniuk, M.; Le Corre, V. Deciphering the evolution of herbicide resistance in weeds. Trends Genet. 2013, 29, 649–658. [Google Scholar] [CrossRef]
- Torra, J.; Montull, J.M.; Taberner, A.; Onkokesung, N.; Boonham, N.; Edwards, R. Target-Site and Non-target-Site Resistance Mechanisms Confer Multiple and Cross-Resistance to ALS and ACCase Inhibiting Herbicides in Lolium rigidum from Spain. Front. Plant Sci. 2021, 12, 625138. [Google Scholar] [CrossRef]
- Bi, Y.; Liu, W.; Guo, W.; Li, L.; Yuan, G.; Du, L.; Wang, J. Molecular basis of multiple resistance to ACCase- and ALS-inhibiting herbicides in Alopecurus japonicus from China. Pestic. Biochem. Physiol. 2016, 126, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed]
- Beckie, H.J.; Heap, I.M.; Smeda, R.J.; Hall, L.M. Screening for herbicide resistance in weeds. Weed Technol. 2000, 14, 428–445. [Google Scholar] [CrossRef]
- Falk, J.S.; Shoup, D.E.; Al-Khatib, K.; Peterson, D.E. Survey of common waterhemp (Amaranthus rudis) response to protox- and ALS inhibiting herbicides in northeast Kansas. Weed Technol. 2005, 19, 838–846. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. Adecimal code for the growth stage of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Seefeldt, S.S.; Jensen, H.; Fuerst, E.P. Log-logistic analysis of herbicide dose-response relationships. Weed Technol. 1995, 9, 218–227. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Boutsalis, P.; Gurjeet, S.G.; Preston, C. Incidence of herbicide resistance in rigid ryegrass (Lolium rigidum) across Southeastern Australia. Weed Technol. 2012, 26, 391–398. [Google Scholar] [CrossRef]
- Matzrafi, M.; Gerson, O.; Rubin, B.; Peleg, Z. Different mutations endowing resistance to Acetyl-CoA carboxylase inhibitors results in changes in ecological fitness of Lolium rigidum populations. Front. Plant Sci. 2017, 8, 1078. [Google Scholar] [CrossRef]
- Papapanagiotou, A.P.; Loukovitis, D.; Anthimidou, E.; Eleftherohorinos, I.G. Impact of ALS Herbicide-Resistant Perennial Ryegrass (Lolium perenne) Population on Growth Rate and Competitive Ability against Wheat. Agronomy 2023, 13, 1641. [Google Scholar] [CrossRef]
- Mucheri, T.; Rugare, J.T.; Bajwa, A.A. Mechanistic understanding and sustainable management of non-target site herbicide resistance in modern day agriculture. Adv. Weed Sci. 2024, 42, e020240056. [Google Scholar] [CrossRef] [PubMed]
- El-Mastouri, Z.; Košnarová, P.; Hamouzova, K.; Alimi, E.; Soukup, J. Insight into the herbicide resistance patterns in Lolium rigidum populations in Tunisian and Moroccan wheat regions. Front. Plant Sci. 2024, 15, 1331725. [Google Scholar] [CrossRef]
- Jugulam, M.; Shyam, C. Non-target-site resistance to herbicides: Recent developments. Plants 2019, 8, 417. [Google Scholar] [CrossRef]
- Christopher, J.T.; Powles, S.B.; Lijegren, D.R.; Holtum, J.A. Cross-resistance to herbicides in annual ryegrass (Lolium rigidum): II. Chlorsulfuron resistance involves a wheat-like detoxification system. Plant Physiol. 1991, 95, 1036–1043. [Google Scholar] [CrossRef]
- Christopher, J.T.; Powles, S.B.; Holtum, J.A.M. Resistance to acetolactate synthase-inhibiting herbicides in annual ryegrass (Lolium rigidum) involves at least two mechanisms. Plant Physiol. 1992, 100, 1909–1913. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-García, J.G.; Alcántara-de la Cruz, R.; Palma-Bautista, C.; Rojano-Delgado, A.M.; Cruz-Hipólito, H.E.; Torra, J.; Barro, F.; De Prado, R. Accumulation of Target Gene Mutations Confers Multiple Resistance to ALS, ACCase, and EPSPS Inhibitors in Lolium Species in Chilie. Front. Plant Sci. 2020, 11, 553948. [Google Scholar] [CrossRef]
- Tavassoli, A.; Gherekhloo, J.; Ghaderi-Far, F.; Zandd, E.; Osunae, M.D.; Prado, R. Ryegrass (Lolium rigidum Gaud) Resistant to ACCase and ALS inhibitors in northeastern Iran. Weed Sci. 2023, 41, e020220069. [Google Scholar] [CrossRef]
- Délye, C.; Zhang, X.-Q.; Chalopin, C.; Michel, S.; Powles, S.B. An isoleucine residue within the carboxyl-transferase domain of multidomain acetyl-CoA carboxylase is a major determinant of sensitivity to aryloxyphenoxypropionate but not to cyclohexanedione inhibitors. Plant Physiol. 2003, 132, 1716–1723. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Powles, S.B. Six amino acid substitutions in the carboxyl-transferase domain of the plastidic acetyl-CoA carboxylase gene are linked with resistance to herbicides in a Lolium rigidum population. New Phytol. 2006, 172, 636–645. [Google Scholar] [CrossRef]
- Scarabel, L.; Panozzo, S.; Varotto, S.; Sattin, M. Allelic variation of the ACCase gene and response to ACCase-inhibiting herbicides in pinoxaden-resistant Lolium spp. Pest Manag. Sci. 2011, 67, 932–941. [Google Scholar]
- Busi, R.; Yu, Q.; Barrett-Lennard, R.; Powles, S. Long distance pollen-mediated flow of herbicide resistance genes in Lolium rigidum. Theor. Appl. Genet. 2008, 117, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Liu, Y.; Bu, D.; Yang, F.; Zhang, Z.; Qiang, S. A double mutation in the ALS gene confers a high level of resistance to mesosulfuron-methyl in shepherd’s-purse. Plants 2023, 12, 2730. [Google Scholar] [CrossRef] [PubMed]
- Gitsopoulos, T.; Georgoulas, I.; Vazanelli, E.; Botsoglou, D. Selectivity of the premixtures flufenacet, diflufenican and flufenacet, diflufenican, metribuzin on bread wheat (Triticum aestivum L.) and barley (Hordeum vulgare L.) and efficacy on ALS/ACCase-resistant populations of Lolium rigidum Gaudin. Agronomy 2024, 14, 949. [Google Scholar] [CrossRef]

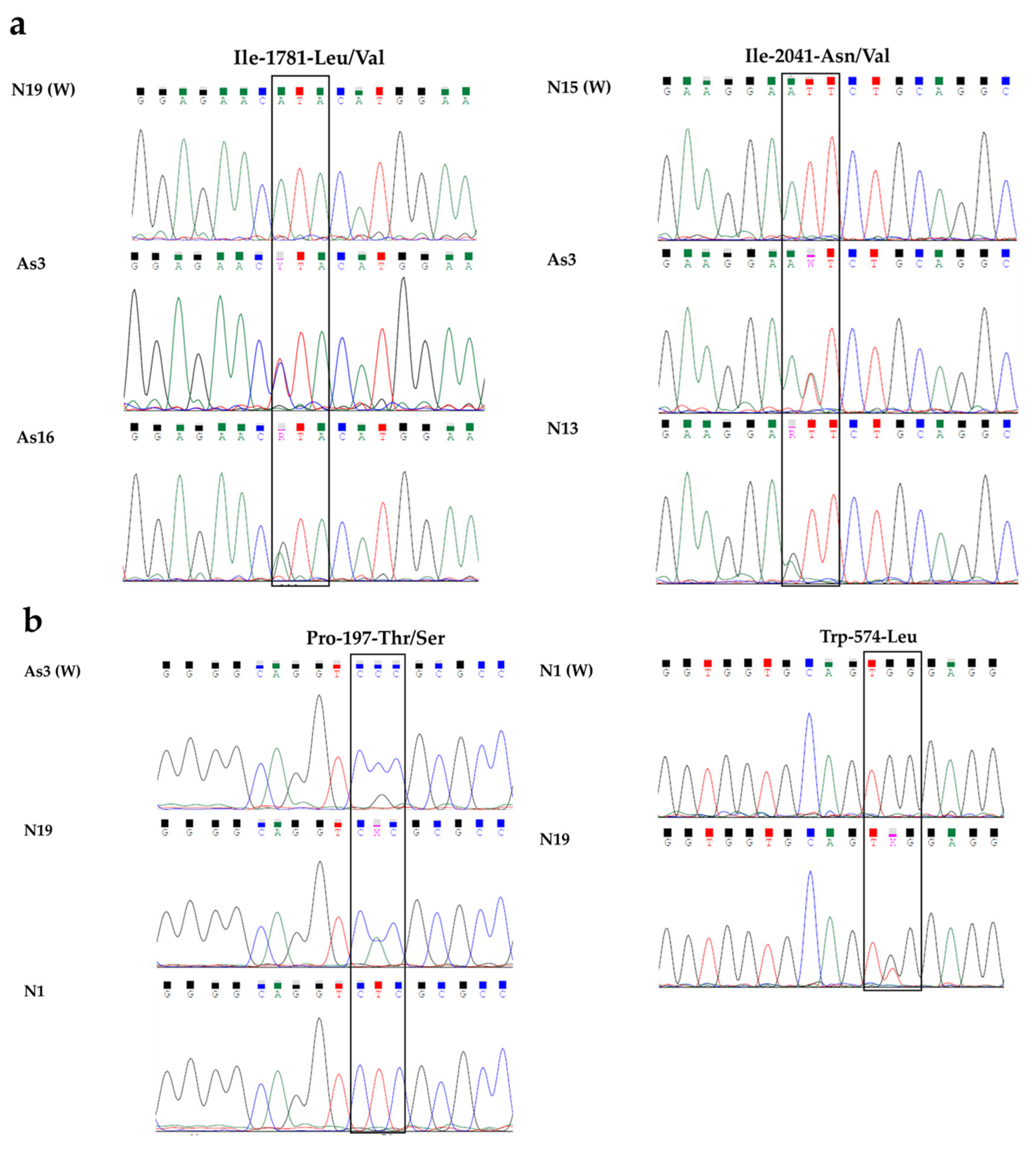

{kind=link}
{kind=link}
| Population | Pinoxaden | Mesosuluron-Methyl + Iodosulfuron-Methyl-Sodium | ||||
|---|---|---|---|---|---|---|
| FWR 1 | GR50 2 | R/S 3 | FWR 1 | GR50 2 | R/S 3 | |
| AS3 5 | 70 bcdf 4 | - | - | 54 bcde 4 | 17.13 | 3.6 |
| AS6 | 61 defg | - | - | 69 ab | 7.2 | 1.51 |
| AS9 | 62 cdefg | 42.40 | 37.24 | 60 bcde | 17.68 | 3.72 |
| AS10 | 44 g | 44.12 | 38.7 | 41 de | 29.04 | 6.11 |
| AS16 | 76 abcd | - | - | 63 bcd | 11.26 | 2.37 |
| AS19 | 93 a | 1.15 | - | 91 a | 4.75 | - |
| N1 | 71 bcde | - | - | 66 b | 17.28 | 3.63 |
| N4 | 72 bcde | - | - | 59 bcde | 13.31 | 2.8 |
| N6 | 81 ab | - | - | 71 ab | 9.12 | 1.92 |
| N11 | 52 fg | 37.30 | 32.71 | 91 a | - | - |
| N13 | 77 abcd | - | - | 55 bcde | 13.37 | 2.8 |
| N15 | 49 g | 52.41 | 45.97 | 37 e | 24.26 | 5.09 |
| N19 | 79 abc | - | - | 64 bc | 9.66 | 2.03 |
| N20 | 57 egf | 33.51 | 29.39 | 42 cde | 25.96 | 5.32 |
| Population | ACCase Mutations | ALS Mutations | ||||
|---|---|---|---|---|---|---|
| Ile-1781 | Ile-2041 | Mutant Plants/ Analyzed Plants | Pro-197 | Trp-574 | Mutant Plants/ Analyzed Plants | |
| AS3 | Leu(3) | Asn(3) | 3/3 | Pro(3) | Trp(3) | 0/3 |
| AS6 | Leu(3) | Asn(1), Ile(2) | 3/3 | Pro(2), Thr(1) | Trp(3) | 1/3 |
| AS9 | Leu(3) | Ile(3) | 3/3 | Pro(3) | Trp(3) | 0/3 |
| AS10 | Leu (3) | Asn(1), Ile(2) | 3/3 | Thr(1), Ser(2) | Leu(3) | 3/3 |
| AS16 | Leu(1) or Val(1) Leu(1), Ile(1) | Ile(3) | 2/3 | Gln(1), Ala(2) | Trp(3) | 3/3 |
| AS19 | Ile(3) | Asn(1), Ile(2) | 1/3 | Ser(1), (2)Pro | Trp(3) | 1/3 |
| N1 | Leu(3) | Asn(1), Ile(2) | 3/3 | Ser(3) | Trp(3) | 3/3 |
| N4 | Leu(2), Ile(1) | Asn(1), Ile(2) | 2/3 | Ser or Thr, Pro(2) | Trp(3) | 1/3 |
| N6 | Leu(3) | Asn(1), Ile(1) Val(1) | 3/3 | Pro(3) | Trp(3) | 0/3 |
| N11 | Leu(3) | Asn(3) | 3/3 | Ser(3) | Trp(3) | 3/3 |
| N13 | Leu(1), Ile(1) Val or Leu | Asn(2), Val(1) | 3/3 | Ser(3) | Trp(3) | 3/3 |
| N15 | Leu(2), Ile(1) | Ile(3) | 2/3 | Ser(2), Pro(1) | Trp(3) | 2/3 |
| N19 | Leu(3) | Asn(1), Ile(2) | 3/3 | Thr(2), Leu(1) | Leu(3) | 3/3 |
| N20 | Leu(3) | Asn(1), Ile(2) | 3/3 | Thr(1), Pro(2) | Trp(3) | 1/3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doulfi, D.; Economou, G.; Madesis, P.; Karapetsi, L.; Eleftherohorinos, I.G. Double Mutations Drive Multiple Resistances to Herbicides in Greek Rigid Ryegrass (Lolium rigidum Gaudin). Agronomy 2025, 15, 1532. https://doi.org/10.3390/agronomy15071532
Doulfi D, Economou G, Madesis P, Karapetsi L, Eleftherohorinos IG. Double Mutations Drive Multiple Resistances to Herbicides in Greek Rigid Ryegrass (Lolium rigidum Gaudin). Agronomy. 2025; 15(7):1532. https://doi.org/10.3390/agronomy15071532
Chicago/Turabian StyleDoulfi, Dimitra, Garyfallia Economou, Panagiotis Madesis, Lefkothea Karapetsi, and Ilias G. Eleftherohorinos. 2025. "Double Mutations Drive Multiple Resistances to Herbicides in Greek Rigid Ryegrass (Lolium rigidum Gaudin)" Agronomy 15, no. 7: 1532. https://doi.org/10.3390/agronomy15071532
APA StyleDoulfi, D., Economou, G., Madesis, P., Karapetsi, L., & Eleftherohorinos, I. G. (2025). Double Mutations Drive Multiple Resistances to Herbicides in Greek Rigid Ryegrass (Lolium rigidum Gaudin). Agronomy, 15(7), 1532. https://doi.org/10.3390/agronomy15071532